
Rare Heterozygous PCSK1 Variants in Human Obesity: The Contribution of the p.Y181H Variant and a Literature Review
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Genotyping
2.3. Haplotype Analysis
2.4. Statistical Analysis
2.5. Literature Search
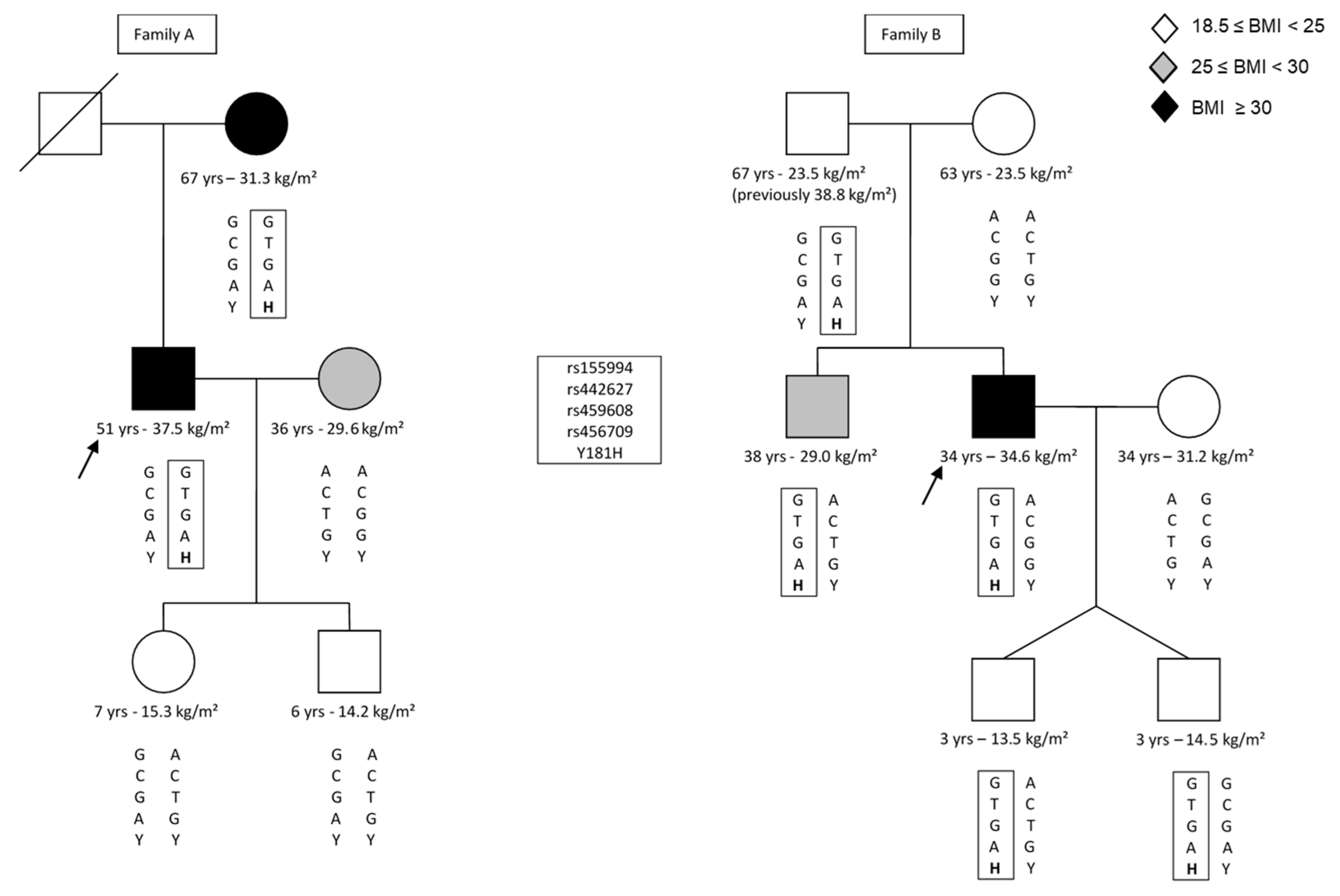
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organisation. Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 3 March 2020).
