

Ancient Components and Recent Expansion in the Eurasian Heartland: Insights into the Revised Phylogeny of Y-Chromosomes from Central Asia
 , , and
, , and
Abstract
:1. Introduction
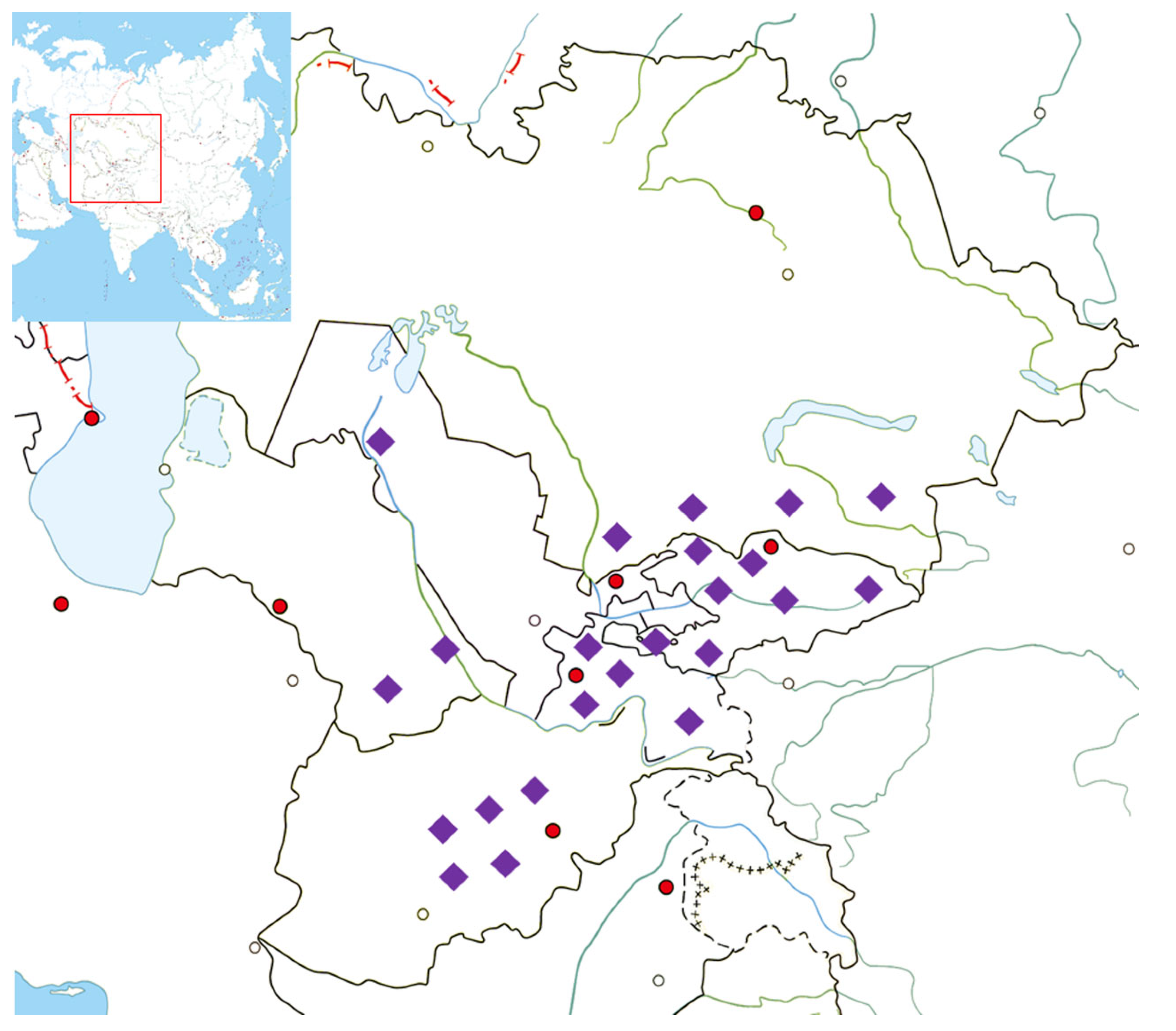
2. Materials and Methods
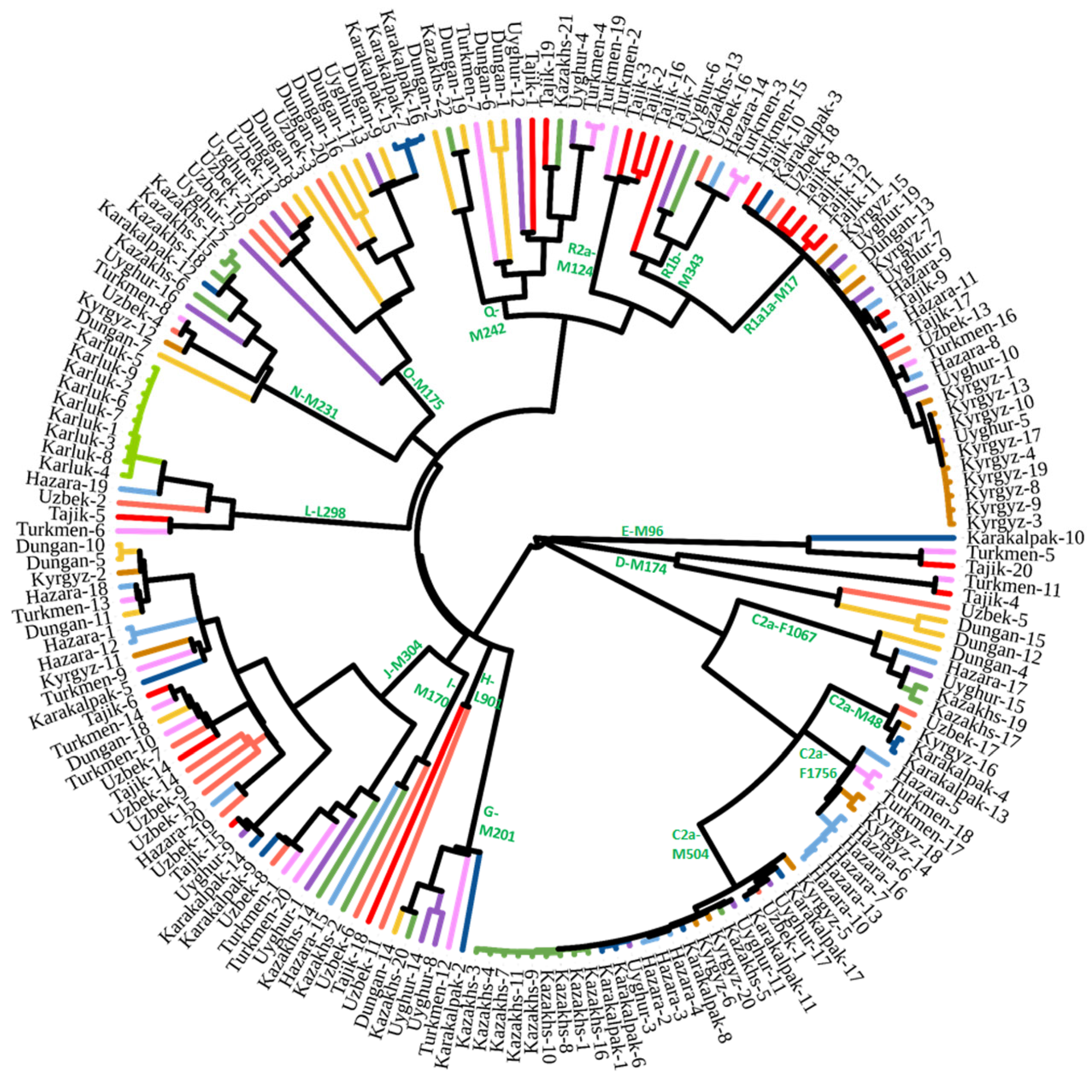
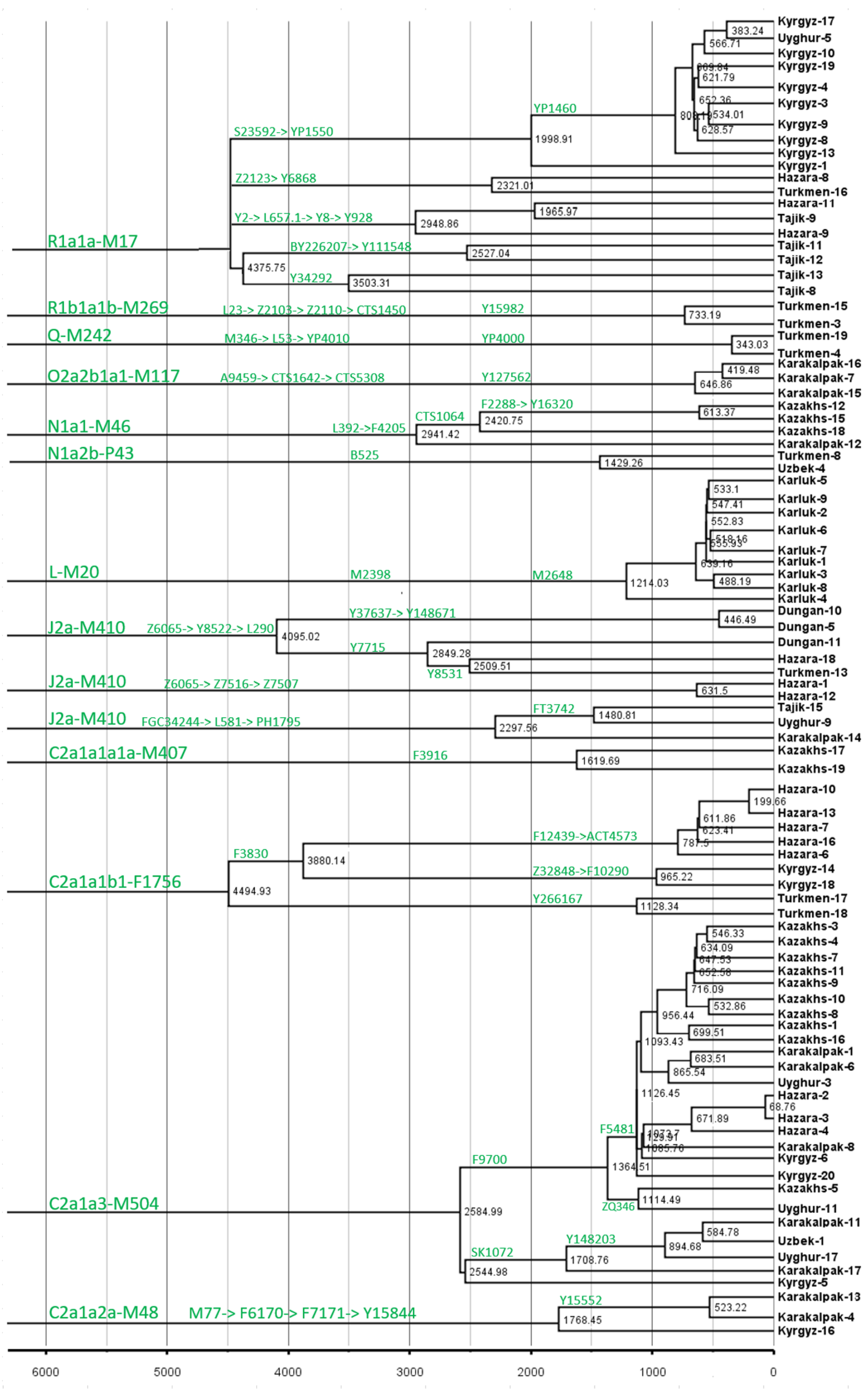
3. Results
4. Discussion
4.1. Multiple Layers of Deposition of Ancient Components
4.2. Recent Expansion and the Formation of Modern Populations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fu, Q.; Li, H.; Moorjani, P.; Jay, F.; Slepchenko, S.M.; Bondarev, A.A.; Johnson, P.L.F.; Aximu-Petri, A.; Prüfer, K.; De Filippo, C.; et al. Genome sequence of a 45,000-year-old modern human from western Siberia. Nature 2014, 514, 445–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghavan, M.; Skoglund, P.; Graf, K.E.; Metspalu, M.; Albrechtsen, A.; Moltke, I.; Rasmussen, S.; Stafford, T.W., Jr.; Orlando, L.; Metspalu, E.; et al. Upper Paleolithic Siberian genome reveals dual ancestry of Native Americans. Nature 2014, 505, 87–91. [Google Scholar] [CrossRef] [Green Version]
- Sikora, M.; Pitulko, V.V.; Sousa, V.C.; Allentoft, M.E.; Vinner, L.; Rasmussen, S.; Margaryan, A.; Damgaard, P.D.B.; De La Fuente, C.; Renaud, G.; et al. The population history of northeastern Siberia since the Pleistocene. Nature 2019, 570, 182–188. [Google Scholar] [CrossRef] [Green Version]
- Derenko, M.; Malyarchuk, B.; Grzybowski, T.; Denisova, G.; Rogalla, U.; Perkova, M.; Dambueva, I.; Zakharov, I. Origin and Post-Glacial Dispersal of Mitochondrial DNA Haplogroups C and D in Northern Asia. PLoS ONE 2010, 5, e15214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pugach, I.; Matveev, R.; Spitsyn, V.; Makarov, S.; Novgorodov, I.; Osakovsky, V.; Stoneking, M.; Pakendorf, B. The Complex Admixture History and Recent Southern Origins of Siberian Populations. Mol. Biol. Evol. 2016, 33, 1777–1795. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.-H.; Wang, L.-X.; Wen, S.-Q.; Yan, S.; Canada, R.; Gurianov, V.; Huang, Y.-Z.; Mallick, S.; Biondo, A.; O’Leary, A.; et al. Paternal origin of Paleo-Indians in Siberia: Insights from Y-chromosome sequences. Eur. J. Hum. Genet. 2018, 26, 1687–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, J.; Ma, P.; Cheng, H.; Wang, C.; Li, Y.; Cui, Y.; Yao, H.; Wen, S.; Wei, L. Post-last glacial maximum expansion of Y-chromosome haplogroup C2a-L1373 in northern Asia and its implications for the origin of Native Americans. Am. J. Phys. Anthr. 