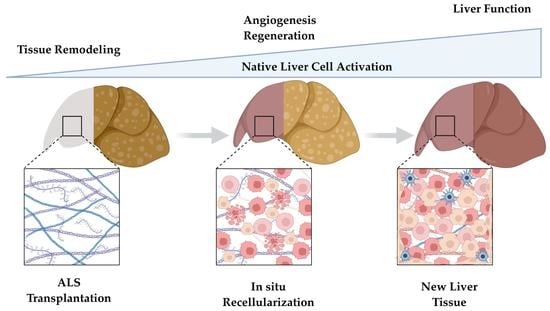
1. Introduction
Liver decellularization produces a non-immunogenic, three-dimensional biological scaffold consisting of an intricate network of structural and non-structural macromolecules from the extracellular matrix (ECM) [
1]. The liver ECM plays an important role in tissue maintenance, homeostasis, and regeneration [
2]. In addition, the ECM orchestrates key cellular behaviors that impact cell support, shape, movement, adhesion, communication, function, and cellular response [
1,
2].
Accordingly, acellular liver scaffolds (ALS) produced by decellularization have been successfully applied to distinct regenerative aims, and beneficial results were achieved when these scaffolds were transplanted in vivo [
3,
4,
5]. While several studies have focused on the recellularization steps before transplantation, ALS transplantation itself has only been partially explored. Although total orthotopic transplantation of ALS would not support the metabolic demands of patients after transplantation, other alternatives can be explored to find new therapeutic tools in the fields of hepatology and regenerative medicine [
6]. For example, ALS could be applied after partial hepatectomy to increase the limit of liver resection [
6,
7] or the scaffold could be used in partial orthotopic transplantation as a way to create a bioartificial liver that ensures metabolic functions while the patient’s own liver regenerates [
6,
8].
The first report of ALS orthotopic transplantation was by Zhang and colleagues in 2015, which provided a proof of concept that decellularized matrices can be transplanted into recipient mice [
9]. Subsequently, Naeem and colleagues performed ALS transplantation after lobectomy and confirmed in situ recellularization when the ALS was transplanted into recipient rats [
10]. Shimoda and colleagues evaluated the potential contribution of ALS transplantation to hepatic regeneration after hepatectomy in recipient pigs [
7]. Taken together, these studies showed that ALS could be recellularized by healthier cells and promoted liver regeneration after hepatectomy when transplanted into mice [
9], rats [
10], and pigs [
7]. Despite these significant advances, the ALS recellularization process and the body’s ability to repopulate the scaffold was investigated only in healthy recipient animals. Therefore, as a next step, it is necessary to investigate cellular recruitment, cell engraftment, post-transplantation angiogenesis and potential therapeutic improvement after ALS transplantation in liver disease models.
To achieve this, we combined in vivo and ex vivo techniques, tissue engineering tools, and a therapeutic approach that involved the granulocyte colony stimulating factor (G-CSF), which is known to stimulate bone marrow cell mobilization. To our knowledge, this study is the first to explore ALS transplantation in G-CSF-pretreated cirrhotic animals. Our strategy is attractive because G-CSF has already been employed in clinical practice for bone marrow cell mobilization, and cumulative evidence has shown that G-CSF exerts beneficial effects in liver disease [
11,
12,
13]. Therefore, the purpose of this study was to investigate whether ALS could be recellularized by healthy cells when transplanted into cirrhotic rats pre-treated with G-CSF, and to elucidate the regenerative events. Accordingly, we also aimed to investigate whether ALS promotes the formation of functional liver tissue in situ and improves the status of the cirrhotic liver after transplantation. In this work, we investigated the hypothesis that ALS could be transplanted directly into a diseased recipient animal, relying entirely on the scaffold’s ability to promote recruitment, adhesion, and repopulation of resident cells to regenerate endogenous liver tissue. We observed that, even under liver disease conditions, the body can act to promote ALS recellularization.
2. Material and Methods
2.1. Experimental Design
Female Wistar rats were subjected to liver cirrhosis induction by intraperitoneal injections of carbon tetrachloride (CCl
4) in association with 5% ethanol in drinking water. At the same time, the donor animals underwent surgical excision of the liver. Each donor liver was decellularized to generate an ALS. Five consecutive days before transplantation, the recipient animals (cirrhotic animals) received subcutaneous injections of granulocyte colony-stimulating factor (G-CSF). Each ALS was then transplanted into a cirrhotic recipient, and ultrasonographic, microcirculation, biochemical, and histological analyses were performed 7, 15, and 30 days after transplantation. The experimental design is illustrated in
Figure 1.
2.2. Animals
This study was approved (097/20) and followed the animal care guidelines of the Animal Ethics Committee of the Health Science Center of the Federal University of Rio de Janeiro, Brazil. Forty-one female Wistar rats, ranging in age from 8 to 14 weeks, were used. The animals were randomly distributed into the following four groups: liver donor rats (n = 9), control rats (n = 3), cirrhotic transplanted rats (Tx; n = 20), and cirrhotic and partial hepatectomized rats (PH; n = 9). Animals were kept on a 12 h light/dark cycle, 25 °C ambient temperature, and 55 ± 5% humidity. During the experiment, the rats were fed standard pellets and water ad libitum and their body weight was monitored weekly. Anesthesia was provided by inhalation of 3–4% isoflurane (Isoforine®, Cristália, São Paulo, Brazil) and maintained by inhalation of 1–2% isoflurane. All rats were given oxygen at a dose of 0.3–0.5 L/min.
2.3. Liver Cirrhosis Induction
Female Wistar rats received CCl4 1 mL/kg diluted in olive oil (1:1) intraperitoneally, three times a week every other day over 8 weeks. In addition, the animals had access to drinking water containing 5% ethanol (v/v) “Caninha da roça” (Indústria de Bebidas Paris LTDA, Rio de Janeiro, Brazil) ad libitum. The control group received 1 mL/kg of olive oil intraperitoneally, three times a week, for 8 weeks.
2.4. Granulocyte Colony-Stimulating Factor Treatment
Cirrhotic rats received G-CSF (100 μg/kg/day) (Filgrastine, Blau Farmacêutica, Belo Horizonte, Brazil) subcutaneously for five consecutive days before partial hepatectomy or ALS transplantation to stimulate bone marrow cell mobilization. Blood samples (500 µL) were collected in EDTA K2 microtubes (Vacuplast, São Paulo, Brazil) and subjected to hemogram analysis in a hematology analyzer on days 0, 3, and 5 after G-CSF treatment. At the same time points, hematology sections were stained with Panoptic Fast Staining (Laborclin, Paraná, Brazil) using 10 μL of blood. The images were obtained by using a Pannoramic MIDI II microscope and scanner (3DHISTECH Ltd., Budapest, Hungary).
2.5. Biochemical Analysis
Blood samples (500 μL) were collected by cardiac puncture in a clot activator gel microtube (Vacuplast, São Paulo, Brazil) before and after cirrhosis induction. In addition, blood samples were also collected before and after 7, 15 and 30 days post-transplantation. Then, the blood samples were centrifugated at 1400 g for 10 min and the serum was stored at −20 °C. Serum levels of albumin (ALB), alanine aminotransferase (ALT), lactate dehydrogenase (LDH), alkaline phosphatase (AP), urea, total bilirubin (TB), and aspartate aminotransferase (AST) were determined with a semi-automatic biochemical analyzer Bio 200 (Bioplus, Rio de Janeiro, Brazil), using an ALB detection kit (Ref. 19, Labtest, Minas Gerais, Brazil), ALT kit (Ref. 108, Labtest, Minas Gerais, Brazil), LDH kit (Ref. 138-1/50, Labtest, Minas Gerais, Brazil), AP kit (Ref. 79-4, Labtest, Minas Gerais, Brazil), Urea kit (Ref. 104-4/50, Labtest, Minas Gerais, Brazil), TB kit (Ref. 94-1, Labtest, Minas Gerais, Brazil) and AST detection kit (Ref. 109, Labtest, Minas Gerais, Brazil).
