
Comprehensive Analysis of BrHMPs Reveals Potential Roles in Abiotic Stress Tolerance and Pollen–Stigma Interaction in Brassica rapa
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification and Physicochemical Characterization of BrHMPs
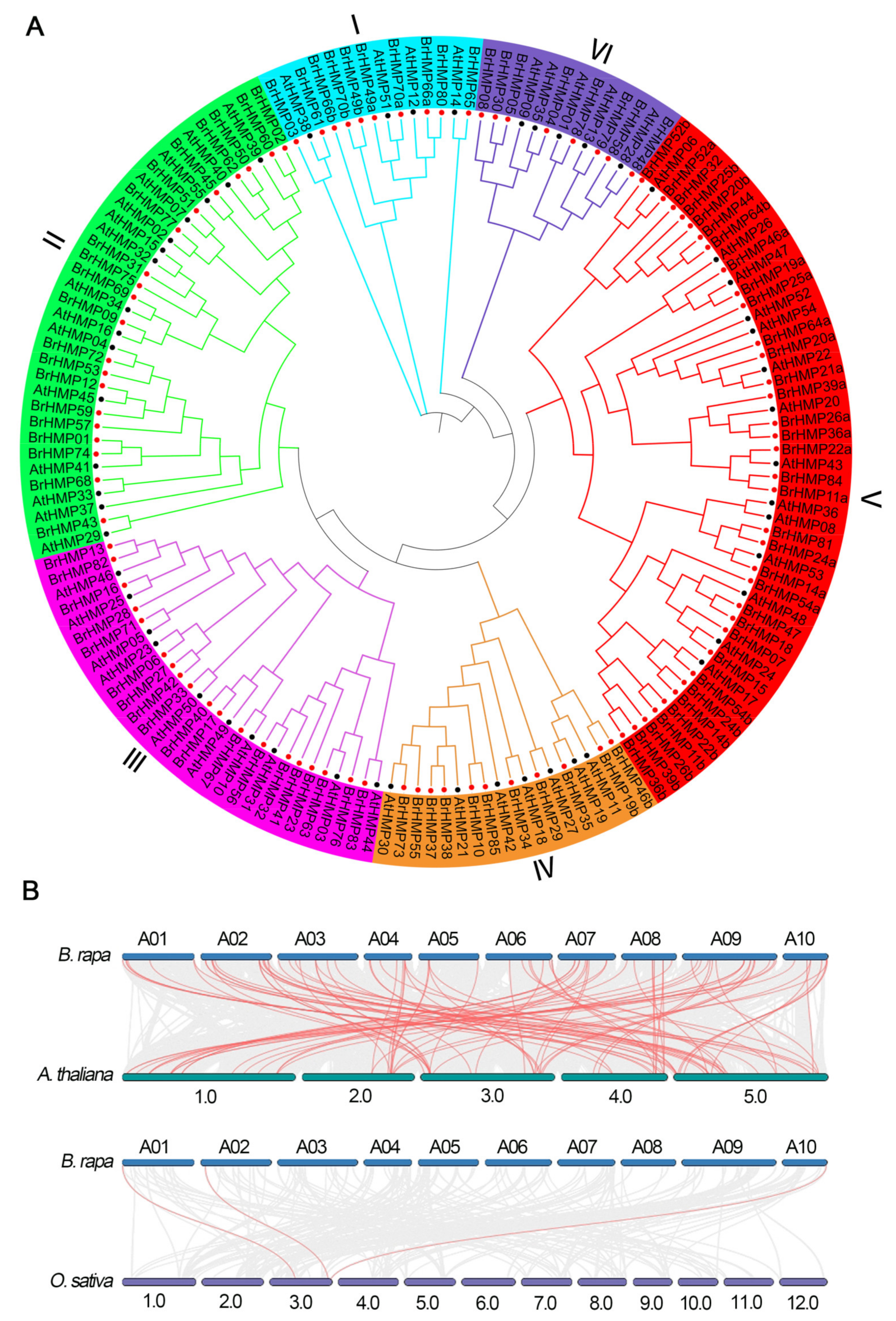
2.2. Chromosome Localization, Synteny Analysis, and Phylogenetic Tree Construction
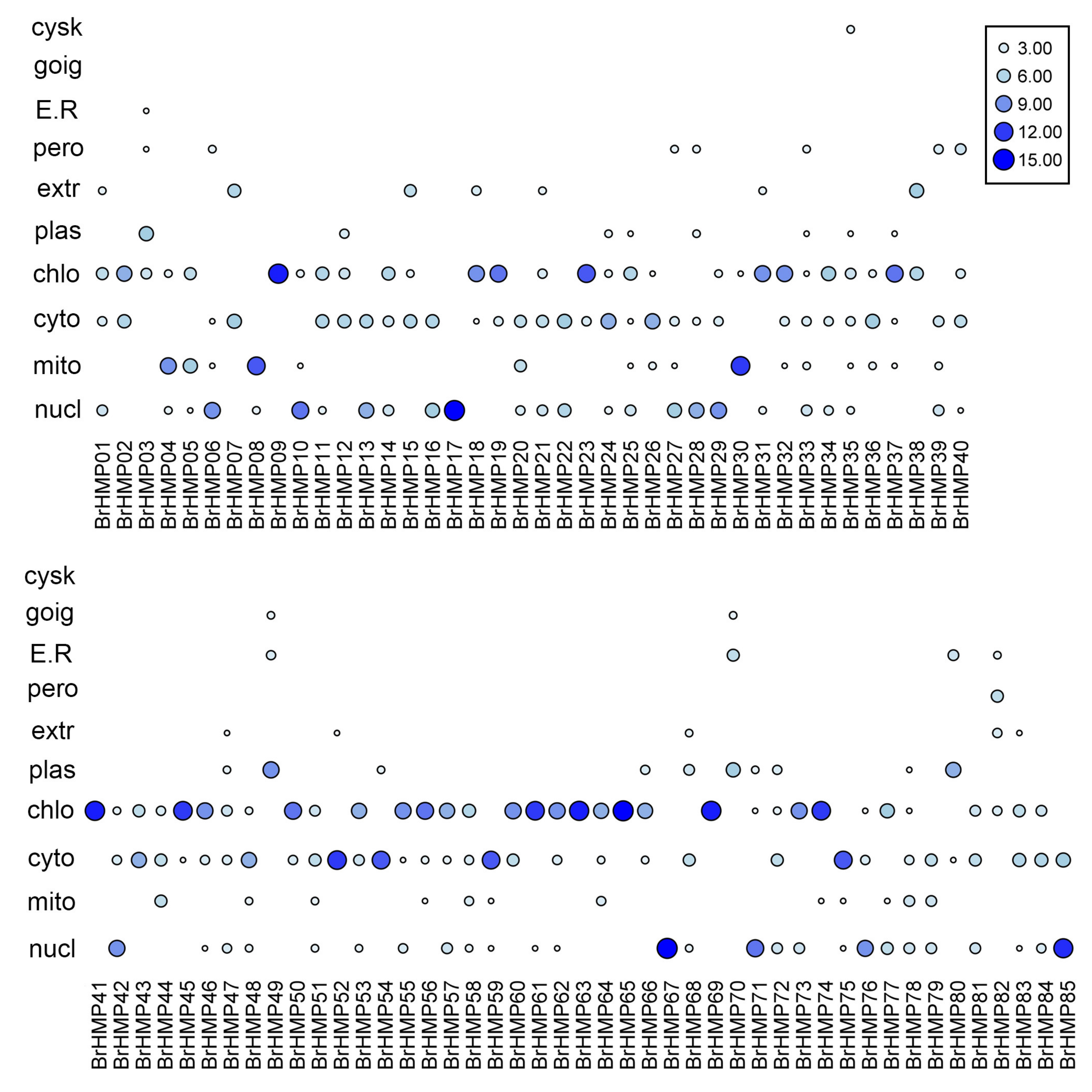
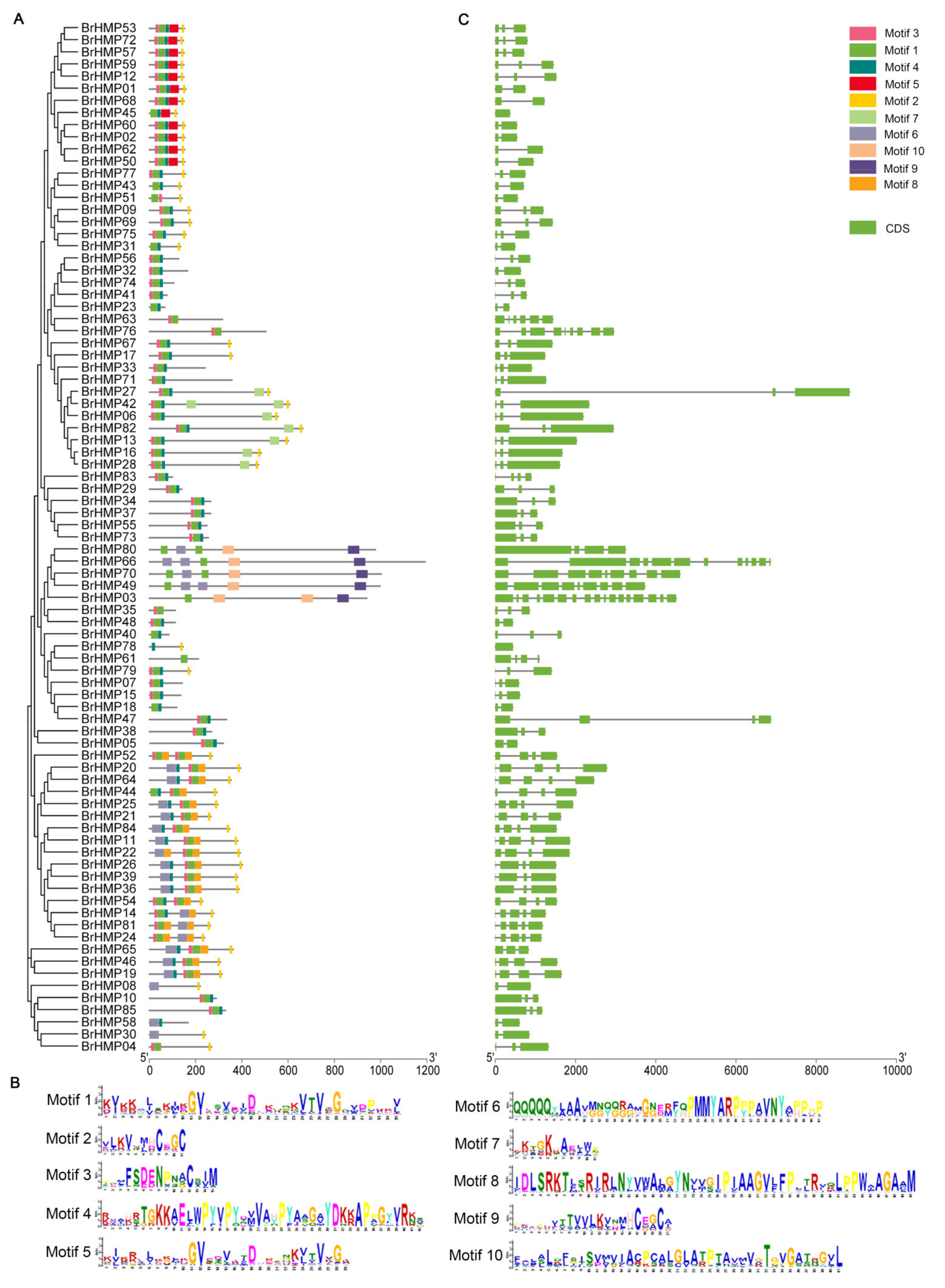
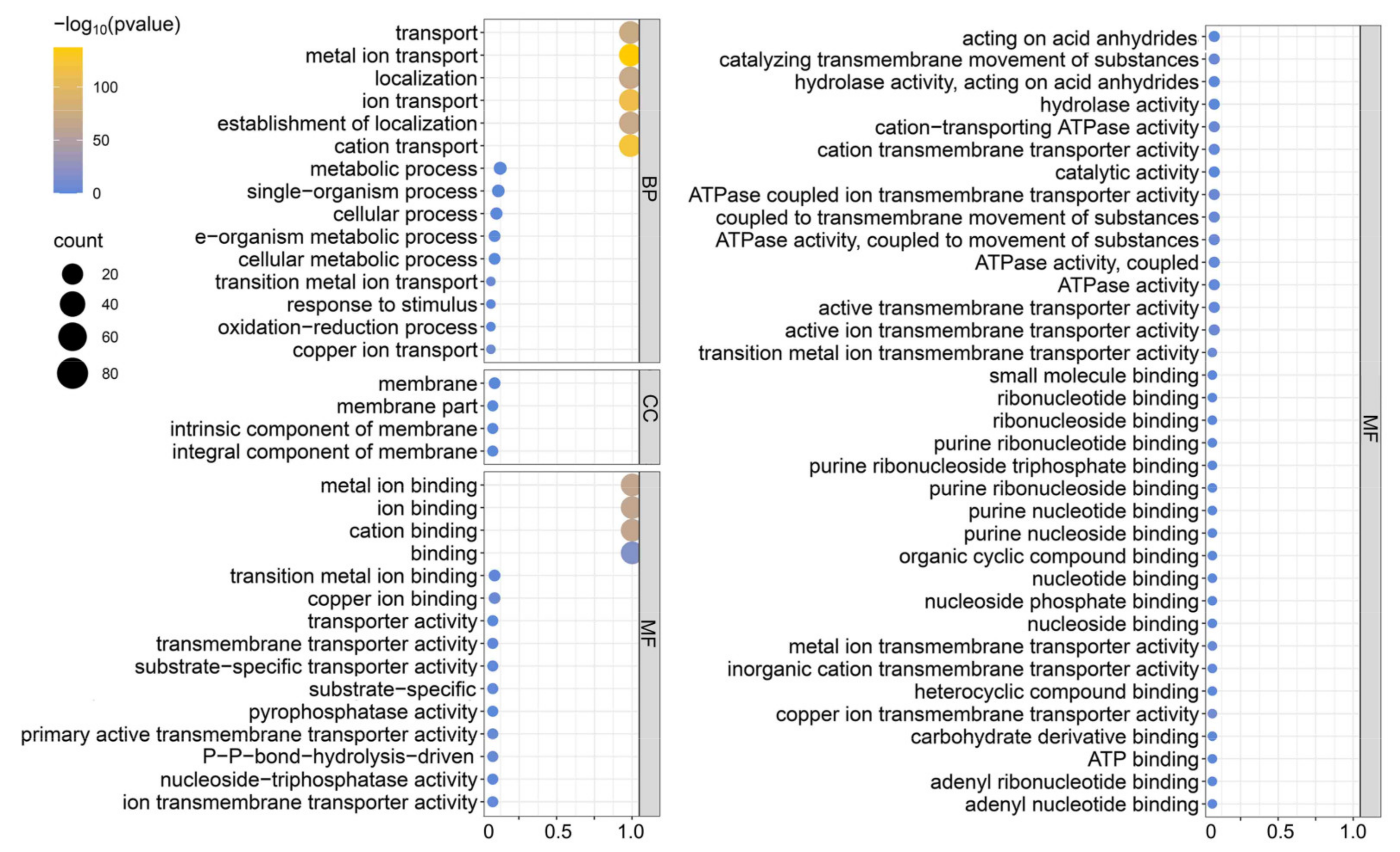
2.3. Conserved Motifs, Subcellular Localization, and GO Analysis
2.4. Promoter Element Analysis and Expression Analysis
2.5. Plant Material, Stress and Pollination Treatments
2.6. Total RNA Extraction and qRT-PCR
2.7. Protein–Protein Interaction Networks and Phosphorylation Site Prediction
3. Results
3.1. Basic Information and Chromosome Distribution Analysis
3.2. Phylogenetic Tree and Synteny Analysis
3.3. Subcellular Localization Analysis
3.4. Gene Structure and Conserved Motif Analysis
3.5. GO Analysis
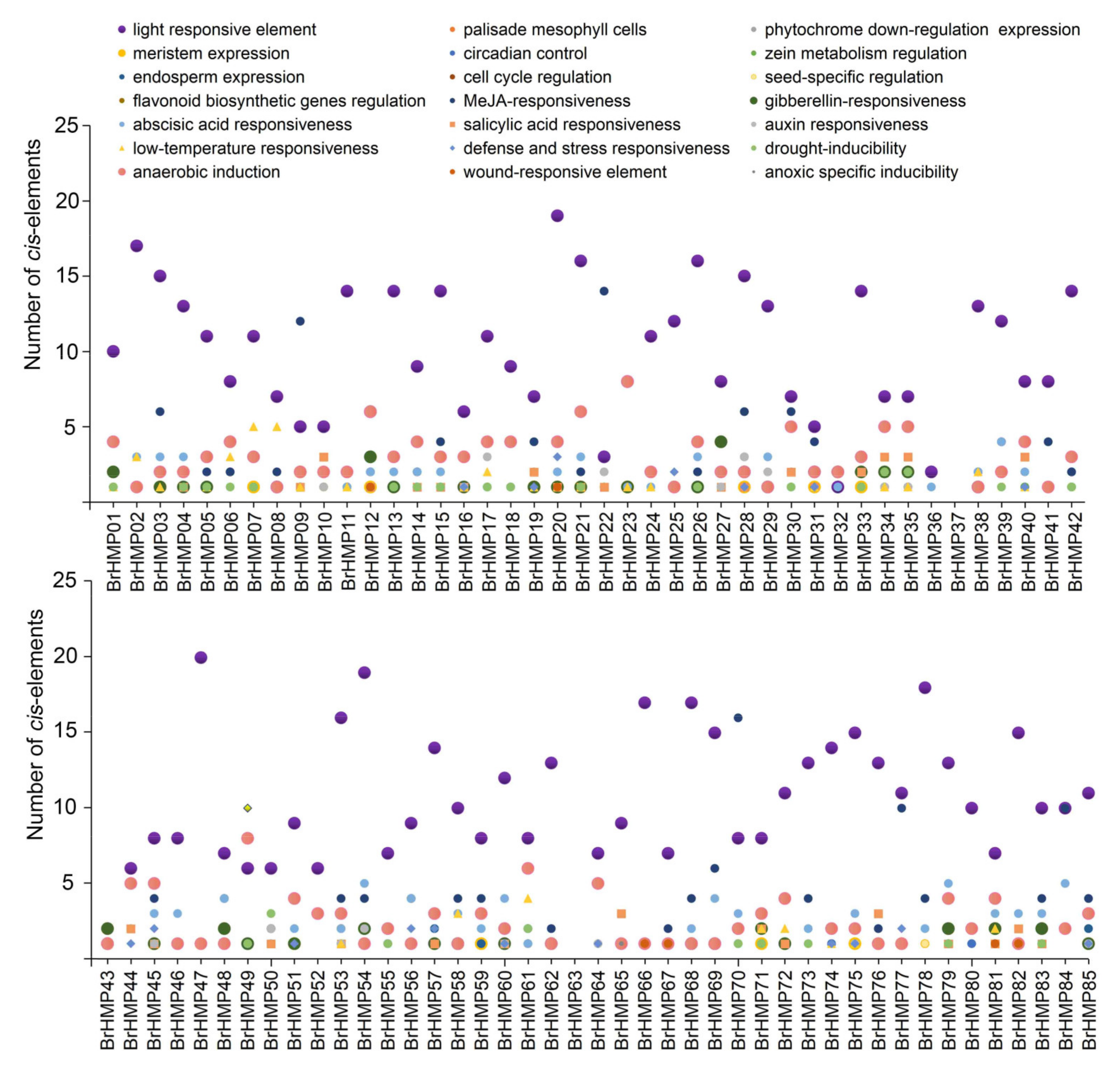
3.6. Promoter Cis-Regulatory Elements Analysis
3.7. Gene Expression Analysis
3.7.1. Expression Patterns of the BrHMPs in Different Tissues
3.7.2. Expression Patterns of the BrHMPs in Response to Heavy Metal Stress
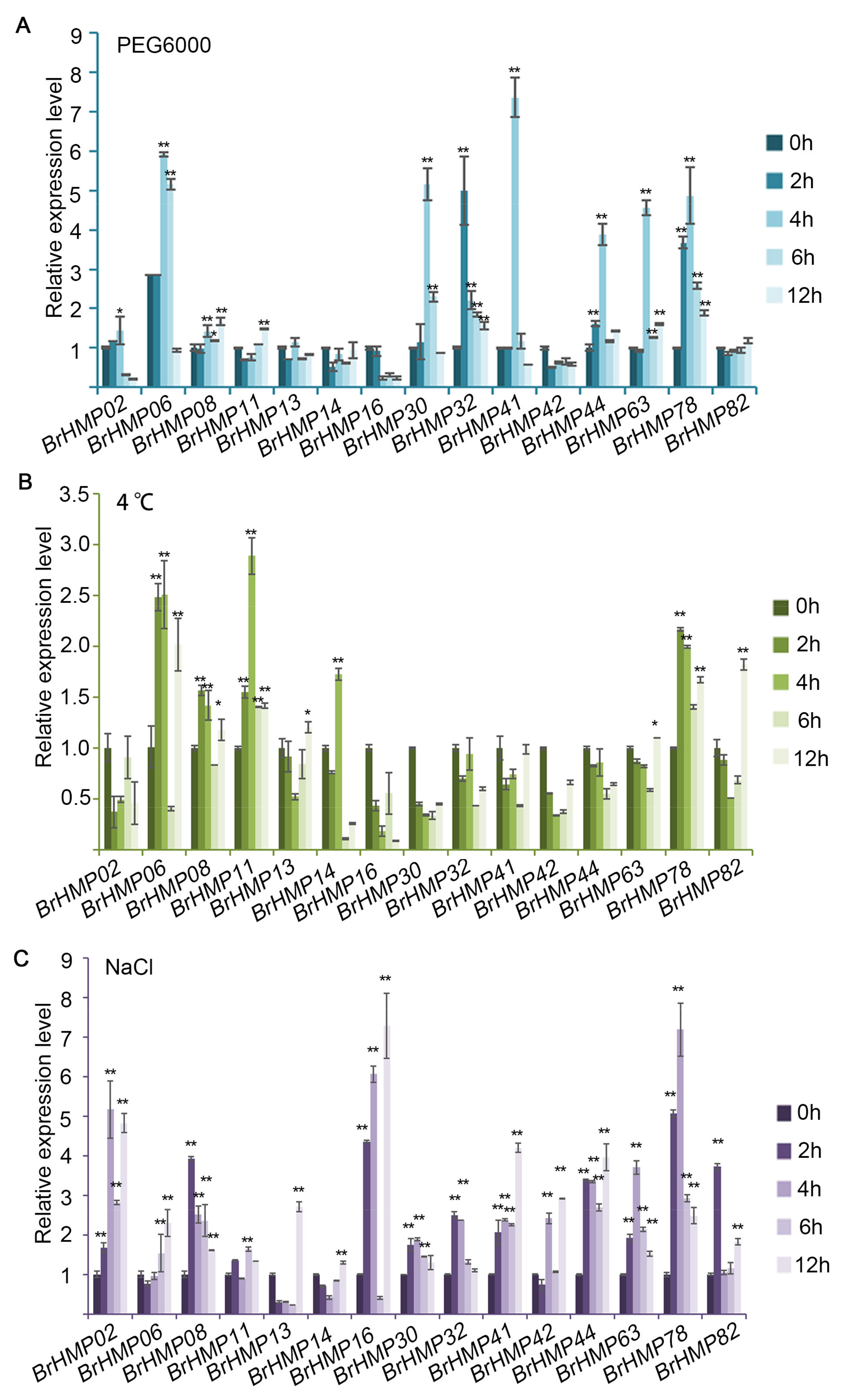
3.7.3. Expression Patterns of the BrHMPs in Abiotic Stress
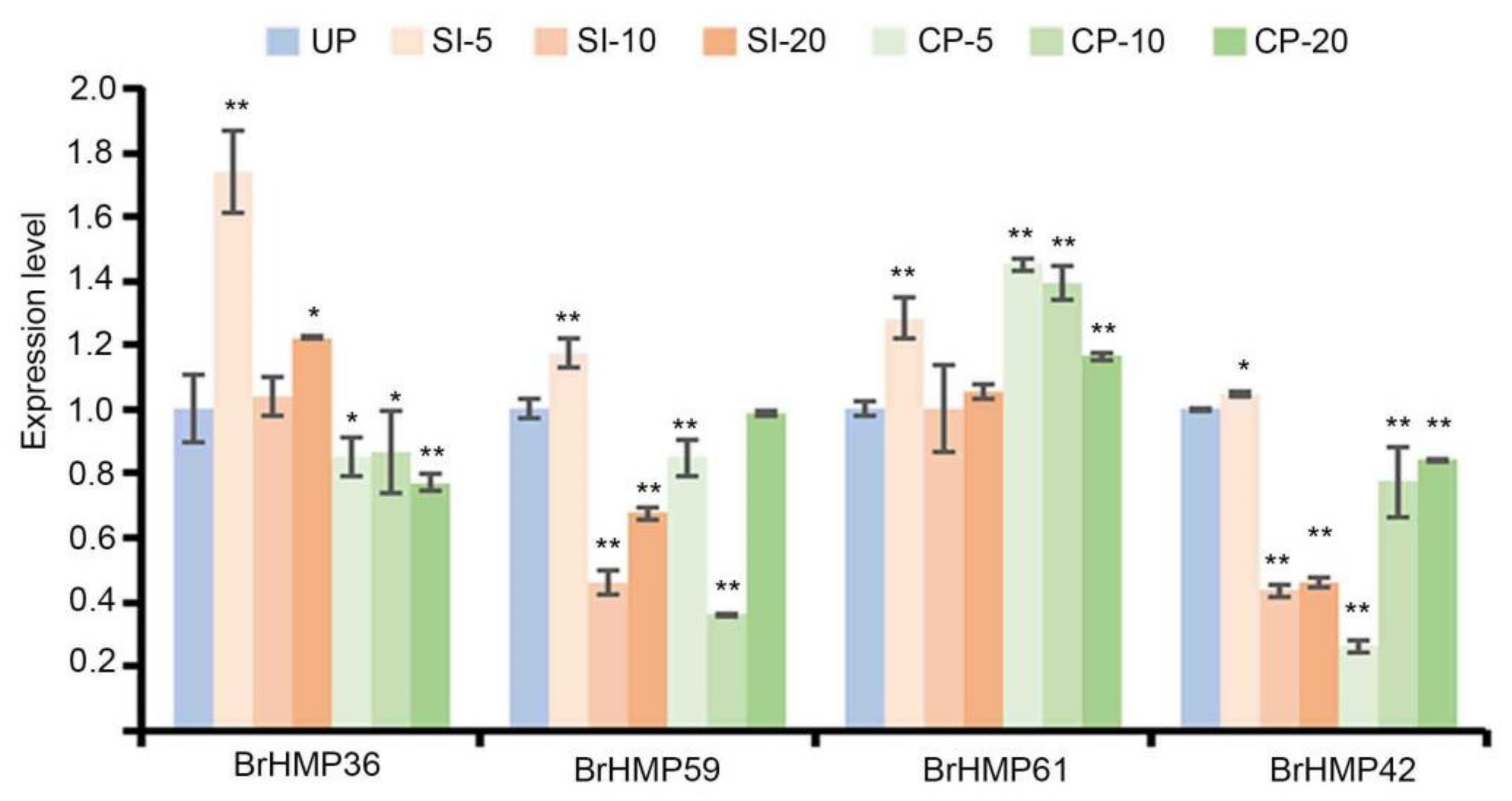
3.7.4. Expression of BrHMPs during SI and CP Responses
3.8. Protein–Protein Interaction (PPI) Networks
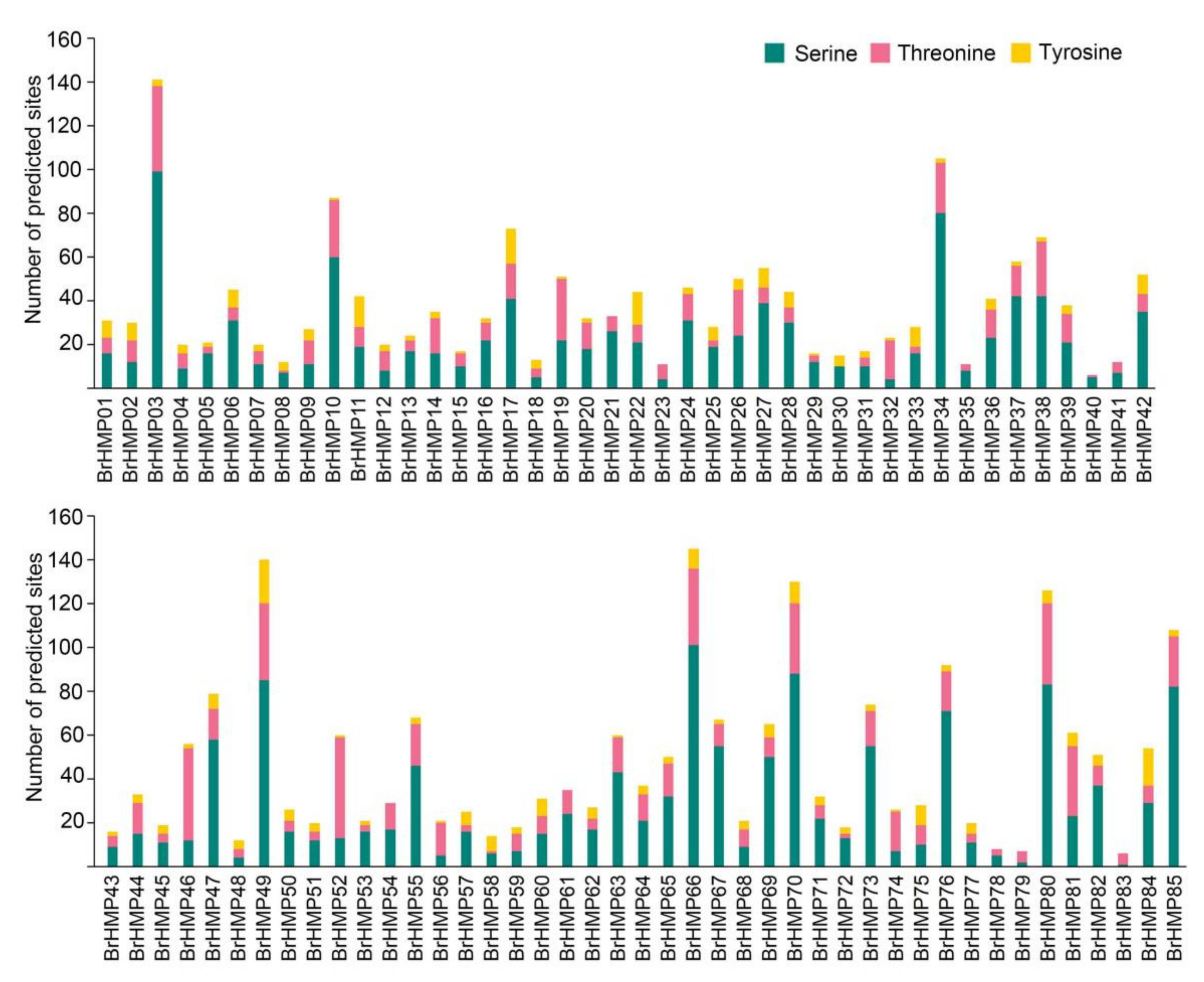
3.9. Phosphorylation Site Prediction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Petersen, P.E. World Health Organization. Economist 2009, 391, 102. [Google Scholar]
- Khan, M.A.; Castro-Guerrero, N.; Mendoza-Cozatl, D.G. Moving toward a precise nutrition: Preferential loading of seeds with essential nutrients over non-essential toxic elements. Front. Plant Sci. 2014, 5, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stohs, S.J.; Bagchi, D. Oxidative mechanisms in the toxicity of metal ions. Free Radic. Biol. Med. 1995, 18, 321–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naithani, S.C.; Chandrakar, V.; Keshavikant, S. Arsenic-induced metabolic disturbances and their mitigation mechanisms in crop plants: A review. Biol. Cas. Slov. Akad. Vied 2016, 71, 367–377. [Google Scholar]
- Chen, T.; Su, Y. Uptake by rice seedlings and in-plant degradation of atrazine as influenced by the oxidative stress induced by added arsenic or phosphate deficiency. Hum. Ecol. Risk Assess. 2018, 24, 1550–1564. [Google Scholar] [CrossRef]