- Beckers, S.; Zegers, D.; Van Gaal, L.F.; Van Hul, W. The role of the leptin-melanocortin signalling pathway in the control of food intake. Crit. Rev. Eukaryot. Gene Expr. 2009, 19, 267–287. [Google Scholar] [CrossRef] [PubMed]
- Benjannet, S.; Rondeau, N.; Day, R.; Chrétien, M.; Seidah, N.G. PC1 and PC2 are proprotein convertases capable of cleaving proopiomelanocortin at distinct pairs of basic residues. Proc. Natl. Acad. Sci. USA 1991, 88, 3564–3568. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Bloomquist, B.T.; Mains, R.E. The prohormone convertases PC1 and PC2 mediate distinct endoproteolytic cleavages in a strict temporal order during proopiomelanocortin biosynthetic processing. J. Biol. Chem. 1993, 268, 1763–1769. [Google Scholar] [CrossRef]
- O’Rahilly, S.; Gray, H.; Humphreys, P.J.; Krook, A.; Polonsky, K.S.; White, A.; Gibson, S.; Taylor, K.; Carr, C. Brief report: Impaired processing of prohormones associated with abnormalities of glucose homeostasis and adrenal function. New Engl. J. Med. 1995, 333, 1386–1390. [Google Scholar] [CrossRef]
- Jackson, R.S.; Creemers, J.W.; Ohagi, S.; Raffin-Sanson, M.L.; Sanders, L.; Montague, C.T.; Hutton, J.C.; O’Rahilly, S. Obesity and impaired prohormone processing associated with mutations in the human prohormone convertase 1 gene. Nat. Genet. 1997, 16, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Pepin, L.; Colin, E.; Tessarech, M.; Rouleau, S.; Bouhours-Nouet, N.; Bonneau, D.; Coutant, R. A New Case of PCSK1 Pathogenic Variant With Congenital Proprotein Convertase 1/3 Deficiency and Literature Review. J. Clin. Endocrinol. Metab. 2019, 104, 985–993. [Google Scholar] [CrossRef]
- Serra, E.G.; Schwerd, T.; Moutsianas, L.; Cavounidis, A.; Fachal, L.; Pandey, S.; Kammermeier, J.; Croft, N.M.; Posovszky, C.; Rodrigues, A.; et al. Somatic mosaicism and common genetic variation contribute to the risk of very-early-onset inflammatory bowel disease. Nat. Commun. 2020, 11, 995. [Google Scholar] [CrossRef]
- Karacan Küçükali, G.; Savaş Erdeve, Ş.; Çetinkaya, S.; Keskin, M.; Buluş, A.D.; Aycan, Z. A case of prohormone convertase deficiency diagnosed with type 2 diabetes. Turk. Arch. Pediatr 2021, 56, 81–84. [Google Scholar] [CrossRef]
- Duclaux-Loras, R.; Bourgeois, P.; Lavrut, P.M.; Charbit-Henrion, F.; Bonniaud-Blot, P.; Maudinas, R.; Bournez, M.; Faure, M.; Cerf-Bensussan, N.; Lachaux, A.; et al. A novel mutation of PCSK1 responsible for PC1/3 deficiency in two siblings. Clin. Res. Hepatol. Gastroenterol. 2021, 45, 101640. [Google Scholar] [CrossRef]
- Ahmed, A.B.M.; Alsaleem, B.M.R. Enteroendocrine Dysfunction in Two Saudi Sisters. Case Rep. Gastroenterol. 2021, 15, 290–295. [Google Scholar] [CrossRef]
- Qian, Y.; Wu, B.; Liu, R.; Lu, Y.; Zhang, P.; Shao, C.; Huang, Y.; Wang, H. Case Report: Complete Maternal Uniparental Isodisomy of Chromosome 5 (iUPD(5)mat) With PCSK1 Nonsense Variant in an Infant With Recurrent Diarrhea. Front. Genet. 2021, 12, 668326. [Google Scholar] [CrossRef] [PubMed]
- Aerts, L.; Terry, N.A.; Sainath, N.N.; Torres, C.; Martín, M.G.; Ramos-Molina, B.; Creemers, J.W. Novel Homozygous Inactivating Mutation in the PCSK1 Gene in an Infant with Congenital Malabsorptive Diarrhea. Genes 2021, 12, 710. [Google Scholar] [CrossRef] [PubMed]