2.6. Liver Procurement
The animals (n = 9) were heparinized with 100 UI of heparin (Hemofol®, Cristália, São Paulo, Brazil) 15 min before the surgical procedures. A transverse abdominal incision followed by laparotomy was performed. Then, the portal vein (PV) was isolated and cannulated with the aid of a 24-gauge catheter (Angiocatch®, BD, São Paulo, Brazil). After PV cannulation, 5 mL of Custodiol (Contatti Medical, Porto Alegre, Brazil) solution was perfused into the liver. Subsequently, the liver was carefully excised and placed in a 60 mm sterile Petri dish.
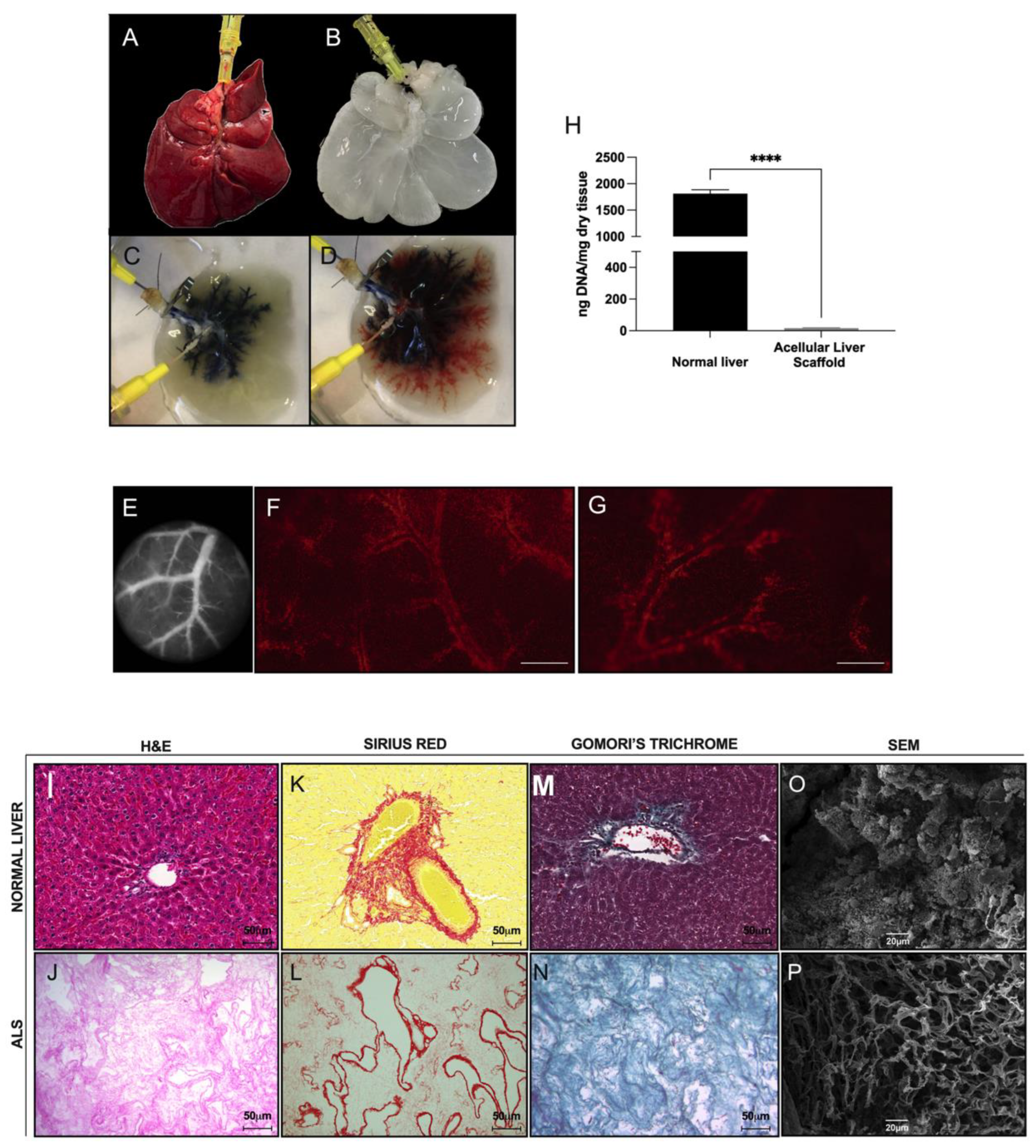
2.7. Liver Decellularization
Harvested rat livers (
n = 9) were decellularized as previously described [
14] using a peristaltic pump (Masterflex Cole Parmer L/S, Model 7522-20) at a speed of 3 mL/min. Briefly, the liver was subjected to continuous perfusion with water (2 h) and 1% (
v/
v) Triton X-100 solution (Sigma-Aldrich, Saint Louis, MO, USA) (2 h) through the PV. Subsequently, a 1% sodium dodecyl sulfate (SDS) (Synth, São Paulo, Brazil) solution was perfused for 18–24 h. After washing with distilled water (overnight), the ALS was perfused with a solution containing 1% amphotericin b, 1% penicillin, and streptomycin (Sigma-Aldrich, Saint Louis, MO, USA) for 1 h.
2.8. Vascular Tree Evaluation
Macroscopic analysis of the ALS vasculature was performed by perfusion of toluidine blue and phenol red. Initially, the portal vein and bile duct were cannulated with the aid of a 24G catheter (Angiocatch®, BD, São Paulo, Brazil). Then, 1 mL of toluidine blue was infused through the portal vein and 1 mL of phenol red through the bile duct. The microscopic analysis was performed by using an Olympus BX150WI microscope (Center Valley, PA, USA).
