
Insights into the Evolution of Spermatogenesis-Related Ubiquitin–Proteasome System Genes in Abdominal Testicular Laurasiatherians

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Selected Species
2.2. Candidate Genes and Sequence Acquisition
2.3. Selective Pressure Analysis
2.4. Labeling Positive Selection Sites on the Three-Dimensional Structure of Proteins
2.5. Identification of Convergent Amino Acids among Abdominal Testicular Laurasiatherians
2.6. Functional Assays of RNF8
2.6.1. Plasmid Construction and Transient Transfection
2.6.2. Cell Culture, Protein Extraction and Western Blot Analysis
2.6.3. Statistical Analysis
3. Results
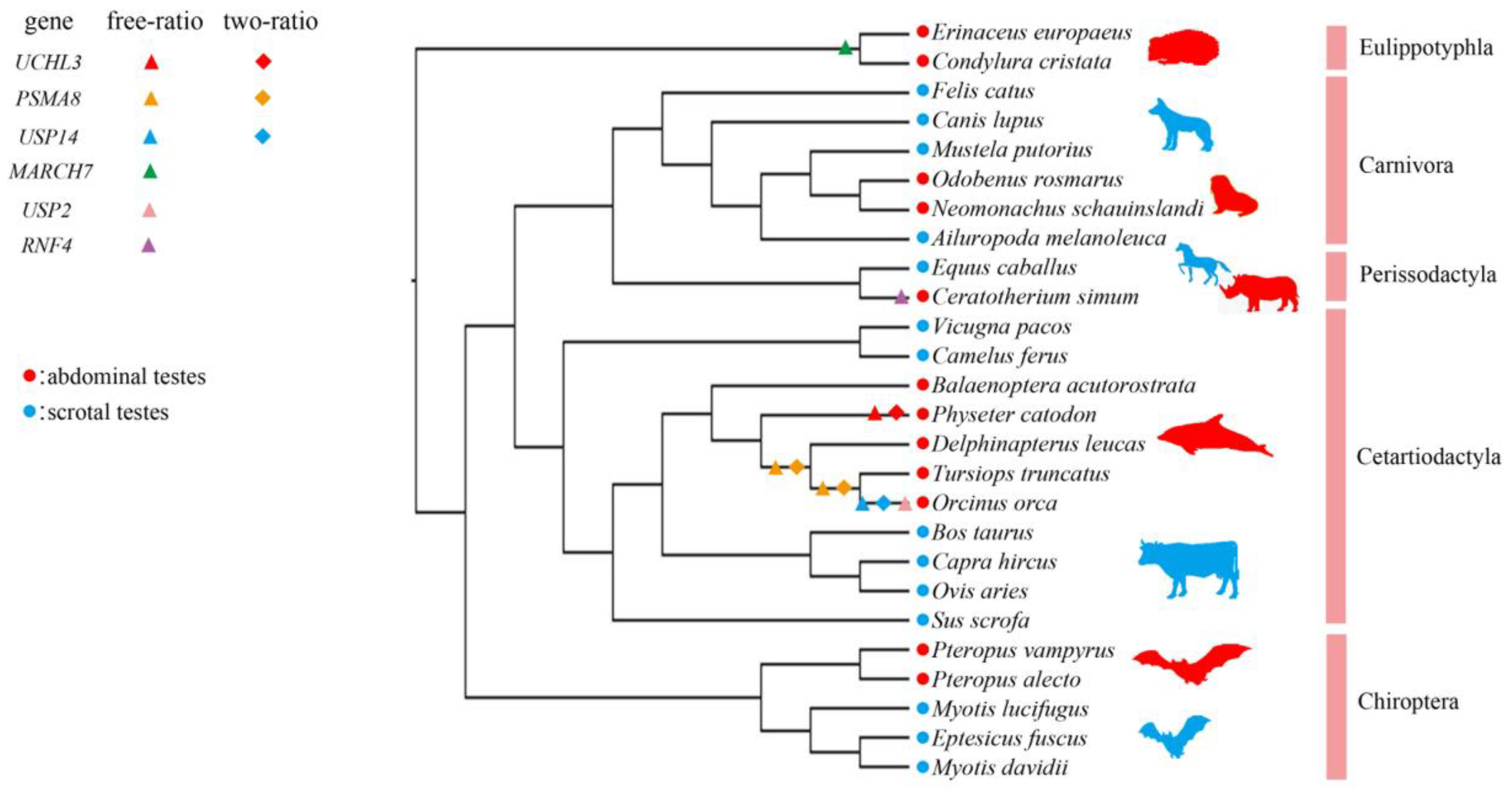
3.1. Positive Selection in Abdominal Testicular Mammals
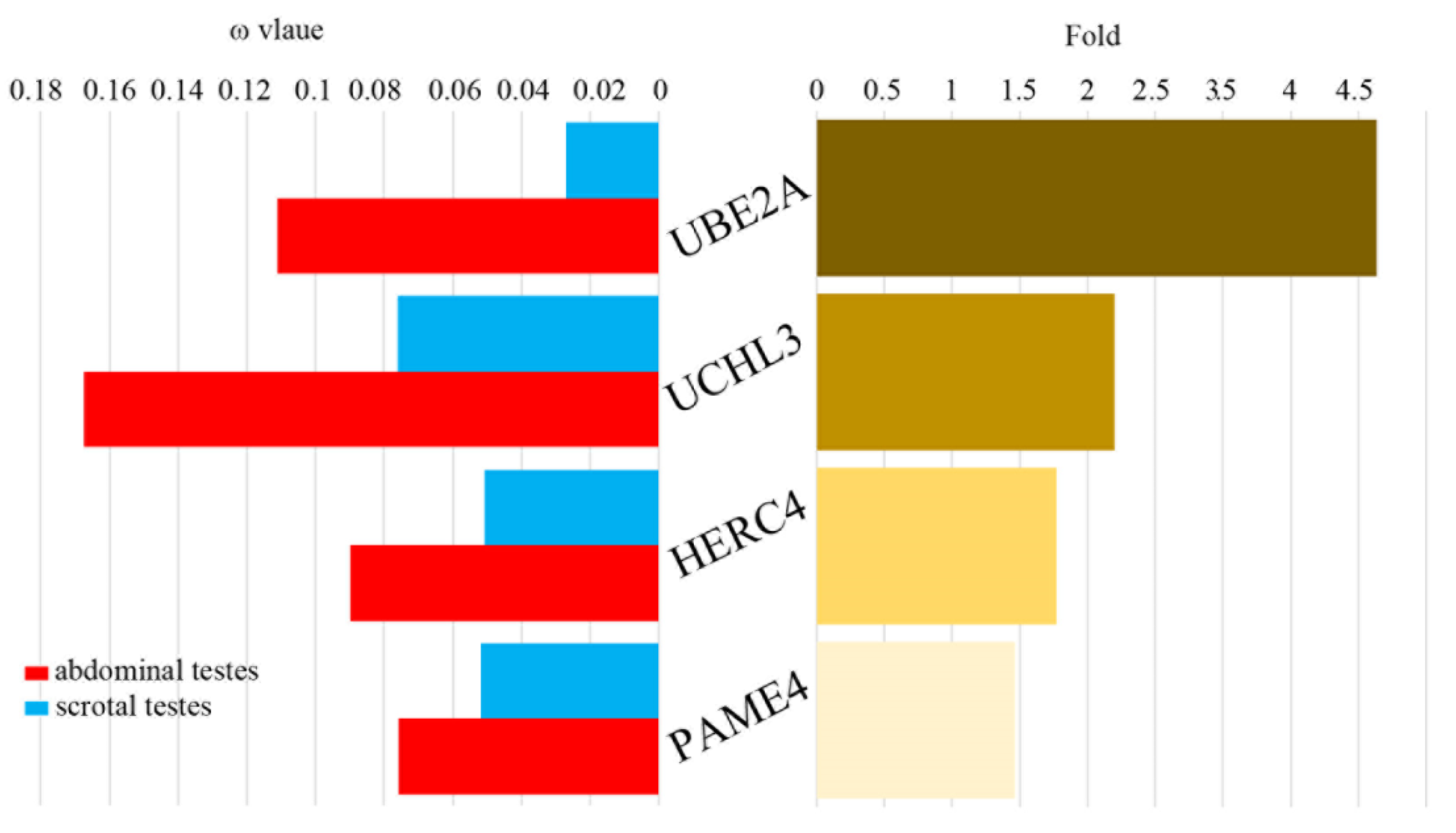
3.2. Rapid Evolution Rates in the Abdominal Testicular Lineages
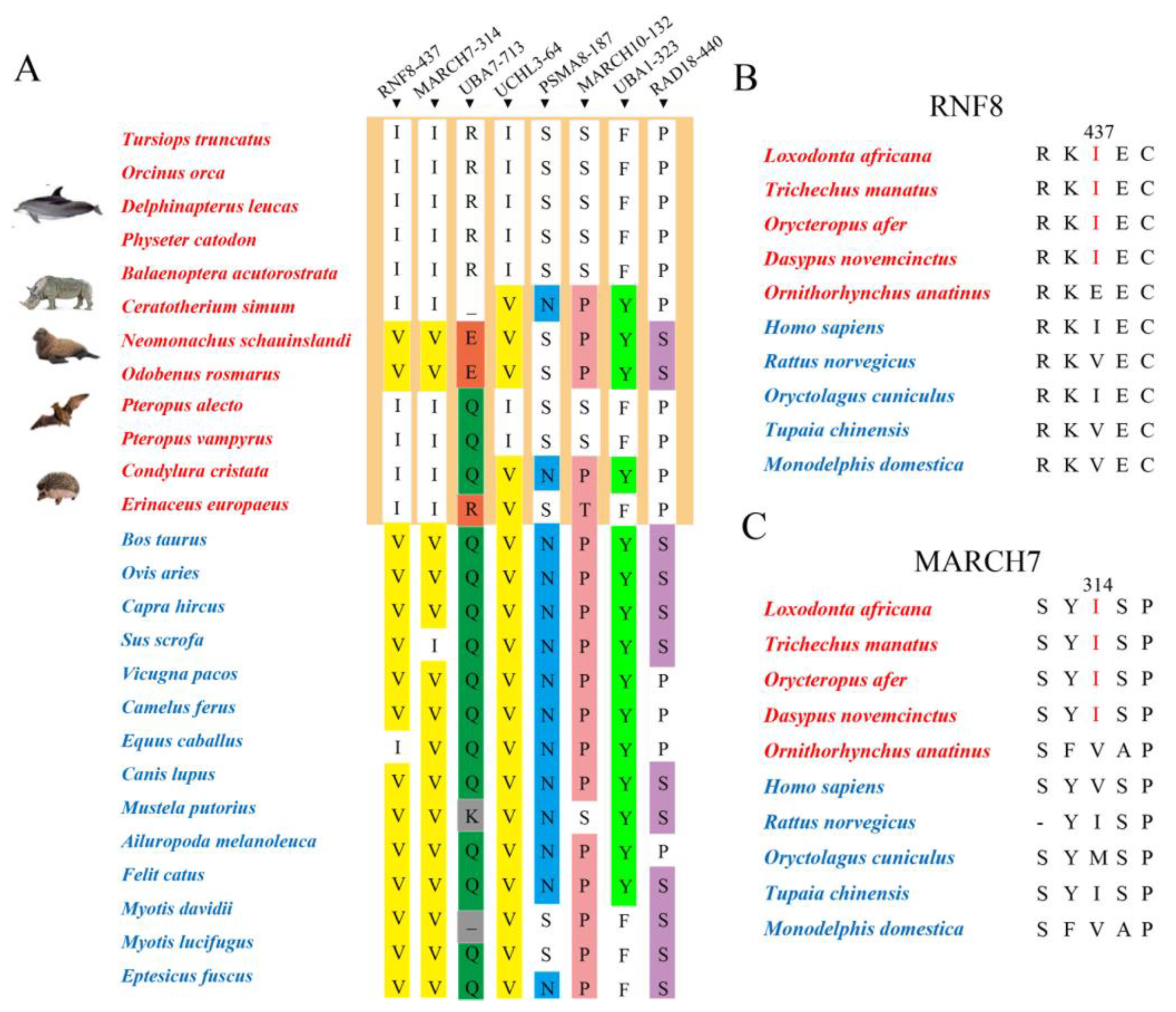
3.3. Potential Molecular Convergence between Abdominal Testicular Species
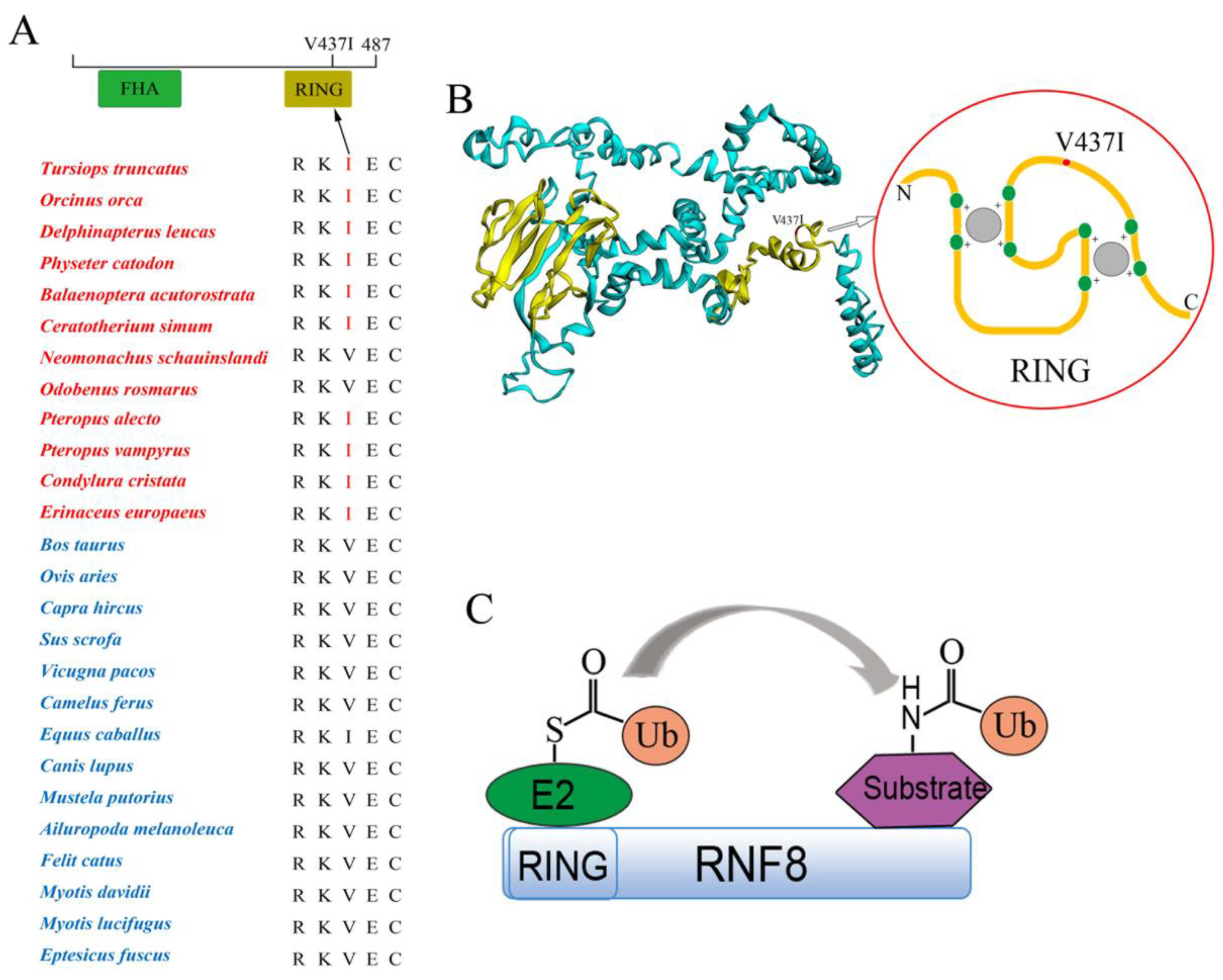
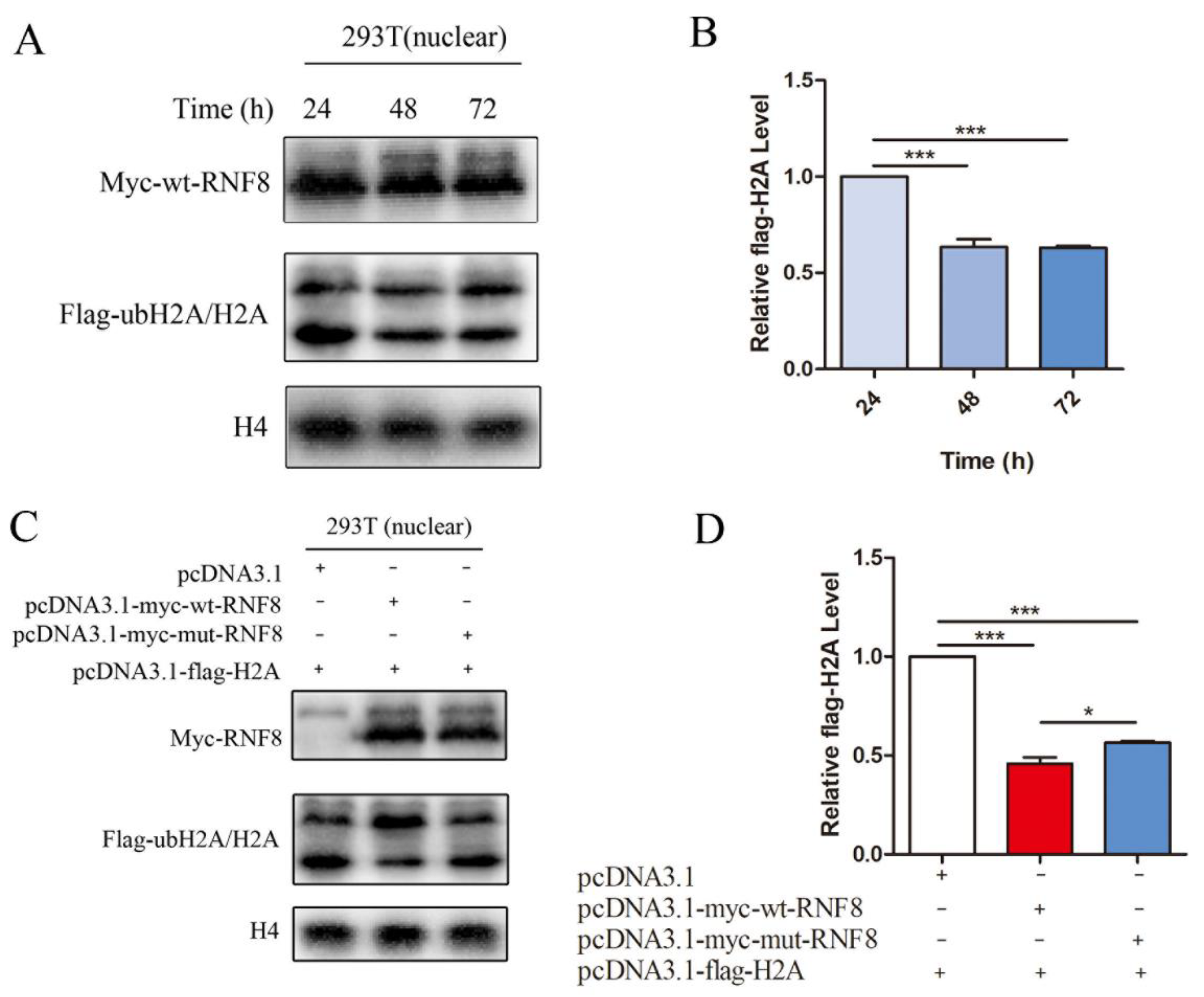
3.4. Functional Convergence of the RNF8 among Abdominal Testicular Species
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mamoulakis, C.; Antypas, S.; Sofras, F.; Takenaka, A.; Sofikitis, N. Testicular Descent. Hormones 2015, 14, 515–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariotti, A.; Di Carlo, L.; Orlando, G.; Corradini, M.L.; Di Donato, L.; Pompa, P.; Iezzi, R.; Cotroneo, A.R.; Romani, G.L.; Merla, A. Scrotal Thermoregulatory Model and Assessment of the Impairment of Scrotal Temperature Control in Varicocele. Ann. Biomed. Eng. 2011, 39, 664–673. [Google Scholar] [CrossRef] [PubMed]
