
Characterization, Comparison of Two New Mitogenomes of Crocodile Newts Tylototriton (Caudata: Salamandridae), and Phylogenetic Implications

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Sequencing
2.2. Sequence Assembly, Annotation, and Analysis
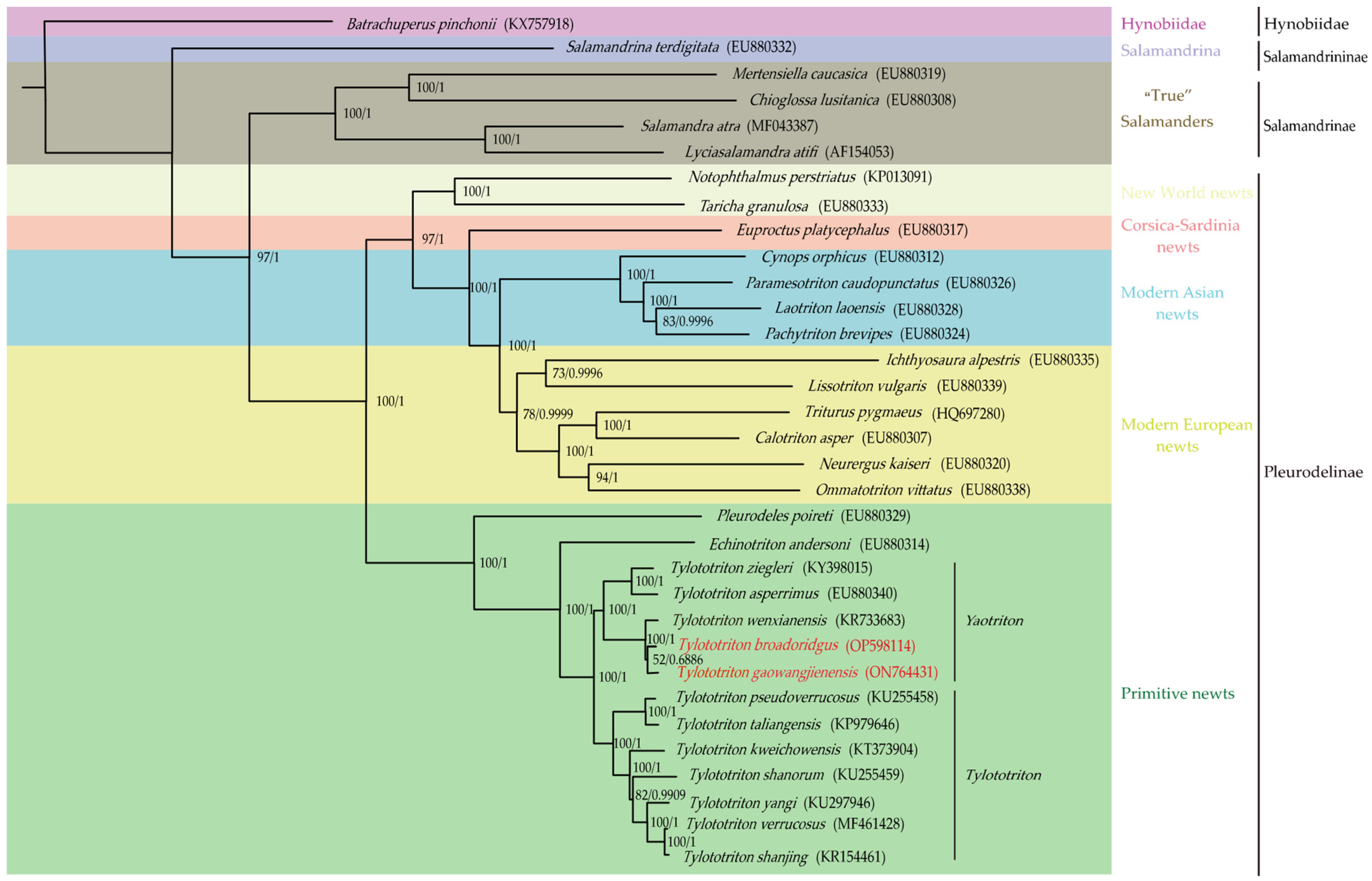
2.3. Phylogenetic Analysis
3. Results
3.1. Mitogenome Assembly and Undefined Sites Identification
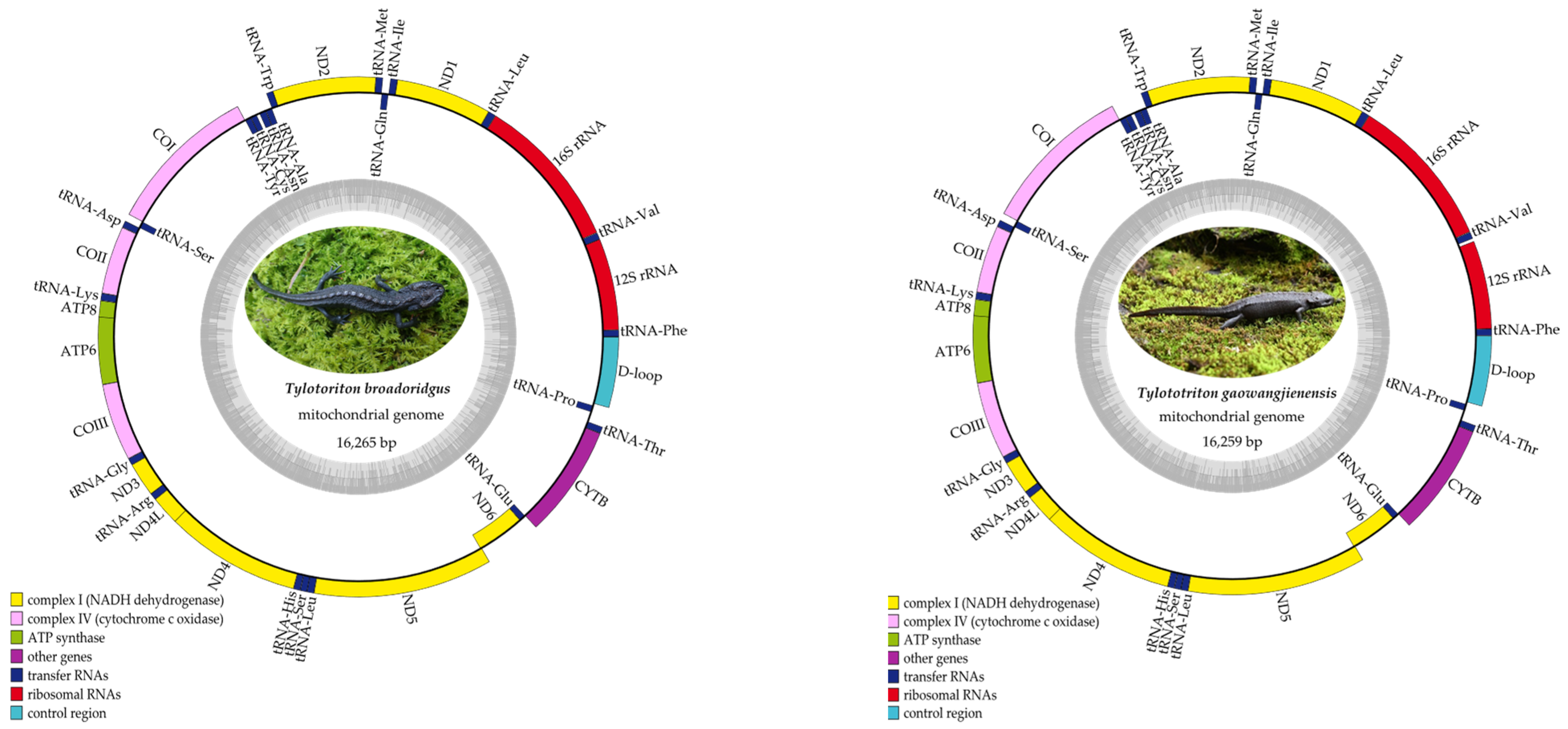
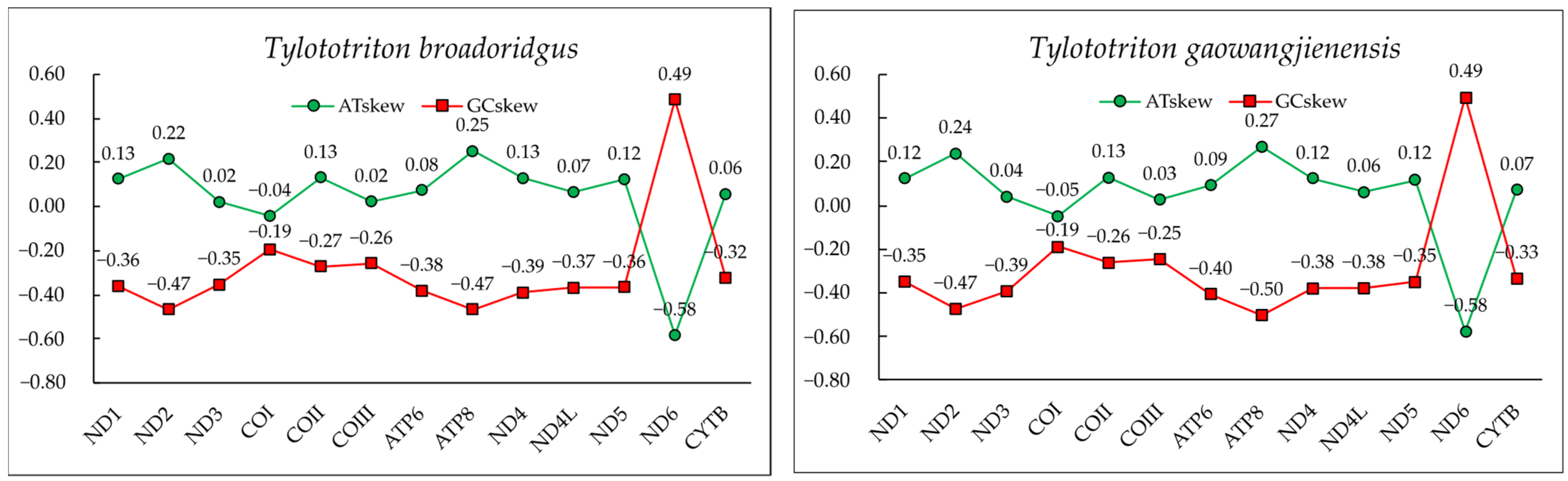
3.2. Mitogenome Annotation and Nucleotide Composition
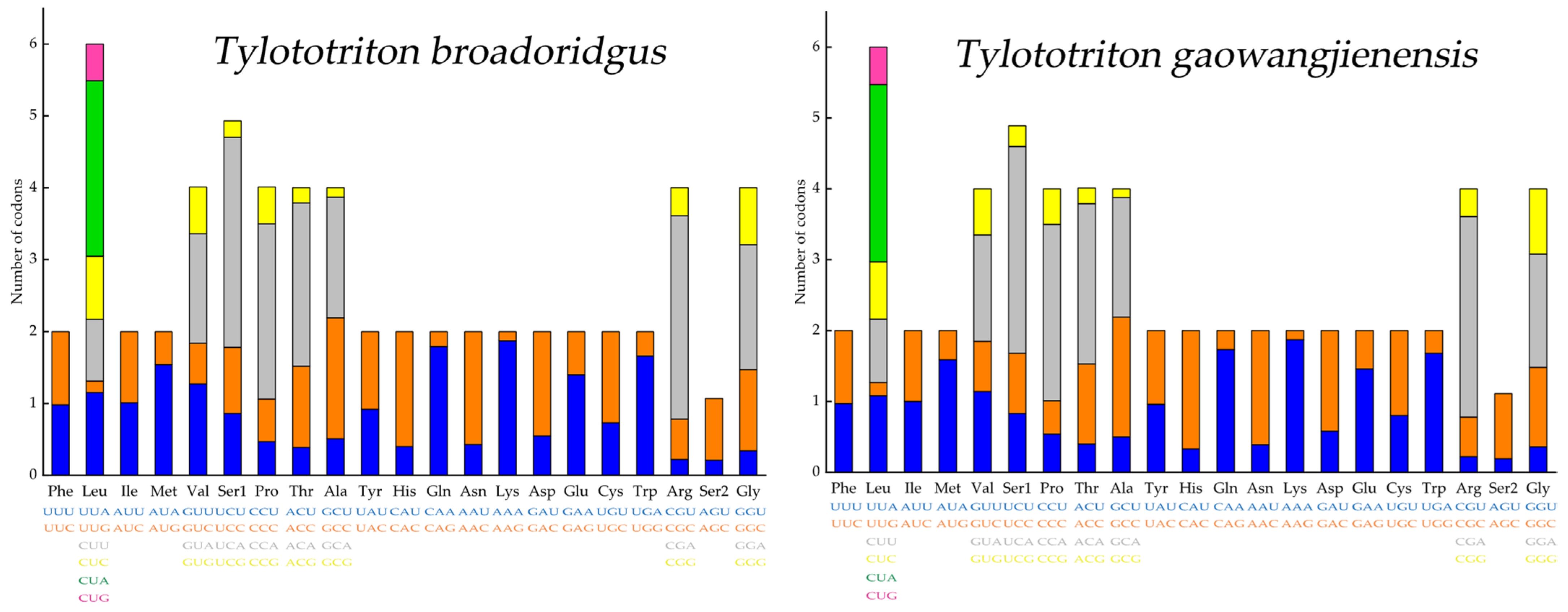
3.3. Characteristics of PCGs and Codon Usage
3.4. Characteristics of rRNAs, tRNAs, and the Control Region
3.5. Phylogenetic Analysis
3.6. Species Verification from ND2 and 16S rRNA Gene
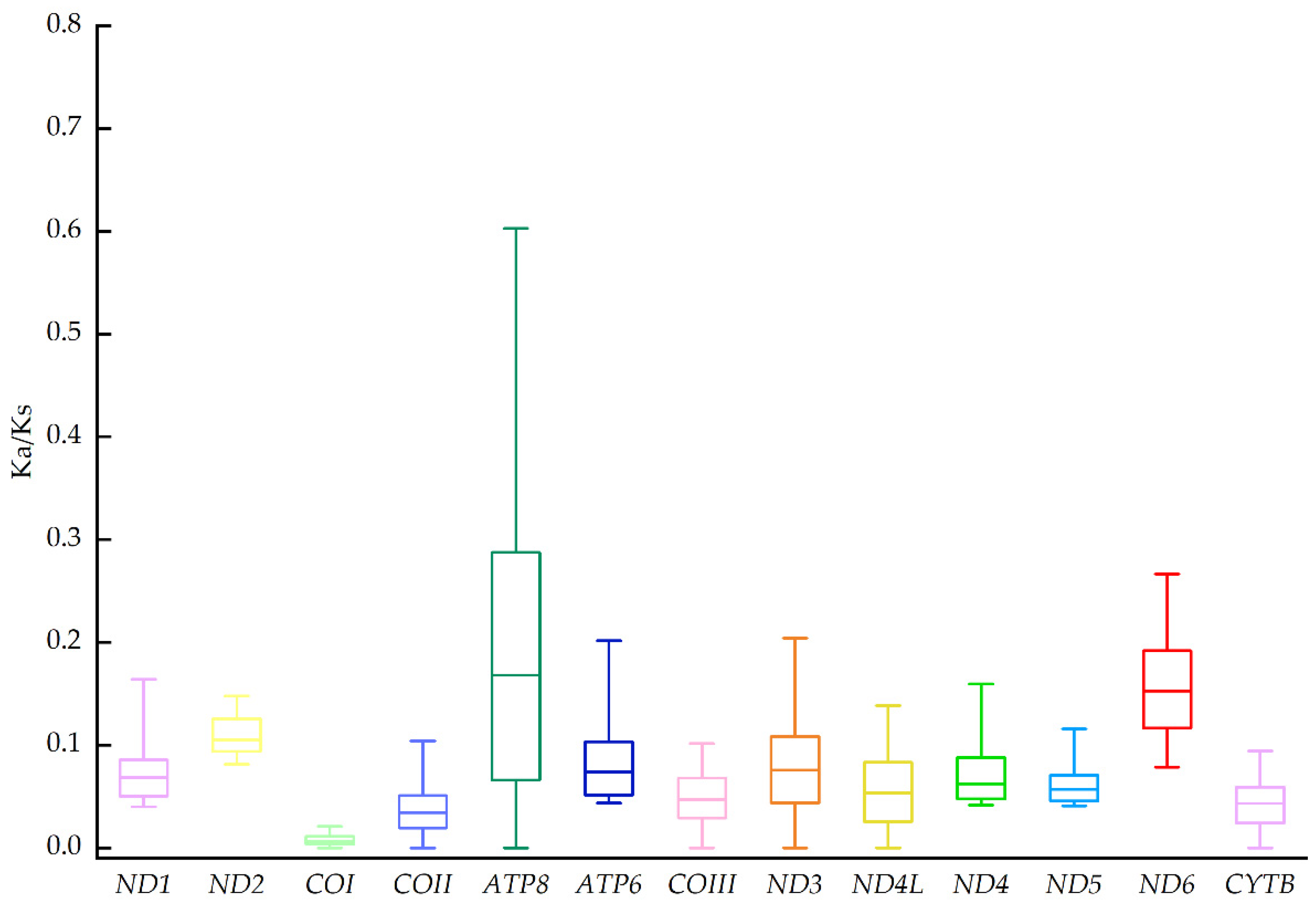
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef]
- Peng, R.; Zhang, P.; Xiong, J.L.; Gu, H.J.; Zeng, X.M.; Zou, F.D. Rediscovery of Protohynobius puxiongensis (Caudata: Hynobiidae) and its phylogenetic position based on complete mitochondrial genomes. Mol. Phylogenet Evol. 2010, 56, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.Y.; Lin, B.J.; Guo, C.; Tang, Z.; Wu, Z.J. The complete mitochondrial genome of the Hynobius maoershanensis (Caudata: Hynobiidae). Mitochondrial DNA A 2016, 27, 173–174. [Google Scholar] [CrossRef]
- Zhang, M.W.; Han, F.Y.; Ye, J.; Ni, Q.Y.; Li, Y.; Yao, Y.F.; Xu, H.L. The entire mitochondrial genome of Vietnam Warty Newt Paramesotriton deloustali (Salamandridae: Paramesotriton) with a new distribution record from China. Conserv. Genet. Resour. 2018, 10, 551–554. [Google Scholar] [CrossRef]
- Yi, M.R.; Hsu, K.C.; Gu, S.; He, X.B.; Luo, Z.S.; Lin, H.D.; Yan, Y.R. Complete mitogenomes of four Trichiurus species: A taxonomic review of the T.lepturus species complexe. ZooKeys 2022, 1084, 1–26. [Google Scholar] [CrossRef]
- Zardoya, R.; Meyer, A. Phylogenetic performance of mitochondrial protein-coding genes in resolving relationships among vertebrates. Mol. Biol. Evol. 1996, 13, 933–942. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Nishikawa, K.; Matsui, M.; Nguyen, T.Q.; Xie, F.; Li, C.; Khatiwada, J.R.; Zhang, B.W.; Gong, D.J.; Mo, Y.M.; et al. Phylogenetic surveys on the newt genus Tylototriton sensu lato (Salamandridae, Caudata) reveal cryptic diversity and novel diversification promoted by historical climatic shifts. PeerJ 2018, 6, e4384. [Google Scholar] [CrossRef] [Green Version]