2.9. Histological Analysis
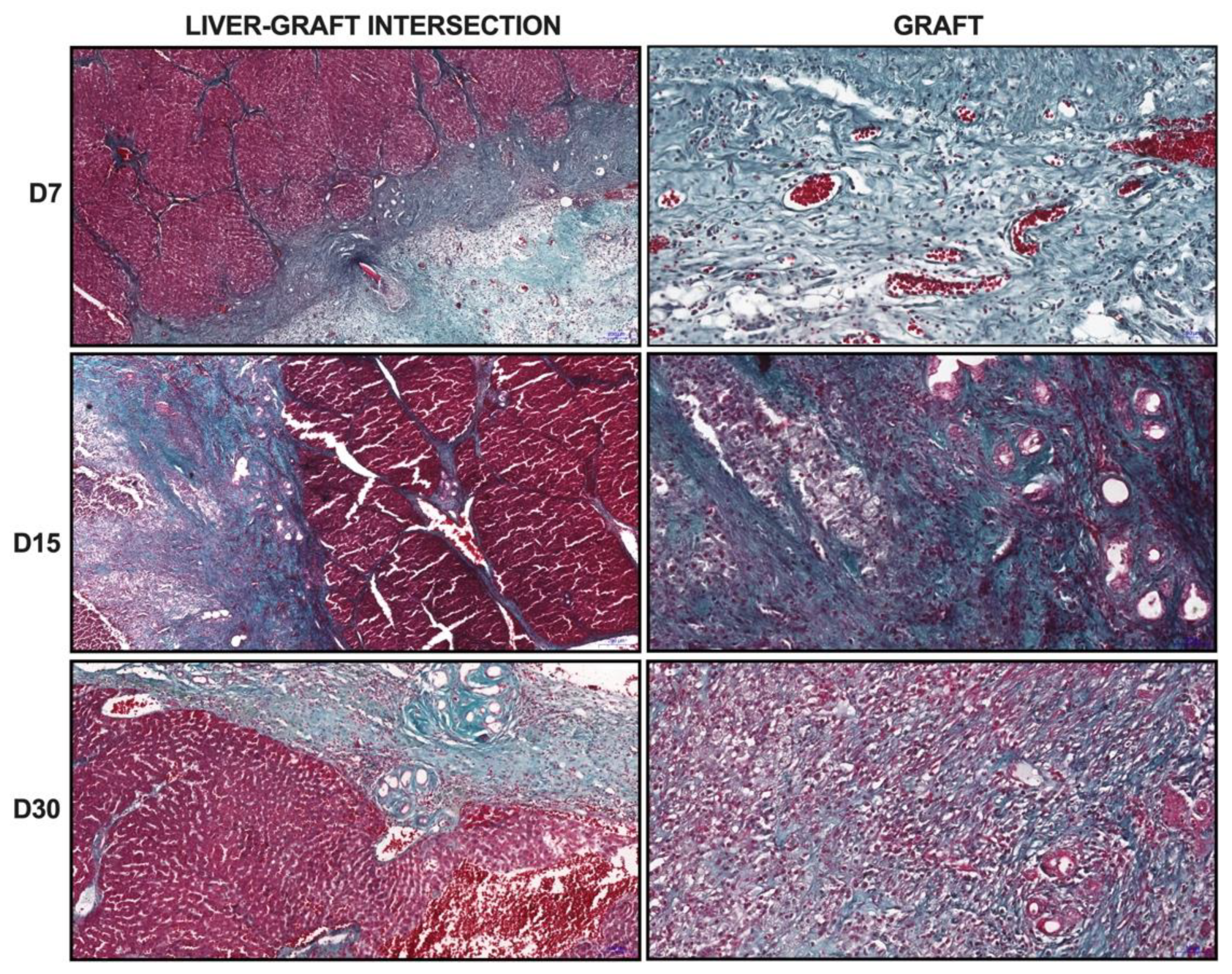
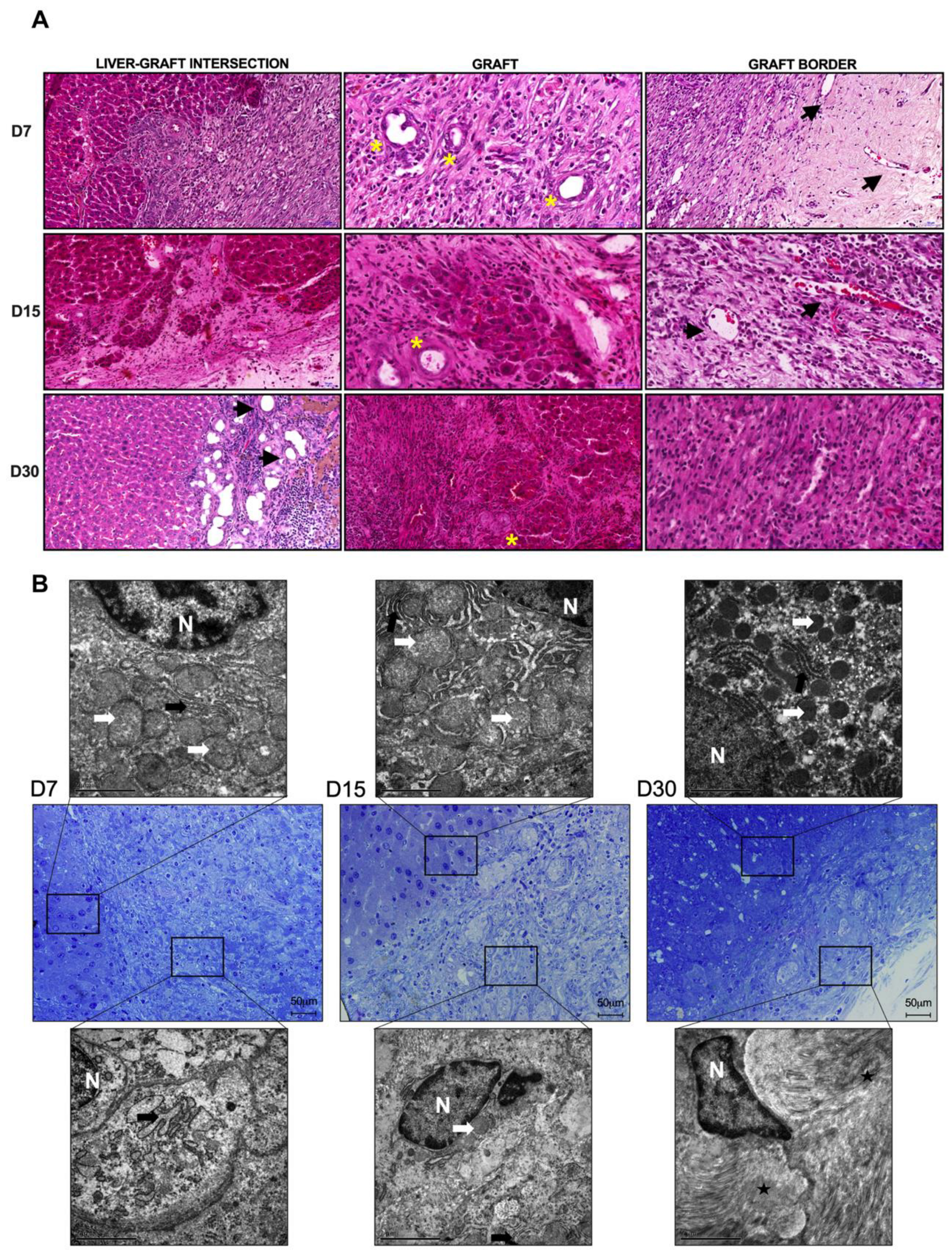
Biopsies from normal livers and ALSs were fixed in formalin (4%) for 48 h. Then, the biopsies were paraffin-embedded and sectioned (5 μm) for the following analyses (n = 3 each): to investigate the morphology, the sections were stained with hematoxylin and eosin (H&E) (Merck, São Paulo, Brazil); to investigate the deposition of the ECM, tissue sections were stained with Gomori’s trichrome, Sirius red, and reticulin. In addition, to investigate glycogen storage, tissue sections were stained with periodic acid–Schiff (PAS) (Merck, São Paulo, Brazil). The images were obtained by using a Pannoramic MIDI II microscope and scanner (3DHISTECH Ltd., Hungary) or an Olympus BX53 polarizing microscope (Olympus Corporation, Japan).
