
Current Knowledge about the Impact of Microgravity on Gene Regulation
 , ,
, ,  , , , ,
, , , ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
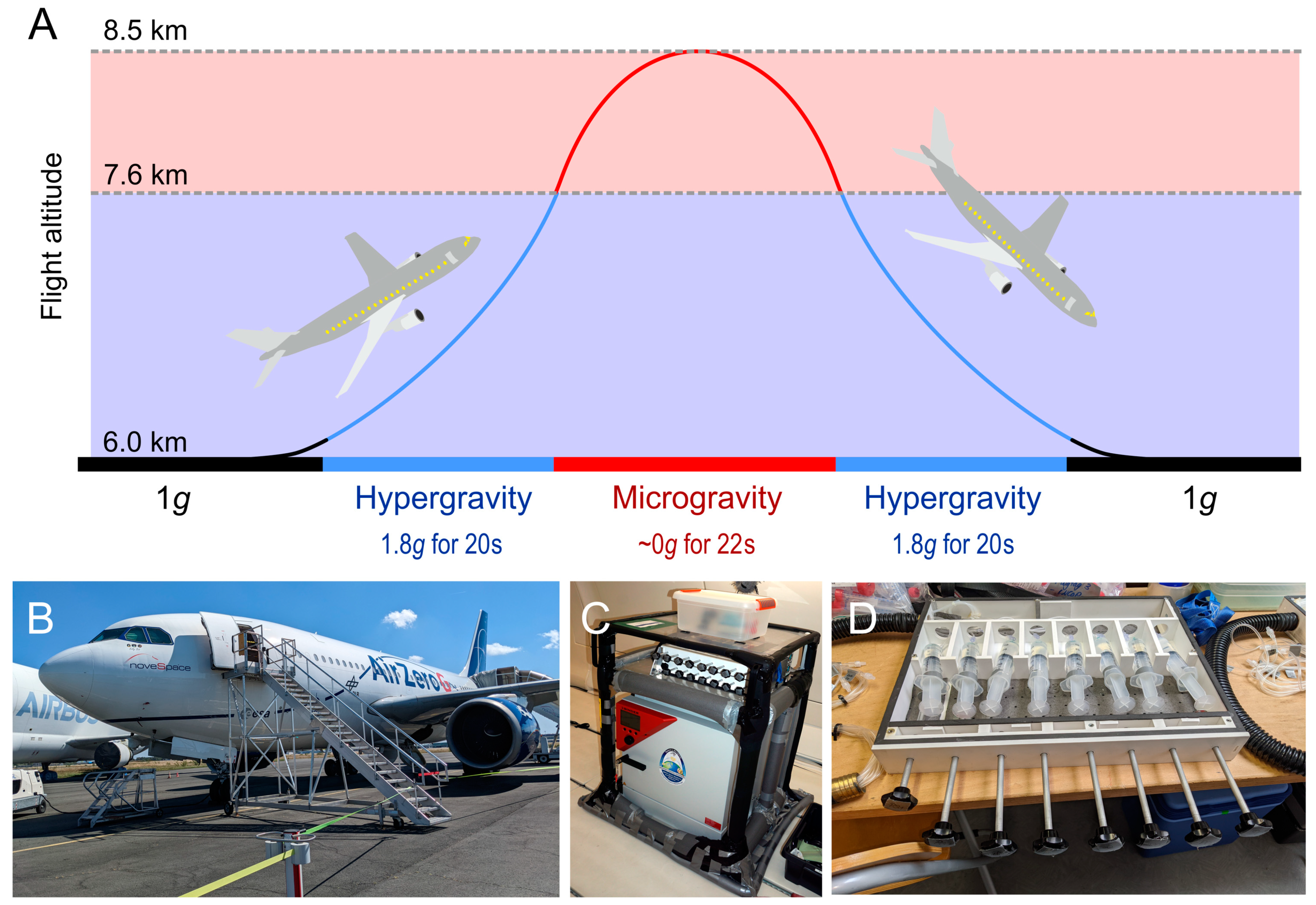
3.1. Microgravity Platforms
3.2. Recent Reports on Microgravity Effects on Bacteria, Protists, and Fungi
3.2.1. Microgravity Affects Homeostasis between Microbiome and Host
3.2.2. Microgravity and Virulence
3.2.3. Physiological Effects of Microgravity on Bacteria and Fungi
3.3. Effects of Real or Simulated Microgravity on the Gene Expression in Animals
3.4. Effects of Microgravity on the Eye and Brain
3.4.1. Effects of Microgravity on Cells with Eye Origin
3.4.2. Effects of Microgravity on Eye in Animals

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Kind | Gene Regulation | Microgravity | Reference |
|---|---|---|---|---|
| (i) Cell line | ARPE-19 |
| Spaceflight to the ISS, 3 d | [99] |
| (ii) Mice | C57BL/6 Male 9-week-old |
| Spaceflight to the ISS, 35 d | [101] |
| (ii) Mice | C57BL/6 Male 10-week-old |
| Spaceflight to the ISS, 35 d | [102] |
| (ii) Mice | C57BL/6 Male 10-week-old |
| Spaceflight to the ISS, 35 d | [103] |
| (ii) Mice | C57BL/6 Female 16-week-old |
| Spaceflight to the ISS, 37 d | [104] |
| (ii) Mice | Strain N/A Female |
| Hindlimb unloading | [105] |
| (iii) Human healthy volunteers | Eight male subjects* |
| 1 h settings of seated, HDT, and HDT + CO2 | [106] |
| (iii) Human healthy volunteers | Six men and five women * |
| 30 d of 6° HDT bed rest with 0.5% CO2 exposure | [107] |
3.4.3. Effects of Microgravity on Eye and Brain in Human Subjects
3.5. Effects of Microgravity on Endothelial Cells
3.6. Microgravity Affects the Immune System in Space
3.7. Effects of Microgravity on Cartilage
3.8. Effects of Microgravity on Muscle Cells
3.9. Effects of Microgravity on Bone
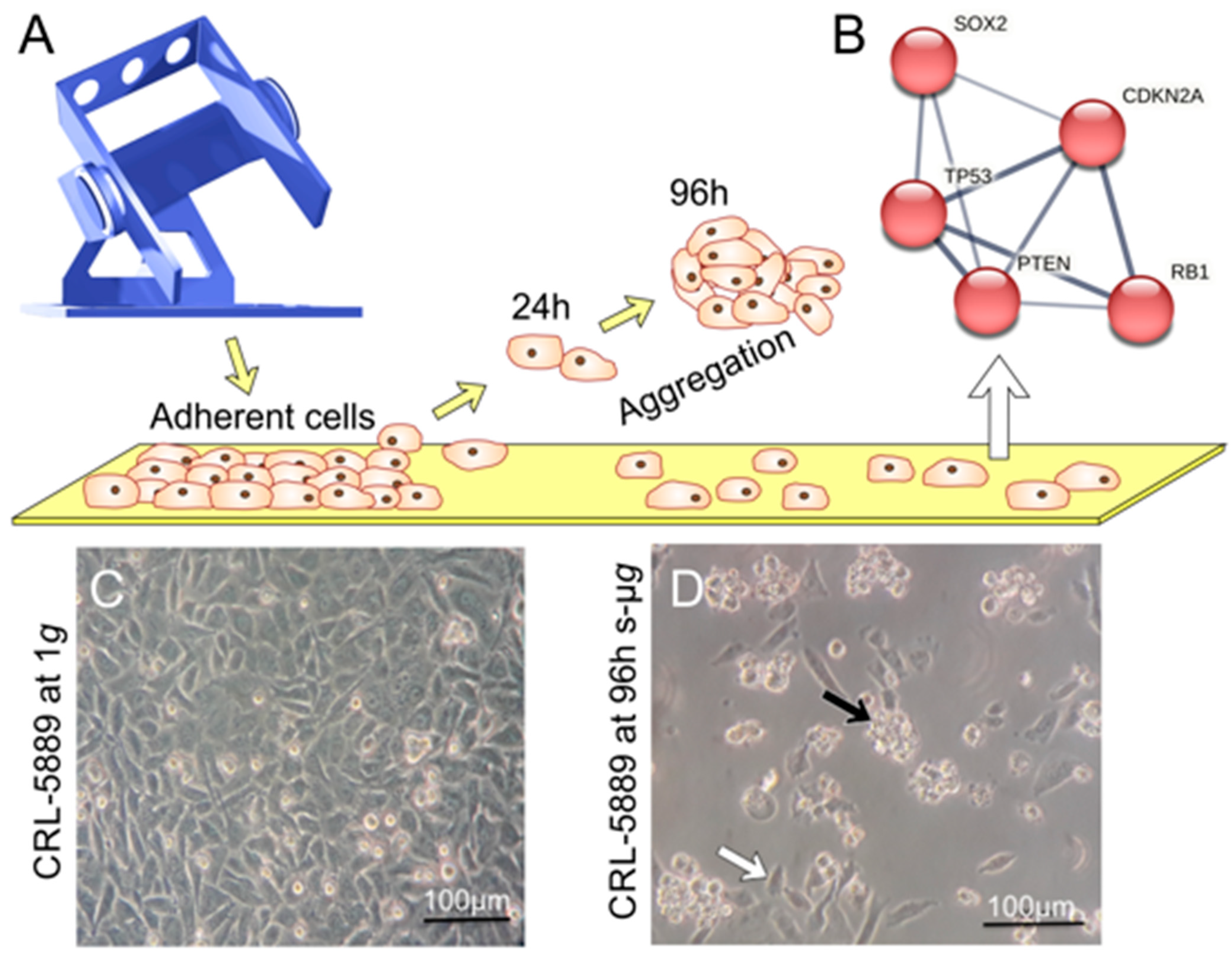
3.10. The Impact of Microgravity on Cancer Cells
3.10.1. Gene Expression in Thyroid Cancer Cells under the Influence of Gravitational Changes
| Cancer Type | Cell Line | Gene Regulation | Microgravity | Reference |
|---|---|---|---|---|
| Thyroid Cancer | FTC-133 |
| Spaceflight to the ISS, 5 d | [182] |
| FTC-133 |
| RPM, 4 h, 3 d | [183] | |
| FTC-133 |
| Spaceflight to the ISS, 12 d | [184] | |
| FTC-133 WRO ML-1 Nthy-ori 3-1 |
| RPM 3 d | [185] | |
| Breast cancer | CRL2351 |
| RPM, 5 d | [186] |
| MCF-7 |
| RPM, 24 h | [187] | |
| MCF-7, MDA-MB-231 |
| RPM, 24 h | [188] | |
| MCF-7 |
| TEXUS-54, 6 min of r-µg, PFC, 31 parabolas | [180] | |
| MDA-MB-231 |
