
Perivascular Adipose Tissue and Vascular Smooth Muscle Tone: Friends or Foes?

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
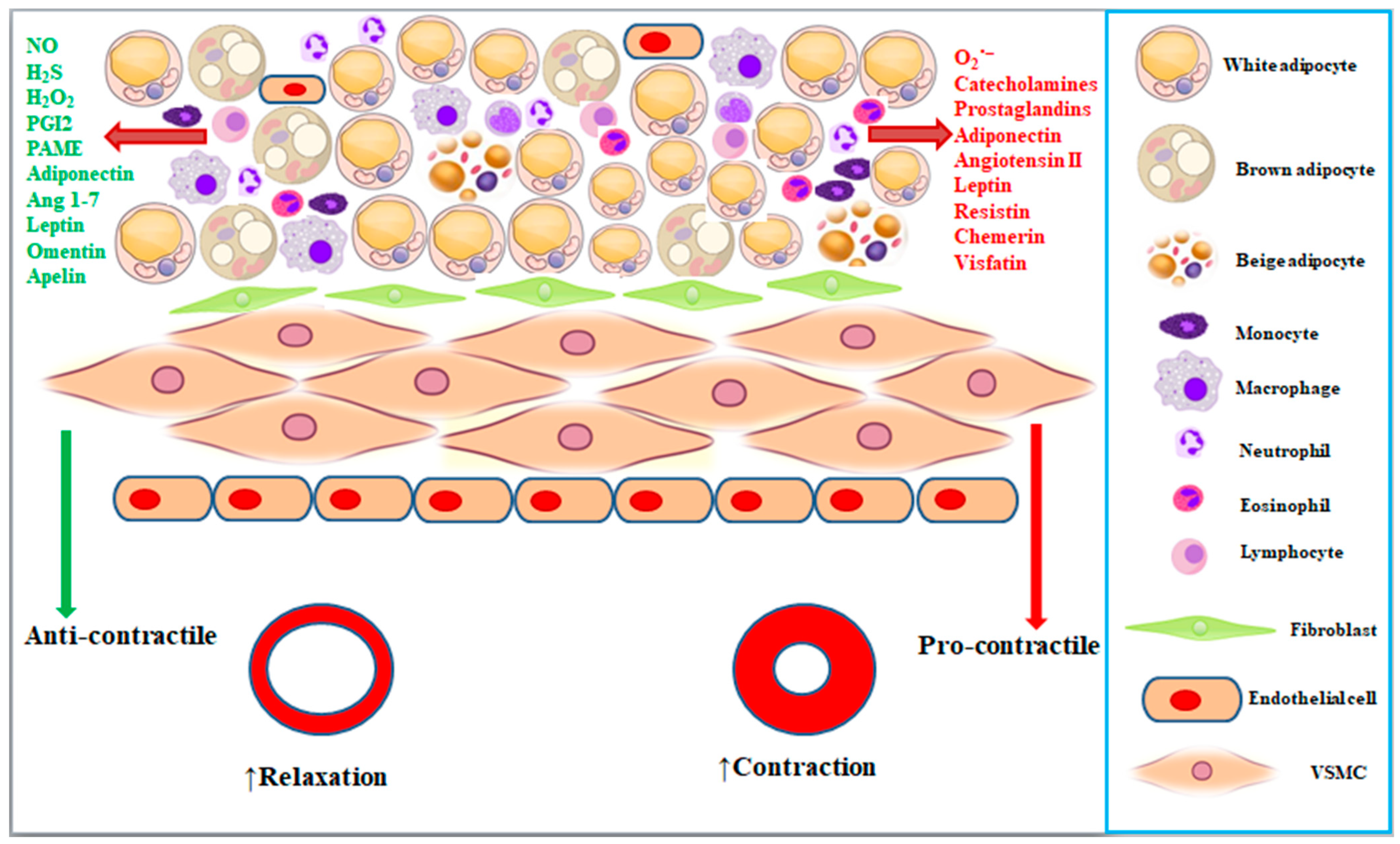
2. Characteristics of Perivascular Adipose Tissues
3. Perivascular Adipose Tissue Regulates Vascular Tone

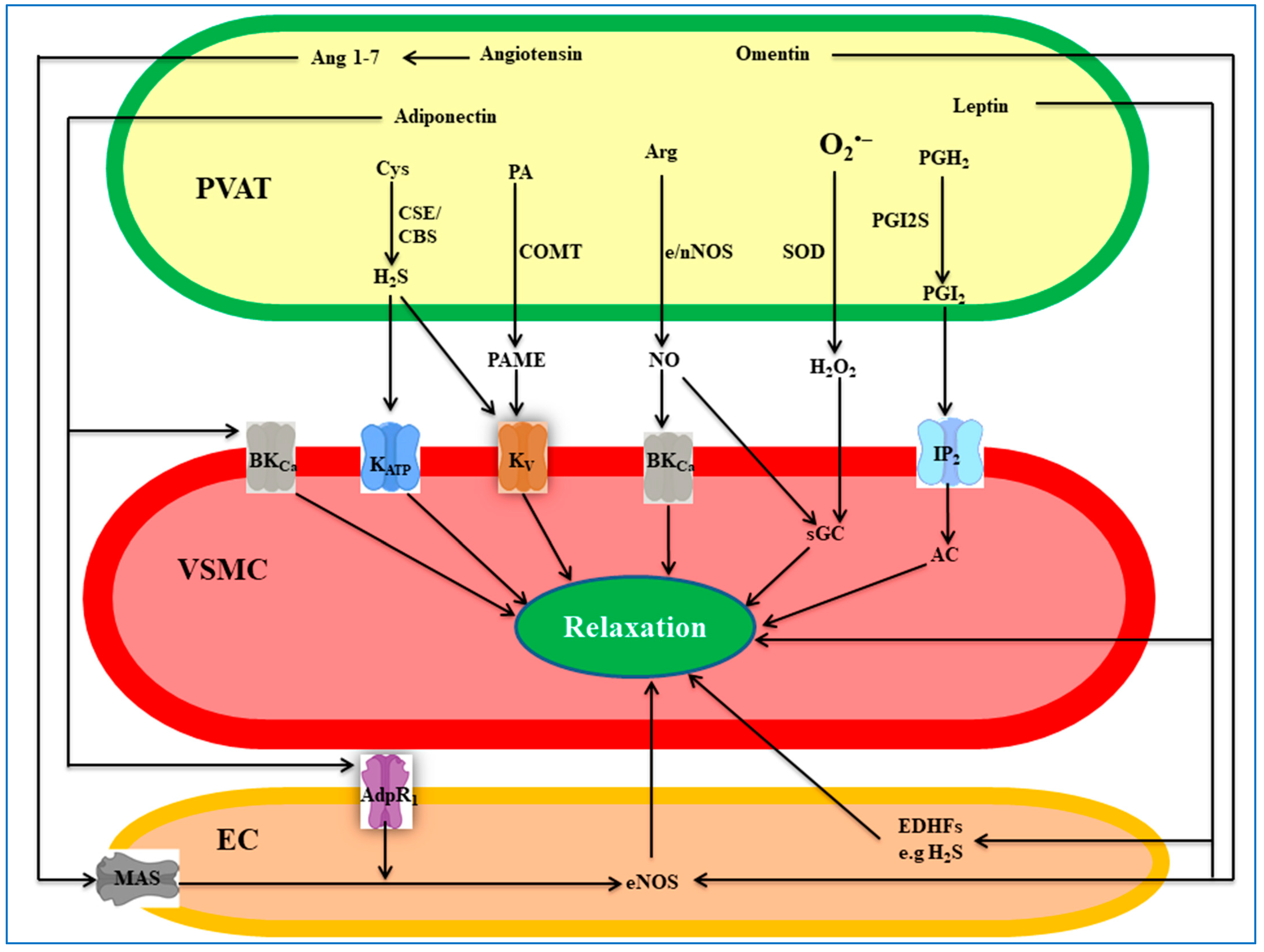
4. PVAT-Derived Anti-Contractile Factors
4.1. Nitric Oxide
4.2. Hydrogen Sulfide
4.3. Hydrogen Peroxide
4.4. Prostanoids
4.5. Palmitic Acid Methyl Ester
4.6. Angiotensin 1-7
4.7. Adiponectin
4.8. Leptin
4.9. Omentin
4.10. Other Factors/Mechanisms
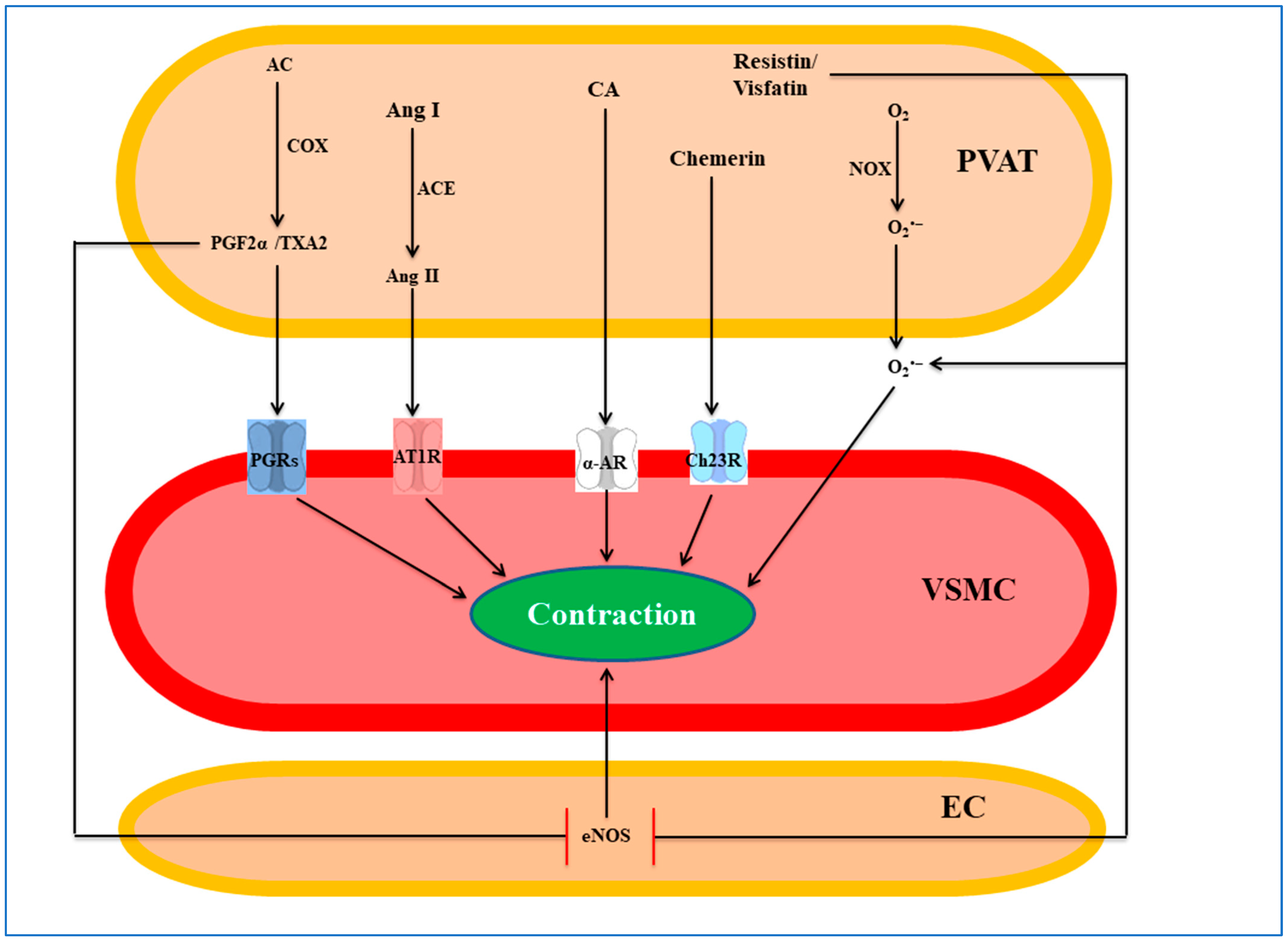
5. PVAT-Derived Pro-Contractile Factors
5.1. Superoxide Anion
5.2. Angiotensin II
5.3. Prostaglandins
5.4. Catecholamines
5.5. Chemerin
5.6. Resistin
5.7. Visfatin
5.8. Other Factors/Mechanisms
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Milutinović, A.; Šuput, D.; Zorc-Pleskovič, R. Pathogenesis of atherosclerosis in the tunica intima, media, and adventitia of coronary arteries: An updated review. Bosn. J. Basic Med. Sci. 2020, 20, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Hillock-Watling, C.; Gotlieb, A.I. The pathobiology of perivascular adipose tissue (PVAT), the fourth layer of the blood vessel wall. Cardiovasc. Pathol. 2022, 61, 107459. [Google Scholar] [CrossRef]
- Nava, E.; Llorens, S. The Local Regulation of Vascular Function: From an Inside-Outside to an Outside-Inside Model. Front. Physiol. 2019, 10, 729. [Google Scholar] [CrossRef]
- Sheng, Y.; Zhu, L. The crosstalk between autonomic nervous system and blood vessels. Int. J. Physiol. Pathophysiol. Pharmacol. 2018, 10, 17–28. [Google Scholar]
- Nava, E.; Llorens, S. The paracrine control of vascular motion. A historical perspective. Pharmacol. Res. 2016, 113, 125–145. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.K.; Zhou, Z.; Zhang, J.; Zeng, R.; Wu, J.; Eitzman, D.T.; Chen, Y.E.; Chang, L. Perivascular Adipose Tissue in Vascular Function and Disease. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1621–1630. [Google Scholar] [CrossRef]
- Soltis, E.E.; Cassis, L.A. Influence of perivascular adipose tissue on rat aortic smooth muscle responsiveness. Clin. Exp. Hypertens. A 1991, 13, 277–296. [Google Scholar] [CrossRef]
- Löhn, M.; Dubrovska, G.; Lauterbach, B.; Luft, F.C.; Gollasch, M.; Sharma, A.M. Periadventitial fat releases a vascular relaxing factor. FASEB J. 2002, 16, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.J.; Lu, C.; Su, L.Y.; Sharma, A.M.; Lee, R.M. Modulation of vascular function by perivascular adipose tissue: The role of endothelium and hydrogen peroxide. Br. J. Pharmacol. 2007, 151, 323–331. [Google Scholar] [CrossRef]
- Ozen, G.; Daci, A.; Norel, X.; Topal, G. Human perivascular adipose tissue dysfunction as a cause of vascular disease: Focus on vascular tone and wall remodeling. Eur. J. Pharmacol. 2015, 766, 16–24. [Google Scholar] [CrossRef]
- Ahmed, A.; Fusi, F.; Valoti, M. Perivascular adipose tissue modulates the effects of flavonoids on rat aorta rings: Role of superoxide anion and β(3) receptors. Pharmacol. Res. 2022, 180, 21. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.K.; Bakar, H.A.; Gollasch, M.; Huang, Y. Perivascular Adipose Tissue: The Sixth Man of the Cardiovascular System. Cardiovasc. Drugs Ther. 2018, 32, 481–502. [Google Scholar] [CrossRef] [PubMed]
- Zaborska, K.E.; Wareing, M.; Edwards, G.; Austin, C. Loss of anti-contractile effect of perivascular adipose tissue in offspring of obese rats. Int. J. Obes. 2016, 40, 1205–1214. [Google Scholar] [CrossRef]
- Ramirez, J.G.; O’Malley, E.J.; Ho, W.S.V. Pro-contractile effects of perivascular fat in health and disease. Br. J. Pharmacol. 2017, 174, 3482–3495. [Google Scholar] [CrossRef] [PubMed]
- Leandro, A.; Azul, L.; Fernandes, R.; Seiça, R.; Sena, C. Contribution of perivascular adipose tissue to vascular dysfunction in type 2 diabetes. Free Rad. Biol. Med. 2018, 120, S162. [Google Scholar] [CrossRef]
- Da Costa, R.M.; Fais, R.S.; Dechandt, C.R.P.; Louzada-Junior, P.; Alberici, L.C.; Lobato, N.S.; Tostes, R.C. Increased mitochondrial ROS generation mediates the loss of the anti-contractile effects of perivascular adipose tissue in high-fat diet obese mice. Br. J. Pharmacol. 2017, 174, 3527–3541. [Google Scholar] [CrossRef] [PubMed]
- Nosalski, R.; Guzik, T.J. Perivascular adipose tissue inflammation in vascular disease. Br. J. Pharmacol. 2017, 174, 3496–3513. [Google Scholar] [CrossRef]
- Chang, L.; Garcia-Barrio, M.T.; Chen, Y.E. Perivascular Adipose Tissue Regulates Vascular Function by Targeting Vascular Smooth Muscle Cells. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 1094–1109. [Google Scholar] [CrossRef]
- Adachi, Y.; Ueda, K.; Nomura, S.; Ito, K.; Katoh, M.; Katagiri, M.; Yamada, S.; Hashimoto, M.; Zhai, B.; Numata, G.; et al. Beiging of perivascular adipose tissue regulates its inflammation and vascular remodeling. Nat. Commun. 2022, 13, 5117. [Google Scholar] [CrossRef]
- Miao, C.Y.; Li, Z.Y. The role of perivascular adipose tissue in vascular smooth muscle cell growth. Br. J. Pharmacol. 2012, 165, 643–658. [Google Scholar] [CrossRef]
- Qi, X.Y.; Qu, S.L.; Xiong, W.H.; Rom, O.; Chang, L.; Jiang, Z.S. Perivascular adipose tissue (PVAT) in atherosclerosis: A double-edged sword. Cardiovasc. Diabetol. 2018, 17, 134. [Google Scholar] [CrossRef]
- Cao, Z.F.H.; Stoffel, E.; Cohen, P. Role of Perivascular Adipose Tissue in Vascular Physiology and Pathology. Hypertension 2017, 69, 770–777. [Google Scholar] [CrossRef]
- Stanek, A.; Brożyna-Tkaczyk, K.; Myśliński, W. The Role of Obesity-Induced Perivascular Adipose Tissue (PVAT) Dysfunction in Vascular Homeostasis. Nutrients 2021, 13, 3843. [Google Scholar] [CrossRef]
- Gil-Ortega, M.; Somoza, B.; Huang, Y.; Gollasch, M.; Fernández-Alfonso, M.S. Regional differences in perivascular adipose tissue impacting vascular homeostasis. Trends Endocrinol. Metab. 2015, 26, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Rajsheker, S.; Manka, D.; Blomkalns, A.L.; Chatterjee, T.K.; Stoll, L.L.; Weintraub, N.L. Crosstalk between perivascular adipose tissue and blood vessels. Curr. Opin. Pharmacol. 2010, 10, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Margaritis, M.; Antonopoulos, A.S.; Digby, J.; Lee, R.; Reilly, S.; Coutinho, P.; Shirodaria, C.; Sayeed, R.; Petrou, M.; De Silva, R.; et al. Interactions Between Vascular Wall and Perivascular Adipose Tissue Reveal Novel Roles for Adiponectin in the Regulation of Endothelial Nitric Oxide Synthase Function in Human Vessels. Circulation 2013, 127, 2209–2221. [Google Scholar] [CrossRef] [PubMed]
- Corvera, S. Cellular Heterogeneity in Adipose Tissues. Annu. Rev. Physiol. 2021, 83, 257–278. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.K.; Jin, Y.; Watts, S.W.; Rockwell, C.E. Naïve, Regulatory, Activated, and Memory Immune Cells Co-exist in PVATs That Are Comparable in Density to Non-PVAT Fats in Health. Front. Physiol. 2020, 11, 58. [Google Scholar] [CrossRef] [PubMed]
- Farias-Itao, D.S.; Pasqualucci, C.A.; Nishizawa, A.; da Silva, L.F.F.; Campos, F.M.; Bittencourt, M.S.; da Silva, K.C.S.; Leite, R.E.P.; Grinberg, L.T.; de Lucena Ferretti-Rebustini, R.E.; et al. B Lymphocytes and Macrophages in the Perivascular Adipose Tissue Are Associated With Coronary Atherosclerosis: An Autopsy Study. J. Am. Heart Assoc. 2019, 8, e013793. [Google Scholar] [CrossRef] [PubMed]
- Farias-Itao, D.S.; Pasqualucci, C.A.; Andrade, R.; da Silva, L.F.F.; Yahagi-Estevam, M.; Lage, S.H.G.; Leite, R.E.P.; Campo, A.B.; Suemoto, C.K. Macrophage Polarization in the Perivascular Fat Was Associated With Coronary Atherosclerosis. J. Am. Heart Assoc. 2022, 11, e023274. [Google Scholar] [CrossRef]
- Qiu, T.; Li, M.; Tanner, M.A.; Yang, Y.; Sowers, J.R.; Korthuis, R.J.; Hill, M.A. Depletion of dendritic cells in perivascular adipose tissue improves arterial relaxation responses in type 2 diabetic mice. Metab. Clin. Exp. 2018, 85, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Shi, H.; Winkler, M.A.; Lee, R.; Weintraub, N.L. Perivascular Adipose Tissue and Vascular Perturbation/Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2020, 40, 2569–2576. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ma, Z.; Zhu, Y.Z. Regional Heterogeneity of Perivascular Adipose Tissue: Morphology, Origin, and Secretome. Front. Pharmacol. 2021, 12, 697720. [Google Scholar] [CrossRef]
- Fu, M.; Xu, L.; Chen, X.; Han, W.; Ruan, C.; Li, J.; Cai, C.; Ye, M.; Gao, P. Neural Crest Cells Differentiate Into Brown Adipocytes and Contribute to Periaortic Arch Adipose Tissue Formation. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1629–1644. [Google Scholar] [CrossRef]
- Chang, L.; Villacorta, L.; Li, R.; Hamblin, M.; Xu, W.; Dou, C.; Zhang, J.; Wu, J.; Zeng, R.; Chen, Y.E. Loss of perivascular adipose tissue on peroxisome proliferator-activated receptor-γ deletion in smooth muscle cells impairs intravascular thermoregulation and enhances atherosclerosis. Circulation 2012, 126, 1067–1078. [Google Scholar] [CrossRef]