2.10. Immunohistochemical and TUNEL Analyses
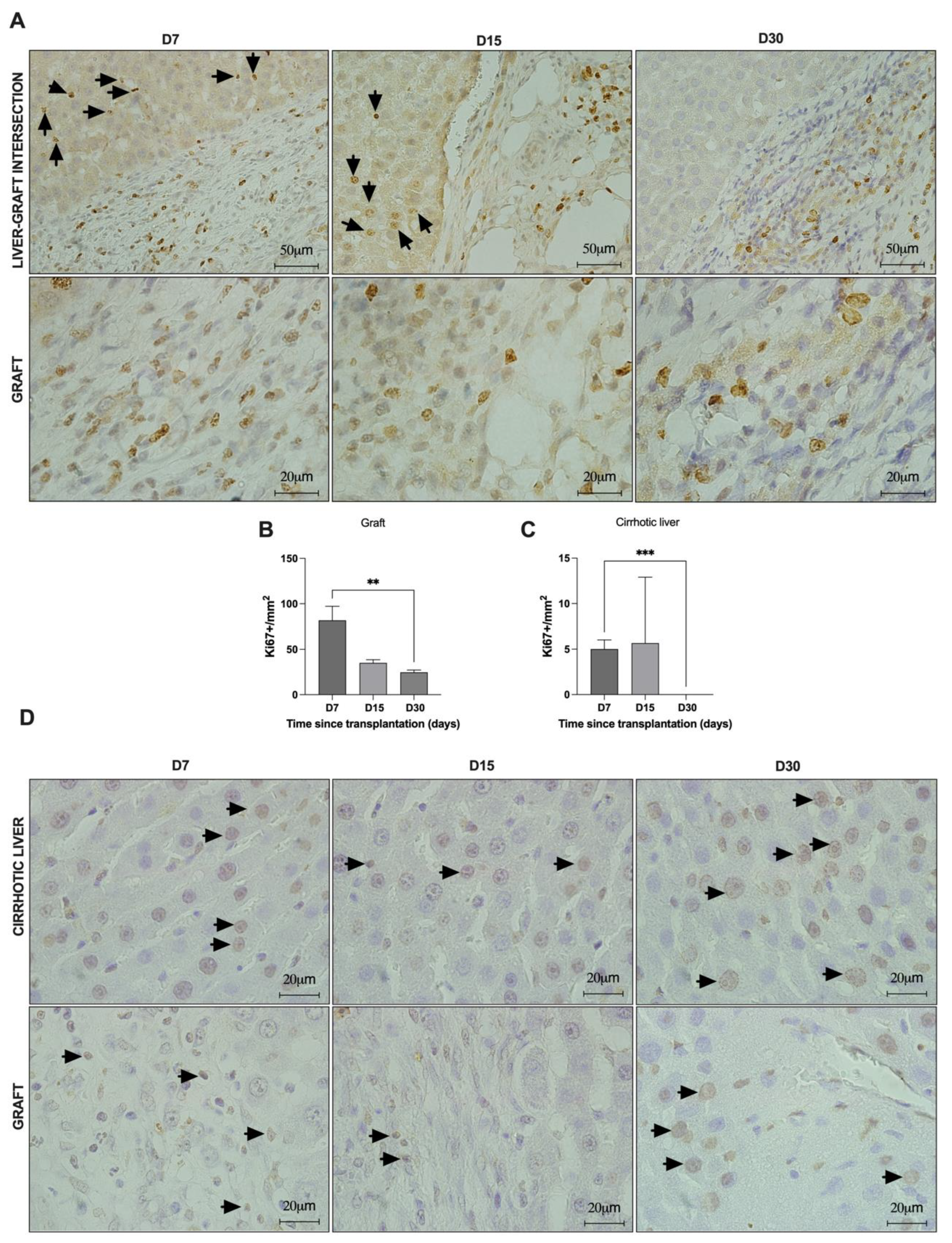
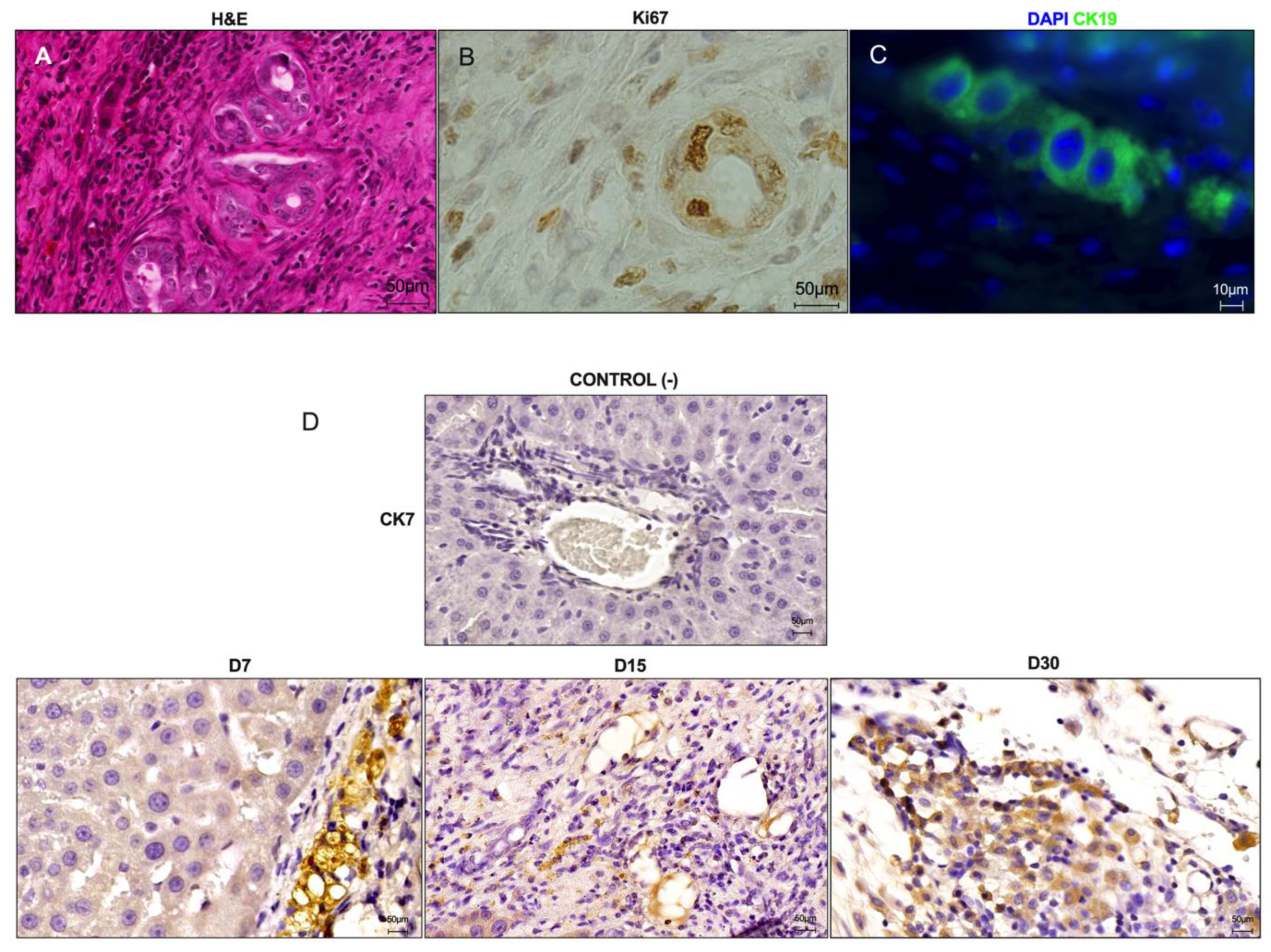
For the immunohistochemical analyses, following deparaffinization and rehydration, the sections were exposed to hydrogen peroxide (3%) diluted in phosphate-buffered saline (PBS) (30 min). Excess peroxide was removed with PBS + Tween 0.2% (2 min). Antigen retrieval was achieved by immersing the slides in sodium citrate buffer (pH 6.0) in a steam cooking appliance at 96 °C (Fun Kitchen, São Paulo, Brazil) (30 min). Then, the slices were immersed in ice for 30 min, followed by two wash steps with PBS + Tween 0.2% (2 min each). The tissue sections were incubated in bovine serum albumin (3%) in PBS for 1 h to block non-specific antibody binding. The slides were then incubated with primary antibodies for Ki67 (1:100; M3064; Spring Bioscience, Pleasanton, CA, USA) and anti-Cytokeratin 7 (CK7) (1:100; M3524, Spring Bioscience, Pleasanton, CA, USA) overnight at 4 °C. Tissue sections with the absence of primary antibodies were used as a control. The next day, the slides were washed with PBS + Tween 0.2% (3 min) and then incubated with a biotin-conjugated secondary antibody (Vectastain universal quick HRP Kit, Vector, Newark, CA, USA) for 10 min. After incubation with streptavidin (Vectastain universal quick HRP Kit, Vector, USA) for 5 min, the reaction was revealed with 3,3-diaminobenzidine (DAB) (SPD-060- Spring Bioscience, Pleasanton, CA, USA) for 2 min and stopped with water. Finally, the sections were stained with hematoxylin for 1 min. The images were obtained by using a Pannoramic MIDI II microscope and scanner (3DHISTECH Ltd., Hungary) or an Olympus BX53 microscope (Olympus Corporation, Japan). TUNEL analysis was used for apoptotic nuclei detection, using the ApopTag® In Situ Peroxidase Detection Kit (Merck Millipore, Saint Louis, MO, USA), according to the manufacturer’s recommendations. For quantification of Ki67, the area of each region (graft/ALS and cirrhotic liver) of immunostained tissue sections was measured using the free-drawing tool of Image J software (National Institutes of Health, Bethesda, MD, USA). Five representative images (100× objective) derived from three experimental tissue sections (n = 3 each) were used for quantification analysis. The positive nuclei were quantified using CountThings software. The quantifications were conducted by a blinded examiner.