| PFC, 31 parabolas | [189] | |
| MDA-MB-231 |
| RPM 24 h, 72 h | [190] | |
| MCF-7, MDA-MB-231 |
| RPM 14 d | [191] | |
| Prostate cancer | PC-3 |
| RPM, 3 d, 5 d | [192] |
| PC-3 |
| RPM, 30 min, 2 d, 4 d, and 24 h | [193] | |
| PC-3 |
| PFC, 31 parabolas | [194] | |
| Lung cancer | A549, H1703 |
| 3D clinostat, 24 h, 38 h | [195] |
| CRL-5889 |
| RPM, 72 h | [196] | |
| A549 |
| 2D clinostat 24 h, 48 h, 72 h | [197] | |
| Colorectal cancer | DLD1 cells |
| RCCS-HARV | [198] |
| Gastric Cancer | EPG85-257 RDB EPG85-257 P |
| RCCS, 72 h | [199] |
| Liver cancer | HepG2 cells hBTSCs |
| RCCS, 15 d | [200] |
| Pancreas Cancer | PaCa-44 |
| RPM, 1 d, 7 d, 9 d | [201] |
| Skin cancer (Melanoma) | BLM |
| Fast rotating 2D clinostat | [202] |
3.10.2. Breast Cancer Cells in Microgravity with a Focus on Gene Expression
3.10.3. Gene Expression in Prostate Cancer Cells under the Influence of Gravitational Changes
3.10.4. Gene Expression Changes of Lung Cancer Cells Exposed to Microgravity Conditions
3.10.5. Cancer Cells of the Gastrointestinal Tract Exposed to Microgravity
3.10.6. Gene Expression Changes of Skin Cancer Cells Exposed to Microgravity
3.11. Recent Findings in Plants under Microgravity Conditions
3.11.1. Growth and Development
3.11.2. Cell Wall Reorganization
3.11.3. Plastid Dysregulation
3.11.4. Post-Transcriptional and Translational Regulation
3.11.5. Experimental Setup and Controls
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moroni, L.; Tabury, K.; Stenuit, H.; Grimm, D.; Baatout, S.; Mironov, V. What can biofabrication do for space and what can space do for biofabrication? Trends Biotechnol. 2022, 40, 398–411. [Google Scholar] [CrossRef]
- White, R.J.; Averner, M. Humans in space. Nature 2001, 409, 1115–1118. [Google Scholar] [CrossRef] [PubMed]
- Blaber, E.; Marçal, H.; Burns, B.P. Bioastronautics: The influence of microgravity on astronaut health. Astrobiology 2010, 10, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Baran, R.; Marchal, S.; Garcia Campos, S.; Rehnberg, E.; Tabury, K.; Baselet, B.; Wehland, M.; Grimm, D.; Baatout, S. The cardiovascular system in space: Focus on in vivo and in vitro studies. Biomedicines 2021, 10, 59. [Google Scholar] [CrossRef] [PubMed]
- Russomano, T.; da Rosa, M.; Dos Santos, M.A. Space motion sickness: A common neurovestibular dysfunction in microgravity. Neurol. India 2019, 67, S214–S218. [Google Scholar] [CrossRef] [Green Version]
- Nelson, E.S.; Mulugeta, L.; Myers, J.G. Microgravity-induced fluid shift and ophthalmic changes. Life 2014, 4, 621–665. [Google Scholar] [CrossRef]