- Courbage, S.; Poitou, C.; Le Beyec-Le Bihan, J.; Karsenty, A.; Lemale, J.; Pelloux, V.; Lacorte, J.M.; Carel, J.C.; Lecomte, N.; Storey, C.; et al. Implication of Heterozygous Variants in Genes of the Leptin-Melanocortin Pathway in Severe Obesity. J. Clin. Endocrinol. Metab. 2021, 106, 2991–3006. [Google Scholar] [CrossRef]
- Jackson, R.S.; Creemers, J.W.; Farooqi, I.S.; Raffin-Sanson, M.L.; Varro, A.; Dockray, G.J.; Holst, J.J.; Brubaker, P.L.; Corvol, P.; Polonsky, K.S.; et al. Small-intestinal dysfunction accompanies the complex endocrinopathy of human proprotein convertase 1 deficiency. J. Clin. Investig. 2003, 112, 1550–1560. [Google Scholar] [CrossRef]
- Farooqi, I.S.; Keogh, J.M.; Yeo, G.S.; Lank, E.J.; Cheetham, T.; O’Rahilly, S. Clinical spectrum of obesity and mutations in the melanocortin 4 receptor gene. N. Engl. J. Med. 2003, 348, 1085–1095. [Google Scholar] [CrossRef] [PubMed]
- Frank, G.R.; Fox, J.; Candela, N.; Jovanovic, Z.; Bochukova, E.; Levine, J.; Papenhausen, P.R.; O’Rahilly, S.; Farooqi, I.S. Severe obesity and diabetes insipidus in a patient with PCSK1 deficiency. Mol. Genet. Metab. 2013, 110, 191–194. [Google Scholar] [CrossRef]
- Martín, M.G.; Lindberg, I.; Solorzano-Vargas, R.S.; Wang, J.; Avitzur, Y.; Bandsma, R.; Sokollik, C.; Lawrence, S.; Pickett, L.A.; Chen, Z.; et al. Congenital proprotein convertase 1/3 deficiency causes malabsorptive diarrhea and other endocrinopathies in a pediatric cohort. Gastroenterology 2013, 145, 138–148. [Google Scholar] [CrossRef]
- Gönç, E.N.; Özön, A.; Alikaşifoğlu, A.; Kandemir, N. Long-Term Follow-up of a Case with Proprotein Convertase 1/3 Deficiency: Transient Diabetes Mellitus with Intervening Diabetic Ketoacidosis During Growth Hormone Therapy. J. Clin. Res. Pediatric Endocrinol. 2017, 9, 283–287. [Google Scholar] [CrossRef]
- Bandsma, R.H.; Sokollik, C.; Chami, R.; Cutz, E.; Brubaker, P.L.; Hamilton, J.K.; Perlman, K.; Zlotkin, S.; Sigalet, D.L.; Sherman, P.M.; et al. From diarrhea to obesity in prohormone convertase 1/3 deficiency: Age-dependent clinical, pathologic, and enteroendocrine characteristics. J. Clin. Gastroenterol. 2013, 47, 834–843. [Google Scholar] [CrossRef]
- Yourshaw, M.; Solorzano-Vargas, R.S.; Pickett, L.A.; Lindberg, I.; Wang, J.; Cortina, G.; Pawlikowska-Haddal, A.; Baron, H.; Venick, R.S.; Nelson, S.F.; et al. Exome sequencing finds a novel PCSK1 mutation in a child with generalized malabsorptive diarrhea and diabetes insipidus. J. Pediatric Gastroenterol. Nutr. 2013, 57, 759–767. [Google Scholar] [CrossRef] [Green Version]
- Wilschanski, M.; Abbasi, M.; Blanco, E.; Lindberg, I.; Yourshaw, M.; Zangen, D.; Berger, I.; Shteyer, E.; Pappo, O.; Bar-Oz, B.; et al. A novel familial mutation in the PCSK1 gene that alters the oxyanion hole residue of proprotein convertase 1/3 and impairs its enzymatic activity. PLoS ONE 2014, 9, e108878. [Google Scholar] [CrossRef]
- Härter, B.; Fuchs, I.; Müller, T.; Akbulut, U.E.; Cakir, M.; Janecke, A.R. Early Clinical Diagnosis of PC1/3 Deficiency in a Patient With a Novel Homozygous PCSK1 Splice-Site Mutation. J. Pediatric Gastroenterol. Nutr. 2016, 62, 577–580. [Google Scholar] [CrossRef]
