
HD-ZIP Gene Family: Potential Roles in Improving Plant Growth and Regulating Stress-Responsive Mechanisms in Plants


 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
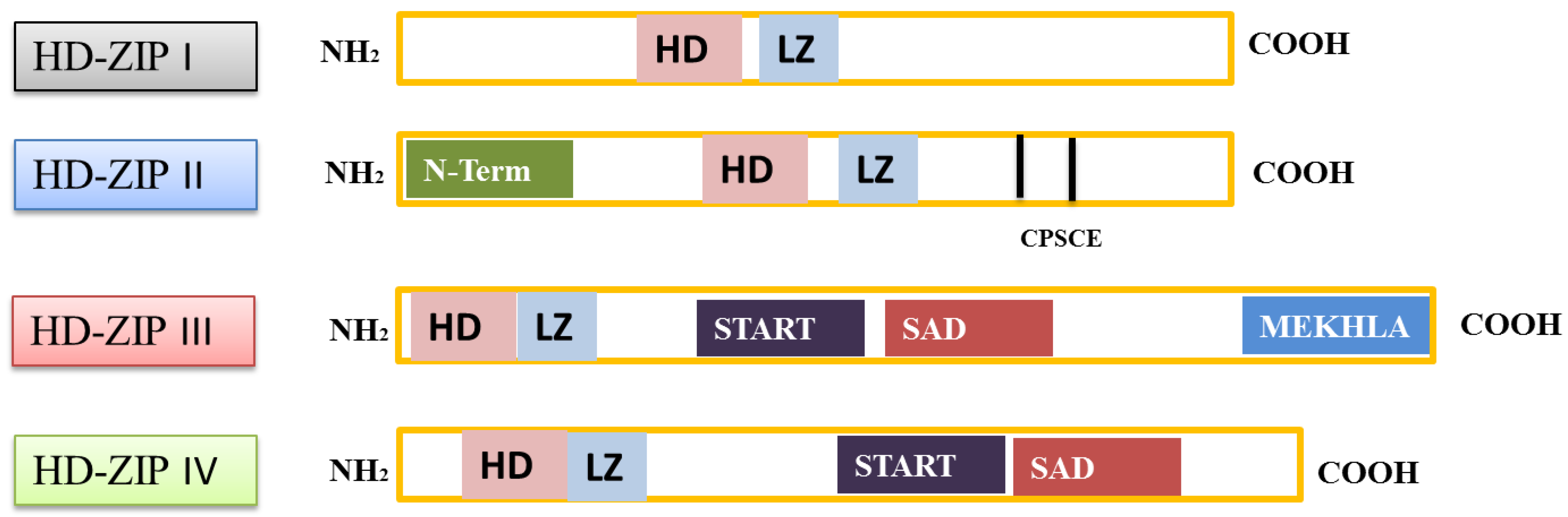
2. Structural Characteristics of HD-ZIP Gene Family
3. Role of HD-ZIP Genes Family in Plant Growth and Regulation
4. The Crucial Role of HD-ZIP Gene Family in Regulating Abiotic Stress
4.1. Role of HD-ZIP I Subfamily in Abiotic Stress Control
4.1.1. Drought Stress
4.1.2. Salinity Stress
4.1.3. Low-Temperature Stress
4.1.4. Heavy Metal Stress
4.1.5. Heat Stress
4.1.6. Flooding Stress
4.1.7. Nutrient Stress
4.2. Role of HD-ZIP II Subfamily in Abiotic Stress Control
4.2.1. Drought Stress
4.2.2. Light Stress
4.2.3. Salinity Stress
4.3. Role of HD-ZIP III Subfamily in Abiotic Stress Control
4.3.1. Drought Stress
4.3.2. Salinity Stress
4.3.3. Heavy Metal Stress
4.4. Role of HD-ZIP IV Subfamily in Abiotic Stress Control
4.4.1. Drought Stress
4.4.2. Salt Stress
4.4.3. Osmotic Stress
5. Role of HD-ZIP Gene Family in Regulating Biotic Stress
5.1. HD-ZIP I: Role in Coping Biotic Stress
5.2. HD-ZIP II: Role in Coping Biotic Stress
5.3. HD-ZIP III: Role in Coping Biotic Stress
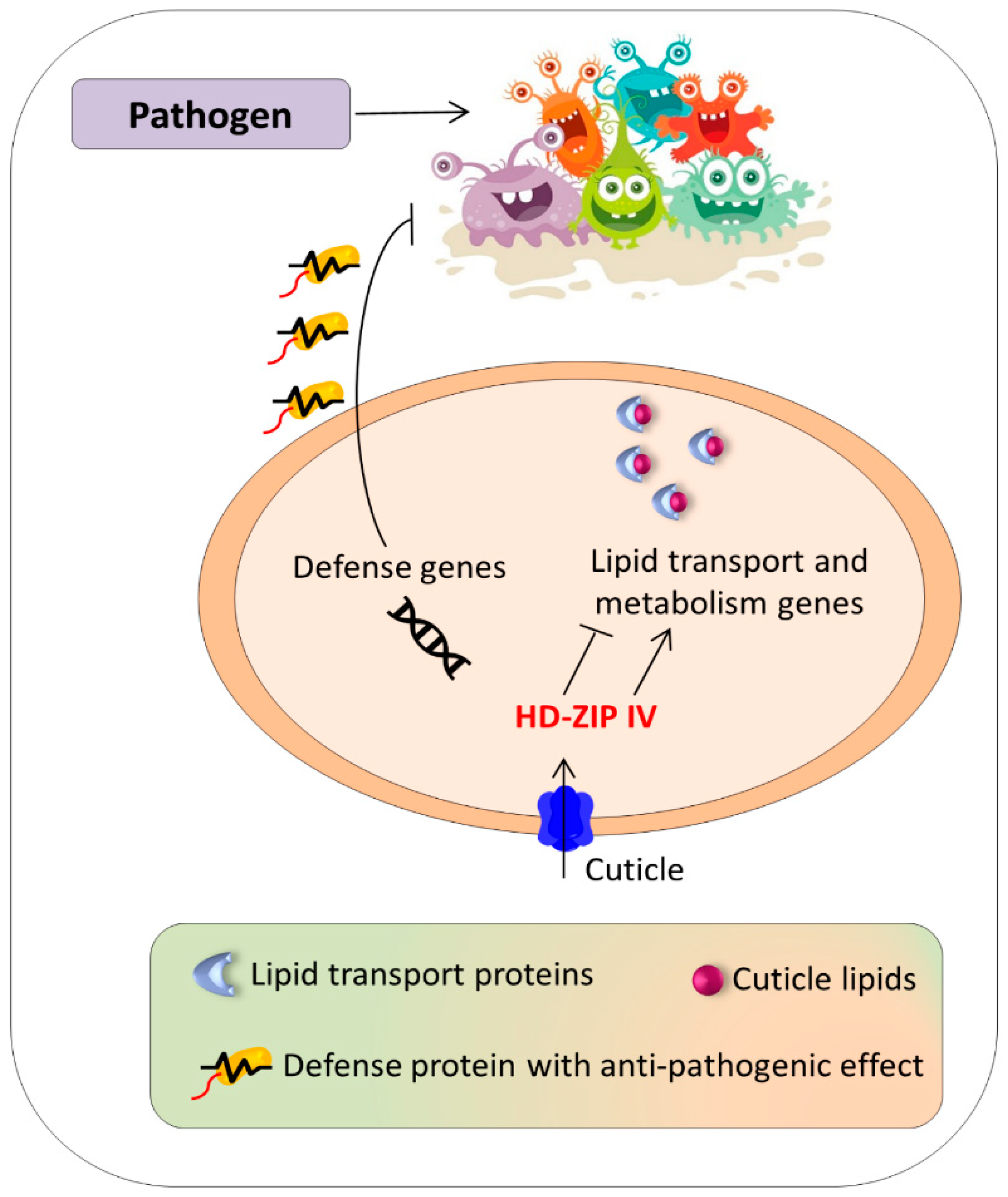
5.4. HD-ZIP IV: Role in Coping Biotic Stress
6. Conclusions and Future Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Garber, R.; Kuroiwa, A.; Gehring, W.J. Genomic and cDNA clones of the homeotic locus Antennapedia in Drosophila. EMBO J. 1983, 2, 2027–2036. [Google Scholar] [CrossRef]
- Ariel, F.D.; Manavella, P.A.; Dezar, C.A.; Chan, R.L. The true story of the HD-Zip family. Trends Plant Sci. 2007, 12, 419–426. [Google Scholar] [CrossRef]
- Moens, C.B.; Selleri, L. Hox cofactors in vertebrate development. Dev. Biol. 2006, 291, 193–206. [Google Scholar] [CrossRef] [Green Version]
- Vollbrecht, E.; Veit, B.; Sinha, N.; Hake, S. The developmental gene Knotted-1 is a member of a maize homeobox gene family. Nature 1991, 350, 241–243. [Google Scholar] [CrossRef] [PubMed]
- Bharathan, G.; Janssen, B.-J.; Kellogg, E.A.; Sinha, N. Did homeodomain proteins duplicate before the origin of angiosperms, fungi, and metazoa? Proc. Natl. Acad. Sci. USA 1997, 94, 13749–13753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Q.; Gao, Y.; Li, J.; Yang, Q.; Qu, X.; Li, H.; Zhang, J.; Wang, T.; Ye, Z.; Yang, C. The HD-Zip IV transcription factor SlHDZIV8 controls multicellular trichome morphology by regulating the expression of Hairless-2. J. Exp. Bot. 2020, 71, 7132–7145. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Ye, J.; Tao, P.; Li, H.; Zhang, J.; Zhang, Y.; Ye, Z. The tomato HD-Zip I transcription factor SlHZ24 modulates ascorbate accumulation through positive regulation of the d-mannose/l-galactose pathway. Plant J. 2016, 85, 16–29. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.-Z.; Chang, X.; Donnelly, L.; Reid, M.S.; Jiang, C.-Z. A HD-ZIP Transcription Factor Regulates Flower Senescence via Ethylene and ABA Cross-talks in Petunia. In Hortscience; 113 S WEST ST, STE 200; American Society Horticultural Science: Alexandria, VA, USA, 2014; p. S138. [Google Scholar]
- Li, S.; Chen, N.; Li, F.; Mei, F.; Wang, Z.; Cheng, X.; Kang, Z.; Mao, H. Characterization of wheat homeodomain-leucine zipper family genes and functional analysis of TaHDZ5-6A in drought tolerance in transgenic Arabidopsis. BMC Plant Biol. 2020, 20, 50. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.-K.; Yoon, J.; Choi, G.J.; Jang, H.A.; Kwon, S.-Y.; Choi, D. Capsicum annuum homeobox 1 (CaHB1) is a nuclear factor that has roles in plant development, salt tolerance, and pathogen defense. Biochem. Biophys. Res. Commun. 2013, 442, 116–121. [Google Scholar] [CrossRef]
- Ribichich, K.F.; Chiozza, M.; Ávalos-Britez, S.; Cabello, J.V.; Arce, A.L.; Watson, G.; Arias, C.; Portapila, M.; Trucco, F.; Otegui, M.E. Successful field performance in warm and dry environments of soybean expressing the sunflower transcription factor HaHB4. J. Exp. Bot. 2020, 71, 3142–3156. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Gong, S.; Wang, Y.; Wang, F.; Bao, H.; Sun, J.; Cai, C.; Yi, K.; Chen, Z.; Zhu, C. MicroRNA166 modulates cadmium tolerance and accumulation in rice. Plant Physiol. 2018, 177, 1691–1703. [Google Scholar] [CrossRef] [Green Version]
- Elhiti, M.; Stasolla, C. Structure and function of homodomain-leucine zipper (HD-Zip) proteins. Plant Signal. Behav. 2009, 4, 86–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harris, J.C.; Hrmova, M.; Lopato, S.; Langridge, P. Modulation of plant growth by HD-Zip class I and II transcription factors in response to environmental stimuli. New Phytol. 2011, 190, 823–837. [Google Scholar] [CrossRef]
- Henriksson, E.; Olsson, A.S.; Johannesson, H.; Johansson, H.; Hanson, J.; Engström, P.; Söderman, E. Homeodomain leucine zipper class I genes in Arabidopsis. Expression patterns and phylogenetic relationships. Plant Physiol. 2005, 139, 509–518. [Google Scholar] [CrossRef] [Green Version]
- Agalou, A.; Purwantomo, S.; Övernäs, E.; Johannesson, H.; Zhu, X.; Estiati, A.; de Kam, R.J.; Engström, P.; Slamet-Loedin, I.H.; Zhu, Z. A genome-wide survey of HD-Zip genes in rice and analysis of drought-responsive family members. Plant Mol. Biol. 2008, 66, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Tron, A.E.; Bertoncini, C.W.; Chan, R.L.; Gonzalez, D.H. Redox regulation of plant homeodomain transcription factors. J. Biol. Chem. 2002, 277, 34800–34807. [Google Scholar] [CrossRef] [Green Version]
- Tron, A.E.; Comelli, R.N.; Gonzalez, D.H. Structure of homeodomain-leucine zipper/DNA complexes studied using hydroxyl radical cleavage of DNA and methylation interference. Biochemistry 2005, 44, 16796–16803. [Google Scholar] [CrossRef]
- Palena, C.M.; Tron, A.E.; Bertoncini, C.W.; Gonzalez, D.H.; Chan, R.L. Positively charged residues at the N-terminal arm of the homeodomain are required for efficient DNA binding by homeodomain-leucine zipper proteins. J. Mol. Biol. 2001, 308, 39–47. [Google Scholar] [CrossRef]
- Tron, A.E.; Welchen, E.; Gonzalez, D.H. Engineering the loop region of a homeodomain-leucine zipper protein promotes efficient binding to a monomeric DNA binding site. Biochemistry 2004, 43, 15845–15851. [Google Scholar] [CrossRef]