- Penchev, R.; Scheuring, R.A.; Soto, A.T.; Miletich, D.M.; Kerstman, E.; Cohen, S.P. Back pain in outer space. Anesthesiology 2021, 135, 384–395. [Google Scholar] [CrossRef]
- Baran, R.; Wehland, M.; Schulz, H.; Heer, M.; Infanger, M.; Grimm, D. Microgravity-related changes in bone density and treatment options: A systematic review. Int. J. Mol. Sci. 2022, 23, 8650. [Google Scholar] [CrossRef] [PubMed]
- Riwaldt, S.; Corydon, T.J.; Pantalone, D.; Sahana, J.; Wise, P.; Wehland, M.; Krüger, M.; Melnik, D.; Kopp, S.; Infanger, M.; et al. Role of apoptosis in wound healing and apoptosis alterations in microgravity. Front. Bioeng. Biotechnol. 2021, 9, 679650. [Google Scholar] [CrossRef] [PubMed]
- Schulz, H.; Strauch, S.M.; Richter, P.; Wehland, M.; Krüger, M.; Sahana, J.; Corydon, T.J.; Wise, P.; Baran, R.; Lebert, M.; et al. Latest knowledge about changes in the proteome in microgravity. Expert Rev. Proteom. 2022, 19, 43–59. [Google Scholar] [CrossRef]
- Krüger, J.; Richter, P.; Stoltze, J.; Strauch, S.M.; Krüger, M.; Daiker, V.; Prasad, B.; Sonnewald, S.; Reid, S.; Lebert, M. Changes of gene expression in euglena gracilis obtained during the 29(th) dlr parabolic flight campaign. Sci. Rep. 2019, 9, 14260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, X.; Pietsch, J.; Wehland, M.; Schulz, H.; Saar, K.; Hübner, N.; Bauer, J.; Braun, M.; Schwarzwälder, A.; Segerer, J.; et al. Differential gene expression profile and altered cytokine secretion of thyroid cancer cells in space. FASEB J. 2014, 28, 813–835. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Wehland, M.; Schulz, H.; Saar, K.; Hübner, N.; Infanger, M.; Bauer, J.; Grimm, D. Genomic approach to identify factors that drive the formation of three-dimensional structures by ea.Hy926 endothelial cells. PLoS ONE 2013, 8, e64402. [Google Scholar] [CrossRef] [Green Version]
- Corydon, T.J.; Mann, V.; Slumstrup, L.; Kopp, S.; Sahana, J.; Askou, A.L.; Magnusson, N.E.; Echegoyen, D.; Bek, T.; Sundaresan, A.; et al. Reduced expression of cytoskeletal and extracellular matrix genes in human adult retinal pigment epithelium cells exposed to simulated microgravity. Cell. Physiol. Biochem. 2016, 40, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Berrios, D.C.; Galazka, J.; Grigorev, K.; Gebre, S.; Costes, S.V. Nasa genelab: Interfaces for the exploration of space omics data. Nucleic Acids Res. 2021, 49, D1515–D1522. [Google Scholar] [CrossRef]
