Revisiting the Karyotype Evolution of Neotropical Boid Snakes: A Puzzle Mediated by Chromosomal Fissions

 , , ,
, , ,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Sampling and Mitotic Chromosome Preparation
2.2. Microdissection and Chromosome Painting Probe Preparation
2.3. Microscopy and Image Analyses
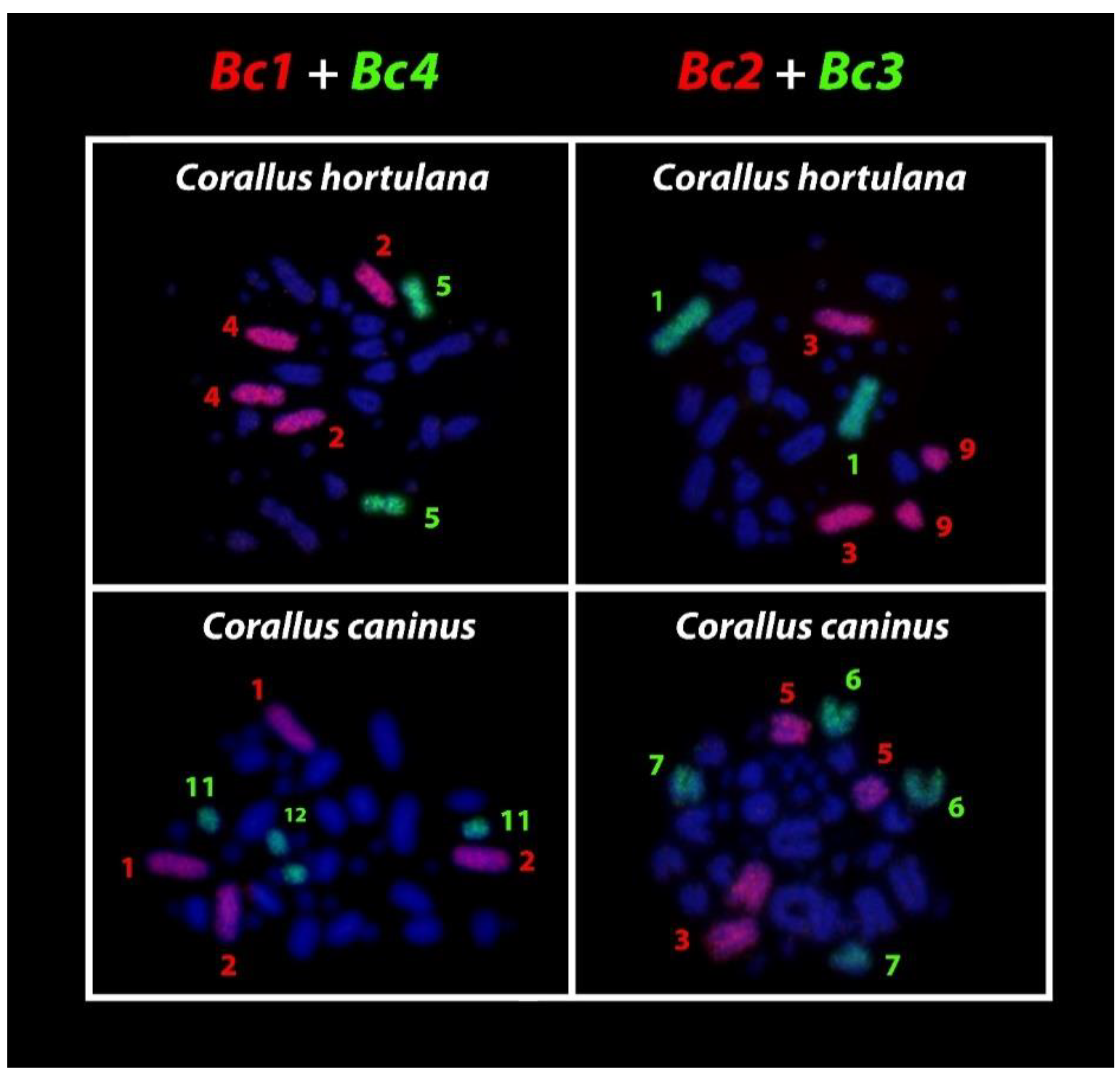
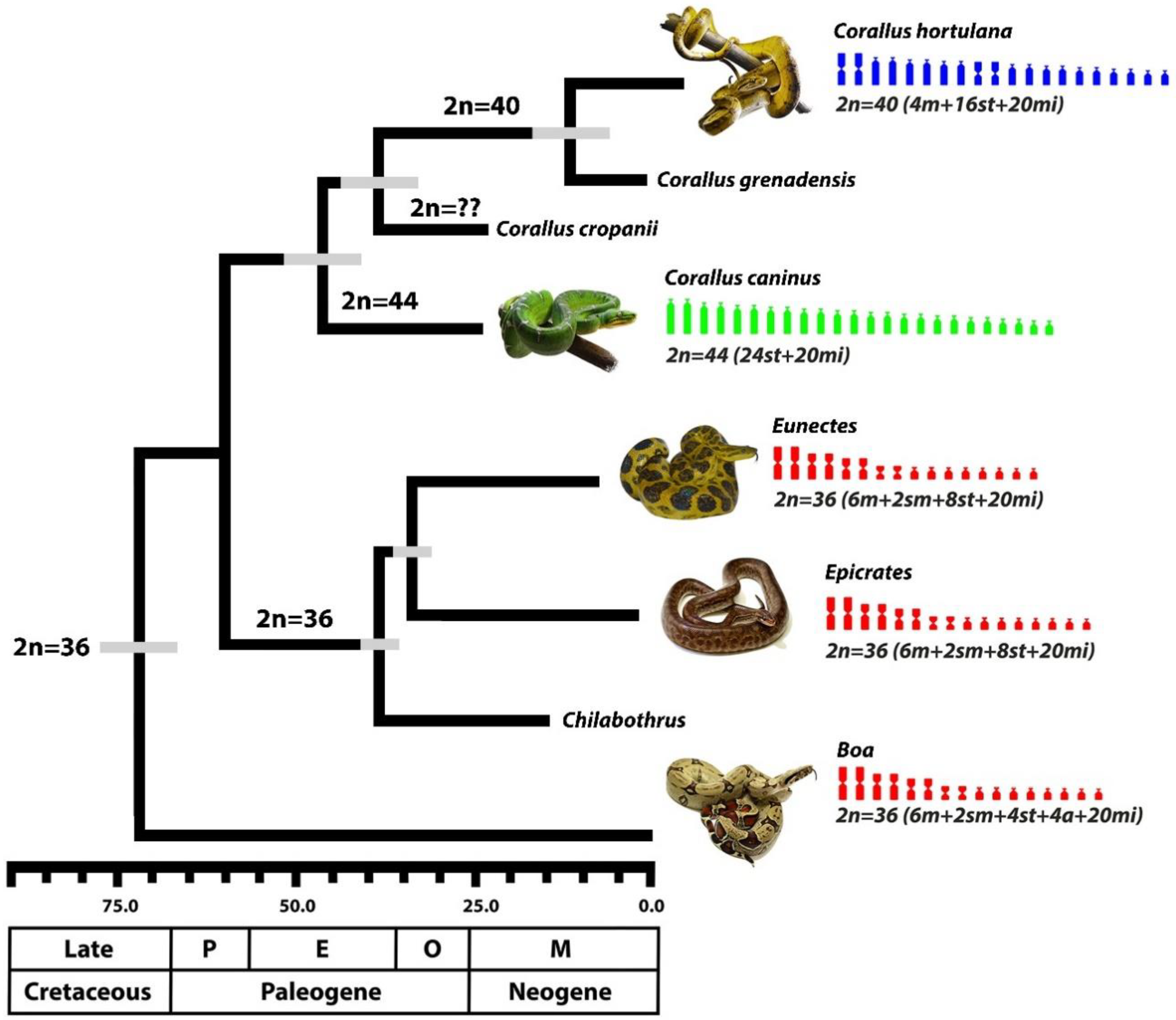