- Thonneau, P.; Bujan, L.; Multigner, L.; Mieusset, R. Occupational heat exposure and male fertility: A review. Hum. Reprod. 1998, 13, 2122–2125. [Google Scholar] [CrossRef] [Green Version]
- Cobellis, G.; Noviello, C.; Nino, F.; Romano, M.; Mariscoli, F.; Martino, A.; Parmeggiani, P.; Papparella, A. Spermatogenesis and Cryptorchidism. Front. Endocrinol. 2014, 5, 63. [Google Scholar] [CrossRef] [PubMed]
- Durairajanayagam, D.; Agarwal, A.; Ong, C. Causes, effects and molecular mechanisms of testicular heat stress. Reprod. BioMedicine Online 2015, 30, 14–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleisner, K.; Ivell, R.; Flegr, J. The evolutionary history of testicular externalization and the origin of the scrotum. J. Biosci. 2010, 35, 27–37. [Google Scholar] [CrossRef]
- Gurney, J.K.; McGlynn, K.A.; Stanley, J.; Merriman, T.; Signal, V.; Shaw, C.; Edwards, R.; Richiardi, L.; Hutson, J.; Sarfati, D. Risk factors for cryptorchidism. Nat. Rev. Urol. 2017, 14, 534–548. [Google Scholar] [CrossRef]
- Sharma, V.; Lehmann, T.; Stuckas, H.; Funke, L.; Hiller, M. Loss of RXFP2 and INSL3 genes in Afrotheria shows that testicular descent is the ancestral condition in placental mammals. PLoS Biol. 2018, 16, e2005293. [Google Scholar] [CrossRef]
- Werdelin, L.; Nilsonne, Å. The Evolution of the Scrotum and Testicular Descent in Mammals: A Phylogenetic View. J. Theor. Biol. 1999, 196, 61–72. [Google Scholar] [CrossRef]
- Rommel, S.A.; Early, G.A.; Matassa, K.A.; Pabst, D.A.; Mclellan, W.A. Venous structures associated with thermoregulation of phocid seal reproductive organs. Anat. Rec. 1995, 243, 390–402. [Google Scholar] [CrossRef]