2.11. Immunofluorescence Analysis
For immunofluorescence analysis, paraffin-embedded tissue sections were subjected to deparaffinization and rehydration steps. Then, antigen retrieval was achieved by immersing the slides in sodium citrate buffer (pH 6.0) in a steam cooking appliance at 96 °C (Fun Kitchen, São Paulo, Brazil) (30 min). Then, the slides were immersed in ice for 30 min, followed by two wash steps with PBS + Tween 0.2% (2 min each). Autofluorescence block was achieved by ammonium chloride 0.5 mM. After that, the tissue sections were washed with PBS three times. The tissue sections were incubated in bovine serum albumin (3%) in PBS for 1 h to block non-specific antibody binding. Tissue slices were then incubated with primary antibodies for anti-Cytokeratin 19 (CK19) (1:100; Ab15463, Abcam, Boston, MA, USA), anti F4/80 (1:100; Ab6640, Abcam, Boston, MA, USA) and anti CD11b (1:100; ab8878, Abcam, Boston, MA, USA) overnight, followed by washing and secondary incubation in secondary antibodies diluted in PBS/BSA 3% (Alexa Fluor 488 anti-rat, 1:500; A11006, Life technologies; Alexa Fluor 488 anti-mouse, 1:500, A11017, Invitrogen; Alexa Fluor 488 anti-sheep, 1:500, A11015, Invitrogen and Alexa Fluor 594 anti-rabbit, 1:500, A11012, Molecular Probes). The tissue sections were washed and counterstained with 4′,6-diamidino-2-phenylindole (DAPI; 1:500). Images were acquired by using a Zeiss Apotome confocal microscope (Zeiss, Germany).
2.12. Scanning and Transmission Electron Microscopy (SEM and TEM) Analysis
Normal livers or the ALS sections were subjected to washing steps (3x) with 0.2 M PBS (pH 7.2) (LGC Biotecnologia, Rio de Janeiro, Brazil) and fixed in 0.2 M sodium cacodylate buffer (Electron Microscopy Science, USA) containing 2.5% glutaraldehyde (Sigma-Aldrich, USA) at 4 °C for 24 h prior to electron microscopic observation. To remove the glutaraldehyde, the specimens were washed (3 times) in sodium cacodylate buffer (pH 7.2), and then they were dehydrated through an ascending alcohol series (30%, 50%, 70%, 90%, and 100%). Subsequently, the specimens were dried in a critical point dryer (CPD2; Tousimis, Cambridge, MA, USA) and coated with gold in an ion-sputtering apparatus (Sputter Coater 108; Cressington, Watford, UK). The fragments were observed using a scanning electron microscope (1450 VP, LEO Electron Microscopy Ltd., Clifton Road, UK). In addition, semi-thin tissue sections (1 μm) were prepared and stained with toluidine blue and then images were obtained by using an Olympus BX53 microscope (Olympus Corporation, Japan). The acquired images were qualitatively analyzed. For TEM analysis, the recipient cirrhotic liver and graft tissues were processed and then quantitatively analyzed using a JEM1011 (Jeol, Akishima, Tokyo, Japan) microscope. The cirrhotic liver and graft specimens were washed in PBS and fixed for 1 h in a solution containing 2.5% glutaraldehyde in 0.1 M sodium cacodylate buffer (pH 7.2) plus 3.5% sucrose. Then, the samples were washed for 10 min in the same buffer. This washing step was repeated 3 times. The tissue was fixed for 1 h in a 1% osmium tetroxide (OsO4) solution in 0.1 M sodium cacodylate buffer (pH 7.2) plus 3.5% sucrose, dehydrated in an acetone series (30%, 50%, 70%, 90% and 100%) and embedded in Poly/Bed(r) 812 resin (Ted Pella Inc., Redding, CA, USA). After polymerization, ultrathin sections were obtained and contrasted with uranyl acetate–lead citrate for ultrastructural observation.
2.13. DNA Content Analysis
DNA was isolated from 25 mg (dry tissue) of control liver, acellular liver scaffold and transplanted liver tissue (n = 3) and detected by DNeasy® Blood & Tissue Kit (Qiagen, Hilden, Germany). Then, the samples were read at Nanophotometer Pearl® (IMPLEN, München, Germany).