- Aoyama, T.; Dong, C.-H.; Wu, Y.; Carabelli, M.; Sessa, G.; Ruberti, I.; Morelli, G.; Chua, N.-H. Ectopic expression of the Arabidopsis transcriptional activator Athb-1 alters leaf cell fate in tobacco. Plant Cell 1995, 7, 1773–1785. [Google Scholar] [PubMed] [Green Version]
- Mattsson, J.; Ckurshumova, W.; Berleth, T. Auxin signaling in Arabidopsis leaf vascular development. Plant Physiol. 2003, 131, 1327–1339. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Hong, Y.; Yin, M.; Li, C.; Zhang, K.; Grierson, D. A tomato HD-Zip homeobox protein, LeHB-1, plays an important role in floral organogenesis and ripening. Plant J. 2008, 55, 301–310. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhang, S.; Dong, R.; Wang, L.; Yao, J.; van Nocker, S.; Wang, X. The grapevine homeobox gene VvHB58 influences seed and fruit development through multiple hormonal signaling pathways. BMC Plant Biol. 2019, 19, 523. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Al-Baidhani, H.H.J.; Harris, J.; Riboni, M.; Li, Y.; Mazonka, I.; Bazanova, N.; Chirkova, L.; Sarfraz Hussain, S.; Hrmova, M. DREB/CBF expression in wheat and barley using the stress-inducible promoters of HD-Zip I genes: Impact on plant development, stress tolerance and yield. Plant Biotechnol. J. 2020, 18, 829–844. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.-J.; Li, P.-T.; Sun, L.-M.; Zhou, H.; Zeng, R.-F.; Ai, X.-Y.; Zhang, J.-Z.; Hu, C.-G. HD-ZIP I Transcription Factor (PtHB13) Negatively Regulates Citrus Flowering through Binding to FLOWERING LOCUS C Promoter. Plants 2020, 9, 114. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.H.; Ito, S.; Imaizumi, T. Flowering time regulation: Photoperiod-and temperature-sensing in leaves. Trends Plant Sci. 2013, 18, 575–583. [Google Scholar] [CrossRef] [Green Version]
- Gaudinier, A.; Blackman, B.K. Evolutionary processes from the perspective of flowering time diversity. New Phytol. 2020, 225, 1883–1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sessa, G.; Carabelli, M.; Sassi, M.; Ciolfi, A.; Possenti, M.; Mittempergher, F.; Becker, J.; Morelli, G.; Ruberti, I. A dynamic balance between gene activation and repression regulates the shade avoidance response in Arabidopsis. Genes Dev. 2005, 19, 2811–2815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morelli, G.; Ruberti, I. Light and shade in the photocontrol of Arabidopsis growth. Trends Plant Sci. 2002, 7, 399–404. [Google Scholar] [CrossRef]
- Sessa, G.; Carabelli, M.; Possenti, M.; Morelli, G.; Ruberti, I. Multiple pathways in the control of the shade avoidance response. Plants 2018, 7, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rueda, E.C.; Dezar, C.A.; Gonzalez, D.H.; Chan, R.L. Hahb-10, a sunflower homeobox-leucine zipper gene, is regulated by light quality and quantity, and promotes early flowering when expressed in Arabidopsis. Plant Cell Physiol. 2005, 46, 1954–1963. [Google Scholar] [CrossRef]
- Sawa, S.; Ohgishi, M.; Goda, H.; Higuchi, K.; Shimada, Y.; Yoshida, S.; Koshiba, T. The HAT2 gene, a member of the HD-Zip gene family, isolated as an auxin inducible gene by DNA microarray screening, affects auxin response in Arabidopsis. Plant J. 2002, 32, 1011–1022. [Google Scholar] [CrossRef] [Green Version]
- Delarue, M.; Prinsen, E.; Va, H.; Caboche, M.; Bellini, C. Sur2 mutations of Arabidopsis thaliana define a new locus involved in the control of auxin homeostasis. Plant J. 1998, 14, 603–611. [Google Scholar] [CrossRef]
- Gu, C.; Guo, Z.-H.; Cheng, H.-Y.; Zhou, Y.-H.; Qi, K.-J.; Wang, G.-M.; Zhang, S.-L. A HD-ZIP II HOMEBOX transcription factor, PpHB.G7, mediates ethylene biosynthesis during fruit ripening in peach. Plant Sci. 2019, 278, 12–19. [Google Scholar] [CrossRef]
- Chen, W.; Cheng, Z.; Liu, L.; Wang, M.; You, X.; Wang, J.; Zhang, F.; Zhou, C.; Zhang, Z.; Zhang, H. Small Grain and Dwarf 2, encoding an HD-Zip II family transcription factor, regulates plant development by modulating gibberellin biosynthesis in rice. Plant Sci. 2019, 288, 110208. [Google Scholar] [CrossRef]
- Prigge, M.J.; Otsuga, D.; Alonso, J.M.; Ecker, J.R.; Drews, G.N.; Clark, S.E. Class III homeodomain-leucine zipper gene family members have overlapping, antagonistic, and distinct roles in Arabidopsis development. Plant Cell 2005, 17, 61–76. [Google Scholar] [CrossRef] [Green Version]
- Emery, J.F.; Floyd, S.K.; Alvarez, J.; Eshed, Y.; Hawker, N.P.; Izhaki, A.; Baum, S.F.; Bowman, J.L. Radial patterning of Arabidopsis shoots by class III HD-ZIP and KANADI genes. Curr. Biol. 2003, 13, 1768–1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baima, S.; Possenti, M.; Matteucci, A.; Wisman, E.; Altamura, M.M.; Ruberti, I.; Morelli, G. The Arabidopsis ATHB-8 HD-zip protein acts as a differentiation-promoting transcription factor of the vascular meristems. Plant Physiol. 2001, 126, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Hawker, N.P.; Bowman, J.L. Roles for class III HD-Zip and KANADI genes in Arabidopsis root development. Plant Physiol. 2004, 135, 2261–2270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damodaran, S.; Dubois, A.; Xie, J.; Ma, Q.; Hindié, V.; Subramanian, S. GmZPR3d Interacts with GmHD-ZIP III Proteins and Regulates Soybean Root and Nodule Vascular Development. Int. J. Mol. Sci. 2019, 20, 827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Javelle, M.; Vernoud, V.; Depege-Fargeix, N.; Arnould, C.; Oursel, D.; Domergue, F.; Sarda, X.; Rogowsky, P.M. Overexpression of the epidermis-specific homeodomain-leucine zipper IV transcription factor Outer Cell Layer1 in maize identifies target genes involved in lipid metabolism and cuticle biosynthesis. Plant Physiol. 2010, 154, 273–286. [Google Scholar] [CrossRef] [Green Version]
- Chew, W.; Hrmova, M.; Lopato, S. Role of homeodomain leucine zipper (HD-Zip) IV transcription factors in plant development and plant protection from deleterious environmental factors. Int. J. Mol. Sci. 2013, 14, 8122–8147. [Google Scholar] [CrossRef] [Green Version]
- Kamata, N.; Okada, H.; Komeda, Y.; Takahashi, T. Mutations in epidermis-specific HD-ZIP IV genes affect floral organ identity in Arabidopsis thaliana. Plant J. 2013, 75, 430–440. [Google Scholar] [CrossRef]