- Ye, M.; Ruan, C.C.; Fu, M.; Xu, L.; Chen, D.; Zhu, M.; Zhu, D.; Gao, P. Developmental and functional characteristics of the thoracic aorta perivascular adipocyte. Cell. Mol. Life Sci. 2019, 76, 777–789. [Google Scholar] [CrossRef]
- Angueira, A.R.; Sakers, A.P.; Holman, C.D.; Cheng, L.; Arbocco, M.N.; Shamsi, F.; Lynes, M.D.; Shrestha, R.; Okada, C.; Batmanov, K.; et al. Defining the lineage of thermogenic perivascular adipose tissue. Nat. Metabol. 2021, 3, 469–484. [Google Scholar] [CrossRef]
- Saxton, S.N.; Withers, S.B.; Heagerty, A.M. Perivascular Adipose Tissue Anticontractile Function Is Mediated by Both Endothelial and Neuronal Nitric Oxide Synthase Isoforms. J. Vasc. Res. 2022, 59, 288–302. [Google Scholar] [CrossRef]
- Araujo, H.N.; Victório, J.A.; Valgas da Silva, C.P.; Sponton, A.C.S.; Vettorazzi, J.F.; de Moraes, C.; Davel, A.P.; Zanesco, A.; Delbin, M.A. Anti-contractile effects of perivascular adipose tissue in thoracic aorta from rats fed a high-fat diet: Role of aerobic exercise training. Clin. Exp. Pharmacol. Physiol. 2018, 45, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Kociszewska, K.; Deja, M.A.; Malinowski, M.; Kowalówka, A. Vasorelaxing properties of the perivascular tissue of the human radial artery. Eur. J. Cardiothorac. Surg. 2022, 61, 1423–1429. [Google Scholar] [CrossRef]
- Malinowski, M.; Deja, M.A.; Gołba, K.S.; Roleder, T.; Biernat, J.; Woś, S. Perivascular tissue of internal thoracic artery releases potent nitric oxide and prostacyclin-independent anticontractile factor. Eur. J. Cardiothorac. Surg. 2008, 33, 225–231. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.-J.; Zeng, Z.-h.; Teoh, K.; Sharma, A.M.; Abouzahr, L.; Cybulsky, I.; Lamy, A.; Semelhago, L.; Lee, R.M.K.W. Perivascular adipose tissue modulates vascular function in the human internal thoracic artery. J. Thorac. Cardiovasc. Surg. 2005, 130, 1130–1136. [Google Scholar] [CrossRef]
- Agabiti-Rosei, C.; De Ciuceis, C.; Rossini, C.; Porteri, E.; Rodella, L.F.; Withers, S.B.; Heagerty, A.M.; Favero, G.; Agabiti-Rosei, E.; Rizzoni, D.; et al. Anticontractile activity of perivascular fat in obese mice and the effect of long-term treatment with melatonin. J. Hypertens. 2014, 32, 1264–1274. [Google Scholar] [CrossRef]
- Bussey, C.E.; Withers, S.B.; Saxton, S.N.; Bodagh, N.; Aldous, R.G.; Heagerty, A.M. β(3)-Adrenoceptor stimulation of perivascular adipocytes leads to increased fat cell-derived NO and vascular relaxation in small arteries. Br. J. Pharmacol. 2018, 175, 3685–3698. [Google Scholar] [CrossRef]
- Dubrovska, G.; Verlohren, S.; Luft, F.C.; Gollasch, M. Mechanisms of ADRF release from rat aortic adventitial adipose tissue. Am. J. Physiol. Heart Circul. Physiol. 2004, 286, H1107–H1113. [Google Scholar] [CrossRef] [PubMed]
- Withers, S.B.; Simpson, L.; Fattah, S.; Werner, M.E.; Heagerty, A.M. cGMP-dependent protein kinase (PKG) mediates the anticontractile capacity of perivascular adipose tissue. Cardiovasc. Res. 2014, 101, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Lynch, F.M.; Withers, S.B.; Yao, Z.; Werner, M.E.; Edwards, G.; Weston, A.H.; Heagerty, A.M. Perivascular adipose tissue-derived adiponectin activates BKCa channels to induce anticontractile responses. Am. J. Physiol. Heart Circul. Physiol. 2013, 304, H786–H795. [Google Scholar] [CrossRef]
- Tsvetkov, D.; Tano, J.Y.; Kassmann, M.; Wang, N.; Schubert, R.; Gollasch, M. The Role of DPO-1 and XE991-Sensitive Potassium Channels in Perivascular Adipose Tissue-Mediated Regulation of Vascular Tone. Front. Physiol. 2016, 7, 335. [Google Scholar] [CrossRef]
- Owen, M.K.; Witzmann, F.A.; McKenney, M.L.; Lai, X.; Berwick, Z.C.; Moberly, S.P.; Alloosh, M.; Sturek, M.; Tune, J.D. Perivascular adipose tissue potentiates contraction of coronary vascular smooth muscle: Influence of obesity. Circulation 2013, 128, 9–18. [Google Scholar] [CrossRef]
- Noblet, J.N.; Owen, M.K.; Goodwill, A.G.; Sassoon, D.; Tune, J.D. Lean and Obese Coronary Perivascular Adipose Tissue Impairs Vasodilation via Differential Inhibition of Vascular Smooth Muscle K+ Channels. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1393–1400. [Google Scholar] [CrossRef]
- Al-Jarallah, A.; Oommen, E.; Chacko Verghese, L.; Oriowo, M.A. Role of Extracellular Calcium and Calcium Sensitization in the Anti-Contractile Effect of Perivascular Adipose Tissue in Pregnant Rat Aorta. Pharmacology 2019, 104, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Victorio, J.A.; Fontes, M.T.; Rossoni, L.V.; Davel, A.P. Different Anti-Contractile Function and Nitric Oxide Production of Thoracic and Abdominal Perivascular Adipose Tissues. Front. Physiol. 2016, 7, 295. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.A.; Randall, M.D.; Roberts, R.E. Sex differences in the regulation of porcine coronary artery tone by perivascular adipose tissue: A role of adiponectin? Br. J. Pharmacol. 2017, 174, 2773–2783. [Google Scholar] [CrossRef]
- Szasz, T.; Carrillo-Sepulveda, M.A.; Webb, R.C. Abstract 260: Aging Decreases the Anticontractile Effect of Perivascular Adipose Tissue in the Mouse Aorta. Hypertension 2012, 60, A260. [Google Scholar] [CrossRef]
- Li, R.; Andersen, I.; Aleke, J.; Golubinskaya, V.; Gustafsson, H.; Nilsson, H. Reduced anti-contractile effect of perivascular adipose tissue on mesenteric small arteries from spontaneously hypertensive rats: Role of Kv7 channels. Eur. J. Pharmacol. 2012, 698, 310–315. [Google Scholar] [CrossRef]
- Zhao, Y.; Vanhoutte, P.M.; Leung, S.W.S. Vascular nitric oxide: Beyond eNOS. J. Pharmacol. Sci. 2015, 129, 83–94. [Google Scholar] [CrossRef]
- Gil-Ortega, M.; Stucchi, P.; Guzmán-Ruiz, R.; Cano, V.; Arribas, S.; González, M.C.; Ruiz-Gayo, M.; Fernandez-Alfonso, M.S.; Somoza, B. Adaptative nitric oxide overproduction in perivascular adipose tissue during early diet-induced obesity. Endocrinology 2010, 151, 3299–3306. [Google Scholar] [CrossRef]
- Virdis, A.; Duranti, E.; Rossi, C.; Dell’Agnello, U.; Santini, E.; Anselmino, M.; Chiarugi, M.; Taddei, S.; and Solini, A. Tumour necrosis factor-alpha participates on the endothelin-1/nitric oxide imbalance in small arteries from obese patients: Role of perivascular adipose tissue. Eur. Heart J. 2015, 36, 784–794. [Google Scholar] [CrossRef] [PubMed]
- Aghamohammadzadeh, R.; Unwin, R.D.; Greenstein, A.S.; Heagerty, A.M. Effects of Obesity on Perivascular Adipose Tissue Vasorelaxant Function: Nitric Oxide, Inflammation and Elevated Systemic Blood Pressure. J. Vasc. Res. 2015, 52, 299–305. [Google Scholar] [CrossRef]
- Man, A.W.C.; Zhou, Y.; Xia, N.; Li, H. Endothelial Nitric Oxide Synthase in the Perivascular Adipose Tissue. Biomedicines 2022, 10, 1754. [Google Scholar] [CrossRef]
- Dashwood, M.R.; Dooley, A.; Shi-Wen, X.; Abraham, D.J.; Souza, D.S. Does periadventitial fat-derived nitric oxide play a role in improved saphenous vein graft patency in patients undergoing coronary artery bypass surgery? J. Vasc. Res. 2007, 44, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Bussey, C.E.; Withers, S.B.; Aldous, R.G.; Edwards, G.; Heagerty, A.M. Obesity-Related Perivascular Adipose Tissue Damage Is Reversed by Sustained Weight Loss in the Rat. Arterioscler. Thromb. Vasc. Biol. 2016, 36, 1377–1385. [Google Scholar] [CrossRef] [PubMed]
- Nóbrega, N.; Araújo, N.F.; Reis, D.; Facine, L.M.; Miranda, C.A.S.; Mota, G.C.; Aires, R.D.; Capettini, L.D.S.A.; dos Santos Cruz, J.; Bonaventura, D. Hydrogen peroxide and nitric oxide induce anticontractile effect of perivascular adipose tissue via renin angiotensin system activation. Nitric Oxide 2019, 84, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Barp, C.G.; Bonaventura, D.; Assreuy, J. NO, ROS, RAS, and PVAT: More Than a Soup of Letters. Front. Physiol. 2021, 12, 640021. [Google Scholar] [CrossRef]
- Sousa, A.S.; Sponton, A.C.S.; Trifone, C.B.; Delbin, M.A. Aerobic Exercise Training Prevents Perivascular Adipose Tissue-Induced Endothelial Dysfunction in Thoracic Aorta of Obese Mice. Front. Physiol. 2019, 10, 640021. [Google Scholar] [CrossRef] [PubMed]
- Weston, A.H.; Egner, I.; Dong, Y.; Porter, E.L.; Heagerty, A.M.; Edwards, G. Stimulated release of a hyperpolarizing factor (ADHF) from mesenteric artery perivascular adipose tissue: Involvement of myocyte BKCa channels and adiponectin. Br. J. Pharmacol. 2013, 169, 1500–1509. [Google Scholar] [CrossRef]
- Xia, N.; Li, H. The role of perivascular adipose tissue in obesity-induced vascular dysfunction. Br. J. Pharmacol. 2017, 174, 3425–3442. [Google Scholar] [CrossRef]
- Withers, S.B.; Bussey, C.E.; Saxton, S.N.; Melrose, H.M.; Watkins, A.E.; Heagerty, A.M. Mechanisms of adiponectin-associated perivascular function in vascular disease. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1637–1642. [Google Scholar] [CrossRef]