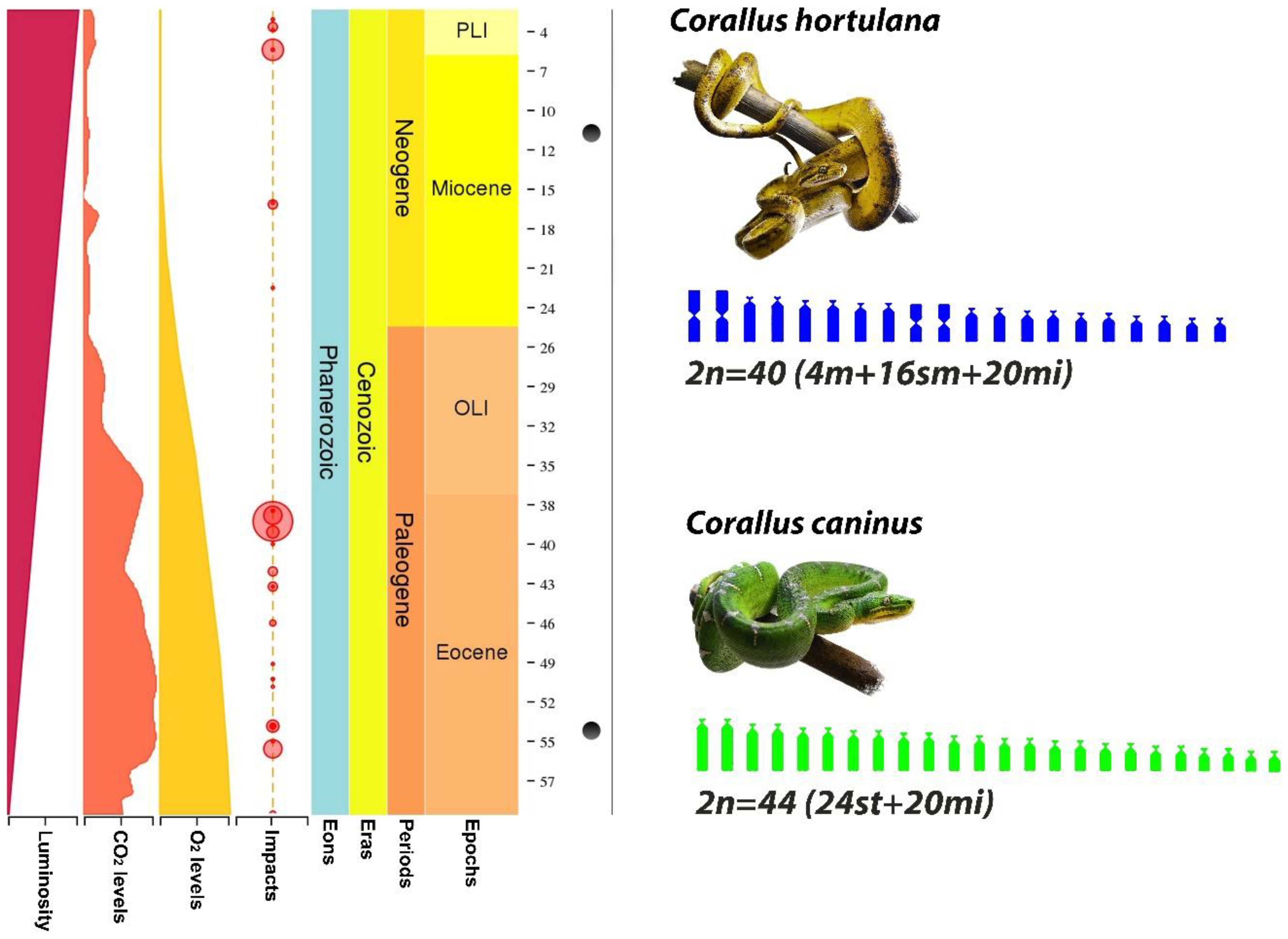
3. Results
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol. 2013, 13, 1–54. [Google Scholar] [CrossRef] [Green Version]
- Hsiang, A.Y.; Field, D.J.; Webster, T.H.; Behlke, A.D.B.; Davis, M.B.; Racicot, R.A.; Gauthier, J. The origin of snakes: Revealing the ecology, behavior, and evolutionary history of early snakes using genomics, phenomics, and the fossil record. BMC Evol. Biol. 2015, 15, 8. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Wiens, J.J. Combining phylogenomic and supermatrix approaches, and a time-calibrated phylogeny for squamate reptiles (lizards and snakes) based on 52 genes and 4162 species. Mol. Phylogenet. Evol. 2016, 94, 537–547. [Google Scholar] [CrossRef]
- The Reptile Database. Available online: http://www.reptile-database.org (accessed on 22 May 2020).