- Rancilhac, L.; Irisarri, I.; Angelini, C.; Arntzen, J.W.; Babik, W.; Bossuyt, F.; Kunzel, S.; Luddecke, T.; Pasmans, F.; Sanchez, E.; et al. Phylotranscriptomic evidence for pervasive ancient hybridization among Old World salamanders. Mol. Phylogenet Evol. 2021, 155, 106967. [Google Scholar] [CrossRef] [PubMed]
- Amphibian Species of the World: An Online Reference. Version 6.1. Available online: https://amphibiansoftheworld.amnh.org/index.php (accessed on 1 August 2022).
- Weisrock, D.W.; Papenfuss, T.J.; Macey, J.R.; Litvinchuk, S.N.; Polymeni, R.; Ugurtas, I.H.; Zhao, E.; Jowkar, H.; Larson, A. A molecular assessment of phylogenetic relationships and lineage accumulation rates within the family Salamandridae (Amphibia, Caudata). Mol. Phylogenet Evol. 2006, 41, 368–383. [Google Scholar] [CrossRef]
- Zhang, P.; Papenfuss, T.J.; Wake, M.H.; Qu, L.; Wake, D.B. Phylogeny and biogeography of the family Salamandridae (Amphibia: Caudata) inferred from complete mitochondrial genomes. Mol. Phylogenet Evol. 2008, 49, 586–597. [Google Scholar] [CrossRef]
- Veith, M.; Bogaerts, S.; Pasmans, F.; Kieren, S. The changing views on the evolutionary relationships of extant Salamandridae (Amphibia: Urodela). PLoS ONE 2018, 13, e0198237. [Google Scholar] [CrossRef]
- Dubois, A.; Raffaëlli, J. A new ergotaxonomy of the family Salamandridae Goldfuss, 1820 (Amphibia, Urodela). Alytes 2009, 26, 1–85. [Google Scholar]
- Li, S.; Wei, G.; Cheng, Y.N.; Zhang, B.W.; Wang, B. Description of a New Species of the Asian Newt Genus Tylototriton sensu lato (Amphibia: Urodela: Salamandridae) from Southwest China. Asian Herpetol. Res. 2020, 11, 282–296. [Google Scholar]
- Phimmachak, S.; Aowphol, A.; Stuart, B.L. Morphological and molecular variation in Tylototriton (Caudata: Salamandridae) in Laos, with description of a new species. Zootaxa 2015, 4006, 285–310. [Google Scholar] [CrossRef] [Green Version]
- Nishikawa, K.; Matsui, M.; Nguyen, T.T. A New Species of Tylototriton from Northern Vietnam (Amphibia: Urodela: Salamandridae). Curr. Herpetol. 2013, 32, 34–49. [Google Scholar] [CrossRef]
- Yang, D.; Jiang, J.P.; Shen, Y.H.; Fei, D.B. A new species of the genus Tylototriton (Urodela: Salamandridae) from northeastern Hunan Province, China. Asian Herpetol. Res. 2014, 5, 1–11. [Google Scholar]
- Poyarkov, N.A.; Nguyen, T.V.; Arkhipov, D.V. A new species of the genus Tylototriton (Amphibia, Caudata, Salamandridae) from Central Vietnam. Taprobanica 2021, 10, 4–22. [Google Scholar] [CrossRef]