- Cao, X.; Ding, L.; Xie, Z.Z.; Yang, Y.; Whiteman, M.; Moore, P.K.; Bian, J.S. A Review of Hydrogen Sulfide Synthesis, Metabolism, and Measurement: Is Modulation of Hydrogen Sulfide a Novel Therapeutic for Cancer? Antioxid. Redox Signal. 2019, 31, 1–38. [Google Scholar] [CrossRef]
- Bełtowski, J. Endogenous hydrogen sulfide in perivascular adipose tissue: Role in the regulation of vascular tone in physiology and pathology. Can. J. Physiol. Pharmacol. 2013, 91, 889–898. [Google Scholar] [CrossRef]
- Streeter, E.; Hart, J.; Badoer, E. An investigation of the mechanisms of hydrogen sulfide-induced vasorelaxation in rat middle cerebral arteries. Naunyn-Schmiedeb. Arch. Pharmacol. 2012, 385, 991–1002. [Google Scholar] [CrossRef] [PubMed]
- Lv, B.; Chen, S.; Tang, C.; Jin, H.; Du, J.; Huang, Y. Hydrogen sulfide and vascular regulation—An update. J. Adv. Res. 2021, 27, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Cacanyiova, S.; Krskova, K.; Zórad, S.; Frimmel, K.; Drobna, M.; Valaskova, Z.; Misak, A.; Golas, S.; Breza, J.; Breza, J., Jr.; et al. Arterial Hypertension and Plasma Glucose Modulate the Vasoactive Effects of Nitroso-Sulfide Coupled Signaling in Human Intrarenal Arteries. Molecules 2020, 25, 2886. [Google Scholar] [CrossRef] [PubMed]
- Schleifenbaum, J.; Köhn, C.; Voblova, N.; Dubrovska, G.; Zavarirskaya, O.; Gloe, T.; Crean, C.S.; Luft, F.C.; Huang, Y.; Schubert, R.; et al. Systemic peripheral artery relaxation by KCNQ channel openers and hydrogen sulfide. J. Hypertens. 2010, 28, 1875–1882. [Google Scholar] [CrossRef]
- Fang, L.; Zhao, J.; Chen, Y.; Ma, T.; Guoheng, X.; Tang, C.; Liu, X.; Geng, B. Hydrogen sulfide derived from periadventitial adipose tissue is a vasodilator. J. Hypertens. 2009, 27, 2174–2185. [Google Scholar] [CrossRef]
- Emilova, R.; Dimitrova, D.; Mladenov, M.; Daneva, T.; Schubert, R.; Gagov, H. Cystathionine gamma-lyase of perivascular adipose tissue with reversed regulatory effect in diabetic rat artery. Biotechnol. Biotechnol. Equip. 2015, 29, 147–151. [Google Scholar] [CrossRef]
- Cacanyiova, S.; Majzunova, M.; Golas, S.; Berenyiova, A. The role of perivascular adipose tissue and endogenous hydrogen sulfide in vasoactive responses of isolated mesenteric arteries in normotensive and spontaneously hypertensive rats. J. Physiol. Pharmacol. 2019, 70, 20. [Google Scholar] [CrossRef]
- Köhn, C.; Schleifenbaum, J.; Szijártó, I.A.; Markó, L.; Dubrovska, G.; Huang, Y.; Gollasch, M. Differential Effects of Cystathionine-γ-lyase–Dependent Vasodilatory H2S in Periadventitial Vasoregulation of Rat and Mouse Aortas. PLoS ONE 2012, 7, e41951. [Google Scholar] [CrossRef]
- Donovan, J.; Wong, P.S.; Garle, M.J.; Alexander, S.P.H.; Dunn, W.R.; Ralevic, V. Coronary artery hypoxic vasorelaxation is augmented by perivascular adipose tissue through a mechanism involving hydrogen sulphide and cystathionine-β-synthase. Acta Physiol. 2018, 224, 11. [Google Scholar] [CrossRef]
- Bełtowski, J.; Guranowski, A.; Jamroz-Wiśniewska, A.; Wolski, A.; Hałas, K. Hydrogen-sulfide-mediated vasodilatory effect of nucleoside 5′-monophosphorothioates in perivascular adipose tissue. Can. J. Physiol. Pharmacol. 2015, 93, 585–595. [Google Scholar] [CrossRef]
- Liao, J.; Yin, H.; Huang, J.; Hu, M. Dysfunction of perivascular adipose tissue in mesenteric artery is restored by aerobic exercise in high-fat diet induced obesity. Clin. Exp. Pharmacol. Physiol. 2021, 48, 697–703. [Google Scholar] [CrossRef] [PubMed]
- Wójcicka, G.; Jamroz-Wiśniewska, A.; Atanasova, P.; Chaldakov, G.N.; Chylińska-Kula, B.; Bełtowski, J. Differential effects of statins on endogenous H2S formation in perivascular adipose tissue. Pharmacol. Res. 2011, 63, 68–76. [Google Scholar] [CrossRef]
- Bełtowski, J.; Jamroz-Wiśniewska, A. Modulation of h(2)s metabolism by statins: A new aspect of cardiovascular pharmacology. Antioxid. Red. Signal. 2012, 17, 81–94. [Google Scholar] [CrossRef]
- Perez-Vizcaino, F.; Cogolludo, A.; Moreno, L. Reactive oxygen species signaling in pulmonary vascular smooth muscle. Respir. Physiol. Neurobiol. 2010, 174, 212–220. [Google Scholar] [CrossRef] [PubMed]
- Bretón-Romero, R.; Lamas, S. Hydrogen peroxide signaling in vascular endothelial cells. Redox Biol. 2014, 2, 529–534. [Google Scholar] [CrossRef]
- Byon, C.H.; Heath, J.M.; Chen, Y. Redox signaling in cardiovascular pathophysiology: A focus on hydrogen peroxide and vascular smooth muscle cells. Redox Biol. 2016, 9, 244–253. [Google Scholar] [CrossRef]
- Park, S.W.; Noh, H.J.; Sung, D.J.; Kim, J.G.; Kim, J.M.; Ryu, S.-Y.; Kang, K.; Kim, B.; Bae, Y.M.; Cho, H. Hydrogen peroxide induces vasorelaxation by enhancing 4-aminopyridine-sensitive Kv currents through S-glutathionylation. Pflügers Arch. Eur. J. Physiol. 2015, 467, 285–297. [Google Scholar] [CrossRef]
- Wei, E.P.; Kontos, H.A.; Beckman, J.S. Mechanisms of cerebral vasodilation by superoxide, hydrogen peroxide, and peroxynitrite. Am. J. Physiol.-Heart Circul. Physiol. 1996, 271, H1262–H1266. [Google Scholar] [CrossRef]
- Dou, D.; Zheng, X.; Liu, J.; Xu, X.; Ye, L.; Gao, Y. Hydrogen peroxide enhances vasodilatation by increasing dimerization of cGMP-dependent protein kinase type Iα. Circ. J. 2012, 76, 1792–1798. [Google Scholar] [CrossRef] [PubMed]
- Iida, Y.; Katusic, Z.S. Mechanisms of Cerebral Arterial Relaxations to Hydrogen Peroxide. Stroke 2000, 31, 2224–2230. [Google Scholar] [CrossRef]
- Fujimoto, S.; Asano, T.; Sakai, M.; Sakurai, K.; Takagi, D.; Yoshimoto, N.; Itoh, T. Mechanisms of hydrogen peroxide-induced relaxation in rabbit mesenteric small artery. Eur. J. Pharmacol. 2001, 412, 291–300. [Google Scholar] [CrossRef]
- Phillips, S.A.; Somberg, L.B.; Hatoum, O.A.; Gutterman, D.D. Mechanisms of hydrogen peroxide induced vasoconstriction in human adipose resistance arteries. FASEB J. 2007, 21, A491. [Google Scholar] [CrossRef]
- Gao, Y.J.; Lee, R.M.K.W. Hydrogen peroxide induces a greater contraction in mesenteric arteries of spontaneously hypertensive rats through thromboxane A2 production. Br. J. Pharmacol. 2001, 134, 1639–1646. [Google Scholar] [CrossRef]
- Yang, Z.; Zheng, T.; Wang, J.; Zhang, A.; Altura, B.T.; Altura, B.M. Hydrogen peroxide induces contraction and raises [Ca2+]i in canine cerebral arterial smooth muscle: Participation of cellular signaling pathways. Naunyn-Schmiedeb. Arch. Pharmacol. 1999, 360, 646–653. [Google Scholar] [CrossRef]
- Costa, R.M.; Filgueira, F.P.; Tostes, R.C.; Carvalho, M.H.C.; Akamine, E.H.; Lobato, N.S. H2O2 generated from mitochondrial electron transport chain in thoracic perivascular adipose tissue is crucial for modulation of vascular smooth muscle contraction. Vasc. Pharmacol. 2016, 84, 28–37. [Google Scholar] [CrossRef]
- Ayala-Lopez, N.; Thompson, J.M.; Watts, S.W. Perivascular Adipose Tissue’s Impact on Norepinephrine-Induced Contraction of Mesenteric Resistance Arteries. Front. Physiol. 2017, 8, 37. [Google Scholar] [CrossRef] [PubMed]
- Reis Costa, D.E.F.; Silveira, A.L.M.; Campos, G.P.; Nóbrega, N.R.C.; de Araújo, N.F.; de Figueiredo Borges, L.; dos Capettini, L.S.A.; Ferreira, A.V.M.; Bonaventura, D. High-Carbohydrate Diet Enhanced the Anticontractile Effect of Perivascular Adipose Tissue Through Activation of Renin-Angiotensin System. Front. Physiol. 2021, 11, 628101. [Google Scholar] [CrossRef] [PubMed]
- Gonzaga, N.A.; Awata, W.M.C.; do Vale, G.T.; Marchi, K.C.; Muniz, J.J.; Tanus-Santos, J.E.; Tirapelli, C.R. Perivascular adipose tissue protects against the vascular dysfunction induced by acute ethanol intake: Role of hydrogen peroxide. Vascul. Pharmacol. 2018, 111, 44–53. [Google Scholar] [CrossRef] [PubMed]
- Kassam, S.I.; Lu, C.; Buckley, N.; Lee, R.M. The mechanisms of propofol-induced vascular relaxation and modulation by perivascular adipose tissue and endothelium. Anesth. Analg. 2011, 112, 1339–1345. [Google Scholar] [CrossRef]
- Li, Y.; Mihara, K.; Saifeddine, M.; Krawetz, A.; Lau, D.C.; Li, H.; Ding, H.; Triggle, C.R.; Hollenberg, M.D. Perivascular adipose tissue-derived relaxing factors: Release by peptide agonists via proteinase-activated receptor-2 (PAR2) and non-PAR2 mechanisms. Br. J. Pharmacol. 2011, 164, 1990–2002. [Google Scholar] [CrossRef]
- Emilova, R.; Dimitrova, D.Z.; Mladenov, M.; Hadzi-Petrushev, N.; Daneva, T.; Padeshki, P.; Schubert, R.; Chichova, M.; Lubomirov, L.; Simeonovska-Nikolova, D.; et al. Diabetes converts arterial regulation by perivascular adipose tissue from relaxation into H(2)O(2)-mediated contraction. Physiol. Res. 2016, 65, 799–807. [Google Scholar] [CrossRef]
- Stitham, J.; Midgett, C.; Martin, K.; Hwa, J. Prostacyclin: An Inflammatory Paradox. Front. Pharmacol. 2011, 2, 24. [Google Scholar] [CrossRef] [PubMed]