2020, 174, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Spyrou, M.A.; Karapetian, M.; Shnaider, S.; Radzevičiūtė, R.; Nägele, K.; Neumann, G.U.; Penske, S.; Zech, J.; Lucas, M.; et al. Paleolithic to Bronze Age Siberians Reveal Connections with First Americans and across Eurasia. Cell 2020, 181, 1232–1245.e20. [Google Scholar] [CrossRef]
- Jeong, C.; Wilkin, S.; Amgalantugs, T.; Bouwman, A.S.; Taylor, W.T.T.; Hagan, R.W.; Bromage, S.; Tsolmon, S.; Trachsel, C.; Grossmann, J.; et al. Bronze Age population dynamics and the rise of dairy pastoralism on the eastern Eurasian steppe. Proc. Natl. Acad. Sci. USA 2018, 115, E11248–E11255. [Google Scholar] [CrossRef] [Green Version]
- Jeong, C.; Balanovsky, O.; Lukianova, E.; Kahbatkyzy, N.; Flegontov, P.; Zaporozhchenko, V.; Immel, A.; Wang, C.-C.; Ixan, O.; Khussainova, E.; et al. The genetic history of admixture across inner Eurasia. Nat. Ecol. Evol. 2019, 3, 966–976. [Google Scholar] [CrossRef]
- Narasimhan, V.M.; Patterson, N.; Moorjani, P.; Rohland, N.; Bernardos, R.; Mallick, S.; Lazaridis, I.; Nakatsuka, N.; Olalde, I.; Lipson, M.; et al. The formation of human populations in South and Central Asia. Science 2019, 365, eaat7487. [Google Scholar] [CrossRef] [PubMed]
- Gnecchi-Ruscone, G.A.; Khussainova, E.; Kahbatkyzy, N.; Musralina, L.; Spyrou, M.A.; Bianco, R.A.; Radzeviciute, R.; Martins, N.F.G.; Freund, C.; Iksan, O.; et al. Ancient genomic time transect from the Central Asian Steppe unravels the history of the Scythians. Sci. Adv. 2021, 7, 4414. [Google Scholar] [CrossRef]
- Damgaard, P.D.B.; Martiniano, R.; Kamm, J.; Moreno-Mayar, J.V.; Kroonen, G.; Peyrot, M.; Barjamovic, G.; Rasmussen, S.; Zacho, C.; Baimukhanov, N.; et al. The first horse herders and the impact of early Bronze Age steppe expansions into Asia. Science 2018, 360, 7711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Järve, M.; Saag, L.; Scheib, C.L.; Pathak, A.K.; Montinaro, F.; Pagani, L.; Flores, R.; Guellil, M.; Saag, L.; Tambets, K.; et al. Shifts in the Genetic Landscape of the Western Eurasian Steppe Associated with the Beginning and End of the Scythian Dominance. Curr. Biol. 2019, 29, 2430–2441.e10. [Google Scholar] [CrossRef] [PubMed]
- Unterländer, M.; Palstra, F.; Lazaridis, I.; Pilipenko, A.; Hofmanová, Z.; Groß, M.; Sell, C.; Blöcher, J.; Kirsanow, K.; Rohland, N.; et al. Ancestry and demography and descendants of Iron Age nomads of the Eurasian Steppe. Nat. Commun. 2017, 8, 14615. [Google Scholar] [CrossRef]