- Pletser, V.; Migeotte, P.F.; Legros, J.C.; Deneyer, B.; Caron, R. The suborbital research association: Using suborbital platforms for scientific and student experiments. Microgravity Sci. Technol. 2016, 28, 529–544. [Google Scholar] [CrossRef]
- von Kampen, P.; Kaczmarczik, U.; Rath, H.J. The new drop tower catapult system. Acta Astronaut. 2006, 59, 278–283. [Google Scholar] [CrossRef]
- Selig, H.; Dittus, H.; Lämmerzahl, C. Drop tower microgravity improvement towards the nano-g level for the microscope payload tests. Microgravity Sci. Technol. 2010, 22, 539–549. [Google Scholar] [CrossRef]
- Thompson, D.M. Zero Gravity Research Facility User’s Guide. Available online: https://ntrs.nasa.gov/archive/nasa/casi.ntrs.nasa.gov/20000017932.pdf (accessed on 14 November 2022).
- Bliumentalis, D. Zero Gravity Research Facility. Available online: https://www1.grc.nasa.gov/facilities/zero-g/ (accessed on 10 December 2022).
- Lotz, C.; Froböse, T.; Wanner, A.; Overmeyer, L.; Ertmer, W. Einstein-elevator: A new facility for research from μ to 5. Gravit. Space Res. 2017, 5, 11–27. [Google Scholar] [CrossRef]
- Sabbatini, M. Parabolic Flights. Available online: http://wsn.spaceflight.esa.int/docs/EUG2LGPr3/EUG2LGPr3-5-ParabolicFlights.pdf (accessed on 15 December 2022).
- Sabbatini, M. Sounding Rockets. Available online: http://wsn.spaceflight.esa.int/docs/EUG2LGPr3/EUG2LGPr3-6-SoundingRockets.pdf (accessed on 15 December 2022).
- Wagner, E.B. Research flights on blue origin’s new shepard. Gravit. Space Res. 2021, 9, 62–67. [Google Scholar] [CrossRef]
- Kirschnick, U.; Agricola, H.; Horn, E. Effects of altered gravity on identified peptidergic neurons of the cricket Acheta Domesticus. Gravit. Space Res. 2006, 19, 135–136. [Google Scholar]
- Colucci, S.; Colaianni, G.; Brunetti, G.; Ferranti, F.; Mascetti, G.; Mori, G.; Grano, M. Irisin prevents microgravity-induced impairment of osteoblast differentiation in vitro during the space flight crs-14 mission. FASEB J. 2020, 34, 10096–10106. [Google Scholar] [CrossRef] [PubMed]
- Ceglia, E. European Users Guide to Low Gravity Platforms; Erasmus User Center and Communication Office, ESA: Noordwijk, The Netherlands, 2005; Volume 1, p. 21. [Google Scholar]
- Hu, W.; Kang, Q.; Duan, E.; Long, M. SJ-10 Recoverable Satellite for Space Microgravity Experiments. Chin. J. Space Sci. 2020, 40, 648–654. [Google Scholar] [CrossRef]
- Eigenbrod, C.; Könemann, T.; Kaczmarczik, U. Drop Tower Bremen—User’s Guide. Available online: https://www.zarm.uni-bremen.de/fileadmin/user_upload/drop_tower/Users_Manual_0412.pdf (accessed on 16 November 2022).