- Pluchart, H.; Khouri, C.; Blaise, S.; Roustit, M.; Cracowski, J.L. Targeting the Prostacyclin Pathway: Beyond Pulmonary Arterial Hypertension. Trends Pharmacol. Sci. 2017, 38, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.A.; Ali, F.; Bailey, L.; Moreno, L.; Harrington, L.S. Role of nitric oxide and prostacyclin as vasoactive hormones released by the endothelium. Exp. Physiol. 2008, 93, 141–147. [Google Scholar] [CrossRef]
- Zeng, Z.H.; Zhang, Z.H.; Luo, B.H.; He, W.K.; Liang, L.Y.; He, C.C.; Su, C.J. The functional changes of the perivascular adipose tissue in spontaneously hypertensive rats and the effects of atorvastatin therapy. Clin. Exp. Hypertens. 2009, 31, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Awata, W.M.C.; Gonzaga, N.A.; Borges, V.F.; Silva, C.B.P.; Tanus-Santos, J.E.; Cunha, F.Q.; Tirapelli, C.R. Perivascular adipose tissue contributes to lethal sepsis-induced vasoplegia in rats. Eur. J. Pharmacol. 2019, 863, 27. [Google Scholar] [CrossRef]
- Ozen, G.; Topal, G.; Gomez, I.; Ghorreshi, A.; Boukais, K.; Benyahia, C.; Kanyinda, L.; Longrois, D.; Teskin, O.; Uydes-Dogan, B.S.; et al. Control of human vascular tone by prostanoids derived from perivascular adipose tissue. Prostaglandins Other Lipid Mediat. 2013, 107, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Mendizábal, Y.; Llorens, S.; Nava, E. Vasoactive effects of prostaglandins from the perivascular fat of mesenteric resistance arteries in WKY and SHROB rats. Life Sci. 2013, 93, 1023–1032. [Google Scholar] [CrossRef]
- Lee, R.H.-C.; Couto e Silva, A.; Possoit, H.E.; Lerner, F.M.; Chen, P.-Y.; Azizbayeva, R.; Citadin, C.T.; Wu, C.Y.C.; Neumann, J.T.; Lin, H.W. Palmitic acid methyl ester is a novel neuroprotective agent against cardiac arrest. Prostaglandins Leukot. Essent. Fat. Acids 2019, 147, 6–14. [Google Scholar] [CrossRef]
- Liu, C.H.; Hsu, H.J.; Tseng, T.L.; Lin, T.; Weng, W.H.; Chen, M.F.; Lee, T.J.F. COMT-Catalyzed Palmitic Acid Methyl Ester Biosynthesis in Perivascular Adipose Tissue and its Potential Role Against Hypertension. J. Pharmacol. Exp. Ther. 2020, 373, 175–183. [Google Scholar] [CrossRef]
- Lee, Y.-C.; Chang, H.-H.; Chiang, C.-L.; Liu, C.-H.; Yeh, J.-I.; Chen, M.-F.; Chen, P.Y.; Kuo, J.S.; Lee, T.J. Role of Perivascular Adipose Tissue–Derived Methyl Palmitate in Vascular Tone Regulation and Pathogenesis of Hypertension. Circulation 2011, 124, 1160–1171. [Google Scholar] [CrossRef]
- Wang, N.; Kuczmanski, A.; Dubrovska, G.; Gollasch, M. Palmitic Acid Methyl Ester and Its Relation to Control of Tone of Human Visceral Arteries and Rat Aortas by Perivascular Adipose Tissue. Front. Physiol. 2018, 9, 583. [Google Scholar] [CrossRef] [PubMed]
- Ferrario, C.M.; Trask, A.J.; Jessup, J.A. Advances in biochemical and functional roles of angiotensin-converting enzyme 2 and angiotensin-(1-7) in regulation of cardiovascular function. Am. J. Physiol. Heart Circ. Physiol. 2005, 289, 29. [Google Scholar] [CrossRef]
- Santos, R.A.S.; Ferreira, A.J.; Verano-Braga, T.; Bader, M. Angiotensin-converting enzyme 2, angiotensin-(1–7) and Mas: New players of the renin–angiotensin system. J. Endocrinol. 2013, 216, R1–R17. [Google Scholar] [CrossRef] [PubMed]
- Gálvez-Prieto, B.; Bolbrinker, J.; Stucchi, P.; de Las Heras, A.I.; Merino, B.; Arribas, S.; Ruiz-Gayo, M.; Huber, M.; Wehland, M.; Kreutz, R.; et al. Comparative expression analysis of the renin-angiotensin system components between white and brown perivascular adipose tissue. J. Endocrinol. 2008, 197, 55–64. [Google Scholar] [CrossRef]
- Lee, R.M.; Lu, C.; Su, L.Y.; Gao, Y.J. Endothelium-dependent relaxation factor released by perivascular adipose tissue. J. Hypertens. 2009, 27, 782–790. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Su, L.Y.; Lee, R.M.; Gao, Y.J. Alterations in perivascular adipose tissue structure and function in hypertension. Eur. J. Pharmacol. 2011, 656, 68–73. [Google Scholar] [CrossRef]
- Lu, C.; Zhao, A.X.; Gao, Y.-J.; Lee, R.M.K.W. Modulation of vein function by perivascular adipose tissue. Eur. J. Pharmacol. 2011, 657, 111–116. [Google Scholar] [CrossRef]
- Lee, R.M.; Bader, M.; Alenina, N.; Santos, R.A.; Gao, Y.J.; Lu, C. Mas receptors in modulating relaxation induced by perivascular adipose tissue. Life Sci. 2011, 89, 467–472. [Google Scholar] [CrossRef]
- Santos, R.A.; Simoes e Silva, A.C.; Maric, C.; Silva, D.M.; Machado, R.P.; de Buhr, I.; Heringer-Walther, S.; Pinheiro, S.V.B.; Lopes, M.T.; Bader, M.; et al. Angiotensin-(1-7) is an endogenous ligand for the G protein-coupled receptor Mas. Proc. Natl. Acad. Sci. USA 2003, 100, 8258–8263. [Google Scholar] [CrossRef]
- Ayala-Lopez, N.; Pereira-Hicks, C.; Burnett, R.; Fink, G.D.; Watts, S.W. Human Perivascular Adipose Tissue Contains an Adrenergic System. FASEB J. 2016, 30, 712–719. [Google Scholar] [CrossRef]
- Achari, A.E.; Jain, S.K. Adiponectin, a Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Iwabu, M.; Okada-Iwabu, M.; Kadowaki, T. Adiponectin receptors: A review of their structure, function and how they work. Best Pract. Res. Clin. Endocrinol. Metab. 2014, 28, 15–23. [Google Scholar] [CrossRef]
- Fésüs, G.; Dubrovska, G.; Gorzelniak, K.; Kluge, R.; Huang, Y.; Luft, F.C.; Gollasch, M. Adiponectin is a novel humoral vasodilator. Cardiovas. Res. 2007, 75, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Greenstein, A.S.; Khavandi, K.; Withers, S.B.; Sonoyama, K.; Clancy, O.; Jeziorska, M.; Laing, I.; Yates, A.P.; Pemberton, P.W.; Malik, R.A.; et al. Local inflammation and hypoxia abolish the protective anticontractile properties of perivascular fat in obese patients. Circulation 2009, 119, 1661–1670. [Google Scholar] [CrossRef] [PubMed]
- Withers, S.B.; Forman, R.; Meza-Perez, S.; Sorobetea, D.; Sitnik, K.; Hopwood, T.; Lawrence, C.B.; Agace, W.W.; Else, K.J.; Heagerty, A.M.; et al. Eosinophils are key regulators of perivascular adipose tissue and vascular functionality. Sci. Rep. 2017, 7, 44571. [Google Scholar] [CrossRef]
- Almabrouk, T.A.M.; White, A.D.; Ugusman, A.B.; Skiba, D.S.; Katwan, O.J.; Alganga, H.; Guzik, T.J.; Touyz, R.M.; Salt, I.P.; Kennedy, S. High Fat Diet Attenuates the Anticontractile Activity of Aortic PVAT via a Mechanism Involving AMPK and Reduced Adiponectin Secretion. Front. Physiol. 2018, 9, 51. [Google Scholar] [CrossRef]
- Almabrouk, T.A.M.; Ugusman, A.B.; Katwan, O.J.; Salt, I.P.; Kennedy, S. Deletion of AMPKα1 attenuates the anticontractile effect of perivascular adipose tissue (PVAT) and reduces adiponectin release. Br. J. Pharmacol. 2017, 174, 3398–3410. [Google Scholar] [CrossRef]
- Sousa, A.S.; Sponton, A.C.S.; Delbin, M.A. Perivascular adipose tissue and microvascular endothelial dysfunction in obese mice: Beneficial effects of aerobic exercise in adiponectin receptor (AdipoR1) and peNOS(Ser1177). Clin. Exp. Pharmacol. Physiol. 2021, 48, 1430–1440. [Google Scholar] [CrossRef]
- Meziat, C.; Boulghobra, D.; Strock, E.; Battault, S.; Bornard, I.; Walther, G.; Reboul, C. Exercise training restores eNOS activation in the perivascular adipose tissue of obese rats: Impact on vascular function. Nitric Oxide 2019, 86, 63–67. [Google Scholar] [CrossRef]
- Loria, A.S.; Spradley, F.T.; Obi, I.E.; Becker, B.K.; De Miguel, C.; Speed, J.S.; Pollock, D.M.; Pollock, J.S. Maternal separation enhances anticontractile perivascular adipose tissue function in male rats on a high-fat diet. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2018, 315, R1085–R1095. [Google Scholar] [CrossRef] [PubMed]
- Badran, M.; Yassin, B.A.; Lin, D.T.S.; Kobor, M.S.; Ayas, N.; Laher, I. Gestational intermittent hypoxia induces endothelial dysfunction, reduces perivascular adiponectin and causes epigenetic changes in adult male offspring. J. Physiol. 2019, 597, 5349–5364. [Google Scholar] [CrossRef]
- Han, F.; Li, K.; Pan, R.; Xu, W.; Han, X.; Hou, N.; Sun, X. Calycosin directly improves perivascular adipose tissue dysfunction by upregulating the adiponectin/AMPK/eNOS pathway in obese mice. Food Funct. 2018, 9, 2409–2415. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Montagnani, M.; Funahashi, T.; Shimomura, I.; Quon, M.J. Adiponectin stimulates production of nitric oxide in vascular endothelial cells. J. Biol. Chem. 2003, 278, 45021–45026. [Google Scholar] [CrossRef] [PubMed]
- Antonopoulos, A.S.; Margaritis, M.; Coutinho, P.; Shirodaria, C.; Psarros, C.; Herdman, L.; Sanna, F.; De Silva, R.; Petrou, M.; Sayeed, R.; et al. Adiponectin as a link between type 2 diabetes and vascular NADPH oxidase activity in the human arterial wall: The regulatory role of perivascular adipose tissue. Diabetes 2015, 64, 2207–2219. [Google Scholar] [CrossRef]
- Gu, P.; Xu, A. Interplay between adipose tissue and blood vessels in obesity and vascular dysfunction. Rev. Endocr. Metab. Disord. 2013, 14, 49–58. [Google Scholar] [CrossRef]
- Vecchione, C.; Maffei, A.; Colella, S.; Aretini, A.; Poulet, R.; Frati, G.; Gentile, M.T.; Fratta, L.; Trimarco, V.; Trimarco, B.; et al. Leptin effect on endothelial nitric oxide is mediated through Akt-endothelial nitric oxide synthase phosphorylation pathway. Diabetes 2002, 51, 168–173. [Google Scholar] [CrossRef]