- Damgaard, P.D.B.; Marchi, N.; Rasmussen, S.; Peyrot, M.; Renaud, G.; Korneliussen, T.; Moreno-Mayar, J.V.; Pedersen, M.W.; Goldberg, A.; Usmanova, E.; et al. 137 ancient human genomes from across the Eurasian steppes. Nature 2018, 557, 369–374. [Google Scholar] [CrossRef]
- Dani, A.H.; Masson, V.M. History of Civilizations of Central Asia, Vol. I: The Dawn of Civilization: Earliest Times to 700 B.C. History of Civilizations of Central Asia; UNESCO Publishing: Paris, France, 1999; Volume 1. [Google Scholar]
- Jettmar, K. The Karasuk Culture and its South-eastern Affinities. Bull. Mus. Far East. Antiq. 1950, 22, 83–126. [Google Scholar]
- Harmatta, J.; Puri, B.N.; Etemadi, G.F. History of Civilizations of Central Asia, Vol. II: The Development of Sedentary and Nomadic Civilizations: 700 B.C. to A.D. History of Civilizations of Central Asia; Motilal Banarsidass Publishers: Delhi, India, 1992; Volume 2. [Google Scholar]
- Thompson, E.A. The Huns; Wiley-Blackwell: Hoboken, NJ, USA, 1996; p. 340. [Google Scholar]
- Asimov, M.S.; Bosworth, C.E. History of Civilizations of Central Asia, Vol. IV: The Age of Achievement: A.D. 750 to the End of Fifteenth Century, Part One: The Historical, Social and Economic Setting. History of Civilizations of Central Asia; UNESCO Publishing: Paris, France, 1998; p. 485. [Google Scholar]
- Adle, C.; Habib, I.; Baipakov, K.M. History of Civilizations of Central Asia, Vol. V: Development in Contrast: From the Sixteenth to the Mid-Nineteenth Century. History of Civilizations of Central Asia; UNESCO Publishing: Paris, France, 1998; p. 485. [Google Scholar]
- Zhabagin, M.; Balanovska, E.; Sabitov, Z.; Kuznetsova, M.; Agdzhoyan, A.; Balaganskaya, O.; Chukhryaeva, M.; Markina, N.; Romanov, A.; Skhalyakho, R.; et al. The Connection of the Genetic, Cultural and Geographic Landscapes of Transoxiana. Sci. Rep. 2017, 7, 3085. [Google Scholar] [CrossRef] [Green Version]
- He, G.; Adnan, A.; Rakha, A.; Yeh, I.H.-Y.; Wang, M.; Zou, X.; Guo, J.; Rehman, M.; Fawad, A.; Chen, P.; et al. A comprehensive exploration of the genetic legacy and forensic features of Afghanistan and Pakistan Mongolian-descent Hazara. Forensic Sci. Int. Genet. 2019, 42, e1–e12. [Google Scholar] [CrossRef]
- Wells, R.S.; Yuldasheva, N.; Ruzibakiev, R.; Underhill, P.A.; Evseeva, I.; Blue-Smith, J.; Jin, L.; Su, B.; Pitchappan, R.; Shanmugalakshmi, S.; et al. The Eurasian Heartland: A continental perspective on Y-chromosome diversity. Proc. Natl. Acad. Sci. USA 2001, 98, 10244–10249. [Google Scholar] [CrossRef] [Green Version]