- Baarends, W.M.; van der Laan, R.; Grootegoed, J.A. Specific aspects of the ubiquitin system in spermatogenesis. J. Endocrinol. Investig. 2000, 23, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Sutovsky, P.; Aarabi, M.; Miranda-Vizuete, A.; Oko, R. Negative biomarker based male fertility evaluation: Sperm phenotypes associated with molecular-level anomalies. Asian J. Androl. 2015, 17, 554–560. [Google Scholar] [CrossRef] [PubMed]
- de Rooij, D.G. Proliferation and differentiation of spermatogonial stem cells. Reproduction 2001, 121, 347–354. [Google Scholar] [CrossRef]
- Meistrich, M.L.; Mohapatra, B.; Shirley, C.R.; Zhao, M. Roles of transition nuclear proteins in spermiogenesis. Chromosoma 2003, 111, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Baarends, W.M.; Hoogerbrugge, J.W.; Roest, H.P.; Ooms, M.; Vreeburg, J.; Hoeijmakers, J.H.J.; Grootegoed, J.A. Histone Ubiquitination and Chromatin Remodeling in Mouse Spermatogenesis. Dev. Biol. 1999, 207, 322–333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryu, K.Y.; Sinnar, S.A.; Reinholdt, L.G.; Vaccari, S.; Hall, S.; Garcia, M.A.; Zaitseva, T.S.; Bouley, D.M.; Boekelheide, K.; Handel, M.A. The mouse polyubiquitin gene Ubb is essential for meiotic progression. Mol. Cell. Biol. 2008, 28, 1136–1146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pickart, C.M. Mechanisms Underlying Ubiquitination. Annu. Rev. Biochem. 2001, 70, 503–533. [Google Scholar] [CrossRef]
- Nandi, D.; Tahiliani, P.; Kumar, A.; Chandu, D. The ubiquitin-proteasome system. J. Biosci. 2006, 31, 137–155. [Google Scholar] [CrossRef] [PubMed]