- Ogawa, E.; Yamada, Y.; Sezaki, N.; Kosaka, S.; Kondo, H.; Kamata, N.; Abe, M.; Komeda, Y.; Takahashi, T. ATML1 and PDF2 play a redundant and essential role in Arabidopsis embryo development. Plant Cell Physiol. 2015, 56, 1183–1192. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Kwak, S.-H.; Zeng, Q.; Ellis, B.E.; Chen, X.-Y.; Schiefelbein, J.; Chen, J.-G. TRICHOMELESS1 regulates trichome patterning by suppressing GLABRA1 in Arabidopsis. Development 2007, 134, 3873–3882. [Google Scholar] [CrossRef] [Green Version]
- Landi, M.; Tattini, M.; Gould, K.S. Multiple functional roles of anthocyanins in plant-environment interactions. Environ. Exp. Bot. 2015, 119, 4–17. [Google Scholar] [CrossRef]
- Kubo, H.; Peeters, A.J.; Aarts, M.G.; Pereira, A.; Koornneef, M. ANTHOCYANINLESS2, a homeobox gene affecting anthocyanin distribution and root development in Arabidopsis. Plant Cell 1999, 11, 1217–1226. [Google Scholar] [CrossRef] [Green Version]
- Kubo, H.; Hayashi, K. Characterization of root cells of anl2 mutant in Arabidopsis thaliana. Plant Sci. 2011, 180, 679–685. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Chen, X.; Hong, Y.-Y.; Wang, Y.; Xu, P.; Ke, S.-D.; Liu, H.-Y.; Zhu, J.-K.; Oliver, D.J.; Xiang, C.-B. Activated expression of an Arabidopsis HD-START protein confers drought tolerance with improved root system and reduced stomatal density. Plant Cell 2008, 20, 1134–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Chen, X.; Wang, Z.; Wang, S.; Wang, Y.; Zhu, Q.; Li, S.; Xiang, C. Arabidopsis enhanced drought tolerance1/HOMEODOMAIN GLABROUS11 confers drought tolerance in transgenic rice without yield penalty. Plant Physiol. 2013, 162, 1378–1391. [Google Scholar] [CrossRef] [Green Version]
- Vernoud, V.; Laigle, G.; Rozier, F.; Meeley, R.B.; Perez, P.; Rogowsky, P.M. The HD-ZIP IV transcription factor OCL4 is necessary for trichome patterning and anther development in maize. Plant J. 2009, 59, 883–894. [Google Scholar] [CrossRef]
- Wei, J.; Choi, H.; Jin, P.; Wu, Y.; Yoon, J.; Lee, Y.-S.; Quan, T.; An, G. GL2-type homeobox gene Roc4 in rice promotes flowering time preferentially under long days by repressing Ghd7. Plant Sci. 2016, 252, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Raza, A. Eco-physiological and biochemical responses of rapeseed (Brassica napus L.) to abiotic stresses: Consequences and mitigation strategies. J. Plant Growth Regul. 2020, 40, 1368–1388. [Google Scholar] [CrossRef]
- Raza, A.; Ashraf, F.; Zou, X.; Zhang, X.; Tosif, H. Plant Adaptation and Tolerance to Environmental Stresses: Mechanisms and Perspectives. In Plant Ecophysiology and Adaptation under Climate Change: Mechanisms and Perspectives I; Springer: Berlin/Heidelberg, Germany, 2020; pp. 117–145. [Google Scholar]
- Zandalinas, S.I.; Mittler, R.; Balfagón, D.; Arbona, V.; Gómez-Cadenas, A. Plant adaptations to the combination of drought and high temperatures. Physiol. Plant 2018, 162, 2–12. [Google Scholar] [CrossRef] [Green Version]
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Perotti, M.F.; Ribone, P.A.; Chan, R.L. Plant transcription factors from the homeodomain-leucine zipper family I. Role in development and stress responses. IUBMB Life 2017, 69, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Gong, S.; Ding, Y.; Hu, S.; Ding, L.; Chen, Z.; Zhu, C. The role of HD-Zip class I transcription factors in plant response to abiotic stresses. Physiol. Plant 2019, 167, 516–525. [Google Scholar] [CrossRef]
- Zhou, W.; Malabanan, P.B.; Abrigo, E. OsHox4 regulates GA signaling by interacting with DELLA-like genes and GA oxidase genes in rice. Euphytica 2015, 201, 97–107. [Google Scholar] [CrossRef]
- Colombo, R.P.; Ibarra, J.G.; Bidondo, L.F.; Silvani, V.A.; Bompadre, M.J.; Pergola, M.; Lopez, N.I.; Godeas, A.M. Arbuscular mycorrhizal fungal association in genetically modified drought-tolerant corn. J. Environ. Qual. 2017, 46, 227–231. [Google Scholar] [CrossRef]
- Re, D.A.; Dezar, C.A.; Chan, R.L.; Baldwin, I.T.; Bonaventure, G. Nicotiana attenuata NaHD20 plays a role in leaf ABA accumulation during water stress, benzylacetone emission from flowers, and the timing of bolting and flower transitions. J. Exp. Bot. 2011, 62, 155–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharif, I.; Aleem, S.; Farooq, J.; Rizwan, M.; Younas, A.; Sarwar, G.; Chohan, S.M. Salinity stress in cotton: Effects, mechanism of tolerance and its management strategies. Physiol. Mol. Biol. Plants 2019, 25, 807–820. [Google Scholar] [CrossRef]
- Isayenkov, S.V.; Maathuis, F.J. Plant salinity stress: Many unanswered questions remain. Front. Plant Sci. 2019, 10, 80. [Google Scholar] [CrossRef] [Green Version]
- Negrão, S.; Schmöckel, S.; Tester, M. Evaluating physiological responses of plants to salinity stress. Ann. Bot. 2017, 119, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Haider, I.; Kohlen, W.; Jiang, L.; Bouwmeester, H.; Meijer, A.H.; Schluepmann, H.; Liu, C.-M.; Ouwerkerk, P.B. Function of the HD-Zip I gene Oshox22 in ABA-mediated drought and salt tolerances in rice. Plant Mol. Biol. 2012, 80, 571–585. [Google Scholar] [CrossRef]
- Deng, X.; Phillips, J.; Meijer, A.H.; Salamini, F.; Bartels, D. Characterization of five novel dehydration-responsive homeodomain leucine zipper genes from the resurrection plant Craterostigma plantagineum. Plant Mol. Biol. 2002, 49, 601–610. [Google Scholar] [CrossRef]
- Ni, Y.; Wang, X.; Li, D.; Wu, Y.; Xu, W.; Li, X. Novel cotton homeobox gene and its expression profiling in root development and in response to stresses and phytohormones. Acta Biochim. Biophys. Sin. 2008, 40, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Ma, Q.; Jin, X.; Peng, X.; Liu, J.; Deng, L.; Yan, H.; Sheng, L.; Jiang, H.; Cheng, B. A novel maize homeodomain–leucine zipper (HD-Zip) I gene, Zmhdz10, positively regulates drought and salt tolerance in both rice and arabidopsis. Plant Cell Physiol. 2014, 55, 1142–1156. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Bao, X.; Wang, S.; Liu, Y.; Tan, J.; Yang, M.; Zhang, M.; Dai, R.; Yu, X. A physic nut stress-responsive HD-Zip transcription factor, JcHDZ07, confers enhanced sensitivity to salinity stress in transgenic Arabidopsis. Front. Plant Sci. 2019, 10, 942. [Google Scholar] [CrossRef] [Green Version]