- Ponce, M.; Navarro, D.; Morales, R.; Batista, A. A new salamander of the genus Bolitoglossa (Caudata: Plethodontidae) from the highlands of western Panama. Zootaxa 2022, 5129, 543–556. [Google Scholar] [CrossRef]
- Yuan, Z.Y.; Jiang, K.; Lü, S.Q.; Yang, J.X.; Nguyen, Q.T.; Nguyen, T.T.; Jin, J.Q.; Che, J. A phylogeny of the Tylototriton asperrimus group (Caudata: Salamandridae) based on a mitochondrial study: Suggestions for a taxonomic revision. Zool. Res. 2011, 32, 577–584. [Google Scholar]
- Gao, Z.W.; Qian, T.Y.; Jiang, J.P.; Hou, D.J.; Deng, X.J.; Yang, D.D. Species diversity and distribution of amphibians and reptiles in Hunan Province, China. Biodivers. Sci. 2022, 30, 21290. [Google Scholar] [CrossRef]
- Shen, Y.H.; Yang, D.D.; Mo, X.Y.; Li, H.H.; Chen, D. The Fauna of Hunan: Amphibia; Hunan Science and Technology Press: Changsha, China, 2014. (In Chinese) [Google Scholar]
- Xiang, Y.; Wu, T.; Zhang, Z.L.; Huang, Y.P.; Huang, X.L.; Wang, B.Z.; Jiang, W.S.; Zhang, Y.X. Tylototriton gaowangjienensis, a New Species of Crocodile Newts (Caudata: Salamandridae: Tylototriton) from Hunan, China; Jishou University: Jishou, China, 2022; (to be submitted). [Google Scholar]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar]
- Meng, G.; Li, Y.Y.; Yang, C.T.; Liu, S.L. MitoZ: A toolkit for animal mitochondrial genome assembly, annotation and visualization. Nucleic Acids Res. 2019, 47, e63. [Google Scholar] [CrossRef] [Green Version]
- Song, M.H.; Yan, C.C.; Li, J.T. MEANGS: An efficient seed-free tool for de novo assembling animal mitochondrial genome using whole genome NGS data. Brief. Bioinform. 2022, 23, bbab538. [Google Scholar] [CrossRef]
- Donath, A.; Jühling, F.; Al-Arab, M.; Bernhart, S.H.; Reinhardt, F.; Stadler, P.F.; Middendorf, M.; Bernt, M. Improved annotation of protein-coding genes boundaries in metazoan mitochondrial genomes. Nucleic Acids Res. 2019, 47, 10543–10552. [Google Scholar] [CrossRef]
- Tillich, M.P.; Lehwark, T.; Pellizzer, E.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq-versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, 6–11. [Google Scholar] [CrossRef] [Green Version]
- Chan, P.P.; Lin, B.Y.; Mak, A.J.; Lowe, T.M. tRNAscan-SE 2.0: Improved detection and functional classification of transfer RNA genes. Nucleic Acids Res. 2021, 49, 9077–9096. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Moberly, J.G.; Bernards, M.T.; Waynant, K.V. Key features and updates for Origin 2018. J. Cheminform. 2018, 10, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. Using RAxML to infer phylogenies. Curr. Protoc. Bioinform. 2015, 51, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics, 1st ed.; Oxford University Press: Oxford, MI, USA, 2000; pp. 1–333. [Google Scholar]