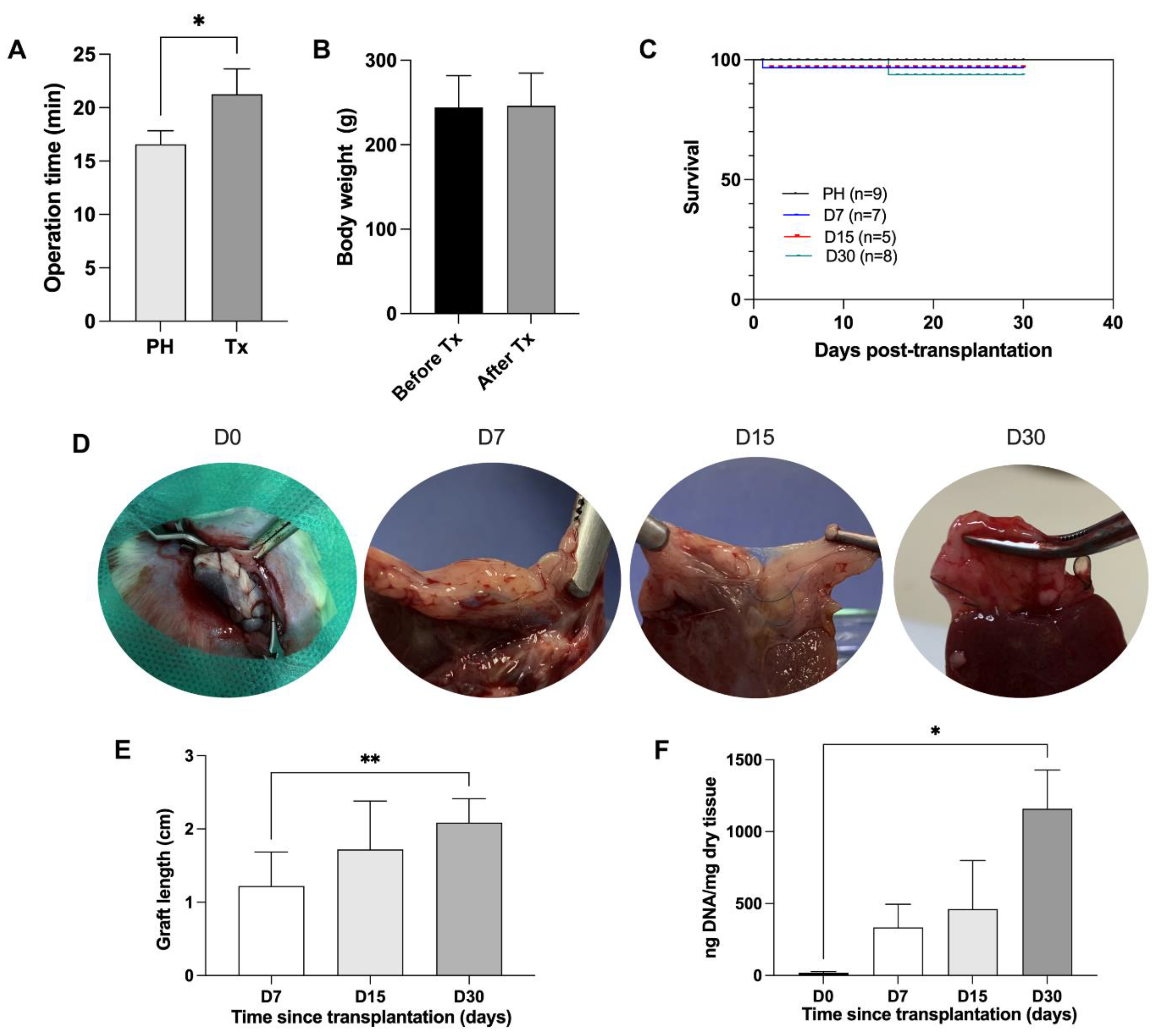
2.14. ALS Orthotopic Transplantation
The ALS was first perfused with Custodiol (5 mL) and 50 μg/mL hepatocyte growth factor (HGF, Sigma-Aldrich, USA) and then orthotopically transplanted into a cirrhotic recipient Wistar rat, as previously described [
12]. The recipient rat previously received granulocyte colony-stimulating factor G-CSF (100 μg/kg/day) (Filgrastine, Blau Farmacêutica, Belo Horizonte, Brazil) subcutaneously for five consecutive days before ALS transplantation to stimulate bone marrow cell mobilization. Blood samples (500 µL) were collected in EDTA K2 microtubes (Vacuplast, Brazil) and subjected to hemogram analysis in a hematology analyzer on days 0, 3, and 5 after G-CSF treatment. At the same time points, hematology sections were stained with Panoptic Fast Staining (Laborclin, Paraná, Brazil) using 10 μL of blood. The images were obtained by using a Pannoramic MIDI II microscope and scanner (3DHISTECH Ltd., Hungary). To perform orthotopic transplantation, recipient animals were subjected to partial median lobe hepatectomy (10%) and subsequently ALS transplantation, after continuous suture with 6–0 silk suture. After transplantation, the animal received 100 μL of tramadol for three days. Only partial hepatectomized rats (
n = 9) were used as a control group. The hepatectomized and transplanted animals were euthanized 7, 15 and 30 days after transplantation. After euthanasia, blood samples were collected, the graft length was measured by a pachymeter, and then tissue sections derived from the cirrhotic liver and the graft were submitted for histological analysis.
2.15. Ultrasound Analysis
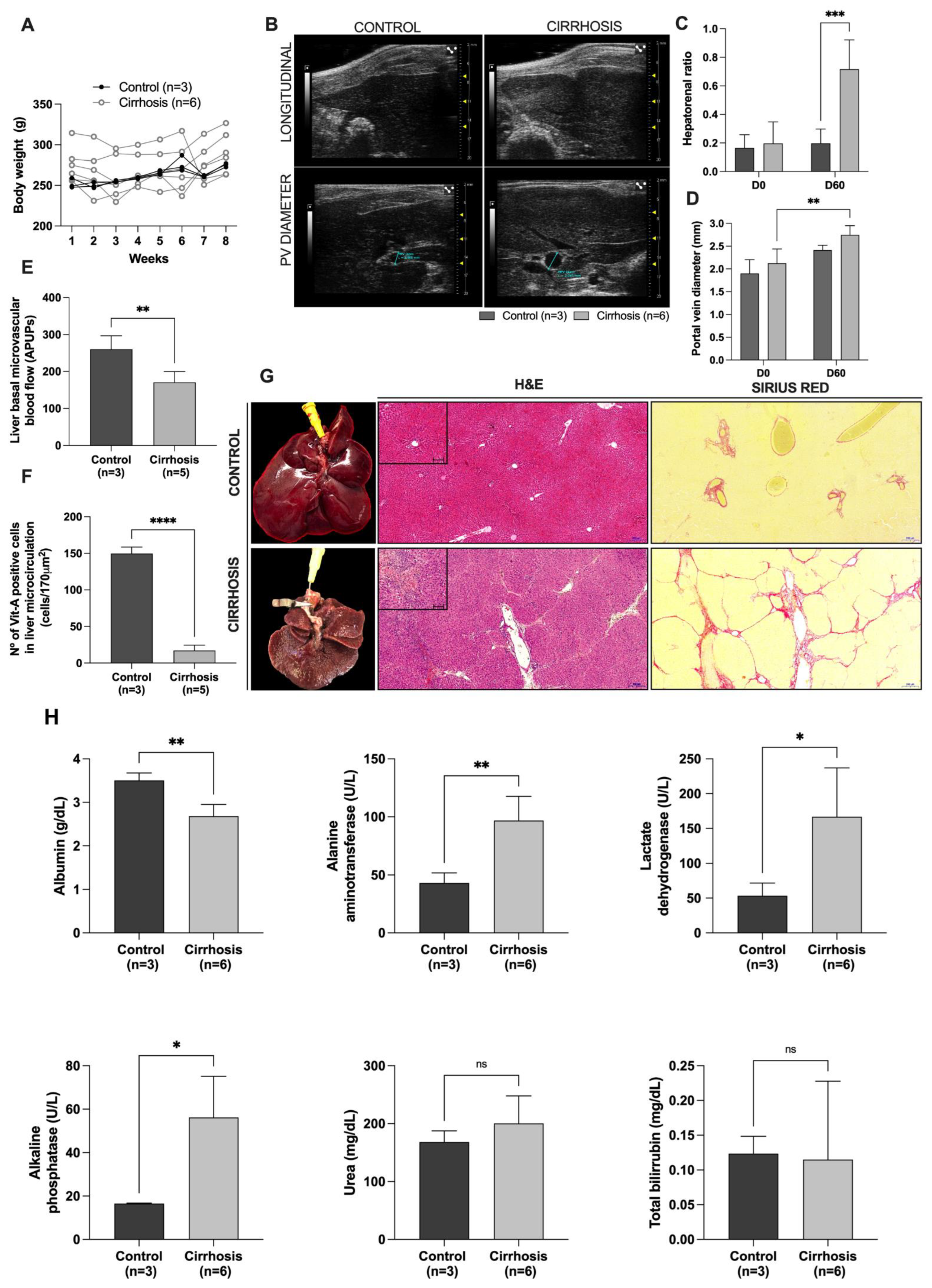
The animals were anesthetized, subjected to abdominal hair shaving, and then examined in the supine position using a Vevo 2100 with a transducer with the frequency 13–24 MHz (MS250; Visual Sonics, Toronto, ON, Canada). The liver and kidneys were evaluated using transverse and longitudinal scans. The evaluated parameters were the portal vein diameter and blood flow velocity. To assess the organ status, liver aspects, echogenicity and the echographic relationship between the liver and renal cortex were analyzed. Ultrasound analyses were performed before and 8 weeks after cirrhosis induction and at 7, 15, and 30 days after transplantation or partial hepatectomy.