- Ye, Y.; Ding, Y.; Jiang, Q.; Wang, F.; Sun, J.; Zhu, C. The role of receptor-like protein kinases (RLKs) in abiotic stress response in plants. Plant Cell Rep. 2017, 36, 235–242. [Google Scholar] [CrossRef]
- Mehmood, S.S.; Lu, G.; Luo, D.; Hussain, M.A.; Raza, A.; Zafar, Z.; Zhang, X.; Cheng, Y.; Zou, X.; Lv, Y. Integrated Analysis of Transcriptomics and Proteomics provides insights into the molecular regulation of cold response in Brassica napus. Environ. Exp. Bot. 2021, 187, 104480. [Google Scholar] [CrossRef]
- He, H.; Lei, Y.; Yi, Z.; Raza, A.; Zeng, L.; Yan, L.; Xiaoyu, D.; Yong, C.; Xiling, Z. Study on the mechanism of exogenous serotonin improving cold tolerance of rapeseed (Brassica napus L.) seedlings. Plant Growth Regul. 2021, 94, 161–170. [Google Scholar] [CrossRef]
- Kovalchuk, N.; Chew, W.; Sornaraj, P.; Borisjuk, N.; Yang, N.; Singh, R.; Bazanova, N.; Shavrukov, Y.; Guendel, A.; Munz, E. The homeodomain transcription factor TaHDZipI-2 from wheat regulates frost tolerance, flowering time and spike development in transgenic barley. New Phytol. 2016, 211, 671–687. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Luang, S.; Harris, J.; Riboni, M.; Li, Y.; Bazanova, N.; Hrmova, M.; Haefele, S.; Kovalchuk, N.; Lopato, S. Overexpression of the class I homeodomain transcription factor TaHDZipI-5 increases drought and frost tolerance in transgenic wheat. Plant Biotechnol. J. 2018, 16, 1227–1240. [Google Scholar] [CrossRef] [PubMed]
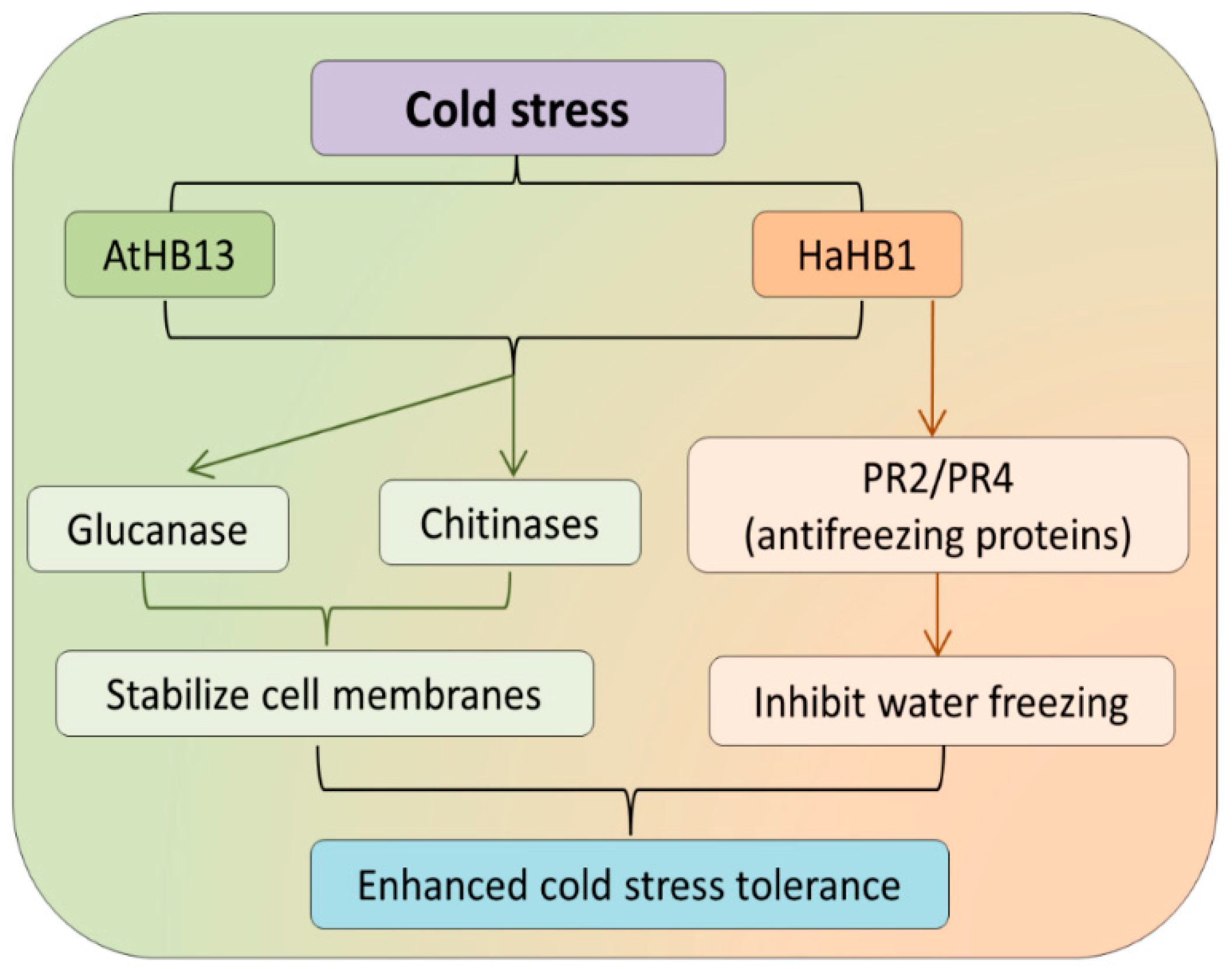
- Cabello, J.V.; Arce, A.L.; Chan, R.L. The homologous HD-Zip I transcription factors HaHB1 and AtHB13 confer cold tolerance via the induction of pathogenesis-related and glucanase proteins. Plant J. 2012, 69, 141–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raza, A.; Hussain, S.; Javed, R.; Hafeez, M.B.; Hasanuzzaman, M. Antioxidant Defense Systems and Remediation of Metal Toxicity in Plants. In Approaches to the Remediation of Inorganic Pollutants; Springer: Singapore, 2021; pp. 91–124. [Google Scholar]
- Raza, A.; Habib, M.; Kakavand, S.N.; Zahid, Z.; Zahra, N.; Sharif, R.; Hasanuzzaman, M. Phytoremediation of Cadmium: Physiological, Biochemical, and Molecular Mechanisms. Biology 2020, 9, 177. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Habib, M.; Charagh, S.; Kakavand, S.N. Genetic engineering of plants to tolerate toxic metals and metalloids. In Handbook of Bioremediation; Elsevier: Amsterdam, The Netherlands, 2021; pp. 411–436. [Google Scholar]
- Zhou, C.-P.; Li, C.-P.; Liang, W.-W.; Guo, P.; Yang, L.-T.; Chen, L.-S. Identification of manganese-toxicity-responsive genes in roots of two citrus species differing in manganese tolerance using cDNA-AFLP. Trees 2017, 31, 813–831. [Google Scholar] [CrossRef]
- Hu, S.; Ding, Y.; Zhu, C. Sensitivity and responses of chloroplasts to heat stress in plants. Front. Plant Sci. 2020, 11, 375. [Google Scholar] [CrossRef] [Green Version]
- Raza, A.; Tabassum, J.; Kudapa, H.; Varshney, R.K. Can omics deliver temperature resilient ready-to-grow crops? Crit. Rev. Biotechnol. 2021, 94, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Sharif, R.; Xie, C.; Wang, J.; Cao, Z.; Zhang, H.; Chen, P.; Yuhong, L. Genome wide identification, characterization and expression analysis of HD-ZIP gene family in Cucumis sativus L. under biotic and various abiotic stresses. Int. J. Biol. Macromol. 2020, 158, 502–520. [Google Scholar] [CrossRef]
- Wang, J.; Zhuang, L.; Zhang, J.; Yu, J.; Yang, Z.; Huang, B. Identification and characterization of novel homeodomain leucine zipper (HD-Zip) transcription factors associated with heat tolerance in perennial ryegrass. Environ. Exp. Bot. 2019, 160, 1–11. [Google Scholar] [CrossRef]
- Striker, G.G. Flooding stress on plants: Anatomical, morphological and physiological responses. Botany 2012, 1, 3–28. [Google Scholar]
- Pan, J.; Sharif, R.; Xu, X.; Chen, X. Waterlogging Response Mechanisms in Plants: Research Progress and Prospects. Front. Plant Sci. 2020, 11, 2319. [Google Scholar]
- Cabello, J.V.; Giacomelli, J.I.; Piattoni, C.V.; Iglesias, A.A.; Chan, R.L. The sunflower transcription factor HaHB11 improves yield, biomass and tolerance to flooding in transgenic Arabidopsis plants. J. Biotechnol. 2016, 222, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.-Y.; Li, J.; Wang, P.-Y.; Qin, F. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef] [PubMed]
- Gulzar, S.; Hassan, A.; Nawchoo, I.A. A Review of Nutrient Stress Modifications in Plants, Alleviation Strategies, and Monitoring through Remote Sensing. In Plant Micronutrients; Springer: Berlin/Heidelberg, Germany, 2020; pp. 331–343. [Google Scholar]
- Aznar, A.; Chen, N.W.; Thomine, S.; Dellagi, A. Immunity to plant pathogens and iron homeostasis. Plant Sci. 2015, 240, 90–97. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nozoye, T.; Nishizawa, N.K. Iron transport and its regulation in plants. Free Radic. Biol. Med. 2019, 133, 11–20. [Google Scholar] [CrossRef]
- Romani, F.; Capella, M.; Ribone, P.; Chan, R.L. The Arabidopsis transcription factor AtHB1 plays a role in iron homeostasis. In Proceedings of the 11th International Congress of Plant Molecular Biology, Iguazú Falls, Brazil, 25–30 October 2015. [Google Scholar]
- Wei, M.; Liu, A.; Zhang, Y.; Zhou, Y.; Li, D.; Dossa, K.; Zhou, R.; Zhang, X.; You, J. Genome-wide characterization and expression analysis of the HD-Zip gene family in response to drought and salinity stresses in sesame. BMC Genom. 2019, 20, 748. [Google Scholar] [CrossRef]
- Yue, H.; Shu, D.; Wang, M.; Xing, G.; Zhan, H.; Du, X.; Song, W.; Nie, X. Genome-Wide Identification and Expression Analysis of the HD-Zip Gene Family in Wheat (Triticum aestivum L.). Genes 2018, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.; Wu, W.; Abrams, S.R.; Cutler, A.J. The relationship of drought-related gene expression in Arabidopsis thaliana to hormonal and environmental factors. J. Exp. Bot. 2008, 59, 2991–3007. [Google Scholar] [CrossRef] [Green Version]
- Stape, J.L.; Binkley, D.; Ryan, M.G. Eucalyptus production and the supply, use and efficiency of use of water, light and nitrogen across a geographic gradient in Brazil. For. Ecol. Manag. 2004, 193, 17–31. [Google Scholar] [CrossRef]