- Jamroz-Wiśniewska, A.; Gertler, A.; Solomon, G.; Wood, M.E.; Whiteman, M.; Bełtowski, J. Leptin-Induced Endothelium-Dependent Vasorelaxation of Peripheral Arteries in Lean and Obese Rats: Role of Nitric Oxide and Hydrogen Sulfide. PLoS ONE 2014, 9, e86744. [Google Scholar] [CrossRef]
- Momin, A.U.; Melikian, N.; Shah, A.M.; Grieve, D.J.; Wheatcroft, S.B.; John, L.; El Gamel, A.; Desai, J.B.; Nelson, T.; Driver, C.; et al. Leptin is an endothelial-independent vasodilator in humans with coronary artery disease: Evidence for tissue specificity of leptin resistance. Eur. Heart J. 2006, 27, 2294–2299. [Google Scholar] [CrossRef]
- Gálvez-Prieto, B.; Somoza, B.; Gil-Ortega, M.F.; García-Prieto, C.; de las Heras, A.; González, M.; Arribas, S.; Aranguez, I.; Bolbrinker, J.; Kreutz, R.; et al. Anticontractile Effect of Perivascular Adipose Tissue and Leptin are Reduced in Hypertension. Front. Pharmacol. 2012, 3, 103. [Google Scholar] [CrossRef]
- Abu Bakar, H.; Robert Dunn, W.; Daly, C.; Ralevic, V. Sensory innervation of perivascular adipose tissue: A crucial role in artery vasodilatation and leptin release. Cardiovasc. Res. 2017, 113, 962–972. [Google Scholar] [CrossRef]
- Atawia, R.; Bruder-Nascimento, T.; Horimatsu, T.; Li, X.; Kennard, S.; Antonova, G.; Chen, W.; Patel, V.; Weintraub, N.; Fulton, D.; et al. Perivascular adipose tissue (PVAT)-derived leptin improves aortic endothelial function via attenuating endothelial glycolysis in a mouse model of lipodystrophy. FASEB J. 2021, 35. [Google Scholar] [CrossRef]
- Schroeter, M.R.; Eschholz, N.; Herzberg, S.; Jerchel, I.; Leifheit-Nestler, M.; Czepluch, F.S.; Chalikias, G.; Konstantinides, S.; Schäfer, K. Leptin-dependent and leptin-independent paracrine effects of perivascular adipose tissue on neointima formation. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 980–987. [Google Scholar] [CrossRef]
- Atawia, R.T.; Bruder-nascimento, T.B.; Horimatsu, T.; Li, X.; Kennard, S.; Chen, W.; Weintraub, N.L.; Fulton, D.; Kim, H.W.W.; De Chantemele, B.E.J. Perivascular Adipose Tissue Regulates Endothelial Function And Glucose Disposal Via Leptin Control Of The 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase 3/NADPH Oxidase 1 Pathways. Hypertension 2020, 76, AP242. [Google Scholar] [CrossRef]
- Spradley, F.T.; Ho, D.H.; Pollock, J.S. Dahl SS rats demonstrate enhanced aortic perivascular adipose tissue-mediated buffering of vasoconstriction through activation of NOS in the endothelium. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2016, 310, 25. [Google Scholar] [CrossRef]
- Payne, G.A.; Borbouse, L.; Kumar, S.; Neeb, Z.; Alloosh, M.; Sturek, M.; Tune, J.D. Epicardial perivascular adipose-derived leptin exacerbates coronary endothelial dysfunction in metabolic syndrome via a protein kinase C-beta pathway. Arterioscler. Thromb. Vasc. Biol. 2010, 30, 1711–1717. [Google Scholar] [CrossRef] [PubMed]
- Ketonen, J.; Shi, J.; Martonen, E.; Mervaala, E. Periadventitial adipose tissue promotes endothelial dysfunction via oxidative stress in diet-induced obese C57Bl/6 mice. Circul. J. 2010, 74, 1479–1487. [Google Scholar] [CrossRef] [PubMed]
- Aghamohammadzadeh, R.; Greenstein, A.S.; Yadav, R.; Jeziorska, M.; Hama, S.; Soltani, F.; Pemberton, P.W.; Ammori, B.; Malik, R.A.; Soran, H. Effects of bariatric surgery on human small artery function: Evidence for reduction in perivascular adipocyte inflammation, and the restoration of normal anticontractile activity despite persistent obesity. J. Am. Coll. Cardiol. 2013, 62, 128–135. [Google Scholar] [CrossRef]
- Li, H.; Wang, Y.P.; Zhang, L.N.; Tian, G. Perivascular adipose tissue-derived leptin promotes vascular smooth muscle cell phenotypic switching via p38 mitogen-activated protein kinase in metabolic syndrome rats. Exp. Biol. Med. 2014, 239, 954–965. [Google Scholar] [CrossRef]
- Noblet, J.N.; Goodwill, A.G.; Sassoon, D.J.; Kiel, A.M.; Tune, J.D. Leptin augments coronary vasoconstriction and smooth muscle proliferation via a Rho-kinase-dependent pathway. Basic. Res. Cardiol. 2016, 111, 25. [Google Scholar] [CrossRef]
- Berti, L.; Hartwig, S.; Irmler, M.; Rädle, B.; Siegel-Axel, D.; Beckers, J.; Lehr, S.; Al-Hasani, H.; Häring, H.U.; Hrabě de Angelis, M.; et al. Impact of fibroblast growth factor 21 on the secretome of human perivascular preadipocytes and adipocytes: A targeted proteomics approach. Arch. Physiol. Biochem. 2016, 122, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Watanabe-Kominato, K.; Takahashi, Y.; Kojima, M.; Watanabe, R. Adipose Tissue-Derived Omentin-1 Function and Regulation. Compr. Physiol. 2017, 7, 765–781. [Google Scholar] [CrossRef]
- Yang, R.Z.; Lee, M.J.; Hu, H.; Pray, J.; Wu, H.B.; Hansen, B.C.; Shuldiner, A.R.; Fried, S.K.; McLenithan, J.C.; Gong, D.W. Identification of omentin as a novel depot-specific adipokine in human adipose tissue: Possible role in modulating insulin action. Am. J. Physiol. Endocrinol. Metab. 2006, 290, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhao, A.; Xiao, H.; Zhu, Y.; Liu, S.; Zhang, S.; Yang, Z.; Du, L.; Li, X.; Niu, X.; Wang, C.; et al. Omentin-1: A newly discovered warrior against metabolic related diseases. Exp. Opin. Therap. Targets 2022, 26, 275–289. [Google Scholar] [CrossRef] [PubMed]
- Hiramatsu-Ito, M.; Shibata, R.; Ohashi, K.; Uemura, Y.; Kanemura, N.; Kambara, T.; Enomoto, T.; Yuasa, D.; Matsuo, K.; Ito, M.; et al. Omentin attenuates atherosclerotic lesion formation in apolipoprotein E-deficient mice. Cardiovasc. Res. 2016, 110, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Watanabe, R.; Konii, H.; Shirai, R.; Sato, K.; Matsuyama, T.A.; Ishibashi-Ueda, H.; Koba, S.; Kobayashi, Y.; Hirano, T.; et al. Counteractive effects of omentin-1 against atherogenesis. Cardiovasc. Res. 2016, 110, 118–128. [Google Scholar] [CrossRef]
- Du, Y.; Ji, Q.; Cai, L.; Huang, F.; Lai, Y.; Liu, Y.; Yu, J.; Han, B.; Zhu, E.; Zhang, J.; et al. Association between omentin-1 expression in human epicardial adipose tissue and coronary atherosclerosis. Cardiovasc. Diabetol. 2016, 15, 90. [Google Scholar] [CrossRef]
- Kowalówka, A.; Machnik, G.; Deja, M.; Okopień, B.; Gąsior, Z.; Haberka, M. Perivascular adipose tissue from the internal mammary artery in patients with severe coronary artery atherosclerosis. Kardiol. Pol. 2020, 78, 1215–1220. [Google Scholar] [CrossRef]
- Yamawaki, H.; Tsubaki, N.; Mukohda, M.; Okada, M.; Hara, Y. Omentin, a novel adipokine, induces vasodilation in rat isolated blood vessels. Biochem. Biophys. Res. Commun. 2010, 393, 668–672. [Google Scholar] [CrossRef]
- Okumura, H.; Nagaya, N.; Enomoto, M.; Nakagawa, E.; Oya, H.; Kangawa, K. Vasodilatory effect of ghrelin, an endogenous peptide from the stomach. J. Cardiovasc. Pharmacol. 2002, 39, 779–783. [Google Scholar] [CrossRef]
- Schinzari, F.; Veneziani, A.; Mores, N.; Barini, A.; Daniele, N.D.; Cardillo, C.; Tesauro, M. Beneficial Effects of Apelin on Vascular Function in Patients With Central Obesity. Hypertension 2017, 69, 942–949. [Google Scholar] [CrossRef]
- Ashton, D.; Hieble, P.; Gout, B.; Aiyar, N. Vasodilatory Effect of Adrenomedullin in Mouse Aorta. Pharmacology 2000, 61, 101–105. [Google Scholar] [CrossRef]
- Heaton, J.; Lin, B.; Chang, J.K.; Steinberg, S.; Hyman, A.; Lippton, H. Pulmonary vasodilation to adrenomedullin: A novel peptide in humans. Am. J. Physiol.-Heart Circul. Physiol. 1995, 268, H2211–H2215. [Google Scholar] [CrossRef] [PubMed]
- Demirel, S.; Sahinturk, S.; Isbil, N.; Ozyener, F. Physiological role of K+ channels in irisin-induced vasodilation in rat thoracic aorta. Peptides 2022, 147, 170685. [Google Scholar] [CrossRef]
- Ye, L.; Xu, M.; Hu, M.; Zhang, H.; Tan, X.; Li, Q.; Shen, B.; Huang, J. TRPV4 is involved in irisin-induced endothelium-dependent vasodilation. Biochem. Biophys. Res. Commun. 2018, 495, 41–45. [Google Scholar] [CrossRef]
- Kameshima, S.; Sakamoto, Y.; Okada, M.; Yamawaki, H. Vaspin prevents elevation of blood pressure through inhibition of peripheral vascular remodelling in spontaneously hypertensive rats. Acta Physiol. 2016, 217, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lyu, W.; Yu, M.; Niu, Y. Sulfur dioxide induces vascular relaxation through PI3K/Akt/eNOS and NO/cGMP signaling pathways in rats. Human Exp. Toxicol. 2020, 39, 1108–1117. [Google Scholar] [CrossRef] [PubMed]
- Meng, Z.; Li, J.; Zhang, Q.; Bai, W.; Yang, Z.; Zhao, Y.; Wang, F. Vasodilator effect of gaseous sulfur dioxide and regulation of its level by Ach in rat vascular tissues. Inhal. Toxicol. 2009, 21, 1223–1228. [Google Scholar] [CrossRef]
- Thorup, C.; Jones, C.L.; Gross, S.S.; Moore, L.C.; Goligorsky, M.S. Carbon monoxide induces vasodilation and nitric oxide release but suppresses endothelial NOS. Am. J. Physiol. 1999, 277, F882–F889. [Google Scholar] [CrossRef]