- Pyron, R.A.; Reynolds, R.G.; Burbrink, F.T. A taxonomic revision of boas (Serpentes: Boidae). Zootaxa 2014, 3846, 249–260. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, R.G.; Niemiller, M.L.; Revell, L.J. Toward a Tree-of-Life for the boas and pythons: Multilocus species-level phylogeny with unprecedented taxon sampling. Mol. Phylogenet. Evol. 2014, 71, 201–213. [Google Scholar] [CrossRef] [PubMed]
- Underwood, G. A Contribution to the Classification of Snakes, 1st ed.; Trustees of the British Museum (Natural History): London, UK, 1967. [Google Scholar]
- Kluge, A.G. Boine snake phylogeny and research cycles. Misc. Publ. Mus. Zool. Univ. Mich. 1991, 178, 1–58. [Google Scholar]
- Reed, R.N.; Rodda, G.H. Giant Constrictors: Biological and Management Profiles and an Establishment Risk Assessment for Nine Large Species of Pythons, Anacondas, and the Boa constrictor, Open-File Report 2009-1202; U.S. Geological Survey: Fort Collins, CO, USA, 2009; 302p. [Google Scholar]
- Rage, J.-C.; Burg, J.P.; Matte, P.; Leyreloup, A.; Marchand, J. Gondwana, Tethys, and terrestrial vertebrates during the Mesozoic and Cainozoic. Geol. Soc. Lond. Spec. Publ. 1988, 37, 255–273. [Google Scholar] [CrossRef]
- Rage, J.C. Fossil snakes from the Paleocene of São José de Itaboraí, Brazil, Part II. Boidae. Palaeovertebrata 2001, 30, 111–150. [Google Scholar]
- Noonan, B.P.; Chippindale, P.T. Dispersal and vicariance: The complex evolutionary history of boid snakes. Mol. Phylogenet. Evol. 2006, 40, 347–358. [Google Scholar] [CrossRef]
- Noonan, B.P.; Chippindale, P.T. Vicariant origin of Malagasy reptiles supports late cretaceous Antarctic land bridge. Am. Nat. 2006, 168, 730–741. [Google Scholar] [CrossRef]
- Reynolds, R.G.; Henderson, R.W. Boas of the World (Superfamily Booidae): A Checklist With Systematic, Taxonomic, and Conservation Assessments. Bull. Mus. Comp. Zool. 2018, 162, 1–58. [Google Scholar] [CrossRef] [Green Version]
- Vidal, N.; Dewynter, M.; Gower, D.J. Dissecting the major American snake radiation: A molecular phylogeny of the Dipsadidae Bonaparte (Serpentes, Caenophidia). Comptes Rendus Biol. 2010, 333, 48–55. [Google Scholar] [CrossRef]
- Colston, T.J.; Grazziotin, F.G.; Shepard, D.B.; Vitt, L.J.; Colli, G.R.; Henderson, R.W.; Blair Hedges, S.; Bonatto, S.; Zaher, H.; Noonan, B.P.; et al. Molecular systematics and historical biogeography of tree boas (Corallus spp.). Mol. Phylogenet. Evol. 2013, 66, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Henderson, R.W.; Micucci, T.W.P.; Puorto, G.; Bourgeois, R.W. Ecological correlates and patterns in the distribution of Neotropical boines (Serpentes: Boidae): A preliminary assessment. Herpetol. Nat. His. 1995, 1, 15–27. [Google Scholar]
- Viana, P.F. Corallus hortulanus (Amazon Tree Boa). Neonate size and timing of reproduction in captivity. Herpetol. Rev. 2017, 48, 87. [Google Scholar]