- Almeida, A.C.; Soares, J.V.; Landsberg, J.J.; Rezende, G.D. Growth and water balance of Eucalyptus grandis hybrid plantations in Brazil during a rotation for pulp production. For. Ecol. Manag. 2007, 251, 10–21. [Google Scholar] [CrossRef]
- Sasaki, K.; Ida, Y.; Kitajima, S.; Kawazu, T.; Hibino, T.; Hanba, Y.T. Overexpressing the HD-Zip class II transcription factor EcHB1 from Eucalyptus camaldulensis increased the leaf photosynthesis and drought tolerance of Eucalyptus. Sci. Rep. 2019, 9, 14121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciarbelli, A.R.; Ciolfi, A.; Salvucci, S.; Ruzza, V.; Possenti, M.; Carabelli, M.; Fruscalzo, A.; Sessa, G.; Morelli, G.; Ruberti, I. The Arabidopsis homeodomain-leucine zipper II gene family: Diversity and redundancy. Plant Mol. Biol. 2008, 68, 465–478. [Google Scholar] [CrossRef]
- Carabelli, M.; Morelli, G.; Whitelam, G.; Ruberti, I. Twilight-zone and canopy shade induction of the Athb-2 homeobox gene in green plants. Proc. Natl. Acad. Sci. USA 1996, 93, 3530–3535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steindler, C.; Matteucci, A.; Sessa, G.; Weimar, T.; Ohgishi, M.; Aoyama, T.; Morelli, G.; Ruberti, I. Shade avoidance responses are mediated by the ATHB-2 HD-zip protein, a negative regulator of gene expression. Development 1999, 126, 4235–4245. [Google Scholar] [CrossRef]
- Lorrain, S.; Allen, T.; Duek, P.D.; Whitelam, G.C.; Fankhauser, C. Phytochrome-mediated inhibition of shade avoidance involves degradation of growth-promoting bHLH transcription factors. Plant J. 2008, 53, 312–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hornitschek, P.; Kohnen, M.V.; Lorrain, S.; Rougemont, J.; Ljung, K.; López-Vidriero, I.; Franco-Zorrilla, J.M.; Solano, R.; Trevisan, M.; Pradervand, S. Phytochrome interacting factors 4 and 5 control seedling growth in changing light conditions by directly controlling auxin signaling. Plant J. 2012, 71, 699–711. [Google Scholar] [CrossRef] [Green Version]
- Leivar, P.; Tepperman, J.M.; Cohn, M.M.; Monte, E.; Al-Sady, B.; Erickson, E.; Quail, P.H. Dynamic antagonism between phytochromes and PIF family basic helix-loop-helix factors induces selective reciprocal responses to light and shade in a rapidly responsive transcriptional network in Arabidopsis. Plant Cell 2012, 24, 1398–1419. [Google Scholar] [CrossRef] [Green Version]
- Shen, W.; Li, H.; Teng, R.; Wang, Y.; Wang, W.; Zhuang, J. Genomic and transcriptomic analyses of HD-Zip family transcription factors and their responses to abiotic stress in tea plant (Camellia sinensis). Genomics 2019, 111, 1142–1151. [Google Scholar] [CrossRef]
- Li, W.; Dong, J.; Cao, M.; Gao, X.; Wang, D.; Liu, B.; Chen, Q. Genome-wide identification and characterization of HD-ZIP genes in potato. Gene 2019, 697, 103–117. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, H.; Srivastava, A.K.; Pan, Y.; Bai, J.; Fang, J.; Shi, H.; Zhu, J.-K. Knockdown of Rice MicroRNA166 Confers Drought Resistance by Causing Leaf Rolling and Altering Stem Xylem Development. Plant Physiol. 2018, 176, 2082–2094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Wang, Y.; Teotia, S.; Wang, Z.; Shi, C.; Sun, H.; Gu, Y.; Zhang, Z.; Tang, G. The interaction between miR160 and miR165/166 in the control of leaf development and drought tolerance in Arabidopsis. Sci. Rep. 2019, 9, 2832. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Gao, Z.; Li, R.; Xu, Y.; Kong, Y.; Zhou, G.; Meng, C.; Hu, R. Genome-wide identification and expression profiling of HD-ZIP gene family in Medicago truncatula. Genomics 2020, 112, 3624–3635. [Google Scholar] [CrossRef] [PubMed]
- DalCorso, G.; Farinati, S.; Furini, A. Regulatory networks of cadmium stress in plants. Plant Signal. Behav. 2010, 5, 663–667. [Google Scholar] [CrossRef]
- Andresen, E.; Küpper, H. Cadmium toxicity in plants. In Cadmium: From Toxicity to Essentiality; Springer: Berlin/Heidelberg, Germany, 2013; pp. 395–413. [Google Scholar]
- Ismael, M.A.; Elyamine, A.M.; Moussa, M.G.; Cai, M.; Zhao, X.; Hu, C. Cadmium in plants: Uptake, toxicity, and its interactions with selenium fertilizers. Metallomics 2019, 11, 255–277. [Google Scholar] [CrossRef] [PubMed]
- Franco, D.M.; Silva, E.M.; Saldanha, L.L.; Adachi, S.A.; Schley, T.R.; Rodrigues, T.M.; Dokkedal, A.L.; Nogueira, F.T.S.; de Almeida, L.F.R. Flavonoids modify root growth and modulate expression of SHORT-ROOT and HD-ZIP III. J. Plant Physiol. 2015, 188, 89–95. [Google Scholar] [CrossRef]
- Singh, A.; Roy, S.; Singh, S.; Das, S.S.; Gautam, V.; Yadav, S.; Kumar, A.; Singh, A.; Samantha, S.; Sarkar, A.K. Phytohormonal crosstalk modulates the expression of miR166/165s, target Class III HD-ZIPs, and KANADI genes during root growth in Arabidopsis thaliana. Sci. Rep. 2017, 7, 3408. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Z.; Sun, B.; Xu, X.; Chen, H.; Zou, L.; Chen, G.; Cao, B.; Chen, C.; Lei, J. Overexpression of AtEDT1/HDG11 in Chinese kale (Brassica oleracea var. alboglabra) enhances drought and osmotic stress tolerance. Front. Plant Sci. 2016, 7, 1285. [Google Scholar] [CrossRef] [Green Version]
- Bang, S.W.; Lee, D.-K.; Jung, H.; Chung, P.J.; Kim, Y.S.; Choi, Y.D.; Suh, J.-W.; Kim, J.-K. Overexpression of OsTF1L, a rice HD-Zip transcription factor, promotes lignin biosynthesis and stomatal closure that improves drought tolerance. Plant Biotechnol. J. 2019, 17, 118–131. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.H.; Wu, S.J.; Peng, Y.S.; Liu, R.N.; Chen, X.; Zhao, P.; Xu, P.; Zhu, J.B.; Jiao, G.L.; Pei, Y. Arabidopsis EDT1/HDG11 improves drought and salt tolerance in cotton and poplar and increases cotton yield in the field. Plant Biotechnol. J. 2016, 14, 72–84. [Google Scholar] [CrossRef]
- Guo, X.Y.; Wang, Y.; Zhao, P.X.; Xu, P.; Yu, G.H.; Zhang, L.Y.; Xiong, Y.; Xiang, C.B. AtEDT1/HDG11 regulates stomatal density and water-use efficiency via ERECTA and E2Fa. New Phytol. 2019, 223, 1478–1488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ma, X.; Li, W.; Niu, D.; Wang, Z.; Yan, X.; Yang, X.; Yang, Y.; Cui, H. Genome-wide characterization of NtHD-ZIP IV: Different roles in abiotic stress response and glandular Trichome induction. BMC Plant Biol. 2019, 19, 444. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, S.; Khan, N.; Iqbal, M.Z.; Hussain, A.; Hassan, M. Salt tolerance of cotton (Gossypium hirsutum L.). Asian J. Plant Sci. 2002, 1, 715–719. [Google Scholar]