- Hurst, L.D. The Ka/Ks ratio: Diagnosing the form of sequence evolution. Trends Genet. 2002, 18, 486–487. [Google Scholar] [CrossRef]
- Sangster, G.; Luksenburg, J.A. Sharp increase of problematic mitogenomes of birds: Causes, consequences, and remedies. Genome Biol. Evol. 2021, 13, evab210. [Google Scholar] [CrossRef] [PubMed]
- Del Angel, V.D.; Hjerde, E.; Sterck, L.; Capella-Gutierrez, S.; Notredame, C.; Pettersson, O.V.; Amselem, J.; Bouri, L.; Bocs, S.; Klopp, C.; et al. Ten steps to get started in Genome Assembly and Annotation. F1000Research 2018, 7, 148. [Google Scholar] [CrossRef] [PubMed]
- Olmo, E. Quantitative variations in the nuclear DNA and phylogenesis of the Amphibia. Caryologia 1973, 26, 43–68. [Google Scholar] [CrossRef]
- Han, F.Y.; Jiang, Y.; Zhang, M.W. The complete mitochondrial genome sequence of Wenxian Knobby Newt Tylototriton wenxianensis (Amphibia: Caudata). Mitochondrial DNA A 2016, 27, 2901–2902. [Google Scholar] [CrossRef]
- Jiang, Y.; Li, Z.Y.; Liu, J.B.; Li, Y.; Ni, Q.Y.; Yao, Y.F.; Xu, H.L.; Li, Y.; Zhang, M.W. The complete mitochondrial genome sequence of Tylototriton taliangensis (Amphibia: Caudata). Mitochondrial DNA A 2016, 27, 2639–2640. [Google Scholar] [CrossRef]
- Jiang, Y.; Yang, M.X.; Han, F.Y.; Li, Y.; Ni, Q.Y.; Yao, Y.F.; Xu, H.; Li, Y.; Zhang, M.W. The complete mitochondrial genome sequence of Red knobby newt Tylototriton shanjing (Amphibia: Caudata). Mitochondrial DNA A 2016, 27, 2773–2774. [Google Scholar] [CrossRef]
- Jiang, Y.; Wei, Z.M.; Han, F.Y.; Ni, Q.Y.; Yao, Y.F.; Xu, H.L.; Li, Y.; Rao, D.Q.; Zhang, M.W. The complete mitogenome sequence of Tylototriton ziegleri (Amphibia: Caudata). Conserv. Genet. Resour. 2017, 9, 503–506. [Google Scholar] [CrossRef]
- Cui, L.; Han, F.Y.; Li, D.Y.; Fan, X.L.; Yang, M.Y.; Yang, D.Y.; Ni, Q.Y.; Lia, Y.; Yao, Y.F.; Xu, H.L.; et al. The complete mitochondrial genome of Cynops cyanurus chuxiongensis (Caudata: Salamandridae). Mitochondrial DNA Part. B 2020, 5, 231–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suk, H.Y.; Kim, D.Y.; Cha, S.H.; Min, M.S. The complete mitochondrial genome information of Hynobius unisacculus (Amphibia, Caudata, Hynobiidae) and the phylogenetic implication. Mitochondrial DNA Part. B 2019, 4, 3705–3706. [Google Scholar] [CrossRef] [PubMed]