- Mills, R.F.; Krijger, G.C.; Baccarini, P.J.; Hall, J.L.; Williams, L.E. Functional expression of AtHMA4, a P1B-type ATPase of the Zn/Co/Cd/Pb subclass. Plant J. 2003, 35, 164–176. [Google Scholar] [CrossRef]
- De Abreu-Neto, J.B.; Turchetto-Zolet, A.C.; de Oliveira, L.F.V.; Zanettini, M.H.B.; Margis-Pinheiro, M. Heavy metal-associated isoprenylated plant protein (HIPP): Characterization of a family of proteins exclusive to plants. FEBS J. 2013, 280, 1604–1616. [Google Scholar] [CrossRef]
- Puig, S.; Mira, H.; Dorcey, E.; Sancenón, V.; Andrés-Colás, N.; Garcia-Molina, A.; Burkhead, J.L.; Gogolin, K.A.; Abdel-Ghany, S.E.; Thiele, D.J.; et al. Higher plants possess two different types of ATX1-like copper chaperones. Biochem. Biophys. Res. Commun. 2007, 354, 385–390. [Google Scholar] [CrossRef]
- Kim, Y.Y.; Choi, H.; Segami, S.; Cho, H.T.; Martinoia, E.; Maeshima, M.; Lee, Y. AtHMA1 contributes to the detoxification of excess Zn(II) in Arabidopsis. Plant J. 2009, 58, 737–753. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.H.; Yu, G.; Li, J.T.; Jia, P.; Zhang, J.C.; Jia, C.G.; Zhang, Y.H.; Pan, H.Y. A heavy metal-associated protein (AcHMA1) from the halophyte, Atriplex canescens (Pursh) Nutt., confers tolerance to iron and other abiotic stresses when expressed in Saccharomyces cerevisiae. Int. J. Mol. Sci. 2014, 15, 14891–14906. [Google Scholar] [CrossRef]
- Wong, C.K.E.; Jarvis, R.S.; Sherson, S.M.; Cobbett, C.S. Functional analysis of the heavy metal binding domains of the Zn/Cd-transporting ATPase, HMA2, in Arabidopsis thaliana. New Phytol. 2009, 181, 79–88. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Kuroda, K.; Kimura, K.; Southron-Francis, J.L.; Furuzawa, A.; Iuchi, S.; Kobayashi, M.; Taylor, G.J.; Koyama, H. Amino Acid Polymorphisms in Strictly Conserved Domains of a P-Type ATPase HMA5 Are Involved in the Mechanism of Copper Tolerance Variation in Arabidopsis. Plant Physiol. 2008, 148, 969–980. [Google Scholar] [CrossRef] [Green Version]
- Shin, L.J.; Lo, J.C.; Yeh, K.C. Copper Chaperone Antioxidant Protein1 Is Essential for Copper Homeostasis. Plant Physiol. 2012, 159, 1099–1110. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.A.; Yoon, E.K.; Heo, J.O.; Lee, M.H.; Hwang, I.; Cheong, H.; Lee, W.S.; Hwang, Y.S.; Lim, J. Analysis of Arabidopsis glucose insensitive growth Mutants Reveals the Involvement of the Plastidial Copper Transporter PAA1 in Glucose-Induced Intracellular Signaling. Plant Physiol. 2012, 159, 1001–1012. [Google Scholar] [CrossRef] [Green Version]