- Balaresque, P.; Poulet, N.; Cussat-Blanc, S.; Gerard, P.; Quintana-Murci, L.; Heyer, E.; Jobling, M.A. Y-chromosome descent clusters and male differential reproductive success: Young lineage expansions dominate Asian pastoral nomadic populations. Eur. J. Hum. Genet. 2015, 23, 1413–1422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balanovsky, O.; Zhabagin, M.; Agdzhoyan, A.; Chukhryaeva, M.; Zaporozhchenko, V.; Utevska, O.; Highnam, G.; Sabitov, Z.; Greenspan, E.; Dibirova, K.; et al. Deep Phylogenetic Analysis of Haplogroup G1 Provides Estimates of SNP and STR Mutation Rates on the Human Y-Chromosome and Reveals Migrations of Iranic Speakers. PLoS ONE 2015, 10, e0122968. [Google Scholar] [CrossRef] [Green Version]
- Di Cristofaro, J.; Pennarun, E.; Mazières, S.; Myres, N.M.; Lin, A.A.; Temori, S.A.; Metspalu, M.; Metspalu, E.; Witzel, M.; King, R.J.; et al. Afghan Hindu Kush: Where Eurasian Sub-Continent Gene Flows Converge. PLoS ONE 2013, 8, e76748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abilev, S.; Malyarchuk, B.; Derenko, M.; Wozniak, M.; Grzybowski, T.; Zakharov, I. The Y-chromosome C3* Star-Cluster Attributed to Genghis Khan’s Descendants is Present at High Frequency in the Kerey Clan from Kazakhstan. Hum. Biol. 2012, 84, 79–89. [Google Scholar] [CrossRef] [Green Version]
- Haber, M.; Platt, D.E.; Bonab, M.A.; Youhanna, S.C.; Soria-Hernanz, D.F.; Martínez-Cruz, B.; Douaihy, B.; Ghassibe-Sabbagh, M.; Rafatpanah, H.; Ghanbari, M.; et al. Afghanistan’s Ethnic Groups Share a Y-Chromosomal Heritage Structured by Historical Events. PLoS ONE 2012, 7, e34288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhabagin, M.; Sabitov, Z.; Tarlykov, P.; Tazhigulova, I.; Junissova, Z.; Yerezhepov, D.; Akilzhanov, R.; Zholdybayeva, E.; Wei, L.-H.; Akilzhanova, A.; et al. The medieval Mongolian roots of Y-chromosomal lineages from South Kazakhstan. BMC Genet. 2020, 21, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhabagin, M.; Sabitov, Z.; Tazhigulova, I.; Alborova, I.; Agdzhoyan, A.; Wei, L.-H.; Urasin, V.; Koshel, S.; Mustafin, K.; Akilzhanova, A.; et al. Medieval Super-Grandfather founder of Western Kazakh Clans from Haplogroup C2a1a2-M48. J. Hum. Genet. 2021, 66, 707–716. [Google Scholar] [CrossRef]