- Vidal, N.; Henderson, R.W.; Delmas, A.S.; Hedges, S.B. A phylogenetic study of the emerald tree boa (Corallus caninus). J. Herpetol. 2005, 39, 500–503. [Google Scholar] [CrossRef]
- Hoorn, C.; Wesselingh, F.P. Introduction: Amazonia, landscape and species evolution. In Amazonia: Landscape and Species Evolution: A Look into the Past, 1st ed.; Hoorn, C., Wesselingh, F.P., Eds.; Wiley: New York, NY, USA, 2010; Volume 1, pp. 1–6. [Google Scholar]
- Hoorn, C.; Wesselingh, F.P.; Ter Steege, H.; Bermúdez, M.; Mora, A.; Sevink, J.; Sanmartin, I.; Sanchez-Meseguer, A.; Anderson, C.L.; Figueiredo, J.P.; et al. Amazonia Through Time: Andean Uplift, Climate Change, Landscape Evolution, and Biodiversity. Science 2010, 330, 927–9311. [Google Scholar] [CrossRef] [Green Version]
- Henderson, R.W.; Powell, R. Natural History of West Indian Reptiles and Amphibians, 1st ed.; University Press of Florida: Gainesville, FL, USA, 2009. [Google Scholar]
- Card, D.C.; Schield, D.R.; Adams, R.H.; Corbin, A.B.; Perry, B.W.; Andrew, A.L.; Pasquesi, G.I.M.; Smith, E.N.; Jezkova, T.; Boback, S.M.; et al. Phylogeographic and population genetic analyses reveal multiple species of Boa and independent origins of insular dwarfism. Mol. Phylogenet. Evol. 2016, 102, 104–116. [Google Scholar] [CrossRef] [Green Version]
- Viana, P.F.; Ribeiro, L.B.; Souza, G.M.; Chalkidis, H.D.M.; Gross, M.C.; Feldberg, E. Is the Karyotype of Neotropical Boid Snakes Really Conserved? Cytotaxonomy, Chromosomal Rearrangements and Karyotype Organization in the Boidae Family. PLoS ONE 2016, 11, e0160274. [Google Scholar] [CrossRef]
- Beçak, W.; Beçak, M.L. Cytotaxonomy and chromosomal evolution in Serpentes. Cytogenet. Genome Res. 1969, 8, 247–262. [Google Scholar] [CrossRef]
- Gorman, G.C.; Gress, F. Chromosome cytology of four boid snakes and a varanid lizard, with comments on the cytosystematics of primitive snakes. Herpetologica 1970, 26, 308–317. [Google Scholar]
- Singh, L. Evolution of karyotypes in snakes. Chromosoma 1972, 38, 185–236. [Google Scholar] [CrossRef] [PubMed]
- Viana, P.F.; Ezaz, T.; Cioffi, M.B.; Almeida, B.J.; Feldberg, E. Evolutionary Insights of the ZW Sex Chromosomes in Snakes: A New Chapter Added by the Amazonian Puffing Snakes of the Genus Spilotes. Genes 2019, 10, 288. [Google Scholar] [CrossRef] [Green Version]
- Gamble, T.; Castoe, T.A.; Nielsen, S.V.; Banks, J.L.; Card, D.C.; Schield, D.R.; Schuett, G.W.; Booth, W. The Discovery of XY Sex Chromosomes in a Boa and Python. Curr. Biol. 2017, 27, 2148–2153.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augstenová, B.; Mazzoleni, S.; Kostmann, A.; Altmanová, M.; Frynta, D.; Kratochvíl, L.; Rovatsos, M. Cytogenetic Analysis Did Not Reveal Differentiated Sex Chromosomes in Ten Species of Boas and Pythons (Reptilia: Serpentes). Genes 2019, 10, 934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trifonov, V.A.; Paoletti, A.; Barucchi, V.C.; Kalinina, T.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Giovannotti, M. Comparative Chromosome Painting and NOR Distribution Suggest a Complex Hybrid Origin of Triploid Lepidodactylus lugubris (Gekkonidae). PLoS ONE 2015, 10, e0132380. [Google Scholar] [CrossRef]