- Kumar, G.; Nair, R.; Sohal, A.P.S. Proprotein Convertase 1/3 Deficiency. Indian J. Pediatrics 2018, 85, 320–321. [Google Scholar] [CrossRef] [PubMed]
- Distelmaier, F.; Herebian, D.; Atasever, C.; Beck-Woedl, S.; Mayatepek, E.; Strom, T.M.; Haack, T.B. Blue Diaper Syndrome and PCSK1 Mutations. Pediatrics 2018, 141, S501–S505. [Google Scholar] [CrossRef]
- Creemers, J.W.M.; Choquet, H.; Stijnen, P.; Vatin, V.; Pigeyre, M.; Beckers, S.; Meulemans, S.; Than, M.E.; Yengo, L.; Tauber, M.; et al. Heterozygous mutations causing partial prohormone convertase 1 deficiency contribute to human obesity. Diabetes 2012, 61, 383–390. [Google Scholar] [CrossRef]
- Hauspie, R.R.M. Flemish Growth Charts. 2004. Available online: http://www.vub.ac.be/groeicurven (accessed on 22 July 2013).
- Roelants, M.; Hauspie, R.; Hoppenbrouwers, K. References for growth and pubertal development from birth to 21 years in Flanders, Belgium. Ann. Hum. Biol. 2009, 36, 680–694. [Google Scholar] [CrossRef]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef]
- Becker, T.; Knapp, M. Maximum-likelihood estimation of haplotype frequencies in nuclear families. Genet. Epidemiol. 2004, 27, 21–32. [Google Scholar] [CrossRef]
- Herold, C.; Becker, T. Genetic association analysis with FAMHAP: A major program update. Bioinformatics 2009, 25, 134–136. [Google Scholar] [CrossRef]
- Menashe, I.; Rosenberg, P.S.; Chen, B.E. PGA: Power calculator for case-control genetic association analyses. BMC Genet. 2008, 9, 36. [Google Scholar] [CrossRef] [Green Version]
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alföldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Philippe, J.; Stijnen, P.; Meyre, D.; De Graeve, F.; Thuillier, D.; Delplanque, J.; Gyapay, G.; Sand, O.; Creemers, J.W.; Froguel, P.; et al. A nonsense loss-of-function mutation in PCSK1 contributes to dominantly inherited human obesity. Int. J. Obes. 2015, 39, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Nordang, G.B.N.; Busk, Ø.L.; Tveten, K.; Hanevik, H.I.; Fell, A.K.M.; Hjelmesæth, J.; Holla, Ø.L.; Hertel, J.K. Next-generation sequencing of the monogenic obesity genes LEP, LEPR, MC4R, PCSK1 and POMC in a Norwegian cohort of patients with morbid obesity and normal weight controls. Mol. Genet. Metab. 2017, 121, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Kleinendorst, L.; Massink, M.P.G.; Cooiman, M.I.; Savas, M.; van der Baan-Slootweg, O.H.; Roelants, R.J.; Janssen, I.C.M.; Meijers-Heijboer, H.J.; Knoers, N.; Ploos van Amstel, H.K.; et al. Genetic obesity: Next-generation sequencing results of 1230 patients with obesity. J. Med. Genet. 2018, 55, 578–586. [Google Scholar] [CrossRef] [PubMed]
- Saeed, S.; Janjua, Q.M.; Haseeb, A.; Khanam, R.; Durand, E.; Vaillant, E.; Ning, L.; Badreddine, A.; Berberian, L.; Boissel, M.; et al. Rare Variant Analysis of Obesity-Associated Genes in Young Adults With Severe Obesity From a Consanguineous Population of Pakistan. Diabetes 2022, 71, 694–705. [Google Scholar] [CrossRef]