- Balanovsky, O.; Gurianov, V.; Zaporozhchenko, V.; Balaganskaya, O.; Urasin, V.; Zhabagin, M.; Grugni, V.; Canada, R.; Al-Zahery, N.; Raveane, A.; et al. Phylogeography of human Y-chromosome haplogroup Q3-L275 from an academic/citizen science collaboration. BMC Evol. Biol. 2017, 17, 18. [Google Scholar] [CrossRef] [Green Version]
- Zhabagin, M.K.; Dibirova, H.D.; Frolova, S.A.; Sabitov, Z. The Relation between the Y-Chromosomal Variation and the Clan Structure: The Gene Pool of the Steppe Aristocracy and the Steppe Clergy of the Kazakhs; Moscow University Anthropology Bulletin: Moscow, Russia, 2014; pp. 96–101. [Google Scholar]
- Dulik, M.C.; Osipova, L.P.; Schurr, T.G. Y-Chromosome Variation in Altaian Kazakhs Reveals a Common Paternal Gene Pool for Kazakhs and the Influence of Mongolian Expansions. PLoS ONE 2011, 6, e17548. [Google Scholar] [CrossRef]
- Turuspekov, Y.; Sabitov, Z.; Daulet, B.; Sadykov, M. The Kazakhstan DNA project hits first hundred Y-profiles for ethnic Kazakhs. Russ. J. Genet. Geneal. 2011, 2, 69–84. [Google Scholar]
- Malyarchuk, B.; Derenko, M.; Wozniak, M.; Grzybowski, T. Y-chromosome variation in Tajiks and Iranians. Ann. Hum. Biol. 2012, 40, 48–54. [Google Scholar] [CrossRef]
- Wen, S.-Q.; Du, P.-X.; Sun, C.; Cui, W.; Xu, Y.-R.; Meng, H.-L.; Shi, M.-S.; Zhu, B.-F.; Li, H. Dual origins of the Northwest Chinese Kyrgyz: The admixture of Bronze age Siberian and Medieval Niru’un Mongolian Y chromosomes. J. Hum. Genet. 2021, 67, 175–180. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics 2010, 26, 589–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows—Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karmin, M.; Saag, L.; Vicente, M.; Sayres, M.A.W.; Järve, M.; Talas, U.G.; Rootsi, S.; Ilumäe, A.-M.; Mägi, R.; Mitt, M.; et al. A recent bottleneck of Y chromosome diversity coincides with a global change in culture. Genome Res. 2015, 25, 459–466. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Li, Y.X.; Ma, P.-C.; Yan, S.; Cheng, H.-Z.; Fan, Z.-Q.; Deng, X.-H.; Ru, K.; Wang, C.-C.; Chen, G.; et al. Shared paternal ancestry of Han, Tai-Kadai-speaking, and Austronesian-speaking populations as revealed by the high resolution phylogeny of O1a-M119 and distribution of its sub-lineages within China. Am. J. Phys. Anthropol. 2021, 174, 686–700. [Google Scholar] [CrossRef]