- Luo, L.; Zhou, H.; Sun, Y.; Zhang, W.; Chen, T.; Chen, S.; Wen, Y.; Xu, S.; Yu, S.; Liu, Y.C. Tsinghua university freefall facility (tuff): A 2.2 second drop tunnel for microgravity research. Microgravity Sci. Technol. 2021, 33, 26. [Google Scholar] [CrossRef]
- Nikhil, V.V.; Nair, A.; Niketh, P.; Kumar, A.; Muruganandam, M.N. The 2.5 s microgravity drop tower at national centre for combustion research and development (nccrd), indian institute of technology madras. Microgravity Sci. Technol. 2018, 30, 663–673. [Google Scholar] [CrossRef]
- Zhang, J.; Dong, W.; Wang, Z.; Zhang, Y.; Zhang, X.; Cai, Z.; Ma, W.; Li, Y.; Li, X.; Cui, X. Development of a new microgravity experiment facility with electromagnetic launch. Microgravity Sci. Technol. 2021, 33, 68. [Google Scholar] [CrossRef]
- Pletser, V.; Rouquette, S.; Friedrich, U.; Clervoy, J.-F.; Gharib, T.; Gai, F.; Mora, C. The first european parabolic flight campaign with the airbus a310 zero-g. Microgravity Sci. Technol. 2016, 28, 587–601. [Google Scholar] [CrossRef]
- TASS. Russia Starts Work on Bion-m Biosatellite to Deliver Mice into Outer Space. Available online: https://tass.com/science/1259627 (accessed on 18 June 2021).
- Poghosyan, A.; Golkar, A. Cubesat evolution: Analyzing cubesat capabilities for conducting science missions. Prog. Aerosp. Sci. 2017, 88, 59–83. [Google Scholar] [CrossRef]
- Padgen, M.R.; Liddell, L.C.; Bhardwaj, S.R.; Gentry, D.; Marina, D.; Parra, M.; Boone, T.; Tan, M.; Ellingson, L.; Rademacher, A.; et al. Biosentinel: A biofluidic nanosatellite monitoring microbial growth and activity in deep space. Astrobiology 2021. online ahead of print. [Google Scholar] [CrossRef]
- Massaro Tieze, S.; Liddell, L.C.; Santa Maria, S.R.; Bhattacharya, S. Biosentinel: A biological cubesat for deep space exploration. Astrobiology 2020. online ahead of print. [Google Scholar] [CrossRef] [Green Version]
- Thirsk, R.; Kuipers, A.; Mukai, C.; Williams, D. The space-flight environment: The International Space Station and beyond. CMAJ 2009, 180, 1216–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shiba, D.; Mizuno, H.; Yumoto, A.; Shimomura, M.; Kobayashi, H.; Morita, H.; Shimbo, M.; Hamada, M.; Kudo, T.; Shinohara, M.; et al. Development of new experimental platform ‘mars’—Multiple artificial-gravity research system—To elucidate the impacts of micro/partial gravity on mice. Sci. Rep. 2017, 7, 10837. [Google Scholar] [CrossRef] [Green Version]
- United Nations Office for Outer Space Affairs. Selected Experiment Projects to Be Executed on Board the css for the 1st Cycle. Available online: http://www.unoosa.org/documents/doc/psa/hsti/CSS_1stAO/1stAO_FinSelResults.pdf (accessed on 17 June 2021).
- Wuest, S.L.; Richard, S.; Kopp, S.; Grimm, D.; Egli, M. Simulated microgravity: Critical review on the use of random positioning machines for mammalian cell culture. BioMed Res. Int. 2015, 2015, 971474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.M.; de Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.A.; Lebert, M.; et al. Ground-based facilities for simulation of microgravity: Organism-specific recommendations for their use, and recommended terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dedolph, R.R.; Dipert, M.H. The physical basis of gravity stimulus nullification by clinostat rotation 1. Plant Physiol. 1971, 47, 756–764. [Google Scholar] [CrossRef]
- Sabbatini, M. Ground Based Facilities. Available online: http://wsn.spaceflight.esa.int/docs/EUG2LGPr3/EUG2LGPr3-3-GroundBased.pdf (accessed on 15 December 2022).