- Parasyri, A.; Barth, O.; Zschiesche, W.; Humbeck, K. The Barley Heavy Metal Associated Isoprenylated Plant Protein HvFP1 Is Involved in a Crosstalk between the Leaf Development and Abscisic Acid-Related Drought Stress Responses. Plants 2022, 11, 2851. [Google Scholar] [CrossRef]
- Zschiesche, W.; Barth, O.; Daniel, K.; Bohme, S.; Rausche, J.; Humbeck, K. The zinc-binding nuclear protein HIPP3 acts as an upstream regulator of the salicylate-dependent plant immunity pathway and of flowering time in Arabidopsis thaliana. New Phytol. 2015, 207, 1084–1096. [Google Scholar] [CrossRef]
- Guan, X.X.; Sui, C.L.; Luo, K.C.; Chen, Z.F.; Feng, C.Y.; Dong, X.F.; Zeng, B.P.; Dong, X.; Liu, X.F. Effects of alpha-Naphthylacetic Acid on Cadmium Stress and Related Factors of Tomato by Regulation of Gene Expression. Agronomy 2022, 12, 2141. [Google Scholar] [CrossRef]
- Bradl, H. (Ed.) Heavy Metals in the Environment: Origin, Interaction and Remediation; Elsevier: Amsterdam, The Netherlands, 2005. [Google Scholar]
- Li, J.M.; Zhang, M.H.; Sun, J.; Mao, X.R.; Wang, J.G.; Liu, H.L.; Zheng, H.L.; Li, X.W.; Zhao, H.W.; Zou, D.T. Heavy Metal Stress-Associated Proteins in Rice and Arabidopsis: Genome-Wide Identification, Phylogenetics, Duplication, and Expression Profiles Analysis. Front. Genet. 2020, 11, 477. [Google Scholar] [CrossRef]
- He, G.; Qin, L.; Tian, W.; Meng, L.; He, T.; Zhao, D. Heavy Metal Transporters-Associated Proteins in S. tuberosum: Genome-Wide Identification, Comprehensive Gene Feature, Evolution and Expression Analysis. Genes 2020, 11, 1269. [Google Scholar] [CrossRef]
- Li, D.; Xu, X.; Hu, X.; Liu, Q.; Wang, Z.; Zhang, H.; Wang, H.; Wei, M.; Wang, H.; Liu, H.; et al. Genome-Wide Analysis and Heavy Metal-Induced Expression Profiling of the HMA Gene Family in Populus trichocarpa. Front. Plant Sci. 2015, 6, 1149. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Artimo, P.; Jonnalagedda, M.; Arnold, K.; Baratin, D.; Csardi, G.; de Castro, E.; Duvaud, S.; Flegel, V.; Fortier, A.; Gasteiger, E.; et al. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012, 40, W597–W603. [Google Scholar] [CrossRef] [PubMed]
- Xiong, E.H.; Zheng, C.Y.; Wu, X.L.; Wang, W. Protein Subcellular Location: The Gap Between Prediction and Experimentation. Plant Mol. Biol. Report. 2016, 34, 52–61. [Google Scholar] [CrossRef]