- Kieren, S.; Sparreboom, M.; Hochkirch, A.; Veith, M. A biogeographic and ecological perspective to the evolution of reproductive behaviour in the family Salamandridae. Mol. Phylogenet Evol. 2018, 121, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Pyron, R.A.; Wiens, J.J. A large-scale phylogeny of Amphibia including over 2800 species, and a revised classification of extant frogs, salamanders, and caecilians. Mol. Phylogenet Evol. 2011, 61, 543–583. [Google Scholar] [CrossRef]
- Nishikawa, K.; Khonsue, W.; Pomchote, P.; Matsui, M. Two new species of Tylototriton from Thailand (amphibia: Urodela: Salamandridae). Zootaxa 2013, 3737, 261–279. [Google Scholar] [CrossRef]
- Qian, L.F.; Sun, X.N.; Li, J.Q.; Guo, W.B.; Pan, T.; Kang, X.; Wang, H.; Jiang, J.P.; Wu, J.; Zhang, B.W. A new species of the genus Tylototriton (Amphibia: Urodela: Salamandridae) from the southern Dabie Mountains in Anhui Province. Asian Herpetol. Res. 2017, 8, 151–164. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Position | Length (bp) | Start Codon | Stop Codon | Anticodon | Strand | Intergenic Nucleotide | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TB | TG | TB | TG | TB | TG | TB | TG | |||||||
| From | To | From | To | TB | TG | |||||||||
| tRNAPhe | 1 | 68 | 1 | 69 | 68 | 69 | GAA | H | 0 | 0 | ||||
| 12S rRNA | 69 | 996 | 70 | 996 | 928 | 927 | H | −1 | −1 | |||||
| tRNAVal | 996 | 1065 | 996 | 1065 | 70 | 70 | TAC | H | 2 | 2 | ||||
| 16S rRNA | 1068 | 2630 | 1068 | 2627 | 1563 | 1560 | H | 1 | 1 | |||||
| tRNALeu | 2632 | 2706 | 2629 | 2703 | 75 | 75 | TAA | H | 0 | 0 | ||||
| ND1 | 2707 | 3675 | 2704 | 3672 | 969 | 969 | ATG | ATG | TAG | TAG | H | −1 | −1 | |
| tRNAIle | 3675 | 3745 | 3672 | 3742 | 71 | 71 | GAT | H | 2 | 2 | ||||
| tRNAGln | 3748 | 3818 | 3745 | 3815 | 71 | 71 | CAT | L | 2 | 1 | ||||
| tRNAMet | 3821 | 3890 | 3817 | 3886 | 70 | 70 | TCA | H | 0 | 0 | ||||
| ND2 | 3891 | 4934 | 3887 | 4930 | 1044 | 1044 | ATG | ATG | TAA | TAA | H | −2 | −2 | |
| tRNATrp | 4933 | 5001 | 4929 | 4997 | 69 | 69 | GTC | H | 1 | 1 | ||||
| tRNAAla | 5003 | 5071 | 4999 | 5067 | 69 | 69 | TTT | L | 0 | 0 | ||||
| tRNAAsn | 5072 | 5144 | 5068 | 5140 | 73 | 73 | TCC | L | 2 | 2 | ||||
| OL | 5147 | 5179 | 5143 | 5175 | 33 | 33 | H | −1 | −1 | |||||
| tRNACys | 5179 | 5244 | 5175 | 5240 | 66 | 66 | TCG | L | 0 | 0 | ||||
| tRNATyr | 5245 | 5311 | 5241 | 5307 | 67 | 67 | GTG | L | 1 | 1 | ||||
| COI | 5313 | 6863 | 5309 | 6859 | 1551 | 1551 | GTG | GTG | TAA | TAA | H | 0 | 0 | |
| tRNASer | 6864 | 6934 | 6860 | 6930 | 71 | 71 | GCT | L | 1 | 1 | ||||