- Sun, J.; Wei, L.; Wang, L.; Huang, Y.; Yan, S.; Cheng, H.; Ong, R.T.; Saw, W.; Fan, Z.; Deng, X.; et al. Paternal gene pool of Malays in Southeast Asia and its applications for the early expansion of Austronesians. Am. J. Hum. Biol. 2020, 33, 23486. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, X.; Li, J.; Li, H.; Zhao, Y.; Zhou, H. The Y-chromosome haplogroup C3*-F3918, likely attributed to the Mongol Empire, can be traced to a 2500-year-old nomadic group. J. Hum. Genet. 2017, 63, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y.; Zhao, Y.; Chen, Y.; Ochir, A.; Sarenbilige; Zhu, H.; Zhou, H. The genome of an ancient Rouran individual reveals an important paternal lineage in the Donghu population. Am. J. Phys. Anthr. 2018, 166, 895–905. [Google Scholar] [CrossRef]
- Wang, H.; Chen, L.; Ge, B.; Zhang, Y.; Zhu, H.; Zhou, H. Genetic data suggests that the Jinggouzi people are associated with the Donghu, an ancient nomadic group of North China. Hum. Biol. 2012, 84, 365–378. [Google Scholar] [CrossRef] [PubMed]
- Zerjal, T.; Xue, Y.; Bertorelle, G.; Wells, R.S.; Bao, W.; Zhu, S.; Qamar, R.; Ayub, Q.; Mohyuddin, A.; Fu, S.; et al. The Genetic Legacy of the Mongols. Am. J. Hum. Genet. 2003, 72, 717–721. [Google Scholar] [CrossRef] [Green Version]
- Wei, L.-H.; Yan, S.; Lu, Y.; Wen, S.-Q.; Huang, Y.-Z.; Wang, L.-X.; Li, S.-L.; Yang, Y.-J.; Wang, X.-F.; Zhang, C.; et al. Whole-sequence analysis indicates that the Y chromosome C2*-Star Cluster traces back to ordinary Mongols, rather than Genghis Khan. Eur. J. Hum. Genet. 2018, 26, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Malyarchuk, B.; Derenko, M.; Denisova, G.; Maksimov, A.; Woźniak, M.; Grzybowski, T.; Dambueva, I.; Zakharov, I. Ancient links between Siberians and Native Americans revealed by subtyping the Y chromosome haplogroup Q1a. J. Hum. Genet. 2011, 56, 583–588. [Google Scholar] [CrossRef]
- Huang, Y.-Z.; Pamjav, H.; Flegontov, P.; Stenzl, V.; Wen, S.-Q.; Tong, X.-Z.; Wang, C.-C.; Wang, L.-X.; Wei, L.-H.; Gao, J.-Y.; et al. Dispersals of the Siberian Y-chromosome haplogroup Q in Eurasia. Mol. Genet. Genom. 2018, 293, 107–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Underhill, P.A.; Poznik, G.D.; Rootsi, S.; Järve, M.; Lin, A.A.; Wang, J.; Passarelli, B.; Kanbar, J.; Myres, N.M.; King, R.J.; et al. The phylogenetic and geographic structure of Y-chromosome haplogroup R1a. Eur. J. Hum. Genet. 2014, 23, 124–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myres, N.M.; Rootsi, S.; Lin, A.A.; Järve, M.; King, R.J.; Kutuev, I.; Cabrera, V.M.; Khusnutdinova, E.; Pshenichnov, A.; Yunusbayev, B.; et al. A major Y-chromosome haplogroup R1b Holocene era founder effect in Central and Western Europe. Eur. J. Hum. Genet. 