- Kanu, A.; Whitfield, J.; Leffler, C.W. Carbon monoxide contributes to hypotension-induced cerebrovascular vasodilation in piglets. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, 2. [Google Scholar] [CrossRef]
- Saxton, S.N.; Ryding, K.E.; Aldous, R.G.; Withers, S.B.; Ohanian, J.; Heagerty, A.M. Role of Sympathetic Nerves and Adipocyte Catecholamine Uptake in the Vasorelaxant Function of Perivascular Adipose Tissue. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 880–891. [Google Scholar] [CrossRef]
- Ayala-Lopez, N.; Jackson, W.F.; Burnett, R.; Wilson, J.N.; Thompson, J.M.; Watts, S.W. Organic cation transporter 3 contributes to norepinephrine uptake into perivascular adipose tissue. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, 2. [Google Scholar] [CrossRef]
- Li, X.; Ballantyne, L.L.; Yu, Y.; Funk, C.D. Perivascular adipose tissue-derived extracellular vesicle miR-221-3p mediates vascular remodeling. FASEB J. 2019, 33, 12704–12722. [Google Scholar] [CrossRef] [PubMed]
- Fujii, J.; Homma, T.; Osaki, T. Superoxide Radicals in the Execution of Cell Death. Antioxidants 2022, 11, 501. [Google Scholar] [CrossRef] [PubMed]
- Staiculescu, M.C.; Foote, C.; Meininger, G.A.; Martinez-Lemus, L.A. The role of reactive oxygen species in microvascular remodeling. Int. J. Mol. Sci. 2014, 15, 23792–23835. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.J.; Takemori, K.; Su, L.Y.; An, W.S.; Lu, C.; Sharma, A.M.; Lee, R.M. Perivascular adipose tissue promotes vasoconstriction: The role of superoxide anion. Cardiovasc. Res. 2006, 71, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Zhang, Y.; Shao, M.; Mu, Q.; Jiao, X.; Hou, N.; Sun, X. C1q/TNF-related protein 9 improves the anti-contractile effects of perivascular adipose tissue via the AMPK-eNOS pathway in diet-induced obese mice. Clin. Exp. Pharmacol. Physiol. 2018, 45, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Gonzaga, N.A.; Awata, W.M.C.; Ficher, S.P.; Assis, V.O.; Alves, J.V.; Tostes, R.C.; Tirapelli, C.R. Melatonin reverses the loss of the anticontractile effect of perivascular adipose tissue in obese rats. J. Pineal. Res. 2021, 70, e12710. [Google Scholar] [CrossRef]
- Gil-Ortega, M.; Condezo-Hoyos, L.; García-Prieto, C.F.; Arribas, S.M.; González, M.C.; Aranguez, I.; Ruiz-Gayo, M.; Somoza, B.; Fernandez-Alfonso, M.S. Imbalance between pro and anti-oxidant mechanisms in perivascular adipose tissue aggravates long-term high-fat diet-derived endothelial dysfunction. PLoS ONE 2014, 9, e95312. [Google Scholar] [CrossRef]
- Hou, N.; Du, G.; Han, F.; Zhang, J.; Jiao, X.; Sun, X. Irisin Regulates Heme Oxygenase-1/Adiponectin Axis in Perivascular Adipose Tissue and Improves Endothelial Dysfunction in Diet-Induced Obese Mice. Cell. Physiol. Biochem. 2017, 42, 603–614. [Google Scholar] [CrossRef]
- da Costa, R.M.; Silva, J.F.d; Alves, J.V.; Dias, T.B.; Rassi, D.M.; Garcia, L.V.; Lobato, N.D.S.; Tostes, R.C. Increased O-GlcNAcylation of Endothelial Nitric Oxide Synthase Compromises the Anti-contractile Properties of Perivascular Adipose Tissue in Metabolic Syndrome. Front. Physiol. 2018, 9, 341. [Google Scholar] [CrossRef] [PubMed]
- Fleenor, B.S.; Eng, J.S.; Sindler, A.L.; Pham, B.T.; Kloor, J.D.; Seals, D.R. Superoxide signaling in perivascular adipose tissue promotes age-related artery stiffness. Aging Cell 2014, 13, 576–578. [Google Scholar] [CrossRef]
- Knock, G.A.; Snetkov, V.A.; Shaifta, Y.; Connolly, M.; Drndarski, S.; Noah, A.; Pourmahram, G.E.; Becker, S.; Aaronson, P.I.; Ward, J.P. Superoxide constricts rat pulmonary arteries via Rho-kinase-mediated Ca(2+) sensitization. Free Radic. Biol. Med. 2009, 46, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Bharadwaj, L.A.; Prasad, K. Mechanism of superoxide anion-induced modulation of vascular tone. Int. J. Angiol. 2002, 11, 23–29. [Google Scholar] [CrossRef]
- Laight, D.W.; Kaw, A.V.; Carrier, M.J.; Änggård, E.E. Interaction between superoxide anion and nitric oxide in the regulation of vascular endothelial function. Br. J. Pharmacol. 1998, 124, 238–244. [Google Scholar] [CrossRef]
- Xia, N.; Horke, S.; Habermeier, A.; Closs, E.I.; Reifenberg, G.; Gericke, A.; Mikhed, Y.; Münzel, T.; Daiber, A.; Förstermann, U.; et al. Uncoupling of endothelial nitric oxide synthase in perivascular adipose tissue of diet-induced obese mice. Arterioscl. Thromb. Vasc. Biol. 2016, 36, 78–85. [Google Scholar] [CrossRef]
- Marchesi, C.; Ebrahimian, T.; Angulo, O.; Paradis, P.; Schiffrin, E.L. Endothelial nitric oxide synthase uncoupling and perivascular adipose oxidative stress and inflammation contribute to vascular dysfunction in a rodent model of metabolic syndrome. Hypertension 2009, 54, 1384–1392. [Google Scholar] [CrossRef]
- Mehta, P.K.; Griendling, K.K. Angiotensin II cell signaling: Physiological and pathological effects in the cardiovascular system. Am. J. Physiol. Cell Physiol. 2007, 292, 26. [Google Scholar] [CrossRef] [PubMed]
- Balakumar, P.; Alqahtani, A.; Khan, N.A.; Alqahtani, T.A.T.; Jagadeesh, G. The physiologic and physiopathologic roles of perivascular adipose tissue and its interactions with blood vessels and the renin-angiotensin system. Pharmacol. Res. 2021, 173, 105890. [Google Scholar] [CrossRef]
- Lu, C.; Su, L.Y.; Lee, R.M.; Gao, Y.J. Mechanisms for perivascular adipose tissue-mediated potentiation of vascular contraction to perivascular neuronal stimulation: The role of adipocyte-derived angiotensin II. Eur. J. Pharmacol. 2010, 634, 107–112. [Google Scholar] [CrossRef]
- Lu, C.; Su, L.Y.; Lee, R.M.; Gao, Y.J. Superoxide anion mediates angiotensin II-induced potentiation of contractile response to sympathetic stimulation. Eur. J. Pharmacol. 2008, 589, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Zou, L.; Wang, W.; Liu, S.; Zhao, X.; Lyv, Y.; Du, C.; Su, X.; Geng, B.; Xu, G. Spontaneous hypertension occurs with adipose tissue dysfunction in perilipin-1 null mice. Biochim. Biophys. Acta 2016, 2, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Mu, W.-J.; Song, Y.-J.; Yang, L.-J.; Qian, S.-W.; Yang, Q.-Q.; Liu, Y.; Tang, Q.Q.; Tang, Y. Bone morphogenetic protein 4 in perivascular adipose tissue ameliorates hypertension through regulation of angiotensinogen. Front. Cardiovasc. Med. 2022, 9, 1038176. [Google Scholar] [CrossRef] [PubMed]
- Kassam, S.I.; Lu, C.; Buckley, N.; Gao, Y.J.; Lee, R.M. Modulation of thiopental-induced vascular relaxation and contraction by perivascular adipose tissue and endothelium. Br. J. Anaesth. 2012, 109, 177–184. [Google Scholar] [CrossRef]
- Huang, F.; Lezama, M.A.; Ontiveros, J.A.; Bravo, G.; Villafaña, S.; del-Rio-Navarro, B.E.; Hong, E. Effect of losartan on vascular function in fructose-fed rats: The role of perivascular adipose tissue. Clin. Exp. Hypertens. 2010, 32, 98–104. [Google Scholar] [CrossRef]
- Rosei, C.A.; Withers, S.B.; Belcaid, L.; De Ciuceis, C.; Rizzoni, D.; Heagerty, A.M. Blockade of the renin-angiotensin system in small arteries and anticontractile function of perivascular adipose tissue. J. Hypertens. 2015, 33, 1039–1045. [Google Scholar] [CrossRef]
- Fontes, M.T.; Paula, S.M.; Lino, C.A.; Senger, N.; Couto, G.K.; Barreto-Chaves, M.L.M.; Mill, J.G.; Rossoni, L.V. Renin-angiotensin system overactivation in perivascular adipose tissue contributes to vascular dysfunction in heart failure. Clin. Sci. 2020, 134, 3195–3211. [Google Scholar] [CrossRef]
- Wang, Z.; Lu, H.; Garcia-Barrio, M.; Guo, Y.; Zhang, J.; Chen, Y.E.; Chang, L. RNA sequencing reveals perivascular adipose tissue plasticity in response to angiotensin II. Pharmacol. Res. 2022, 178, 16. [Google Scholar] [CrossRef]
- Kagota, S.; Maruyama-Fumoto, K.; Shimari, M.; McGuire, J.J.; Shinozuka, K. Angiotensin II Type 1 Receptor Antagonist Azilsartan Restores Vascular Reactivity Through a Perivascular Adipose Tissue-Independent Mechanism in Rats with Metabolic Syndrome. Cardiovasc. Drugs Ther. 2019, 33, 501–509. [Google Scholar] [CrossRef]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef]
- Wang, M.; FitzGerald, G.A. Cardiovascular Biology of Microsomal Prostaglandin E Synthase-1. Trends Cardiovasc. Med. 2010, 20, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.R.; Fredette, N.C.; Barton, M.; Prossnitz, E.R. Regulation of vascular smooth muscle tone by adipose-derived contracting factor. PLoS ONE 2013, 8, e79245. [Google Scholar] [CrossRef] [PubMed]
- dos Reis Costa, D.E.F.; de Araújo, N.F.; Nóbrega, N.R.C.; de Assis Rabelo Ribeiro, N.; de Oliveira, A.C.C.; dos Santos Aggum Capettini, L.; Ferreira, A.V.M.; Bonaventura, D. Contribution of RAS, ROS and COX-1-derived prostanoids to the contractile profile of perivascular adipose tissue in cafeteria diet-induced obesity. Life Sci. 2022, 309, 120994. [Google Scholar] [CrossRef]
- Chang, L.; Villacota, L.; Dou, C.; Chen, E.; Zhang, J. Perivascular Adipose Tissue-derived Prostaglandins Constrict Vessel. Hypertension 2012, 60, A662. [Google Scholar] [CrossRef]