- Raza, A.; Charagh, S.; Zahid, Z.; Mubarik, M.S.; Javed, R.; Siddiqui, M.H.; Hasanuzzaman, M. Jasmonic acid: A key frontier in conferring abiotic stress tolerance in plants. Plant Cell Rep. 2021, 40, 1513–1541. [Google Scholar] [CrossRef]
- Mir, M.A.; John, R.; Alyemeni, M.N.; Alam, P.; Ahmad, P. Jasmonic acid ameliorates alkaline stress by improving growth performance, ascorbate glutathione cycle and glyoxylase system in maize seedlings. Sci. Rep. 2018, 8, 2831. [Google Scholar] [CrossRef]
- Cai, X.T.; Xu, P.; Wang, Y.; Xiang, C.B. Activated expression of AtEDT1/HDG11 promotes lateral root formation in Arabidopsis mutant edt1 by upregulating jasmonate biosynthesis. J. Integr. Plant Biol. 2015, 57, 1017–1030. [Google Scholar] [CrossRef]
- Lin, Z.; Li, Y.; Zhang, Z.; Liu, X.; Hsu, C.-C.; Du, Y.; Sang, T.; Zhu, C.; Wang, Y.; Satheesh, V. A RAF-SnRK2 kinase cascade mediates early osmotic stress signaling in higher plants. Nat. Commun. 2020, 11, 613. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.; Zhang, X.; Yang, Z.; Wang, X.; Yang, Z.; Zhang, C.; Wu, Z.; Kong, D.; Liu, Z.; Zhao, G. Genome-wide analysis of the HD-ZIP IV transcription factor family in Gossypium arboreum and GaHDG11 involved in osmotic tolerance in transgenic Arabidopsis. Mol. Genet. Genom. 2017, 292, 593–609. [Google Scholar] [CrossRef]
- Romani, F.; Ribone, P.A.; Capella, M.; Miguel, V.N.; Chan, R.L. A matter of quantity: Common features in the drought response of transgenic plants overexpressing HD-Zip I transcription factors. Plant Sci. 2016, 251, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimian-Motlagh, S.; Ribone, P.A.; Thirumalaikumar, V.P.; Allu, A.D.; Chan, R.L.; Mueller-Roeber, B.; Balazadeh, S. JUNGBRUNNEN1 confers drought tolerance downstream of the HD-Zip I transcription factor AtHB13. Front. Plant Sci. 2017, 8, 2118. [Google Scholar] [CrossRef] [Green Version]
- Cabello, J.V.; Giacomelli, J.I.; Gómez, M.C.; Chan, R.L. The sunflower transcription factor HaHB11 confers tolerance to water deficit and salinity to transgenic Arabidopsis and alfalfa plants. J. Biotechnol. 2017, 257, 35–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdés, A.E.; Övernäs, E.; Johansson, H.; Rada-Iglesias, A.; Engström, P. The homeodomain-leucine zipper (HD-Zip) class I transcription factors ATHB7 and ATHB12 modulate abscisic acid signalling by regulating protein phosphatase 2C and abscisic acid receptor gene activities. Plant Mol. Biol. 2012, 80, 405–418. [Google Scholar] [CrossRef]
- Steindler, C.; Carabelli, M.; Borello, U.; Morelli, G.; Ruberti, I. Phytochrome A, phytochrome B and other phytochrome (s) regulate ATHB-2 gene expression in etiolated and green Arabidopsis plants. Plant Cell Environ. 1997, 20, 759–763. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, S.; Xiao, Y.; Li, Z.; Wu, M.; Xie, X.; Li, H.; Mu, W.; Li, F.; Liu, P. Functional characterization of a HD-ZIP IV transcription factor NtHDG2 in regulating flavonols biosynthesis in Nicotiana tabacum. Plant Physiol. Biochem. 2020, 146, 259–268. [Google Scholar] [CrossRef]
- Ramegowda, V.; Senthil-Kumar, M. The interactive effects of simultaneous biotic and abiotic stresses on plants: Mechanistic understanding from drought and pathogen combination. J. Plant Physiol. 2015, 176, 47–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, M.P.; Rui, T.; Barbetti, M.J. Plant genotype and temperature impact simultaneous biotic and abiotic stress-related gene expression in Pythium-infected plants. Plant Pathol. 2020, 69, 655–668. [Google Scholar] [CrossRef]
- Gull, A.; Lone, A.A.; Wani, N.U.I. Biotic and abiotic stresses in plants. In Abiotic and Biotic Stress in Plants; IntechOpen: London, UK, 2019; pp. 1–19. [Google Scholar] [CrossRef] [Green Version]
- Sharif, R.; Mujtaba, M.; Ur Rahman, M.; Shalmani, A.; Ahmad, H.; Anwar, T.; Tianchan, D.; Wang, X. The Multifunctional Role of Chitosan in Horticultural Crops; A Review. Molecules 2018, 23, 872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharif, R.S.; Xie, C.; Zhang, H.; Arnao, M.B.; Ali, M.; Ali, Q.; Muhammad, I.; Shalmani, A.; Nawaz, M.A.; Chen, P.; et al. Melatonin and Its Effects on Plant Systems. Molecules 2018, 23, 2352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micali, C.; Göllner, K.; Humphry, M.; Consonni, C.; Panstruga, R. The powdery mildew disease of Arabidopsis: A paradigm for the interaction between plants and biotrophic fungi. Am. Soc. Plant Biol. 2008, 6, e0115. [Google Scholar] [CrossRef] [Green Version]
- Gao, D.; Appiano, M.; Huibers, R.P.; Chen, X.; Loonen, A.E.; Visser, R.G.; Wolters, A.-M.A.; Bai, Y. Activation tagging of ATHB13 in Arabidopsis thaliana confers broad-spectrum disease resistance. Plant Mol. Biol. 2014, 86, 641–653. [Google Scholar] [CrossRef]
- Manavella, P.A.; Dezar, C.A.; Bonaventure, G.; Baldwin, I.T.; Chan, R.L. HAHB4, a sunflower HD-Zip protein, integrates signals from the jasmonic acid and ethylene pathways during wounding and biotic stress responses. Plant J. 2008, 56, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Yu, J.; Wang, Y.; Tian, C. The C2H2 transcription factor VdMsn2 controls hyphal growth, microsclerotia formation, and virulence of Verticillium dahliae. Fungal Biol. 2017, 121, 1001–1010. [Google Scholar] [CrossRef]
- He, X.; Wang, T.; Zhu, W.; Wang, Y.; Zhu, L. GhHB12, a HD-ZIP I Transcription Factor, Negatively Regulates the Cotton Resistance to Verticillium dahliae. Int. J. Mol. Sci. 2018, 19, 3997. [Google Scholar] [CrossRef] [Green Version]
- Mulugeta, T.; Abreha, K.; Tekie, H.; Mulatu, B.; Yesuf, M.; Andreasson, E.; Liljeroth, E.; Alexandersson, E. Phosphite protects against potato and tomato late blight in tropical climates and has varying toxicity depending on the Phytophthora infestans isolate. Crop. Prot. 2019, 121, 139–146. [Google Scholar] [CrossRef]