- Rombauts, S.; Déhais, P.; Van Montagu, M.; Rouzé, P. PlantCARE, a plant cis-acting regulatory element database. Nucleic Acids Res. 1999, 27, 295–296. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT. Methods 2013, 25, 402–408. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Bull, P.C.; Cox, D.W. Wilson disease and Menkes disease: New handles on heavy-metal transport. Trends Genet. 1994, 10, 246–252. [Google Scholar] [CrossRef]
- Imran, Q.M.; Falak, N.; Hussain, A.; Mun, B.G.; Sharma, A.; Lee, S.U.; Kim, K.M.; Yun, B.W. Nitric Oxide Responsive Heavy Metal-Associated Gene AtHMAD1 Contributes to Development and Disease Resistance in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 1712. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Zhang, X.; Liu, J.; Niu, Y.; Chen, Y.M.; Hao, Y.L.; Zhao, J.; Sun, L.; Wang, H.Y.; Xiao, J.; et al. Characterization of the Heavy-Metal-Associated Isoprenylated Plant Protein (HIPP) Gene Family from Triticeae Species. Int. J. Mol. Sci. 2020, 21, 6191. [Google Scholar] [CrossRef]
- Yang, L.; Lu, Y.J.; Tian, W.Q.; Feng, Y.L.; Bai, J.L.; Zhang, H. Insights into the functional divergence of the haloacid dehalogenase superfamily from phosphomonoesterase to inorganic pyrophosphatase. Arch. Biochem. Biophys. 2021, 705, 108896. [Google Scholar] [CrossRef]
- Aravind, L.; Galperin, M.Y.; Koonin, E.V. The catalytic domain of the P-type ATPase has the haloacid dehalogenase fold. Trends Biochem. Sci. 1998, 23, 127–129. [Google Scholar] [CrossRef]
- Gohla, A. Do metabolic HAD phosphatases moonlight as protein phosphatases? Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 153–166. [Google Scholar] [CrossRef]
- Zafra, A.; Rejon, J.D.; Hiscock, S.J.; Alche, J.D. Patterns of ROS Accumulation in the Stigmas of Angiosperms and Visions into Their Multi-Functionality in Plant Reproduction. Front. Plant Sci. 2016, 7, 1112. [Google Scholar] [CrossRef] [Green Version]
- Kodera, C.; Just, J.; Da Rocha, M.; Larrieu, A.; Riglet, L.; Legrand, J.; Rozier, F.; Gaude, T.; Fobis-Loisy, I. The molecular signatures of compatible and incompatible pollination in Arabidopsis. BMC Genom. 2021, 22, 268. [Google Scholar] [CrossRef]
- Zhang, L.L.; Huang, J.B.; Su, S.Q.; Wei, X.C.; Yang, L.; Zhao, H.H.; Yu, J.Q.; Wang, J.; Hui, J.Y.; Hao, S.Y.; et al. FERONIA receptor kinase-regulated reactive oxygen species mediate self-incompatibility in Brassica rapa. Curr. Biol. 2021, 31, 3004–3016.E4. [Google Scholar] [CrossRef]