- Yang, T.; Du, Y. Distinct roles of central and peripheral prostaglandin E2 and EP subtypes in blood pressure regulation. Am. J. Hypertens. 2012, 25, 1042–1049. [Google Scholar] [CrossRef]
- Jadhav, V.; Jabre, A.; Lin, S.Z.; Lee, T.J. EP1- and EP3-receptors mediate prostaglandin E2-induced constriction of porcine large cerebral arteries. J. Cereb. Blood Flow Metab. 2004, 24, 1305–1316. [Google Scholar] [CrossRef]
- Ahmad, A.A.; Randall, M.D.; Roberts, R.E. Sex differences in the role of phospholipase A(2) -dependent arachidonic acid pathway in the perivascular adipose tissue function in pigs. J. Physiol. 2017, 595, 6623–6634. [Google Scholar] [CrossRef]
- Spradley, F.T. Sympathetic nervous system control of vascular function and blood pressure during pregnancy and preeclampsia. J. Hypertens. 2019, 37, 476–487. [Google Scholar] [CrossRef]
- Sorriento, D.; Santulli, G.; Del Giudice, C.; Anastasio, A.; Trimarco, B.; Iaccarino, G. Endothelial cells are able to synthesize and release catecholamines both in vitro and in vivo. Hypertension 2012, 60, 129–136. [Google Scholar] [CrossRef]
- Ahmad, M.F.; Ferland, D.; Ayala-Lopez, N.; Contreras, G.A.; Darios, E.; Thompson, J.; Ismail, A.; Thelen, K.; Moeser, A.J.; Burnett, R.; et al. Perivascular Adipocytes Store Norepinephrine by Vesicular Transport. Arterioscl. Thromb. Vasc. Biol. 2019, 39, 188–199. [Google Scholar] [CrossRef]
- Ayala-Lopez, N.; Martini, M.; Jackson, W.F.; Darios, E.; Burnett, R.; Seitz, B.; Fink, G.D.; Watts, S.W. Perivascular adipose tissue contains functional catecholamines. Pharmacol. Res. Perspect. 2014, 2, 41. [Google Scholar] [CrossRef]
- Vargovic, P.; Ukropec, J.; Laukova, M.; Cleary, S.; Manz, B.; Pacak, K.; Kvetnansky, R. Adipocytes as a new source of catecholamine production. FEBS Lett. 2011, 585, 2279–2284. [Google Scholar] [CrossRef] [PubMed]
- Stunes, A.K.; Reseland, J.E.; Hauso, O.; Kidd, M.; Tømmerås, K.; Waldum, H.L.; Syversen, U.; Gustafsson, B.I. Adipocytes express a functional system for serotonin synthesis, reuptake and receptor activation. Diabetes Obes. Metab. 2011, 13, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Leite, F.; Ribeiro, L. Adipocytes and macrophages secretomes coregulate catecholamine-synthesizing enzymes. Int. J. Med. Sci. 2021, 18, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.K.; Darios, E.S.; Burnett, R.; Thompson, J.M.; Watts, S.W. Fenfluramine-induced PVAT-dependent contraction depends on norepinephrine and not serotonin. Pharmacol. Res. 2019, 140, 43–49. [Google Scholar] [CrossRef]
- Kwiecien, K.; Brzoza, P.; Bak, M.; Majewski, P.; Skulimowska, I.; Bednarczyk, K.; Cichy, J.; Kwitniewski, M. The methylation status of the chemerin promoter region located from −252 to + 258 bp regulates constitutive but not acute-phase cytokine-inducible chemerin expression levels. Sci. Rep. 2020, 10, 13702. [Google Scholar] [CrossRef]
- Carracedo, M.; Artiach, G.; Witasp, A.; Clària, J.; Carlström, M.; Laguna-Fernandez, A.; Stenvinkel, P.; Bäck, M. The G-protein coupled receptor ChemR23 determines smooth muscle cell phenotypic switching to enhance high phosphate-induced vascular calcification. Cardiovasc. Res. 2019, 115, 1557–1566. [Google Scholar] [CrossRef]
- Watts, S.W.; Dorrance, A.M.; Penfold, M.E.; Rourke, J.L.; Sinal, C.J.; Seitz, B.; Sullivan, T.J.; Charvat, T.T.; Thompson, J.M.; Burnett, R. Chemerin connects fat to arterial contraction. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1320–1328. [Google Scholar] [CrossRef]
- Darios, E.S.; Winner, B.M.; Charvat, T.; Krasinksi, A.; Punna, S.; Watts, S.W. The adipokine chemerin amplifies electrical field-stimulated contraction in the isolated rat superior mesenteric artery. Am. J. Physiol.-Heart Circul. Physiol. 2016, 311, H498–H507. [Google Scholar] [CrossRef]
- Flood, E.D.; Watts, S.W. Endogenous Chemerin from PVAT Amplifies Electrical Field-Stimulated Arterial Contraction: Use of the Chemerin Knockout Rat. Int. J. Mol. Sci. 2020, 21, 6392. [Google Scholar] [CrossRef]
- Kennedy, A.J.; Yang, P.; Read, C.; Kuc, R.E.; Yang, L.; Taylor, E.J.; Taylor, C.W.; Maguire, J.J.; Davenport, A.P. Chemerin Elicits Potent Constrictor Actions via Chemokine-Like Receptor 1 (CMKLR1), not G-Protein-Coupled Receptor 1 (GPR1), in Human and Rat Vasculature. J. Am. Heart Assoc. 2016, 5, 004421. [Google Scholar] [CrossRef]
- Ferland, D.J.; Darios, E.S.; Neubig, R.R.; Sjögren, B.; Truong, N.; Torres, R.; Dexheimer, T.S.; Thompson, J.M.; Watts, S.W. Chemerin-induced arterial contraction is G(i)- and calcium-dependent. Vascul. Pharmacol. 2017, 88, 30–41. [Google Scholar] [CrossRef]
- Omori, A.; Goshima, M.; Kakuda, C.; Kodama, T.; Otani, K.; Okada, M.; Yamawaki, H. Chemerin-9-induced contraction was enhanced through the upregulation of smooth muscle chemokine-like receptor 1 in isolated pulmonary artery of pulmonary arterial hypertensive rats. Pflugers Arch. 2020, 472, 335–342. [Google Scholar] [CrossRef]
- Son, M.; Oh, S.; Lee, H.S.; Chung, D.M.; Jang, J.T.; Jeon, Y.J.; Choi, C.H.; Park, K.Y.; Son, K.H.; Byun, K. Ecklonia Cava Extract Attenuates Endothelial Cell Dysfunction by Modulation of Inflammation and Brown Adipocyte Function in Perivascular Fat Tissue. Nutrients 2019, 11, 2795. [Google Scholar] [CrossRef] [PubMed]
- Weng, C.; Shen, Z.; Li, X.; Jiang, W.; Peng, L.; Yuan, H.; Yang, K.; Wang, J. Effects of chemerin/CMKLR1 in obesity-induced hypertension and potential mechanism. Am. J. Transl. Res. 2017, 9, 3096–3104. [Google Scholar] [PubMed]
- Jamaluddin, M.S.; Weakley, S.M.; Yao, Q.; Chen, C. Resistin: Functional roles and therapeutic considerations for cardiovascular disease. Br. J. Pharmacol. 2012, 165, 622–632. [Google Scholar] [CrossRef]
- Lee, S.; Lee, H.C.; Kwon, Y.W.; Lee, S.E.; Cho, Y.; Kim, J.; Lee, S.; Kim, J.Y.; Lee, J.; Yang, H.M. Adenylyl cyclase-associated protein 1 is a receptor for human resistin and mediates inflammatory actions of human monocytes. Cell. Metab. 2014, 19, 484–497. [Google Scholar] [CrossRef] [PubMed]
- Tarkowski, A.; Bjersing, J.; Shestakov, A.; Bokarewa, M.I. Resistin competes with lipopolysaccharide for binding to toll-like receptor 4. J. Cell. Mol. Med. 2010, 14, 1419–1431. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Jiang, J.; Lü, J.M.; Chai, H.; Wang, X.; Lin, P.H.; Yao, Q. Resistin decreases expression of endothelial nitric oxide synthase through oxidative stress in human coronary artery endothelial cells. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, 30. [Google Scholar] [CrossRef]
- Rachwalik, M.; Obremska, M.; Zyśko, D.; Matusiewicz, M.; Protasiewicz, M.; Jasiński, M. Resistin levels in perivascular adipose tissue and mid-term mortality in patients undergoing coronary artery bypass grafting. Physiol. Res. 2021, 70, 543–550. [Google Scholar] [CrossRef]
- Rachwalik, M.; Obremska, M.; Zyśko, D.; Matusiewicz, M.; Ściborski, K.; Jasiński, M. The concentration of resistin in perivascular adipose tissue after CABG and postoperative atrial fibrillation. BMC Cardiovasc. Dis. 2019, 19, 294. [Google Scholar] [CrossRef]
- Park, S.Y.; Kim, K.H.; Seo, K.W.; Bae, J.U.; Kim, Y.H.; Lee, S.J.; Lee, W.S.; Kim, C.D. Resistin derived from diabetic perivascular adipose tissue up-regulates vascular expression of osteopontin via the AP-1 signalling pathway. J. Pathol. 2014, 232, 87–97. [Google Scholar] [CrossRef]
- Small, H.Y.; McNeilly, S.; Mary, S.; Sheikh, A.M.; Delles, C. Resistin Mediates Sex-Dependent Effects of Perivascular Adipose Tissue on Vascular Function in the Shrsp. Sci. Rep. 2019, 9, 6897. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Huang, L.; Wang, A.; Liu, Y.; Cai, R.; Li, W.; Zhou, M.S. Resistin-Induced Endoplasmic Reticulum Stress Contributes to the Impairment of Insulin Signaling in Endothelium. Front. Pharmacol. 2018, 9, 1226. [Google Scholar] [CrossRef]
- Gentile, M.T.; Vecchione, C.; Marino, G.; Aretini, A.; Di Pardo, A.; Antenucci, G.; Maffei, A.; Cifelli, G.; Iorio, L.; Landolfi, A. Resistin impairs insulin-evoked vasodilation. Diabetes 2008, 57, 577–583. [Google Scholar] [CrossRef]
- Dick, G.M.; Katz, P.S.; Farias, M.; Morris, M.; James, J.; Knudson, J.D.; Tune, J.D. Resistin impairs endothelium-dependent dilation to bradykinin, but not acetylcholine, in the coronary circulation. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, 11. [Google Scholar] [CrossRef]
- Verma, S.; Li, S.H.; Wang, C.H.; Fedak, P.W.; Li, R.K.; Weisel, R.D.; Mickle, D.A. Resistin promotes endothelial cell activation: Further evidence of adipokine-endothelial interaction. Circulation 2003, 108, 736–740. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.Y.; Chao, Y.W.; Tsai, Y.L.; Lien, C.C.; Chang, C.F.; Deng, M.C.; Ho, L.T.; Kwok, C.F.; Juan, C.C. Resistin induces monocyte-endothelial cell adhesion by increasing ICAM-1 and VCAM-1 expression in endothelial cells via p38MAPK-dependent pathway. J. Cell Physiol. 2011, 226, 2181–2188. [Google Scholar] [CrossRef] [PubMed]