2010, 19, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Sengupta, S.; Zhivotovsky, L.A.; King, R.; Mehdi, S.; Edmonds, C.A.; Chow, C.-E.T.; Lin, A.A.; Mitra, M.; Sil, S.K.; Ramesh, A.; et al. Polarity and Temporality of High-Resolution Y-Chromosome Distributions in India Identify Both Indigenous and Exogenous Expansions and Reveal Minor Genetic Influence of Central Asian Pastoralists. Am. J. Hum. Genet. 2006, 78, 202–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, S.; Singh, A.; Rajkumar, R.; Kumar, K.S.; Samy, S.K.; Nizamuddin, S.; Singh, A.; Sheikh, S.A.; Peddada, V.; Khanna, V.; et al. Dissecting the influence of Neolithic demic diffusion on Indian Y-chromosome pool through J2-M172 haplogroup. Sci. Rep. 2016, 6, 19157. [Google Scholar] [CrossRef] [Green Version]
- Fu, Q.; Posth, C.; Hajdinjak, M.; Petr, M.; Mallick, S.; Fernandes, D.; Furtwängler, A.; Haak, W.; Meyer, M.; Mittnik, A.; et al. The genetic history of Ice Age Europe. Nature 2016, 534, 200–205. [Google Scholar] [CrossRef] [Green Version]
- Haak, W.; Lazaridis, I.; Patterson, N.; Rohland, N.; Mallick, S.; Llamas, B.; Brandt, G.; Nordenfelt, S.; Harney, E.; Stewardson, K.; et al. Massive migration from the steppe was a source for Indo-European languages in Europe. Nature 2015, 522, 207–211. [Google Scholar] [CrossRef] [Green Version]
- Allentoft, M.; Sikora, M.; Sjögren, K.-G.; Rasmussen, S.; Rasmussen, M.; Stenderup, J.; Damgaard, P.D.B.; Schroeder, H.; Ahlström, T.; Vinner, L.; et al. Population genomics of Bronze Age Eurasia. Nature 2015, 522, 167–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Y Chromosome Consortium. A nomenclature system for the tree of human Y-chromosomal binary haplogroups. Genome Res. 2002, 12, 339–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouckaert, R.R.; Drummond, A.J. bModelTest: Bayesian phylogenetic site model averaging and model comparison. BMC Evol. Biol. 2017, 17, 42. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, M.; Anzick, S.L.; Waters, M.R.; Skoglund, P.; DeGiorgio, M.; Stafford, T.W.; Rasmussen, S.; Moltke, I.; Albrechtsen, A.; Doyle, S.M.; et al. The genome of a Late Pleistocene human from a Clovis burial site in western Montana. Nature 2014, 506, 225–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree Download Page. 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 1 September 2022).