- Eletr, Z.M.; Wilkinson, K.D. Regulation of proteolysis by human deubiquitinating enzymes. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2014, 1843, 114–128. [Google Scholar] [CrossRef] [Green Version]
- Ying, D.; Liu, M.L.; Jia, M.C. Identification and characterization of a spermatogenesis-related gene Ube1 in rat testis. Sheng LI Xue Bao 2008, 60, 382–390. [Google Scholar]
- Mulugeta Achame, E.; Wassenaar, E.; Hoogerbrugge, J.W.; Sleddens-Linkels, E.; Ooms, M.; Sun, Z.-W.; van Ijcken, W.F.J.; Grootegoed, J.A.; Baarends, W.M. The ubiquitin-conjugating enzyme HR6B is required for maintenance of X chromosome silencing in mouse spermatocytes and spermatids. BMC Genom. 2010, 11, 367. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.-Y.; Wu, J.; Ye, L.; Gavrilina, G.B.; Saunders, T.L.; Yu, X. RNF8-dependent histone modifications regulate nucleosome removal during spermatogenesis. Dev. Cell 2010, 18, 371–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Song, Y.; Guo, Z.; Hu, M.; Liu, B.; Duan, H.; Wang, L.; Yuan, T.; Wang, D. Function of RAD6B and RNF8 in spermatogenesis. Cell Cycle 2018, 17, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ji, S.-Y.; Busayavalasa, K.; Shao, J.; Yu, C. Meiosis I progression in spermatogenesis requires a type of testis-specific 20S core proteasome. Nat. Commun. 2019, 10, 3387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, J.; Wang, Y.-L.; Setsuie, R.; Sekiguchi, S.; Sakurai, M.; Sato, Y.; Lee, W.-W.; Ishii, Y.; Kyuwa, S.; Noda, M.; et al. Developmental Regulation of Ubiquitin C-terminal Hydrolase Isozyme Expression During Spermatogenesis in Mice. Biol. Reprod. 2004, 71, 515–521. [Google Scholar] [CrossRef]