| tRNAAsp | 6936 | 7005 | 6932 | 7001 | 70 | 70 | TAG | H | 1 | 1 | ||||
| COII | 7007 | 7694 | 7003 | 7690 | 688 | 688 | ATG | ATG | T(AA) | T(AA) | H | 0 | 0 | |
| tRNALys | 7695 | 7767 | 7691 | 7763 | 73 | 73 | TGT | H | 1 | 1 | ||||
| ATP8 | 7769 | 7936 | 7765 | 7932 | 168 | 168 | ATG | ATG | TAA | TAA | H | −10 | −10 | |
| ATP6 | 7927 | 8610 | 7923 | 8606 | 684 | 684 | GTG | ATG | TAA | TAA | H | −1 | −1 | |
| COIII | 8610 | 9393 | 8606 | 9389 | 784 | 784 | ATG | ATG | T(AA) | T(AA) | H | 0 | 0 | |
| tRNAGly | 9394 | 9463 | 9390 | 9458 | 70 | 69 | TGG | H | 0 | 0 | ||||
| ND3 | 9464 | 9811 | 9459 | 9806 | 348 | 348 | ATG | ATG | TAA | TAA | H | −2 | −2 | |
| tRNAArg | 9810 | 9878 | 9805 | 9873 | 69 | 69 | TTC | H | 0 | 0 | ||||
| ND4L | 9879 | 10,175 | 9874 | 10,170 | 297 | 297 | ATG | ATG | TAA | TAA | H | −7 | −7 | |
| ND4 | 10,169 | 11,546 | 10,164 | 11,541 | 1378 | 1378 | ATG | ATG | T(AA) | T(AA) | H | 0 | 0 | |
| tRNAHis | 11,547 | 11,614 | 11,542 | 11,609 | 68 | 68 | TGA | H | 0 | 0 | ||||
| tRNASer | 11,615 | 11,682 | 11,610 | 11,677 | 68 | 68 | GTA | H | −1 | −1 | ||||
| tRNALeu | 11,682 | 11,753 | 11,677 | 11,748 | 72 | 72 | GCA | H | 0 | 0 | ||||
| ND5 | 11,754 | 13,565 | 11,749 | 13,560 | 1812 | 1812 | ATG | ATG | TAA | TAA | H | −15 | −15 | |
| ND6 | 13,551 | 14,069 | 13,546 | 14,064 | 519 | 519 | ATG | ATG | AGA | AGA | L | 0 | 0 | |
| tRNAGlu | 14,070 | 14,137 | 14,065 | 14,132 | 68 | 68 | GTT | L | 2 | 2 | ||||
| CYTB | 14,140 | 15,280 | 14,135 | 15,275 | 1141 | 1141 | ATG | ATG | T(AA) | T(AA) | H | 0 | 0 | |
| tRNAThr | 15,281 | 15,348 | 15,276 | 15,343 | 68 | 68 | TGC | H | 108 | 108 | ||||
| tRNAPro | 15,457 | 15,527 | 15,452 | 15,522 | 71 | 71 | TTG | L | 22 | 22 | ||||
| D-loop | 15,550 | 16,265 | 15,545 | 16,259 | 716 | 715 | H | 0 | 0 | |||||
| Species | Total Length (bp) | T (%) | C (%) | A (%) | G (%) | A + T Content (%) | AT-Skew | GC-Skew | Accession Number |
|---|---|---|---|---|---|---|---|---|---|
| T. broadoridgus | 16,265 | 25.7 | 26.3 | 33.6 | 14.5 | 59.3 | 0.13 | −0.29 | OP598114 |
| T. gaowangjienensis | 16,259 | 25.6 | 26.3 | 33.6 | 14.6 | 59.2 | 0.14 | −0.29 | ON764431 |
| Tylototriton wenxianensis | 16,265 | 25.67 | 26.20 | 33.62 | 14.51 | 59.29 | 0.13 | −0.29 | KR733683 |
| Tylototriton kweichowensis | 16,727 | 25.64 | 26.10 | 33.93 | 14.33 | 59.57 | 0.14 | −0.29 | KT373904 |
| Tylototriton asperrimus | 16,161 | 25.51 | 26.50 | 33.26 | 14.73 | 58.77 | 0.13 | −0.29 | EU880340 |
| Tylototriton pseudoverrucosus | 16,265 | 26.06 | 25.77 | 33.40 | 14.77 | 59.46 | 0.12 | −0.27 | KU255458 |