- Hirai, H.; Satoh, H.; Kudoh, A.; Watanabe, T. Interaction between resistin and adiponectin in the proliferation of rat vascular smooth muscle cells. Mol. Cell. Endocrinol. 2013, 366, 108–116. [Google Scholar] [CrossRef]
- Calabro, P.; Samudio, I.; Willerson, J.T.; Yeh, E.T.H. Resistin Promotes Smooth Muscle Cell Proliferation Through Activation of Extracellular Signal–Regulated Kinase 1/2 and Phosphatidylinositol 3-Kinase Pathways. Circulation 2004, 110, 3335–3340. [Google Scholar] [CrossRef]
- Saddi-Rosa, P.; Oliveira, C.S.V.; Giuffrida, F.M.A.; Reis, A.F. Visfatin, glucose metabolism and vascular disease: A review of evidence. Diabetol. Metab. Syndr. 2010, 2, 21. [Google Scholar] [CrossRef] [PubMed]
- Dakroub, A.; Nasser, S.A.; Younis, N.; Bhagani, H.; Al-Dhaheri, Y.; Pintus, G.; Eid, A.A.; El-Yazbi, A.F.; Eid, A.H. Visfatin: A Possible Role in Cardiovasculo-Metabolic Disorders. Cells 2020, 9, 2444. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.E.; Onyango, D.J.; Ramanjaneya, M.; Conner, A.C.; Patel, S.T.; Dunmore, S.J.; Randeva, H.S. Visfatin regulates insulin secretion, insulin receptor signalling and mRNA expression of diabetes-related genes in mouse pancreatic beta-cells. J. Mol. Endocrinol. 2010, 44, 171–178. [Google Scholar] [CrossRef]
- Gasparrini, M.; Mazzola, F.; Cuccioloni, M.; Sorci, L.; Audrito, V.; Zamporlini, F.; Fortunato, C.; Amici, A.; Cianci, M.; Deaglio, S.; et al. Molecular insights into the interaction between human nicotinamide phosphoribosyltransferase and Toll-like receptor 4. J. Biol. Chem. 2022, 298, 101669. [Google Scholar] [CrossRef]
- Zheng, L.-Y.; Xu, X.; Wan, R.-H.; Xia, S.; Lu, J.; Huang, Q. Association between serum visfatin levels and atherosclerotic plaque in patients with type 2 diabetes. Diabetol. Metab. Syndr. 2019, 11, 60. [Google Scholar] [CrossRef]
- Stastny, J.; Bienertova-Vasku, J.; Vasku, A. Visfatin and its role in obesity development. Diabetes Metab. Syndr. Clin. Res. Rev. 2012, 6, 120–124. [Google Scholar] [CrossRef]
- Dahl, T.B.; Yndestad, A.; Skjelland, M.; Øie, E.; Dahl, A.; Michelsen, A.; Damås, J.K.; Tunheim, S.H.; Ueland, T.; Smith, C.; et al. Increased Expression of Visfatin in Macrophages of Human Unstable Carotid and Coronary Atherosclerosis. Circulation 2007, 115, 972–980. [Google Scholar] [CrossRef]
- Morgan, S.A.; Bringolf, J.B.; Seidel, E.R. Visfatin expression is elevated in normal human pregnancy. Peptides 2008, 29, 1382–1389. [Google Scholar] [CrossRef]
- Akcabag, E.; Bayram, Z.; Kucukcetin, I.O.; Uzun, G.; Ozdem, S.; Ozdem, S.S. Functional effects of visfatin in isolated rat mesenteric small resistance arteries. Eur. J. Pharmacol. 2021, 908, 17. [Google Scholar] [CrossRef]
- Wang, P.; Xu, T.Y.; Guan, Y.F.; Su, D.F.; Fan, G.R.; Miao, C.Y. Perivascular adipose tissue-derived visfatin is a vascular smooth muscle cell growth factor: Role of nicotinamide mononucleotide. Cardiovasc. Res. 2009, 81, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Yamawaki, H.; Hara, N.; Okada, M.; Hara, Y. Visfatin causes endothelium-dependent relaxation in isolated blood vessels. Biochem. Biophys. Res. Commun. 2009, 383, 503–508. [Google Scholar] [CrossRef]
- Vallejo, S.; Romacho, T.; Angulo, J.; Villalobos, L.A.; Cercas, E.; Leivas, A.; Bermejo, E.; Carraro, R.; Sanchez-Ferrer, C.F.; Peiro, C. Visfatin Impairs Endothelium-Dependent Relaxation in Rat and Human Mesenteric Microvessels through Nicotinamide Phosphoribosyltransferase Activity. PLoS ONE 2011, 6, e27299. [Google Scholar] [CrossRef] [PubMed]
- Song, E.; Fan, P.; Huang, B.; Deng, H.B.; Cheung, B.M.; Félétou, M.; Villeneuve, N.; Xu, A.; Vanhoutte, P.M.; Wang, Y. Deamidated lipocalin-2 induces endothelial dysfunction and hypertension in dietary obese mice. J. Am. Heart Assoc. 2014, 3, 000837. [Google Scholar] [CrossRef] [PubMed]
- Grigoras, A.; Amalinei, C.; Balan, R.A.; Giusca, S.E.; Caruntu, I.D. Perivascular adipose tissue in cardiovascular diseases-an update. Anatol. J. Cardiol. 2019, 22, 219–231. [Google Scholar] [CrossRef]
- Sena, C.M.; Pereira, A.; Fernandes, R.; Letra, L.; Seiça, R.M. Adiponectin improves endothelial function in mesenteric arteries of rats fed a high-fat diet: Role of perivascular adipose tissue. Br. J. Pharmacol. 2017, 174, 3514–3526. [Google Scholar] [CrossRef]
- Han, S.; Yang, C.-L.; Chen, X.; Naes, L.; Cox, B.F.; Westfall, T. Direct evidence for the role of neuropeptide Y in sympathetic nerve stimulation-induced vasoconstriction. Am. J. Physiol.-Heart Circul. Physiol. 1998, 274, H290–H294. [Google Scholar] [CrossRef] [PubMed]
- Singer, K.; Morris, D.L.; Oatmen, K.E.; Wang, T.; DelProposto, J.; Mergian, T.; Cho, K.W.; Lumeng, C.N. Neuropeptide Y is produced by adipose tissue macrophages and regulates obesity-induced inflammation. PLoS ONE 2013, 8, 5. [Google Scholar] [CrossRef]
- Giardina, J.B.; Green, G.M.; Cockrell, K.L.; Granger, J.P.; Khalil, R.A. TNF-alpha enhances contraction and inhibits endothelial NO-cGMP relaxation in systemic vessels of pregnant rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, R130–R143. [Google Scholar] [CrossRef]
- Ma, Y.; Li, L.; Shao, Y.; Bai, X.; Bai, T.; Huang, X. Methotrexate improves perivascular adipose tissue/endothelial dysfunction via activation of AMPK/eNOS pathway. Mol. Med. Rep. 2017, 15, 2353–2359. [Google Scholar] [CrossRef]
- Tang, W.B.; Zhou, Y.Q.; Zhao, T.; Shan, J.L.; Sun, P.; Yang, T.T.; Chang, X.W.; Li, S.; Wang, P.S.; Xie, D.P. Effect of interleukin-6 (IL-6) on the vascular smooth muscle contraction in abdominal aorta of rats with streptozotocin-induced diabetes. Chin. J. Physiol. 2011, 54, 318–323. [Google Scholar]
- Dorrance, A.M. Interleukin 1-beta (IL-1beta) enhances contractile responses in endothelium-denuded aorta from hypertensive, but not normotensive, rats. Vascul. Pharmacol. 2007, 47, 160–165. [Google Scholar] [CrossRef]
- Blomkalns, A.L.; Gavrila, D.; Thomas, M.; Neltner, B.S.; Blanco, V.M.; Benjamin, S.B.; McCormick, M.L.; Stoll, L.L.; Denning, G.M.; Collins, S.P.; et al. CD14 directs adventitial macrophage precursor recruitment: Role in early abdominal aortic aneurysm formation. J. Am. Heart Assoc. 2013, 2, e000065. [Google Scholar] [CrossRef] [PubMed]
- Gruzdeva, O.V.; Dyleva, Y.A.; Belik, E.V.; Sinitsky, M.Y.; Stasev, A.N.; Kokov, A.N.; Brel, N.K.; Krivkina, E.O.; Bychkova, E.E.; Tarasov, R.S.; et al. Relationship between Epicardial and Coronary Adipose Tissue and the Expression of Adiponectin, Leptin, and Interleukin 6 in Patients with Coronary Artery Disease. J. Pers. Med. 2022, 12, 129. [Google Scholar] [CrossRef]
- Zhang, Z.B.; Ruan, C.C.; Lin, J.R.; Xu, L.; Chen, X.H.; Du, Y.N.; Fu, M.X.; Kong, L.R.; Zhu, D.L.; Gao, P.J. Perivascular Adipose Tissue-Derived PDGF-D Contributes to Aortic Aneurysm Formation During Obesity. Diabetes 2018, 67, 1549–1560. [Google Scholar] [CrossRef] [PubMed]
- Manka, D.; Chatterjee, T.K.; Stoll, L.L.; Basford, J.E.; Konaniah, E.S.; Srinivasan, R.; Bogdanov, V.Y.; Tang, Y.; Blomkalns, A.L.; Hui, D.Y.; et al. Transplanted perivascular adipose tissue accelerates injury-induced neointimal hyperplasia: Role of monocyte chemoattractant protein-1. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1723–1730. [Google Scholar] [CrossRef]
- Ruan, C.C.; Ma, Y.; Ge, Q.; Li, Y.; Zhu, L.M.; Zhang, Y.; Kong, L.R.; Wu, Q.H.; Li, F.; Cheng, L.; et al. Complement-mediated inhibition of adiponectin regulates perivascular inflammation and vascular injury in hypertension. FASEB J. 2017, 31, 1120–1129. [Google Scholar] [CrossRef] [PubMed]
- Lyngsø, K.S.; Assersen, K.; Dalgaard, E.G.; Skott, O.; Jensen, B.L.; Hansen, P.B. Does Aldosterone Play a Significant Role for Regulation of Vascular Tone? J. Cardiovasc. Pharmacol. 2016, 68, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Emont, M.P.; Jacobs, C.; Essene, A.L.; Pant, D.; Tenen, D.; Colleluori, G.; Vincenzo, A.D.; Jørgensen, A.M.; Dashti, H.; Stefek, A.; et al. A single-cell atlas of human and mouse white adipose tissue. Nature 2022, 603, 926–933. [Google Scholar] [CrossRef]
- Lee, R.M.; Lu, C.; Su, L.-Y.; Werstuck, G.; Gao, Y.-J. Effects of hyperglycemia on the modulation of vascular function by perivascular adipose tissue. J. Hypertens. 2009, 27, 118–131. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, A.; Bibi, A.; Valoti, M.; Fusi, F. Perivascular Adipose Tissue and Vascular Smooth Muscle Tone: Friends or Foes? Cells 2023, 12, 1196. https://doi.org/10.3390/cells12081196
Ahmed A, Bibi A, Valoti M, Fusi F. Perivascular Adipose Tissue and Vascular Smooth Muscle Tone: Friends or Foes? Cells. 2023; 12(8):1196. https://doi.org/10.3390/cells12081196
Chicago/Turabian StyleAhmed, Amer, Aasia Bibi, Massimo Valoti, and Fabio Fusi. 2023. "Perivascular Adipose Tissue and Vascular Smooth Muscle Tone: Friends or Foes?" Cells 12, no. 8: 1196. https://doi.org/10.3390/cells12081196