- Hendricks, A.E.; Bochukova, E.G.; Marenne, G.; Keogh, J.M.; Atanassova, N.; Bounds, R.; Wheeler, E.; Mistry, V.; Henning, E.; Körner, A.; et al. Rare Variant Analysis of Human and Rodent Obesity Genes in Individuals with Severe Childhood Obesity. Sci. Rep. 2017, 7, 4394. [Google Scholar] [CrossRef]
- Serra-Juhé, C.; Martos-Moreno, G.; Bou de Pieri, F.; Flores, R.; Chowen, J.A.; Pérez-Jurado, L.A.; Argente, J. Heterozygous rare genetic variants in non-syndromic early-onset obesity. Int. J. Obes. 2020, 44, 830–841. [Google Scholar] [CrossRef]
- Akbari, P.; Gilani, A.; Sosina, O.; Kosmicki, J.A.; Khrimian, L.; Fang, Y.Y.; Persaud, T.; Garcia, V.; Sun, D.; Li, A.; et al. Sequencing of 640,000 exomes identifies GPR75 variants associated with protection from obesity. Science 2021, 373, eabf8683. [Google Scholar] [CrossRef]
- Curtis, D. Weighted burden analysis in 200 000 exome-sequenced UK Biobank subjects characterises effects of rare genetic variants on BMI. Int. J. Obes. 2022, 46, 782–792. [Google Scholar] [CrossRef]
- AbouHashem, N.; Zaied, R.E.; Al-Shafai, K.; Nofal, M.; Syed, N.; Al-Shafai, M. The Spectrum of Genetic Variants Associated with the Development of Monogenic Obesity in Qatar. Obes. Facts 2022, 15, 357–365. [Google Scholar] [CrossRef]
- Wade, K.H.; Lam, B.Y.H.; Melvin, A.; Pan, W.; Corbin, L.J.; Hughes, D.A.; Rainbow, K.; Chen, J.H.; Duckett, K.; Liu, X.; et al. Loss-of-function mutations in the melanocortin 4 receptor in a UK birth cohort. Nat. Med. 2021, 27, 1088–1096. [Google Scholar] [CrossRef] [PubMed]
- Chami, N.; Preuss, M.; Walker, R.W.; Moscati, A.; Loos, R.J.F. The role of polygenic susceptibility to obesity among carriers of pathogenic mutations in MC4R in the UK Biobank population. PLoS Med. 2020, 17, e1003196. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| ID | c.Mutation ° | p.Mutation ° | Mutation Type ° | Obesity | Carriers in Family | Health Information of Carriers in Family | References |
|---|---|---|---|---|---|---|---|
| 1 | c.1777G>A/c.620 +4A>C | p.Gly593Arg/NA | Missense/Splice Site | yes (<3 years) | 4 heterozygous childrenBREAK(3x c.1777G>A, 1x c.620 +4A>C) | c.1777G>A: “All unaffected”BREAKc.620 +4A>C: NA | [5,6] |
| 2 | c.748G>T/c.638_640delCAG | p.Glu250*/p.Ala213del | Nonsense/Nonsense | Yes (<18 month) | Heterozygous parents BREAK(mother: c.748G>T, father: c.638_640delCAG) | both “clinically normal” | [15] |
| 3 | c.920C>T | p.Ser307Leu | Missense | yes | Heterozygous parents | “not obese” | [16] |
| 4 | c.1024delT/c.-775208_*59002del | n/a | Frameshift/Gene Deletion | yes (2 years) | heterozygous parents BREAK(mother: frameshift, father: gene deletion) | “normal weight” | [17] |
| 5 | c.1777G>A | p.Gly593Arg | Missense | yes (7.7 years) | heterozygous parents | NA | [18] |
| 6 | c.625G>A/c.772C>A | p.Gly209Arg/p.Pro258Thr | Missense | NA | NA | NA | [18] |
| 7 | c.1095+1G>T | NA | Splice Site | yes (4.3 years) | heterozygous parents | NA | [18,19] |
| 8 | c.1009C>T | PGln337* | Nonsense | no (died 8 month) | heterozygous parents | NA | [18] |
| 9, 10 | c.1213C>T | p.Arg405* | Nonsense | yes (3.7 years and 9.3 years) | NA | NA | [18,20] |
| 11,12 | c.1_2delATinsTA | p.Met1* | Nonsense | yes (2.9 years and 12.8 years) | NA | NA | [18,20] |