- Kwon, J. The New Function of Two Ubiquitin C-terminal Hydrolase Isozymes as Reciprocal Modulators of Germ Cell Apoptosis. Exp. Anim. 2007, 56, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z. PAML 4: Phylogenetic Analysis by Maximum Likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z. Likelihood ratio tests for detecting positive selection and application to primate lysozyme evolution. Mol. Biol. Evol. 1998, 15, 568–573. [Google Scholar] [CrossRef]
- Yang, Z.; Nielsen, R. Synonymous and nonsynonymous rate variation in nuclear genes of mammals. J. Mol. Evol. 1998, 46, 409–418. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Yang, Z.; Wong, W.S.W.; Nielsen, R. Bayes Empirical Bayes Inference of Amino Acid Sites Under Positive Selection. Mol. Biol. Evol. 2005, 22, 1107–1118. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.B. FasParser: A package for manipulating sequence data. Zool. Res. 2017, 38, 110–112. [Google Scholar] [PubMed] [Green Version]
- Elert, A.; Jahn, K.; Heidenreich, A.; Hofmann, R. The familial undescended testis. Klin. Pädiatrie 2002, 215, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Ayers, K.; Kumar, R.; Robevska, G.; Bruell, S.; Bell, K.; Malik, M.A.; Bathgate, R.A.; Sinclair, A. Original research: Familial bilateral cryptorchidism is caused by recessive variants in RXFP2. BMJ Open Access 2019, 56, 727–733. [Google Scholar]
- Abou El-Ella, S.S.; Tawfik, M.A.; Abd El-Aziz, T.F.; Shalaby, A.M.A.; Barseem, N.F. The G178A polymorphic variant of INSL3 may be linked to cryptorchidism among Egyptian pediatric cohort. Pediatric Surg. Int. 2020, 36, 1387–1393. [Google Scholar] [CrossRef]