- Klaumann, S.; Nickolaus, S.D.; Furst, S.H.; Starck, S.; Schneider, S.; Neuhaus, H.E.; Trentmann, O. The tonoplast copper transporter COPT5 acts as an exporter and is required for interorgan allocation of copper in Arabidopsis thaliana. New Phytol. 2011, 192, 393–404. [Google Scholar] [CrossRef]
- Shikanai, T.; Müller-Moulé, P.; Munekage, Y.; Niyogi, K.K.; Pilon, M. PAA1, a P-type ATPase of Arabidopsis, functions in copper transport in chloroplasts. Plant Cell 2003, 15, 1333–1346. [Google Scholar] [CrossRef] [Green Version]
- Andrés-Colás, N.; Sancenón, V.; Rodríguez-Navarro, S.; Mayo, S.; Thiele, D.J.; Ecker, J.R.; Puig, S.; Peñarrubia, L. The Arabidopsis heavy metal P-type ATPase HMA5 interacts with metallochaperones and functions in copper detoxification of roots. Plant J. 2006, 45, 225–236. [Google Scholar] [CrossRef]
- Boutigny, S.; Sautron, E.; Finazzi, G.; Rivasseau, C.; Frelet-Barrand, A.; Pilon, M.; Rolland, N.; Seigneurin-Berny, D. HMA1 and PAA1, two chloroplast-envelope PIB-ATPases, play distinct roles in chloroplast copper homeostasis. J. Exp. Bot. 2014, 65, 1529–1540. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Wang, L.; Zhao, F.J.; Wu, L.; Liu, A.; Xu, W. SpHMA1 is a chloroplast cadmium exporter protecting photochemical reactions in the Cd hyperaccumulator Sedum plumbizincicola. Plant Cell Environ. 2019, 42, 1112–1124. [Google Scholar] [CrossRef]
- Zhu, Y.X.; Dai, Y.J.; Jing, X.T.; Liu, X.X.; Jin, C.W. Inhibition of BRUTUS Enhances Plant Tolerance to Zn Toxicity by Upregulating Pathways Related to Iron Nutrition. Life 2022, 12, 216. [Google Scholar] [CrossRef] [PubMed]
- Selote, D.; Matthiadis, A.; Gillikin, J.W.; Sato, M.H.; Long, T.A. The E3 ligase BRUTUS facilitates degradation of VOZ1/2 transcription factors. Plant Cell Environ. 2018, 41, 2463–2474. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Sato, S.; Sasaki, K.; Saburi, W.; Matsui, H.; Imai, R. COLD SHOCK DOMAIN PROTEIN 3 is involved in salt and drought stress tolerance in Arabidopsis. FEBS Open Bio 2013, 3, 438–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granlund, I.; Storm, P.; Schubert, M.; García-Cerdán, J.G.; Funk, C.; Schröder, W.P. The TL29 protein is lumen located, associated with PSII and not an ascorbate peroxidase. Plant Cell Physiol. 2009, 50, 1898–1910. [Google Scholar] [CrossRef] [Green Version]
- Padmanaban, S.; Czerny, D.D.; Levin, K.A.; Leydon, A.R.; Su, R.T.; Maugel, T.K.; Zou, Y.; Chanroj, S.; Cheung, A.Y.; Johnson, M.A.; et al. Transporters involved in pH and K+ homeostasis affect pollen wall formation, male fertility, and embryo development. J. Exp. Bot. 2017, 68, 3165–3178. [Google Scholar] [CrossRef] [Green Version]