- Cioffi, M.B.; Ráb, P.; Ezaz, T.; Bertollo, L.; Lavoué, S.; Oliveira, E.A.; Sember, A.; Molina, W.F.; Souza, F.; Majtánová, Z.; et al. Deciphering the evolutionary history of arowana fishes (Teleostei, Osteoglossiformes, Osteoglossidae): Insight from comparative cytogenomics. Int. J. Mol. Sci. 2019, 20, 4296. [Google Scholar] [CrossRef] [Green Version]
- Lisachov, A.P.; Giovannotti, M.; Pereira, J.C.; Andreyushkova, D.A.; Romanenko, S.A.; Ferguson-Smith, M.A.; Borodin, P.M.; Trifonov, V.A. Chromosome Painting Does Not Support a Sex Chromosome Turnover in Lacerta agilis Linnaeus, 1758. Cytogenet. Genome Res. 2020, 160, 134–140. [Google Scholar] [CrossRef]
- Viana, P.F.; Ribeiro, L.B.; Lima, T.; De Carvalho, V.T.; Vogt, R.C.; Gross, M.C.; Feldberg, E. An optimized protocol for obtaining mitotic chromosomes from cultured reptilian lymphocytes. Nucleus 2016, 59, 191–195. [Google Scholar] [CrossRef]
- Yang, F.; Trifonov, V.; Ng, B.L.; Kosyakova, N.; Carter, N.P. Generation of paint probes by flow-sorted and microdissected chromosomes. In Fluorescence In Situ Hybridization (FISH)—Application Guide, 1st ed.; Liehr, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 1, pp. 35–52. [Google Scholar]
- Zwick, M.S.; Hanson, R.E.; Islam-Faridi, M.N.; Stelly, D.M.; Wing, R.A.; Price, H.J.; McKnight, T.D. A rapid procedure for the isolation of C0t-1 DNA from plants. Genome 1997, 40, 138–142. [Google Scholar] [CrossRef] [Green Version]
- Viana, P.F.; Feldberg, E.; Cioffi, M.B.; De Carvalho, V.T.; Menezes, S.; Vogt, R.C.; Liehr, T.; Ezaz, T. The Amazonian Red Side-Necked Turtle Rhinemys rufipes (Spix, 1824) (Testudines, Chelidae) Has a GSD Sex-Determining Mechanism with an Ancient XY Sex Microchromosome System. Cells 2020, 9, 2088. [Google Scholar] [CrossRef] [PubMed]
- Toma, G.A.; De Moraes, R.L.R.; Sassi, F.D.M.C.; Bertollo, L.A.C.; De Oliveira, E.A.; Rab, P.; Sember, A.; Liehr, T.; Hatanaka, T.; Viana, P.F.; et al. Cytogenetics of the small-sized fish, Copeina guttata (Characiformes, Lebiasinidae): Novel insights into the karyotype differentiation of the family. PLoS ONE 2019, 14, e0226746. [Google Scholar] [CrossRef] [PubMed]
- Sassi, F.M.C.; Hatanaka, T.; Moraes, R.L.R.; Toma, G.A.; Oliveira, E.A.; Liehr, T.; Rab, P.; Bertollo, L.A.C.; Viana, P.F.; Feldberg, E.; et al. An insight into the chromosomal evolution of Lebiasinidae (Teleostei, Characiformes). Genes 2020, 11, 365. [Google Scholar] [CrossRef] [PubMed]
- Sember, A.; Oliveira, E.A.; Ráb, P.; Bertollo, L.; Freitas, N.L.; Viana, P.F.; Yano, C.F.; Hatanaka, T.; Marinho, M.; de Moraes, R.; et al. Centric Fusions behind the Karyotype Evolution of Neotropical Nannostomus Pencilfishes (Characiforme, Lebiasinidae): First Insights from a Molecular Cytogenetic Perspective. Genes 2020, 11, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- Beçak, W. Constituição cromossômica e mecanismo de determinação do sexo em ofídios sul-americanos. I. Aspectos cariotípicos. Mem. Inst. Butantan. 1965, 32, 37–78. [Google Scholar]