- Panu, R.; Main, K.M.; Christine, W.V.; Hannu, K.; Riikka, A.; Terttu, V.; Skakkebæk, N.; Jorma, T.; Virtanen, H.E. Association of placenta organotin concentrations with congenital cryptorchidism and reproductive hormone levels in 280 newborn boys from Denmark and Finland. Hum. Reprod. 2013, 6, 1647–1660. [Google Scholar]
- García-Rodríguez, J.; García-Martín, M.; Nogueras-Ocaña, M.; de Dios Luna-del-Castillo, J.; Espigares García, M.; Olea, N.; Lardelli-Claret, P. Exposure to pesticides and cryptorchidism: Geographical evidence of a possible association. Environ. Health Perspect. 1996, 104, 1090–1095. [Google Scholar]
- Kolon Thomas, F.; Herndon, C.D.A.; Baker Linda, A.; Baskin Laurence, S.; Baxter Cheryl, G.; Cheng Earl, Y.; Diaz, M.; Lee Peter, A.; Seashore Carl, J.; Tasian Gregory, E.; et al. Evaluation and Treatment of Cryptorchidism: AUA Guideline. J. Urol. 2014, 192, 337–345. [Google Scholar] [CrossRef]
- Schrder, A. Ist die Hormontherapie bei Hodenhochstand noch zeitgem? Aktuelle Urol. 2020, 51, 183–185. [Google Scholar]
- Ding, X.; Wei, H.; Zhou, X.; Gu, L.; Yu, F.; Zheng, Y.; Ren, W.; Xu, S.; Yang, G. Molecular evolution of spermatogenesis-related genes in abdominal testicular mammals supports the cooling hypothesis. J. Genet. Genom. 2021. [Google Scholar] [CrossRef]
- Manku, G.; Wing, S.S.; Culty, M. Expression of the Ubiquitin Proteasome System in Neonatal Rat Gonocytes and Spermatogonia: Role in Gonocyte Differentiation1. Biol. Reprod. 2012, 87, 44. [Google Scholar] [CrossRef] [PubMed]
- Hua, Z.; Yu, P. Diversifying Evolution of the Ubiquitin-26S Proteasome System in Brassicaceae and Poaceae. Int. J. Mol. Sci. 2019, 20, 3226. [Google Scholar] [CrossRef] [Green Version]