- Rosenzweig, A.C.; Arguello, J.M. Toward a Molecular Understanding of Metal Transport by P-1B-Type ATPases. In Metal Transporters; Lutsenko, S., Arguello, J.M., Eds.; Current Topics in Membranes Book Series; Academic Press: Cambridge, MA, USA, 2012; Volume 69, pp. 113–136. [Google Scholar]
- Sautron, E.; Mayerhofer, H.; Giustini, C.; Pro, D.; Crouzy, S.; Ravaud, S.; Pebay-Peyroula, E.; Rolland, N.; Catty, P.; Seigneurin-Berny, D. HMA6 and HMA8 are two chloroplast Cu+-ATPases with different enzymatic properties. Biosci. Rep. 2015, 35, e00201. [Google Scholar] [CrossRef]
- Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.H.; Bancroft, I.; Cheng, F.; Li, X.; Hua, W.; Wang, X. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar]
- Bowers, J.E.; Chapman, B.A.; Rong, J.; Paterson, A.H. Unravelling angiosperm genome evolution by phylogenetic analysis of chromosomal duplication events. Nature 2003, 422, 433–438. [Google Scholar] [CrossRef]
- Chen, H.; Wang, T.; He, X.; Cai, X.; Lin, R.; Liang, J.; Wu, J.; King, G.; Wang, X. BRAD V3.0: An upgraded Brassicaceae database. Nucleic Acids Res. 2022, 50, D1432–D1441. [Google Scholar] [CrossRef]
- Delledonne, M.; Xia, Y.; Dixon, R.A.; Lamb, C. Nitric oxide functions as a signal in plant disease resistance. Nature 1998, 394, 585–588. [Google Scholar] [CrossRef]
- Williams, L.E.; Pittman, J.K.; Hall, J.L. Emerging mechanisms for heavy metal transport in plants. Biochim. Biophys. Acta 2000, 1465, 104–126. [Google Scholar] [CrossRef]
- Sheteiwy, M.S.; Gong, D.T.; Gao, Y.; Pan, R.H.; Hu, J.; Guan, Y.J. Priming with methyl jasmonate alleviates polyethylene glycol-induced osmotic stress in rice seeds by regulating the seed metabolic profile. Environ. Exp. Bot. 2018, 153, 236–248. [Google Scholar] [CrossRef]
- Duric, M.; Subotic, A.; Trifunovic-Momcilov, M.; Milosevic, S. Improvement of water deficit stress tolerance of Impatiens walleriana shoots grown in vitro by methyl jasmonate. Plant Cell Tissue Organ Cult. 2022. [Google Scholar] [CrossRef]
- Ma, B.J.; Liu, X.F.; Guo, S.Y.; Xie, X.L.; Zhang, J.; Wang, J.Y.; Zheng, L.L.; Wang, Y.C. RtNAC100 involved in the regulation of ROS, Na plus accumulation and induced salt-related PCD through MeJA signal pathways in recretohalophyte Reaumuria trigyna. Plant Sci. 2021, 310, 110976. [Google Scholar] [CrossRef]