- Schwarz, R.P.; Goodwin, T.J.; Wolf, D.A. Cell culture for three-dimensional modeling in rotating-wall vessels: An application of simulated microgravity. J. Tissue Cult. Methods 1992, 14, 51–57. [Google Scholar] [CrossRef]
- Acres, J.M.; Youngapelian, M.J.; Nadeau, J. The influence of spaceflight and simulated microgravity on bacterial motility and chemotaxis. NPJ Microgravity 2021, 7, 7. [Google Scholar] [CrossRef]
- Duscher, A.A.; Conesa, A.; Bishop, M.; Vroom, M.M.; Zubizarreta, S.D.; Foster, J.S. Transcriptional profiling of the mutualistic bacterium vibrio fischeri and an hfq mutant under modeled microgravity. NPJ Microgravity 2018, 4, 25. [Google Scholar] [CrossRef] [Green Version]
- Topolski, C.; Divo, E.; Li, X.; Hicks, J.; Chavez, A.; Castillo, H. Phenotypic and transcriptional changes in escherichia coli k12 in response to simulated microgravity on the eaglestat, a new 2d microgravity analog for bacterial studies. Life Sci. Space Res. 2022, 34, 1–8. [Google Scholar] [CrossRef]
- Kalichamy, S.S. A simple protocol to analyze the effects of simulated microgravity on nematodes. Biol. Bull. Russ. Acad. Sci. 2021, 48, S22–S33. [Google Scholar] [CrossRef]
- Anil-Inevi, M.; Yilmaz, E.; Sarigil, O.; Tekin, H.C.; Ozcivici, E. Single cell densitometry and weightlessness culture of mesenchymal stem cells using magnetic levitation. In Stem Cell Nanotechnology: Methods and Protocols; Turksen, K., Ed.; Springer: New York, NY, USA, 2020; pp. 15–25. [Google Scholar]
- Herranz, R.; Larkin, O.J.; Dijkstra, C.E.; Hill, R.J.A.; Anthony, P.; Davey, M.R.; Eaves, L.; van Loon, J.J.W.A.; Medina, F.J.; Marco, R. Microgravity simulation by diamagnetic levitation: Effects of a strong gradient magnetic field on the transcriptional profile of drosophila melanogaster. BMC Genom. 2012, 13, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruhaug, G.B.L. Diamagnetic Levitation Using High-Temperature Superconducting Wires for Microgravity Research and Mitigation in Human Spaceflight Applications. Available online: http://arxiv.org/pdf/2004.09683v1 (accessed on 20 December 2022).
- Globus, R.K.; Morey-Holton, E. Hindlimb unloading: Rodent analog for microgravity. J. Appl. Physiol. 2016, 120, 1196–1206. [Google Scholar] [CrossRef] [PubMed]
- Pavy-Le Traon, A.; Heer, M.; Narici, M.V.; Rittweger, J.; Vernikos, J. From space to earth: Advances in human physiology from 20 years of bed rest studies (1986–2006). Eur. J. Appl. Physiol. 2007, 101, 143–194. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, R.; Qaisar, R.; Goswami, N.; Khan, N.A.; Elmoselhi, A. Effect of microgravity environment on gut microbiome and angiogenesis. Life 2021, 11, 1008. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, R.; Akbar, N.; Khan, N.A. Gut microbiome and human health under the space environment. J. Appl. Microbiol. 2021, 130, 14–24. [Google Scholar] [CrossRef]
- Vemuri, R.; Shankar, E.M.; Chieppa, M.; Eri, R.; Kavanagh, K. Beyond just bacteria: Functional biomes in the gut ecosystem including virome, mycobiome, archaeome and helminths. Microorganisms 2020, 8, 483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senatore, G.; Mastroleo, F.; Leys, N.; Mauriello, G. Growth of lactobacillus reuteri dsm17938 under two simulated microgravity systems: Changes in reuterin production, gastrointestinal passage resistance, and stress genes expression response. Astrobiology 2020, 20, 1–14. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, Y.; Guo, J.; Kang, L.; Deng, Y.; Li, Y. Investigation on rat intestinal homeostasis alterations induced by 7-day simulated microgravity effect based on a proteomic approach. Acta Astronaut. 2020, 166, 560–566. [Google Scholar] [CrossRef]
- Shama, S.; Qaisar, R.; Khan, N.A.; Tauseef, I.; Siddiqui, R. The role of 4-phenylbutyric acid in gut microbial dysbiosis in a mouse model of simulated microgravity. Life 2022, 12, 1301. [Google Scholar] [CrossRef]
- Cui, J.; Yi, Z.; Fu, Y.; Liu, H. Simulated microgravity shapes the endophytic bacterial community by affecting wheat root metabolism. Environ. Microbiol. 2022, 24, 3355–3368. [Google Scholar] [CrossRef]
- Green, M.J.; Aylott, J.W.; Williams, P.; Ghaemmaghami, A.M.; Williams, P.M. Immunity in space: Prokaryote adaptations and immune response in microgravity. Life 2021, 11, 112. [Google Scholar] [CrossRef]