{kind=link}
{kind=link}
{kind=link}
| Haplogroup | Dungan | Hazara | Karakalpak | Karluk | Kazakhs | Kyrgyz | Tajik | Turkmen | Uyghur | Uzbek | Sum |
|---|---|---|---|---|---|---|---|---|---|---|---|
| C2a1a1b1-F1756 | 6 (0.3) | 2 (0.1) | 2 (0.1) | 10 | |||||||
| C2a1a2a-M48 | 2 (0.12) | 1 (0.05) | 1 (0.05) | 4 | |||||||
| C2a1a3-M504 | 3 (0.15) | 5 (0.29) | 10 (0.45) | 3 (0.15) | 3 (0.14) | 1 (0.05) | 25 | ||||
| C2b*-F1067xM407 | 1 (0.05) | 1 (0.05) | 2 | ||||||||
| C2b1a1a1a-M407 | 2 (0.09) | 1 (0.05) | 3 | ||||||||
| D1a1a2-N1 | 2 (0.1) | 1 (0.05) | 3 | ||||||||
| D1a2a-P47 | 1 (0.05) | 1 (0.05) | 2 | ||||||||
| E1b1b1-M35.1 | 1 (0.06) | 1 (0.05) | 1 (0.05) | 3 | |||||||
| G-M201 | 1 (0.05) | 1 (0.06) | 1 (0.45) | 1 (0.05) | 2 (0.09) | 6 | |||||
| H-L901 | 1 (0.05) | 1 (0.05) | 2 | ||||||||
| I-M170 | 1 (0.05) | 1 (0.45) | 1 (0.05) | 3 | |||||||
| J1a-CTS5368 | 1 (0.06) | 1 (0.45) | 2 (0.1) | 1 (0.05) | 1 (0.05) | 6 | |||||
| J2a-M410 | 4 (0.2) | 4 (0.2) | 2 (0.12) | 2 (0.1) | 3 (0.15) | 4 (0.2) | 1 (0.05) | 5 (0.25) | 25 | ||
| L-M20 | 1 (0.05) | 9 (1) | 1 (0.05) | 1 (0.05) | 1 (0.05) | 13 | |||||
| N-M231 | 1 (0.05) | 1 (0.06) | 4 (0.18) | 1 (0.05) | 1 (0.05) | 1 (0.05) | 1 (0.05) | 10 | |||
| O-M175 | 6 (0.3) | 3 (0.18) | 3 (0.14) | 4 (0.2) | 16 | ||||||
| Q-M242 | 4 (0.2) | 2 (0.09) | 2 (0.1) | 3 (0.15) | 2 (0.09) | 13 | |||||
| R1a1a-M17 | 1 (0.05) | 3 (0.15) | 1 (0.06) | 11 (0.55) | 7 (0.35) | 1 (0.05) | 4 (0.18) | 2 (0.1) | 30 | ||
| R1b-M343 | 1 (0.05) | 1 (0.45) | 1 (0.05) | 2 (0.1) | 1 (0.05) | 1 (0.05) | 7 | ||||
| R2a-M124 | 3 (0.15) | 1 (0.05) | 4 | ||||||||
| Sum | 20 | 20 | 17 | 9 | 22 | 20 | 20 | 20 | 19 | 20 | 187 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhabagin, M.; Wei, L.-H.; Sabitov, Z.; Ma, P.-C.; Sun, J.; Dyussenova, Z.; Balanovska, E.; Li, H.; Ramankulov, Y. Ancient Components and Recent Expansion in the Eurasian Heartland: Insights into the Revised Phylogeny of Y-Chromosomes from Central Asia. Genes 2022, 13, 1776. https://doi.org/10.3390/genes13101776
Zhabagin M, Wei L-H, Sabitov Z, Ma P-C, Sun J, Dyussenova Z, Balanovska E, Li H, Ramankulov Y. Ancient Components and Recent Expansion in the Eurasian Heartland: Insights into the Revised Phylogeny of Y-Chromosomes from Central Asia. Genes. 2022; 13(10):1776. https://doi.org/10.3390/genes13101776
Chicago/Turabian StyleZhabagin, Maxat, Lan-Hai Wei, Zhaxylyk Sabitov, Peng-Cheng Ma, Jin Sun, Zhanargul Dyussenova, Elena Balanovska, Hui Li, and Yerlan Ramankulov. 2022. "Ancient Components and Recent Expansion in the Eurasian Heartland: Insights into the Revised Phylogeny of Y-Chromosomes from Central Asia" Genes 13, no. 10: 1776. https://doi.org/10.3390/genes13101776