- Sanchez-Fernandez, R.; Davies, T.; Coleman, J.; Rea, P.A. The Arabidopsis thaliana ABC Protein Superfamily, a Complete Inventory. J. Biol. Chem. 2001, 276, 30231–30244. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Song, W.Y.; Ko, D.; Eom, Y.; Hansen, T.H.; Schiller, M.; Lee, T.G.; Martinoia, E.; Lee, Y. The phytochelatin transporters AtABCC1 and AtABCC2 mediate tolerance to cadmium and mercury. Plant J. 2012, 69, 278–288. [Google Scholar] [CrossRef]
- Song, W.Y.; Mendoza-Cozatl, D.G.; Lee, Y.; Schroeder, J.I.; Ahn, S.N.; Lee, H.S.; Wicker, T.; Martinoia, E. Phytochelatin-metal(loid) transport into vacuoles shows different substrate preferences in barley and Arabidopsis. Plant Cell Environ. 2014, 37, 1192–1201. [Google Scholar] [CrossRef] [Green Version]
- Song, W.Y.; Park, J.; Mendoza-Cózatl, D.G.; Suter-Grotemeyer, M.; Shim, D.; Hörtensteiner, S.; Geisler, M.; Weder, B.; Rea, P.A.; Rentsch, D.; et al. Arsenic tolerance in Arabidopsis is mediated by two ABCC-type phytochelatin transporters. Proc. Natl. Acad. Sci. USA 2010, 107, 21187–21192. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.; Cho, H.T. The function of ABCB transporters in auxin transport. Plant Signal. Behav. 2013, 8, e22990. [Google Scholar] [CrossRef] [Green Version]
- Ullah, A.; Zeng, F.J.; Tariq, A.; Asghar, M.A.; Saleem, K.; Raza, A.; Naseer, M.A.; Zhang, Z.H.; Noor, J. Exogenous naphthaleneacetic acid alleviated alkalinity-induced morpho-physio-biochemical damages in Cyperus esculentus L. var. sativus Boeck. Front. Plant Sci. 2022, 13, 1018787. [Google Scholar] [CrossRef]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic acid: Emergence of a core signaling network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama, S.; Isogai, A. Self-incompatibility in plants. Annu. Rev. Plant Biol. 2005, 56, 467–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aist, R.J. Papillae and Related Wound Plugs of Plant Cells. Annu. Rev. Phytopathol. 1976, 141, 145–163. [Google Scholar] [CrossRef]
- Levine, A.; Tenhaken, R.; Dixon, R.; Lamb, C. H2O2 from the oxidative burst orchestrates the plant hypersensitive disease resistance response. Cell 1994, 79, 583–593. [Google Scholar] [CrossRef]
- Huckelhoven, R. Hypersensitive Cell Death and Papilla Formation in Barley Attacked by the Powdery Mildew Fungus Are Associated with Hydrogen Peroxide but Not with Salicylic Acid Accumulation. Plant Physiol. 1999, 119, 1251–1260. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Y.; Hecker, A.G.; Hauser, B.A. The APX4 locus regulates seed vigor and seedling growth in Arabidopsis thaliana. Planta 2014, 239, 909–919. [Google Scholar] [CrossRef]