- Thomas, J.H. Adaptive evolution in two large families of ubiquitin-ligase adapters in nematodes and plants. Genome Res. 2006, 16, 1017–1030. [Google Scholar] [CrossRef] [Green Version]
- Jain, N.; Cho, H.; O’Connell, M.; Lee, J.K. Rank-invariant resampling based estimation of false discovery rate for analysis of small sample microarray data. BMC Bioinform. 2005, 6, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Qian, X. Transferred subgroup false discovery rate for rare post-translational modifications detected by mass spectrometry. Mol. Cell. Proteom. 2014, 13, 1359–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryden, M.M. Testicular temperature in the Southern elephant seal, Mirounga leonina(Linn). Reproduction 1967, 13, 583–584. [Google Scholar] [CrossRef]
- Blix, A.S.; Fay, F.H.; Ronald, K. On testicular cooling in phocid seals. Polar Res. 1983, 1, 231–233. [Google Scholar] [CrossRef]
- Garolla, A.; Torino, M.; Sartini, B.; Cosci, I.; Foresta, C. Seminal and molecular evidence that sauna exposure affects human spermatogenesis. Hum. Reprod. 2013, 28, 877–885. [Google Scholar] [CrossRef] [Green Version]
- Gou, L.-T.; Kang, J.-Y.; Dai, P.; Wang, X.; Li, F.; Zhao, S.; Zhang, M.; Hua, M.-M.; Lu, Y.; Zhu, Y.; et al. Ubiquitination-Deficient Mutations in Human Piwi Cause Male Infertility by Impairing Histone-to-Protamine Exchange during Spermiogenesis. Cell 2017, 169, 1090–1104.e13. [Google Scholar] [CrossRef] [Green Version]