- Bai, P.; Zhang, B.; Zhao, X.; Li, D.; Yu, Y.; Zhang, X.; Huang, B.; Liu, C. Decreased metabolism and increased tolerance to extreme environments in staphylococcus warneri during long-term spaceflight. MicrobiologyOpen 2019, 8, e917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Bai, P.; Zhao, X.; Yu, Y.; Zhang, X.; Li, D.; Liu, C. Increased growth rate and amikacin resistance of salmonella enteritidis after one-month spaceflight on china’s shenzhou-11 spacecraft. MicrobiologyOpen 2019, 8, e00833. [Google Scholar] [CrossRef] [Green Version]
- Crucian, B.E.; Choukèr, A.; Simpson, R.J.; Mehta, S.; Marshall, G.; Smith, S.M.; Zwart, S.R.; Heer, M.; Ponomarev, S.; Whitmire, A.; et al. Immune system dysregulation during spaceflight: Potential countermeasures for deep space exploration missions. Front. Immunol. 2018, 9, 1437. [Google Scholar] [CrossRef] [PubMed]
- Bigley, A.B.; Agha, N.H.; Baker, F.L.; Spielmann, G.; Kunz, H.E.; Mylabathula, P.L.; Rooney, B.V.; Laughlin, M.S.; Mehta, S.K.; Pierson, D.L.; et al. Nk cell function is impaired during long-duration spaceflight. J. Appl. Physiol. 2019, 126, 842–853. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Tian, H.; Wang, P.; Li, L.; Zhang, Z.; Zhang, J.; Zhao, Y. Spaceflight and simulated microgravity suppresses macrophage development via altered ras/erk/nfκb and metabolic pathways. Cell. Mol. Immunol. 2021, 18, 1489–1502. [Google Scholar] [CrossRef] [Green Version]
- Krieger, S.S.; Zwart, S.R.; Mehta, S.; Wu, H.; Simpson, R.J.; Smith, S.M.; Crucian, B. Alterations in saliva and plasma cytokine concentrations during long-duration spaceflight. Front. Immunol. 2021, 12, 725748. [Google Scholar] [CrossRef]
- Barrila, J.; Sarker, S.F.; Hansmeier, N.; Yang, S.; Buss, K.; Briones, N.; Park, J.; Davis, R.R.; Forsyth, R.J.; Ott, C.M.; et al. Evaluating the effect of spaceflight on the host–pathogen interaction between human intestinal epithelial cells and salmonella typhimurium. NPJ Microgravity 2021, 7, 9. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Y.; Wang, L.; Li, Y.; Song, Y.; Yang, R.; Han, Y. Evaluation of pathogenesis and biofilm formation ability of yersinia pestis after 40-day exposure to simulated microgravity. Int. J. Astrobiol. 2022, 21, 96–109. [Google Scholar] [CrossRef]
- Morrison, M.D.; Thissen, J.B.; Karouia, F.; Mehta, S.; Urbaniak, C.; Venkateswaran, K.; Smith, D.J.; Jaing, C. Investigation of spaceflight induced changes to astronaut microbiomes. Front. Microbiol. 2021, 12, 659179. [Google Scholar] [CrossRef]
- Prasad, B.; Richter, P.; Vadakedath, N.; Haag, F.W.M.; Strauch, S.M.; Mancinelli, R.; Schwarzwälder, A.; Etcheparre, E.; Gaume, N.; Lebert, M. How the space environment influences organisms: An astrobiological perspective and review. Int. J. Astrobiol. 2021, 20, 159–177. [Google Scholar] [CrossRef]
- Prasad, B.; Richter, P.; Vadakedath, N.; Mancinelli, R.; Krüger, M.; Strauch, S.M.; Grimm, D.; Darriet, P.; Chapel, J.-P.; Cohen, J.; et al. Exploration of space to achieve scientific breakthroughs. Biotechnol. Adv. 2020, 43, 107572. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, R.; Torres, M.; Clemens, R.; Hateley, S.; Hosamani, R.; Wade, W.; Bhattacharya, S. Spaceflight and simulated microgravity conditions increase virulence of serratia marcescens in the drosophila melanogaster infection model. NPJ Microgravity 2020, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Sheet, S.; Yesupatham, S.; Ghosh, K.; Choi, M.-S.; Shim, K.S.; Lee, Y.S. Modulatory effect of low-shear modeled microgravity on stress resistance, membrane lipid composition, virulence, and relevant gene expression in the food-borne pathogen listeria monocytogenes. Enzym. Microb. Technol. 2020, 133, 109440. [Google Scholar] [CrossRef] [PubMed]
- Fajardo-Cavazos, P.; Leehan, J.D.; Nicholson, W.L. Alterations in the spectrum of spontaneous rifampicin-resistance mutations in the bacillus subtilis rpob gene after cultivation in the human spaceflight environment. Front. Microbiol. 2018, 9, 192. [Google Scholar] [CrossRef] [Green Version]
- Tirumalai, M.R.; Karouia, F.; Tran, Q.; Stepanov, V.G.; Bruce, R.J.; Ott, C.M.; Pierson, D.L.; Fox, G.E. Evaluation of acquired antibiotic resistance in Escherichia coli exposed to long-term low-shear modeled microgravity and background antibiotic exposure. mBio 2019, 10, e02637-18. [Google Scholar] [CrossRef] [Green Version]