- Dickinson, H. Dry stigmas, water and self-incompatibility in Brassica. Sex. Plant Reprod. 1995, 8, 1–10. [Google Scholar] [CrossRef]
- Elleman, C.J.; Franklin-Tong, V.; Dickinson, H.G. Pollination in species with dry stigmas: The nature of the early stigmatic response and the pathway taken by pollen tubes. New Phytol. 1992, 121, 413–424. [Google Scholar] [CrossRef]
- Hiroi, K.; Sone, M.; Sakazono, S.; Osaka, M.; Masuko-Suzuki, H.; Matsuda, T.; Suzuki, G.; Suwabe, K.; Watanabe, M. Time-lapse imaging of self- and cross-pollinations in Brassica rapa. Ann. Bot. 2013, 112, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Wu, Y.; Zhang, G. Formation pattern and regulatory mechanisms of pollen wall in Arabidopsis. J. Plant Physiol. 2021, 260, 153388. [Google Scholar] [CrossRef]
- Huang, X.Y.; Niu, J.; Sun, M.X.; Zhu, J.; Gao, J.F.; Yang, J.; Zhou, Q.; Yang, Z.N. CYCLIN-DEPENDENT KINASE G1 is associated with the spliceosome to regulate CALLOSE SYNTHASE5 splicing and pollen wall formation in Arabidopsis. Plant Cell 2013, 25, 637–648. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.L.; Xue, J.S.; Zhang, F.; Yao, C.; Shen, S.Y.; Sui, C.X.; Peng, Y.J.; Xu, Q.L.; Feng, Y.F.; Hu, W.J.; et al. A dye combination for the staining of pollen coat and pollen wall. Plant Reprod. 2021, 34, 91–101. [Google Scholar] [CrossRef]
- Kazan, K.; Manners, J.M. MYC2: The master in action. Mol. Plant 2013, 6, 686–703. [Google Scholar] [CrossRef] [Green Version]
- Zhai, Q.; Yan, L.; Tan, D.; Chen, R.; Sun, J.; Gao, L.; Dong, M.Q.; Wang, Y.; Li, C. Phosphorylation-coupled proteolysis of the transcription factor MYC2 is important for jasmonate-signaled plant immunity. PLoS Genet. 2013, 9, e1003422. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Wu, X.; Liu, S.; Zhang, L.; Li, T.; Cao, Y.; Duan, Q. Comprehensive Analysis of BrHMPs Reveals Potential Roles in Abiotic Stress Tolerance and Pollen–Stigma Interaction in Brassica rapa. Cells 2023, 12, 1096. https://doi.org/10.3390/cells12071096
Yang L, Wu X, Liu S, Zhang L, Li T, Cao Y, Duan Q. Comprehensive Analysis of BrHMPs Reveals Potential Roles in Abiotic Stress Tolerance and Pollen–Stigma Interaction in Brassica rapa. Cells. 2023; 12(7):1096. https://doi.org/10.3390/cells12071096
Chicago/Turabian StyleYang, Lin, Xiaoyu Wu, Shangjia Liu, Lina Zhang, Ting Li, Yunyun Cao, and Qiaohong Duan. 2023. "Comprehensive Analysis of BrHMPs Reveals Potential Roles in Abiotic Stress Tolerance and Pollen–Stigma Interaction in Brassica rapa" Cells 12, no. 7: 1096. https://doi.org/10.3390/cells12071096