2.16. Microcirculation Analysis
For intravital microscopy of the transplanted ALS, normal and cirrhotic Wistar rats were anesthetized by intraperitoneal (i.p.) injection of ketamine (100 mg/kg) and xylazine (10 mg/kg). To exteriorize the liver, midline and left subcostal incisions were performed. The hepatic ligaments were dissected, and the median liver lobe was then exteriorized, placed on a glass disk, and covered with a glass slide for microcirculation analysis. Using a 10× ocular and 10× objective (Olympus BX150WI; Center Valley, PA, USA), images were displayed on a television monitor and recorded by a digital video recorder (DP73; Olympus, MA, USA) for off-line analysis with the Cellsens standard 1.9 software program (Olympus, MA, USA). Hepatic microvascular blood flow was measured using a Laser Speckle Contrast Imaging apparatus (Pericam PSI system, Perimed, Sweden), which provides a microcirculatory perfusion index proportional to the concentration and mean velocity of red blood cells used to assess microvascular blood flow in real time. To perform this analysis, the animals were held on a stable surface and placed under a laser light system with image contrast at a wavelength of 785 nm for continuous measurement of tissue blood perfusion in real time. The distance between the scan head and the cirrhotic liver and ALS surface was approximately 10 cm. Relative liver blood flow in all animal groups was expressed in arbitrary perfusion units (APUs). In cirrhotic livers and grafts, the number of vitamin A-positive hepatic stellate cells (HSCs) was determined as the number of fluorescent cells derived from vitamin A autofluorescence.
2.17. Statistical Analysis
GraphPad Prism 9 (GraphPad Software, La Jolla, CA, USA) was used to conduct statistical analyses. Descriptive data are presented as means ± standard deviations (SD). Comparisons between the groups were performed using a paired or unpaired Student’s t-test, or one-way ANOVA followed by Tukey’s test or two-way ANOVA followed by Šídák’s multiple comparisons test. A value of p < 0.05 was used as a significance threshold.
4. Discussion
In this study, we focused on applying acellular liver scaffolds obtained through decellularization to investigate ECM-based acellular and non-immunogenic material as a template for the in vivo recellularization of healthy cells after transplantation into cirrhotic rats. Here, we investigated the following open questions: (a) whether the body can recellularize the ALS after in vivo transplantation; (b) whether healthy cells can be recruited; (c) whether the transplanted scaffold can be affected by the cirrhotic liver that received it, either becoming cirrhotic or remaining a healthy template for healthy cell growth; and (d) whether the cirrhotic liver changed after ALS transplantation.
To address these questions, our first set of experiments focused on an efficient method for inducing liver cirrhosis in rats. Using a well-established rodent model of liver cirrhosis based on CCl4 administration in association with ethanol, we found that cirrhotic animals developed an inflammatory process that increased hepatic stellate cell activation and promoted the deposition of fibrous ECM compounds. This increase in ECM deposition and consequent fibrous scar formation leads to vascular alterations in cirrhotic livers, resulting in decreased blood flow flux and an increased portal vein diameter, liver echogenicity, and hepatorenal ratio. In addition to the histological and ultrasonographic alterations, our cirrhosis model also affected the biochemical parameters, resembling human cirrhosis. Subsequently, we produced the ALS to perform transplantation in cirrhotic rats.
Decellularization is an efficient method for generating cell-free tissues [
15]. In this study, we applied continuous perfusion of a chemical detergent to eliminate the cellular components of normal rat livers. Our protocol provided acellular liver tissue with a preserved three-dimensional native liver shape, vascular tree, and ECM components from the native liver. Our ALS was employed in a strategy for in vivo organ engineering using partial orthotopic liver transplantation [
16] to investigate and harness the innate capability of the recipient body to promote cell recruitment and full recellularization, resembling new liver tissue. These aspects were similarly explored by Naeem and colleagues [
7] in a study where the ALS produced by sodium lauryl ether sulfate perfusion was transplanted into Sprague Dawley rats, and by Shimoda and colleagues [
10] to investigate the contribution of transplanted ALSs in regeneration after hepatectomy in mini pigs. In our study, to advance closer to clinical application, we addressed these questions using a liver cirrhosis model. To our knowledge, this is the first study to address the endogenous recruitment of liver cells into cirrhotic recipient rats with transplanted ALSs. Here, we observed that the ALS was recellularized 7, 15, and 30 days post-transplantation. This recellularization was time-dependent and cell migration and recruitment occurred gradually. While cell engraftment was more restricted to the intersection area at 7 days, a greater number of cells was present in the scaffold at 15 days and they were distributed more widely throughout the scaffold, achieving complete recellularization and repopulation of the scaffold edges within 30 days post-transplantation. This rapid recellularization capability could benefit patients who undergo hepatectomy.
The great potential of acellular scaffolds derived from decellularized organs was first demonstrated by Taylor and colleagues in 2018. In vivo organ engineering was achieved by heterotopic transplantation of acellular heart scaffolds into pigs and bovine recipients. They observed that acellular heart scaffold transplantation promoted endothelial cell adhesion to the vessel lumen, in addition to cardiomyocyte engraftment and tissue formation within the graft. In this regard, they harnessed the body’s own recuperative powers to achieve recellularization, one of the greatest hurdles in tissue engineering [
17].
In our study, the transplanted scaffolds were completely recellularized by different cell types. We observed intense leukocyte engraftment, especially at the early time points. We first asked whether this could have been stimulated by the G-CSF treatment that preceded ALS transplantation; however, this cell mobilization was also observed when the transplant was performed in recipient animals that did not receive G-CSF [
7,
10]. These observations led us to anticipate that a granulocyte-specific component may orchestrate ALS recellularization. Our findings agree with those previously reported by Naeem in healthy rats [
7] and Shimoda in healthy mini pigs [
10]. Therefore, a granulocyte-specific component also orchestrated the ALS recellularization in healthy and cirrhotic recipients. In addition to granulocyte-component-mediated recellularization in healthy and liver disease contexts, leukocyte engraftment was also observed in other tissues, such as when an acellular diaphragmatic scaffold was produced and transplanted into recipient C57BL/6 mice [
3] and when an acellular uterus scaffold was transplanted into Sprague Dawley recipient rats [
4]. Although acellular scaffolds are considered to be immune-privileged, it is widely known that acellular scaffold transplantation is characterized by immediate mononuclear cell recruitment and engraftment [
18,
19]. However, cumulative evidence has shown that the presence of growth factors and cytokines within the acellular scaffold is responsible for the rapid shift of this immune response into a pro-regenerative response where inflammatory cells, including monocytes and liver-resident macrophages, restore internal communication with fibroblasts, epithelial, endothelial, and stem or tissue progenitor cells [
20,
21,
22,
23]. Taken together, this pro-regenerative response is amplified by cellular stimulation and recruitment [
3,
24]. It is also worth noting that immune response activation is one of the most important key markers for promoting tissue regeneration [
25,
26].