| 13 | c.1095+1G>A | n/a | Splice Site | no (died 15 month) | heterozygous parents | NA | [18] |
| 14 | c.1348_1353del | p.Val450Valfs*1 | Frameshift | no (5.5 years) | heterozygous parents, 2 heterozygous siblings | NA | [18] |
| 15 | c.1643T>C | p.Phe548Ser | Missense | yes (3.5 years) | NA | NA | [18] |
| 16 | c.693C>A | p.Tyr231* | Nonsense | yes (2.4 years) | heterozygous parents, heterozygous sibling | NA | [18] |
| 17 | c.1269C>G | p.Asn423Lys | Missense | yes (17 years) | NA | NA | [18] |
| 18 | c.1029C>A | p.Tyr343* | Nonsense | NA (polyphagia) | NA | NA | [21] |
| 19, 20, 21 | c.927C>G | p.Asn309Lys | Missense | yes (6 years), no (12 month and died 5 month) | Heterozygous parents | “healthy parents” | [22] |
| 22 | c.544-2A>G | n/a | Splice Site | yes (12 month) | heterozygous parents | “healthy parents” | [23] |
| 23, 24 | c.1323C>T | p.Arg438* | Nonsense | yes (2 years and 4 years) | NA | NA | [24] |
| 25 | c.679del | p.Val227Leufs*12 | Frameshift | yes (9 month) | heterozygous parents | mother BMI 27.5, father BMI 26.1 | [25] |
| 26 | c.595C>T | p.Arg199* | Nonsense | yes (3 years) | heterozygous parents | mother BMI 31.6 father 23.4 | [7] |
| 27 | NA | p.Arg391* | Nonsense | NA (“excessive weight gain”) | NA | NA | [8] |
| 28 | c.685G>T | p.V229F | Missense | yes (14 years) | NA | “obese mother” | [9] |
| 29, 30 | c.500A>C | p.Asp167Ala | Missense | yes (1.5 years and 4.5 years) | heterozygous parents | NA | [10] |
| 31, 32 | c.1350_1353del | p.Asp451fs | Frameshift | no (6 years), yes (12 years) | NA | NA | [11] |
| 33 | c.238C>T | p.Arg80* | Frameshift | no (11 month, weightgain) | heterozygous mother | mother BMI 25.4 | [12] |
| 34 | c.1034A>C | p.E345A | Missense | no (14 month, weightgain) | NA | NA | [13] |
| 35 | c.286-2A>G | n/a | Splice Site | yes | NA | NA | [14] |
| Parameter | Lean Controls | Children/Adolescents with Obesity |
|---|---|---|
| N | 486 | 481 |
| Men/women | 167/319 | 211/270 |
| Age (years) | 35 ± 7 | 12 ± 4 |
| BMI (kg/m²) | 22.1 ± 1.6 | 30.9 ± 5.8 |
| BMI z-score | n/a | 2.59 ± 0.54 |
| Overweight (%) | n/a | 172 ± 25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van Dijck, E.; Beckers, S.; Diels, S.; Huybrechts, T.; Verrijken, A.; Van Hoorenbeeck, K.; Verhulst, S.; Massa, G.; Van Gaal, L.; Van Hul, W. Rare Heterozygous PCSK1 Variants in Human Obesity: The Contribution of the p.Y181H Variant and a Literature Review. Genes 2022, 13, 1746. https://doi.org/10.3390/genes13101746
Van Dijck E, Beckers S, Diels S, Huybrechts T, Verrijken A, Van Hoorenbeeck K, Verhulst S, Massa G, Van Gaal L, Van Hul W. Rare Heterozygous PCSK1 Variants in Human Obesity: The Contribution of the p.Y181H Variant and a Literature Review. Genes. 2022; 13(10):1746. https://doi.org/10.3390/genes13101746
Chicago/Turabian StyleVan Dijck, Evelien, Sigri Beckers, Sara Diels, Tammy Huybrechts, An Verrijken, Kim Van Hoorenbeeck, Stijn Verhulst, Guy Massa, Luc Van Gaal, and Wim Van Hul. 2022. "Rare Heterozygous PCSK1 Variants in Human Obesity: The Contribution of the p.Y181H Variant and a Literature Review" Genes 13, no. 10: 1746. https://doi.org/10.3390/genes13101746