- Ma, T.; Keller, J.A.; Yu, X. RNF8-dependent histone ubiquitination during DNA damage response and spermatogenesis. Acta Biochim. Biophys. Sin. (Shanghai) 2011, 43, 339–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazari, M.; Babakhanzadeh, E.; Mohsen Aghaei Zarch, S.; Talebi, M.; Narimani, N.; Dargahi, M.; Sabbaghian, M.; Ghasemi, N. Upregulation of the RNF8 gene can predict the presence of sperm in azoospermic individuals. Clin. Exp. Reprod. Med. 2020, 47, 61–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plans, V.; Scheper, J.; Soler, M.; Loukili, N.; Okano, Y.; Thomson, T.M. The RING finger protein RNF8 recruits UBC13 for lysine 63-based self polyubiquitylation. J. Cell. Biochem. 2010, 97, 572–582. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Adachi, S.; Iwakami, R.; Yasuda, H.; Muto, Y.; Seki, N.; Okano, Y. N-terminally extended human ubiquitin-conjugating enzymes(E2s) mediate the ubiquitination of RING-finger proteins, ARA54 and RNF8. FEBS J. 2010, 268, 2725–2732. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, X.; Cao, L.; Zheng, Y.; Zhou, X.; He, X.; Xu, S.; Ren, W. Insights into the Evolution of Spermatogenesis-Related Ubiquitin–Proteasome System Genes in Abdominal Testicular Laurasiatherians. Genes 2021, 12, 1780. https://doi.org/10.3390/genes12111780
Ding X, Cao L, Zheng Y, Zhou X, He X, Xu S, Ren W. Insights into the Evolution of Spermatogenesis-Related Ubiquitin–Proteasome System Genes in Abdominal Testicular Laurasiatherians. Genes. 2021; 12(11):1780. https://doi.org/10.3390/genes12111780
Chicago/Turabian StyleDing, Xiaoyue, Li Cao, Yu Zheng, Xu Zhou, Xiaofang He, Shixia Xu, and Wenhua Ren. 2021. "Insights into the Evolution of Spermatogenesis-Related Ubiquitin–Proteasome System Genes in Abdominal Testicular Laurasiatherians" Genes 12, no. 11: 1780. https://doi.org/10.3390/genes12111780