| Tylototriton shanjing | 16,661 | 25.41 | 26.28 | 34.04 | 14.27 | 59.45 | 0.15 | −0.30 | KR154461 |
| Tylototriton taliangensis | 16,265 | 26.03 | 25.76 | 33.42 | 14.79 | 59.45 | 0.12 | −0.27 | KP979646 |
| Tylototriton verrucosus | 16,660 | 25.43 | 26.31 | 33.99 | 14.27 | 59.42 | 0.14 | −0.30 | MF461428 |
| Tylototriton yangi | 16,648 | 25.53 | 26.21 | 33.99 | 14.28 | 59.51 | 0.14 | −0.29 | KU297946 |
| Tylototriton ziegleri | 16,266 | 25.49 | 26.38 | 33.71 | 14.42 | 59.20 | 0.14 | −0.29 | KY398015 |
| Tylototriton shanorum | 17,096 | 25.26 | 26.40 | 34.11 | 14.23 | 59.37 | 0.15 | −0.30 | KU255459 |
| Cynops orphicus | 16,296 | 28.91 | 23.16 | 32.88 | 15.05 | 61.79 | 0.06 | −0.21 | EU880312 |
| Echinotriton andersoni | 16,272 | 26.47 | 25.44 | 34.02 | 14.07 | 60.49 | 0.12 | −0.29 | EU880314 |
| Euproctus platycephalus | 15,799 | 30.14 | 22.09 | 34.15 | 13.61 | 64.30 | 0.06 | −0.24 | EU880317 |
| Calotriton asper | 16,564 | 26.47 | 25.93 | 32.20 | 15.41 | 58.66 | 0.10 | −0.25 | EU880307 |
| Ichthyosaura alpestris | 16,339 | 27.39 | 25.05 | 32.91 | 14.65 | 60.30 | 0.09 | −0.26 | EU880335 |
| Laotriton laoensis | 16,361 | 27.30 | 24.88 | 31.80 | 16.03 | 59.09 | 0.08 | −0.22 | EU880328 |
| Lissotriton vulgaris | 16,310 | 28.34 | 24.74 | 31.76 | 15.16 | 60.10 | 0.06 | −0.24 | EU880339 |
| Neurergus kaiseri | 16,202 | 27.69 | 23.97 | 34.57 | 13.78 | 62.26 | 0.11 | −0.27 | EU880320 |
| Notophthalmus perstriatus | 16,336 | 28.34 | 23.84 | 34.08 | 13.74 | 62.41 | 0.09 | −0.27 | KP013091 |
| Paramesotriton caudopunctatus | 15,968 | 28.49 | 23.57 | 33.73 | 14.21 | 62.22 | 0.08 | −0.25 | EU880326 |
| Pleurodeles poireti | 16,211 | 27.96 | 24.46 | 33.26 | 14.32 | 61.22 | 0.09 | −0.26 | EU880329 |
| Taricha granulosa | 16,151 | 24.99 | 27.67 | 31.37 | 15.97 | 56.36 | 0.11 | −0.27 | EU880333 |
| Lyciasalamandra atifi | 16,650 | 29.07 | 24.13 | 32.34 | 14.47 | 61.41 | 0.05 | −0.25 | AF154053 |
| Mertensiella caucasica | 17,023 | 29.38 | 24.54 | 31.96 | 14.12 | 61.34 | 0.04 | −0.27 | EU880319 |
| Salamandra atra | 15,592 | 30.31 | 23.35 | 32.45 | 13.89 | 62.76 | 0.03 | −0.25 | MF043387 |
| Salamandrina terdigitata | 16,252 | 29.59 | 22.26 | 34.57 | 13.59 | 64.16 | 0.08 | −0.24 | EU880332 |
| Chioglossa lusitanica | 16,417 | 31.03 | 22.80 | 33.01 | 13.16 | 64.04 | 0.03 | −0.27 | EU880308 |
| Pachytriton brevipes | 16,240 | 28.18 | 23.80 | 33.15 | 14.87 | 61.33 | 0.08 | −0.23 | EU880324 |
| Triturus pygmaeus | 16,442 | 27.25 | 25.73 | 31.83 | 15.19 | 59.08 | 0.08 | −0.26 | HQ697280 |