- Hynková, I.; Starostová, Z.; Frynta, D. Mitochondrial DNA Variation Reveals Recent Evolutionary History of Main Boa constrictor Clades. Zool. Sci. 2009, 26, 623–631. [Google Scholar] [CrossRef]
- Suárez-Atilano, M.; Burbrink, F.; Vázquez-Domínguez, E. Phylogeographical structure within Boa constrictor imperator across the lowlands and mountains of Central America and Mexico. J. Biogeogr. 2014, 41, 2371–2384. [Google Scholar] [CrossRef]
- Burbrink, F.T. Inferring the phylogenetic position of Boa constrictor among the Boinae. Mol. Phylogenet. Evol. 2005, 34, 167–180. [Google Scholar] [CrossRef]
- Viana, P.F.; Ezaz, T.; Cioffi, M.D.B.; Liehr, T.; Al-Rikabi, A.; Goll, L.G.; Rocha, A.M.; Feldberg, E. Landscape of snake’ sex chromosomes evolution spanning 85 MYR reveals ancestry of sequences despite distinct evolutionary trajectories. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Booth, W.; Million, L.; Reynolds, R.G.; Burghardt, G.M.; Vargo, E.L.; Schal, C.; Tzika, A.C.; Schuett, G.W. Consecutive Virgin Births in the New World Boid Snake, the Colombian Rainbow Boa, Epicrates maurus. J. Hered. 2011, 102, 759–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibata, H.; Sakata, S.; Hirano, Y.; Nitasaka, E.; Sakabe, A. Facultative parthenogenesis validated by DNA analyses in the green anaconda (Eunectes murinus). PLoS ONE 2017, 12, e0189654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seixas, F.; Morinha, F.; Luis, C.; Alvura, N.; dos Anjos Pires, M. DNA-validated parthenogenesis: First case in a captive Cuban boa (Chilabothrus angulifer). bioRxiv 2020, 751529. [Google Scholar] [CrossRef]
- Augstenová, B.; Pokorná, M.J.; Altmanová, M.; Frynta, D.; Rovatsos, M.; Kratochvíl, L. ZW, XY, and yet ZW: Sex chromosome evolution in snakes even more complicated. Evolution 2018, 72, 1701–1707. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viana, P.F.; Ezaz, T.; Cioffi, M.d.B.; Liehr, T.; Al-Rikabi, A.; Tavares-Pinheiro, R.; Bertollo, L.A.C.; Feldberg, E. Revisiting the Karyotype Evolution of Neotropical Boid Snakes: A Puzzle Mediated by Chromosomal Fissions. Cells 2020, 9, 2268. https://doi.org/10.3390/cells9102268
Viana PF, Ezaz T, Cioffi MdB, Liehr T, Al-Rikabi A, Tavares-Pinheiro R, Bertollo LAC, Feldberg E. Revisiting the Karyotype Evolution of Neotropical Boid Snakes: A Puzzle Mediated by Chromosomal Fissions. Cells. 2020; 9(10):2268. https://doi.org/10.3390/cells9102268
Chicago/Turabian StyleViana, Patrik F., Tariq Ezaz, Marcelo de Bello Cioffi, Thomas Liehr, Ahmed Al-Rikabi, Rodrigo Tavares-Pinheiro, Luiz Antônio Carlos Bertollo, and Eliana Feldberg. 2020. "Revisiting the Karyotype Evolution of Neotropical Boid Snakes: A Puzzle Mediated by Chromosomal Fissions" Cells 9, no. 10: 2268. https://doi.org/10.3390/cells9102268