Despite leukocyte engraftment, other cell types such as fibroblasts, Kupffer cells, and hepatocytes migrated within the graft, showing that the ALS was a good niche for cell repopulation and proliferation of different types of cells at all the analyzed time points. Although engraftment of hepatocytes has been observed, extended follow-up analysis must be performed to determine if the number of these cells increases after transplantation. In addition, we observed the engraftment of hepatic stellate cells. In our study, stellate cells were useful for demonstrating the tissue remodeling that ALS undergoes after transplantation. Interestingly, the number of activated stellate cells was increased at 15 days post-transplantation, suggesting a pro-regenerative phase. In parallel, the number of activated stellate cells was decreased at 30 days post-transplantation, confirming the anti-inflammatory effect of transplanted ALS.
In addition to detecting the presence of these cells, we also observed cholangiocytes in ALS. Similarly, Naeem and Shimoda observed the engraftment of cholangiocytes in transplanted ALS. These cells were organized into duct structures that assembled the bile ducts into the transplanted grafts. Importantly, we observed intense bile duct proliferation in the grafts after transplantation. This finding indicates that a bile duct activation pathway can compensate for ALS recellularization after transplantation in cirrhotic rats. The biliary system is known to be involved in liver regeneration and repair, especially after partial hepatectomy [
27,
28]. Likewise, biliary-derived cells can expand and differentiate into hepatocytes. This intense bile duct proliferation might also contribute to ALS recellularization after transplantation through a reparative complex pathway called the ductular reaction [
29,
30,
31]. This supportive evidence agreed with our histological and immunohistological findings, which showed that bile duct proliferation was more intense 7 days post-transplantation, but decreased at 15 and 30 days. Especially at 30 days, we found that many CK7- and CK19-positive cells were distributed throughout the parenchyma of the ALS and no longer in ductular structures. Therefore, our data suggest a correlation of the ductular reaction with a positive outcome of ALS transplantation in recipient cirrhotic rats in early time points after transplantation. Our results showed that this event was important to drive recellularization by liver parenchyma cells, which may have arisen from these proliferating structures. Furthermore, cumulative evidence has shown that CK19 is also a marker of stem/progenitor cells, suggesting the presence of stem and progenitor cells in ALS after transplantation [
27,
28,
29,
30,
31].
In vivo implantation of the ALS gave rise to changes in the characteristics of the recipient cirrhotic livers. These changes resulted from orchestrated events and processes involved in tissue remodeling, angiogenesis, and the immune response promoted by liver injury. To date, little is known about the origin of the cells after acellular scaffold transplantation. Our findings demonstrated proliferative cells in cirrhotic recipient livers 7 and 15 days after ALS transplantation, suggesting that resident hepatocytes proliferate to ensure ALS recellularization at early time points after transplantation. In addition to proliferative events, we also observed that apoptotic events were present in the transplanted graft and in the cirrhotic recipient livers, suggesting that both cell proliferation and cell death were involved in tissue connection and remodeling. These observations highlight that, in addition to the immune response and bile duct proliferative events, the ALS was also recellularized by resident cells that arose from proliferative hepatocytes in the cirrhotic livers. The pro-regenerative response activated by ALS transplantation also contributes to the attenuation of liver cirrhosis. For example, the ultrasonographic parameters improved, in agreement with the histological observations. Moreover, the biochemical parameters also indicated recovery in liver function and attenuation of liver injury markers. In contrast, cirrhotic animals subjected to partial hepatectomy were not able to restore their biochemical and ultrasonographic parameters.
Our findings showed that at all the analyzed time points, the transplanted ALS ensured a pro-regenerative microenvironment, attracted vessels from the host cirrhotic tissue, and promoted stem and progenitor cell recruitment to the ALS after transplantation. These events were beneficial not only for the ALS, but also for promoting tissue remodeling in cirrhotic livers. We observed well-distributed blood vessel-like structures in the transplanted ALS at all the analyzed time points. In some tissue sections, we also observed red blood cells in the vessels, suggesting the presence of blood flux. In agreement with our histological findings, we observed that the transplanted ALS was able to support blood flow at all analyzed time points. The angiogenic potential was improved at the time of transplantation, reaching a higher level of blood flow circulation at 30 days post-transplantation. These angiogenic events effectively facilitated the migration and repopulation of resident cirrhotic liver cells. Certainly, blood flow microcirculation was important to promote nutrient and oxygen supply to the transplanted ALS, allowing the formation of new liver tissue. Interestingly, the improvement of blood flow microcirculation in the recipient cirrhotic livers was also observed after ALS transplantation. To our knowledge, this is the first study to confirm blood flow circulation into the ALS after transplantation in recipient cirrhotic rats.
To resemble functional liver tissue, the transplanted ALS should undergo remodeling to ensure its tissue mechanical properties; therefore, the production of structural ECM proteins is crucial. The liver contains important ECM constituents, such as fibrillar collagens (types I and III) [
6,
14]. In this study, we observed that cells in the graft produced neo-ECMs (collagen I and reticular fibers) after ALS transplantation. This finding is consistent with those reported by Naeem and colleagues [
7]. Hepatic stellate cells, fibroblasts, and immune cells, such as macrophages and neutrophils are the major cells involved in neo-ECM deposition after ALS transplantation. We also confirmed that the transplanted ALS had the capability of in vivo remodeling, regeneration, and growth. These findings were also observed in the recipient cirrhotic livers after ALS transplantation. More importantly, we confirmed that the transplanted ALS was not affected by the cirrhotic livers and remained a template for healthy cell growth. Some studies have already shown that healthy scaffolds provide a template for the growth and development of healthy cells. Ogiso and colleagues showed that healthy ALSs can sustain fetal hepatocytes and cholangiocytes after in vitro recellularization [
32]. Acun and colleagues observed that healthy ALSs also enhanced the differentiation of induced pluripotent stem cells into hepatocyte-like cells and significantly increased the expression of mature hepatocyte markers [
33]. On the other hand, early studies indicated that acellular cirrhotic scaffolds derived from cirrhotic rats accelerated the epithelial–mesenchymal transition, phenotype, proliferation, and drug resistance of hepatocarcinoma cells in vitro [
34]. In addition, human cirrhotic liver acellular scaffolds also promoted the Smad-dependent TGF-β1 epithelial–mesenchymal transition when HepG2 cells were recellularized in vitro [
35]. Thus, we did not know a priori whether cirrhotic cells derived from cirrhotic livers could repopulate ALS and assemble healthy liver tissue. Histological, biochemical, and ultrasonographic analyses confirmed that the ALS was recellularized by healthy cells to generate tissue resembling healthy liver tissue. Our findings demonstrate that the ALS obtained by decellularization is a potent scaffold for cell proliferation and differentiation induction that is capable of providing an appropriate environment for liver tissue assembly and formation.
In light of the encouraging results obtained with rats in this study, we speculate that ALSs can be readily extended to large animal models of liver disease so that in the near future, this alternative will reach patients. Our data provide proof of concept that ALSs can be transplanted into patients with liver disease, as an alternative to the rapid replacement of liver mass and as metabolic support until the patient’s own liver recovers.



 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}