| Ommatotriton vittatus | 16,193 | 28.80 | 23.84 | 32.38 | 14.98 | 61.18 | 0.06 | −0.23 | EU880338 |
| B. pinchonii | 16,381 | 32.84 | 19.65 | 33.92 | 13.60 | 66.75 | 0.02 | −0.18 | KX757918 |
| Size | A (%) | T (%) | C (%) | G (%) | A + T (%) | AT-Skew | GC-Skew | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TB | TG | TB | TG | TB | TG | TB | TG | TB | TG | TB | TG | TB | TG | TB | TG | |
| D-loop | 716 | 715 | 28.6 | 29.0 | 34.9 | 34.5 | 21.2 | 21.0 | 15.2 | 15.5 | 63.5 | 63.5 | −0.10 | −0.09 | −0.17 | −0.15 |
| 12SrRNA | 928 | 927 | 37.9 | 38.1 | 19.9 | 20.1 | 23.9 | 23.7 | 18.2 | 18.1 | 57.8 | 58.2 | 0.31 | 0.31 | −0.14 | −0.13 |
| 16SrRNA | 1563 | 1560 | 40.2 | 40.1 | 23.3 | 23.4 | 20.3 | 20.1 | 16.3 | 16.4 | 63.5 | 63.5 | 0.27 | 0.26 | −0.11 | −0.10 |
| tRNAs | 1537 | 1537 | 32.3 | 32.1 | 30.1 | 30.0 | 17.7 | 17.9 | 19.8 | 20.0 | 62.4 | 62.1 | 0.04 | 0.03 | 0.06 | 0.05 |
| PCGs-1st | 3795 | 3795 | 30 | 30.3 | 25.4 | 25.1 | 25.1 | 25.2 | 19.4 | 19.3 | 55.4 | 55.4 | 0.08 | 0.09 | −0.13 | −0.13 |
| PCGs-2nd | 3794 | 3794 | 25.1 | 25 | 33.6 | 33.4 | 27.9 | 28 | 13.5 | 13.5 | 58.7 | 58.4 | −0.14 | −0.14 | −0.35 | −0.35 |
| PCGs-3rd | 3794 | 3794 | 37.8 | 37 | 23 | 23 | 28.1 | 28 | 11.1 | 11.4 | 60.8 | 60.0 | 0.24 | 0.23 | −0.43 | −0.42 |
| PCGs | 11,383 | 11,383 | 31 | 31 | 27.3 | 27.2 | 27 | 27.1 | 14.7 | 14.8 | 58.3 | 58.2 | 0.06 | 0.07 | −0.29 | −0.29 |
| Genome | 16,265 | 16,259 | 33.6 | 33.6 | 25.7 | 25.6 | 26.3 | 26.3 | 14.5 | 14.6 | 59.3 | 59.2 | 0.13 | 0.14 | −0.29 | −0.29 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.-X.; Lan, X.-Y.; Luo, Q.-H.; Gu, Z.-R.; Zhou, Q.; Zhang, M.-Y.; Zhang, Y.-X.; Jiang, W.-S. Characterization, Comparison of Two New Mitogenomes of Crocodile Newts Tylototriton (Caudata: Salamandridae), and Phylogenetic Implications. Genes 2022, 13, 1878. https://doi.org/10.3390/genes13101878
Wang J-X, Lan X-Y, Luo Q-H, Gu Z-R, Zhou Q, Zhang M-Y, Zhang Y-X, Jiang W-S. Characterization, Comparison of Two New Mitogenomes of Crocodile Newts Tylototriton (Caudata: Salamandridae), and Phylogenetic Implications. Genes. 2022; 13(10):1878. https://doi.org/10.3390/genes13101878
Chicago/Turabian StyleWang, Jin-Xiu, Xiang-Ying Lan, Qing-Hua Luo, Zhi-Rong Gu, Qiang Zhou, Ming-Yao Zhang, You-Xiang Zhang, and Wan-Sheng Jiang. 2022. "Characterization, Comparison of Two New Mitogenomes of Crocodile Newts Tylototriton (Caudata: Salamandridae), and Phylogenetic Implications" Genes 13, no. 10: 1878. https://doi.org/10.3390/genes13101878