- Ren, Y.; Armstrong, M.; Qi, Y.; McLellan, H.; Zhong, C.; Du, B.; Birch, P.R.; Tian, Z. Phytophthora infestans RXLR effectors target parallel steps in an immune signal transduction pathway. Plant Physiol. 2019, 180, 2227–2239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hausbeck, M.K.; Lamour, K.H. Phytophthora capsici on vegetable crops: Research progress and management challenges. Plant Dis. 2004, 88, 1292–1303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Liu, X.; Han, T.; Li, K.; Qu, Y.; Gao, Z. Differential Potential of Phytophthora capsici Resistance Mechanisms to the Fungicide Metalaxyl in Peppers. Microorganisms 2020, 8, 278. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, Y.L.; Suzuki, R.; Cabrera, J.; Nakagami, S.; Sagara, T.; Ejima, C.; Sano, R.; Aoki, Y.; Olmo, R.; Kurata, T. Root-knot and cyst nematodes activate procambium-associated genes in Arabidopsis roots. Front. Plant Sci. 2017, 8, 1195. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.-Y.; Iwasaki, M.; Machida, C.; Machida, Y.; Zhou, X.; Chua, N.-H. βC1, the pathogenicity factor of TYLCCNV, interacts with AS1 to alter leaf development and suppress selective jasmonic acid responses. Genes Dev. 2008, 22, 2564–2577. [Google Scholar] [CrossRef] [Green Version]
- Lewandowska, M.; Keyl, A.; Feussner, I. Wax biosynthesis in response to danger: Its regulation upon abiotic and biotic stress. New Phytol. 2020, 227, 698–713. [Google Scholar] [CrossRef] [Green Version]
- Ziv, C.; Zhao, Z.; Gao, Y.G.; Xia, Y. Multifunctional roles of plant cuticle during plant-pathogen interactions. Front. Plant Sci. 2018, 9, 1088. [Google Scholar] [CrossRef]
- Tafolla-Arellano, J.C.; Báez-Sañudo, R.; Tiznado-Hernández, M.E. The cuticle as a key factor in the quality of horticultural crops. Sci. Hortic. 2018, 232, 145–152. [Google Scholar] [CrossRef]
- Yan, T.; Li, L.; Xie, L.; Chen, M.; Shen, Q.; Pan, Q.; Fu, X.; Shi, P.; Tang, Y.; Huang, H. A novel HD-ZIP IV/MIXTA complex promotes glandular trichome initiation and cuticle development in Artemisia annua. New Phytol. 2018, 218, 567–578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karabourniotis, G.; Liakopoulos, G.; Nikolopoulos, D.; Bresta, P. Protective and defensive roles of non-glandular trichomes against multiple stresses: Structure–function coordination. J. For. Res. 2019, 31, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Sun, X.; Zhang, Q.; Song, P.; Hu, Q.; Zhang, X.; Li, X.; Hu, J.; Pan, J.; Sun, S. GLABROUS (CmGL) encodes a HD-ZIP IV transcription factor playing roles in multicellular trichome initiation in melon. Theor. Appl. Genet. 2018, 131, 569–579. [Google Scholar] [CrossRef] [PubMed]
- DeBono, A.; Yeats, T.H.; Rose, J.K.; Bird, D.; Jetter, R.; Kunst, L.; Samuels, L. Arabidopsis LTPG is a glycosylphosphatidylinositol-anchored lipid transfer protein required for export of lipids to the plant surface. Plant Cell 2009, 21, 1230–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, H.-W.; Tian, X.-H.; Ma, G.-H.; Li, Z.-X. Isolation and functional analysis of ZmLTP3, a homologue to Arabidopsis LTP3. Int. J. Mol. Sci. 2013, 14, 5025–5035. [Google Scholar] [CrossRef] [Green Version]
- Hairat, S.; Baranwal, V.K.; Khurana, P. Identification of Triticum aestivum nsLTPs and functional validation of two members in development and stress mitigation roles. Plant Physiol. Biochem. 2018, 130, 418–430. [Google Scholar] [CrossRef]
- Kovalchuk, N.; Li, M.; Wittek, F.; Reid, N.; Singh, R.; Shirley, N.; Ismagul, A.; Eliby, S.; Johnson, A.; Milligan, A.S. Defensin promoters as potential tools for engineering disease resistance in cereal grains. Plant Biotechnol. J. 2010, 8, 47–64. [Google Scholar] [CrossRef]
- Kovalchuk, N.; Wu, W.; Bazanova, N.; Reid, N.; Singh, R.; Shirley, N.; Eini, O.; Johnson, A.A.; Langridge, P.; Hrmova, M. Wheat wounding-responsive HD-Zip IV transcription factor GL7 is predominantly expressed in grain and activates genes encoding defensins. Plant Mol. Biol. 2019, 101, 41–61. [Google Scholar] [CrossRef]
- Zottich, U.; Da Cunha, M.; Carvalho, A.O.; Dias, G.B.; Silva, N.C.; Santos, I.S.; do Nacimento, V.V.; Miguel, E.C.; Machado, O.L.; Gomes, V.M. Purification, biochemical characterization and antifungal activity of a new lipid transfer protein (LTP) from Coffea canephora seeds with α-amylase inhibitor properties. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2011, 1810, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Boutrot, F.; Chantret, N.; Gautier, M.-F. Genome-wide analysis of the rice and Arabidopsis non-specific lipid transfer protein (nsLtp) gene families and identification of wheat nsLtp genes by EST data mining. BMC Genom. 2008, 9, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Stress Control | Plant Species | Gene | Functions | References |
|---|---|---|---|---|
| Subfamily I | ||||
| Drought | Arabidopsis thaliana | AtHB7 | Overexpression of AtHB7 regualte the expression of drought stress-specific genes. | [126] |
| Arabidopsis thaliana | AtHB13 | The AtHB13 work upstream of the JUB1 gene to confer drought stress. | [127] | |
| Arabidopsis thaliana | HaHB11 | The HaHB11 transgenic plants closed their stomata faster and lost less water than controls. | [128] | |
| Alfalfa | HaHB11 | Longer roots and rolled leaves in HaHB11 transgenic alfalfa plant. | [128] | |
| Arabidopsis thaliana | AtHB12 | Nullify the negative effects of ABA signaling genes (PYL5 and PYL8). | [129] | |
| Oryza sativa | OsHOX4 | The OsHOX4 modulate GA signaling by interacting with DELLA-like genes and GA oxidase genes. | [60] | |
| Oryza Sativa | OsHOX22 | Higher expression of OsHOX22 gene under drought stress. | [16] | |
| Nicotiana attenuate | NaHD20 | Augmented ABA accumualtion in leaf. | [62] | |
| Triticum aestivum | TaHDZ5-6A | TaHDZ5-6A transgenic plants displayed enhanced drought tolerance by lowering the water loss rates, higher survival rates, and higher proline contents. | [9] | |
| Salinity | Arabidopsis thaliana | HaHB11 | Higher expression of salt stress-related genes. | [128] |
| Alfalfa | HaHB11 | Strong root activities. | [128] | |
| Oryza sativa | OSHOX22 | Regualted ABA signaling. | [66] | |
| Physic nut | JcHDZ07 | Overexpression of JcHDZ07-induced sensitivity to salinity stress. | [70] | |
| Zea mays | ZmHDZ10 | Lower relative electrolyte leakage (REL), lowee MDA and increased proline content in overxpressed ZmHDZ10 transgenic plant. | [69] | |
| Heat stress | Soybean | HaHB4 | HaHB4 transgenic plant possesses larger xylem area, and increased water use efficiency under high temperature stress. | [11] |
| Perennial ryegrass | LpHOX21 | Higher expression of LpHOX21 gene was recorded in heat-tolerant cultivar. | [84] | |
| Heavy metal (manganese) | Citrus sinensis | TDF #170-1, 170-1k | Induced expression of these genes were observed under heavy metal stress. | [80] |
| Cold stress | Triticum aestivum | TaHDZipI-2 | Frost toelrance-related genes were upregualted in TaHDZip1-2 overxpressed plants. | [74] |
| Triticum aestivum | TaHDZipI-5 | Induction in lipid biosynthesis genes induced cold tolerance. | [75] | |
| Arabidopsis thaliana | AtHB13 | Higher antioxidant activities of AtHB13 transgenic plants. | [59] | |
| Arabidopsis thaliana | AtHB1/HaHB1 | Induction of pathogenesis-related and glucanase proteins. | [76] | |
| Flooding stress | Arabidopsis thaliana | HaHB11 | Modulation of genes genes involved in glycolisis and fermentative pathways. | [87] |
| Nutrient stress (iron deficiency) | Arabidopsis thaliana | AtHB1 | Overexpression of AtHB1 regualtes iron homeostasis. | [92] |
| Subfamily II | ||||
| Drought | Sesame | SiHDZ13, SiHDZ42 | Higher expression under drought stress. | [93] |
| Triticum aestivum | Tahdz4-A | Upregualted mRNA level under drought stress. | [94] | |
| Eucalyptus | EcHB1 | Increased the leaf photosynthesis. | [98] | |
| Arabidopsis thaliana | HAT2, HAT22 | High response to hormonal treatment. | [95] | |
| Salinity | Camellia sinensis | CsHDZ15, CsHDZ16 | Augmented expression under salinity stress. | [105] |
| Solanum tuberosum | StHOX17, StHOX20, StHOX27 | Higher expression under salinity stress. | [106] | |
| Capsicum annum | CaHB1 | Upregulation of multiple genes involved in plant osmotic stress resistance. | [10] | |
| Light stress | Arabidopsis thaliana | AtHB2/HAT4 | Stimualted expression of phytochrome genes in overexpressed AtHAT4 gene transgenic plant. | [100,130] |
| Subfamily III | ||||
| Drought | Oryza sativa | OsHB4 | LeafrRolling and altering stem xylem development. | [107] |
| Salinity | Triticum aestivum | Tahdz1, Tahdz23 | Induced mRNA level under salinity stress. | [94] |
| Medicago truncatula | MtHDZ5, MtHDZ13, MtHDZ22 | Higher expression under salinity stress. | [109] | |
| Cadmium stress | Oryza sativa | OsHB4 | Silencing of OsHB4 gene reduced Cd accumulation in the leaves and grains. | [12] |
| Subfamily IV | ||||
| Drought | Oryza sativa | OsHDG11 | Transgenic rice plants had higher levels of abscisic acid, proline, soluble sugar, and reactive oxygen species-scavenging enzyme activities under stress. | [51] |
| Chinese kale | AtEDT1/HDG11 | Induced stomatal closure. | [115] | |
| Gossypium herbaceum | HDG11 | Augmeneted proline content, soluble sugar content, and activities of reactive oxygen species-scavenging enzymes. | [117] | |
| Nicotiana tobaccum | NtHD-ZIP IV 4, NtHD-ZIP IV 10 | Higher expression under drought stress. | [119] | |
| Nicotiana tobaccum | NtHDG2 | Induced flavonoid biosynthesis. | [131] | |
| Oryza sativa | OsTFIL | Promotes lignin biosynthesis and stomatal closure. | [116] | |
| Salinity | Gossypium herbaceum | AtEDT1/HDG11 | Better proline content, soluble sugar content. | [117] |
| Arabidopsis thaliana | EDT1/HDG11 | Promotes lateral root formation in Arabidopsis mutant edt1 by upregulating jasmonate biosynthesis. | [123] | |
| Nicotiana tobaccum | NtHDG2 | Higher antioxidant activities. | [131] | |
| Osmotic | Arabidopsis thaliana | GaHDG11 | Upregualted expression level was observed under osmotic stress. | [125] |
| Subfamilies | Plant | Gene | Pathogen | Functions | Reference |
|---|---|---|---|---|---|
| Subfamily I | Arabidopsis thaliana | AtHB13 | Powdery mildew (Odium neolycopersici), downy mildew (Hyaloperonospora arabidopsidis) | Overexpression of AtHB13 stimualted the expression of various defense related genes. | [138] |
| Zea mays | HaHB4 | Spodoptera littoralis | Modulate signals from the jasmonic acid and ethylene pathways. | [139] | |
| Gossypium hirsutum | GhHB12 | Verticillium dahliae | Increased susceptibility of the cotton plant via suppression of the jasmonic acid (JA)-response genes GhJAZ2 and GhPR3. | [141] | |
| Subfamily II | Solanum tuberosum | StHOX28, StHOX30 | Phytophthora infestans | Induced expression pattern under Phytophora infestans. | [106] |
| Capsicum annuum | CaHB1 | Phytophthora capsici | Overexpression of CaHB1 in tomato resulted in a thicker cell wall. | [10] | |
| Subfamily III | Solanum tuberosum | StHOX7, StHOX16, StHOX26, StHOX38 | Phytophthora infestans | Induced expression pattern under Phytophora infestans. | [106] |
| Arabidopsis thaliana | AtHB8 | Meloidogyne incognita | The promoters of procambial marker gene ATHB8 were activated in M. incognita-induced galls. | [146] | |
| Arabidopsis thaliana | PHB, PHV | TYLCCNV | Suppress selective jasmonic acid responses. | [147] | |
| Subfamily IV | Zea mays | ZmOCL1 | Pseudomonas syringae | Overexpression of ZmOCL1 induced antifungal activity of a lipid transfer proteins. | [42,159,160] |
| Solanum tuberosum | StHOX21, StHOX42 | Phytophthora infestans | Induced expression pattern under Phytophora infestans. | [106] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharif, R.; Raza, A.; Chen, P.; Li, Y.; El-Ballat, E.M.; Rauf, A.; Hano, C.; El-Esawi, M.A. HD-ZIP Gene Family: Potential Roles in Improving Plant Growth and Regulating Stress-Responsive Mechanisms in Plants. Genes 2021, 12, 1256. https://doi.org/10.3390/genes12081256
Sharif R, Raza A, Chen P, Li Y, El-Ballat EM, Rauf A, Hano C, El-Esawi MA. HD-ZIP Gene Family: Potential Roles in Improving Plant Growth and Regulating Stress-Responsive Mechanisms in Plants. Genes. 2021; 12(8):1256. https://doi.org/10.3390/genes12081256
Chicago/Turabian StyleSharif, Rahat, Ali Raza, Peng Chen, Yuhong Li, Enas M. El-Ballat, Abdur Rauf, Christophe Hano, and Mohamed A. El-Esawi. 2021. "HD-ZIP Gene Family: Potential Roles in Improving Plant Growth and Regulating Stress-Responsive Mechanisms in Plants" Genes 12, no. 8: 1256. https://doi.org/10.3390/genes12081256