- Prasad, B.; Grimm, D.; Strauch, S.M.; Erzinger, G.S.; Corydon, T.J.; Lebert, M.; Magnusson, N.E.; Infanger, M.; Richter, P.; Krüger, M. Influence of microgravity on apoptosis in cells, tissues, and other systems in vivo and in vitro. Int. J. Mol. Sci. 2020, 21, 9373. [Google Scholar] [CrossRef]
- Urbaniak, C.; Grams, T.; Mason, C.E.; Venkateswaran, K. Simulated microgravity promotes horizontal gene transfer of antimicrobial resistance genes between bacterial genera in the absence of antibiotic selective pressure. Life 2021, 11, 960. [Google Scholar] [CrossRef]
- Sharma, G.; Curtis, P.D. The impacts of microgravity on bacterial metabolism. Life 2022, 12, 774. [Google Scholar] [CrossRef]
- Vroom, M.M.; Rodriguez-Ocasio, Y.; Lynch, J.B.; Ruby, E.G.; Foster, J.S. Modeled microgravity alters lipopolysaccharide and outer membrane vesicle production of the beneficial symbiont vibrio fischeri. NPJ Microgravity 2021, 7, 8. [Google Scholar] [CrossRef]
- Tesei, D.; Chiang, A.J.; Kalkum, M.; Stajich, J.E.; Mohan, G.B.M.; Sterflinger, K.; Venkateswaran, K. Effects of simulated microgravity on the proteome and secretome of the polyextremotolerant black fungus knufia chersonesos. Front. Genet. 2021, 12, 638708. [Google Scholar] [CrossRef]
- Cordero, R.J.B.; Dragotakes, Q.; Friello, P.J.; Casadevall, A. Melanin protects cryptococcus neoformans from spaceflight effects. Environ. Microbiol. Rep. 2022, 14, 679–685. [Google Scholar] [CrossRef]
- Thombre, R.; Shinde, V.; Dixit, J.; Jagtap, S.; Vidyasagar, P.B. Response of extreme haloarchaeon haloarcula argentinensis rr10 to simulated microgravity in clinorotation. 3 Biotech 2017, 7, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Agostino, M.; Babin, A.-L.; Zaffino, M.; Frippiat, J.-P.; Machouart, M.; Debourgogne, A. Simulated microgravity created using a random positioning machine induces changes in the physiology of the fusarium solani species complex. Microorganisms 2022, 10, 2270. [Google Scholar] [CrossRef]
- Wang, J.; Han, C.; Lu, Z.; Ge, P.; Cui, Y.; Zhao, D.; Yang, X.; Wu, B.; Qiang, L.; Zhang, Y.; et al. Simulated microgravity suppresses mapk pathway-mediated innate immune response to bacterial infection and induces gut microbiota dysbiosis. FASEB J. 2020, 34, 14631–14644. [Google Scholar] [CrossRef] [PubMed]
- Walls, S.; Diop, S.; Birse, R.; Elmen, L.; Gan, Z.; Kalvakuri, S.; Pineda, S.; Reddy, C.; Taylor, E.; Trinh, B.; et al. Prolonged exposure to microgravity reduces cardiac contractility and initiates remodeling in drosophila. Cell Rep. 2020, 33, 108445. [Google Scholar] [CrossRef]
- Vroom, M.M.; Troncoso-Garcia, A.; Duscher, A.A.; Foster, J.S. Modeled microgravity alters apoptotic gene expression and caspase activity in the squid-vibrio symbiosis. BMC Microbiol. 2022, 22, 202. [Google Scholar] [CrossRef] [PubMed]
- Higashibata, A.; Hashizume, T.; Nemoto, K.; Higashitani, N.; Etheridge, T.; Mori, C.; Harada, S.; Sugimoto, T.; Szewczyk, N.J.; Baba, S.A.; et al. Microgravity elicits reproducible alterations in cytoskeletal and metabolic gene and protein expression in space-flown caenorhabditis elegans. NPJ Microgravity 2016, 2, 15022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higashitani, A.; Hashizume, T.; Takiura, M.; Higashitani, N.; Teranishi, M.; Oshima, R.; Yano, S.; Kuriyama, K.; Higashibata, A. Histone deacetylase hda-4-mediated epigenetic regulation in space-flown c. Elegans. NPJ Microgravity 2021, 7, 33. [Google Scholar] [CrossRef]
- Liu, H.; Guo, D.; Kong, Y.; Rui, Q.; Wang, D. Damage on functional state of intestinal barrier by microgravity stress in nematode caenorhabditis elegans. Ecotoxicol. Environ. Saf. 2019, 183, 109554. [Google Scholar] [CrossRef]
- Kong, Y.; Liu, H.; Li, W.; Wang, D. Intestine-specific activity of insulin signaling pathway in response to microgravity stress in caenorhabditis elegans. Biochem. Biophys. Res. Commun. 2019, 517, 278–284. [Google Scholar] [CrossRef]
- Hong, X.; Ratri, A.; Choi, S.Y.; Tash, J.S.; Ronca, A.E.; Alwood, J.S.; Christenson, L.K. Effects of spaceflight aboard the international space station on mouse estrous cycle and ovarian gene expression. NPJ Microgravity 2021, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Horie, K.; Kato, T.; Kudo, T.; Sasanuma, H.; Miyauchi, M.; Akiyama, N.; Miyao, T.; Seki, T.; Ishikawa, T.; Takakura, Y.; et al. Impact of spaceflight on the murine thymus and mitigation by exposure to artificial gravity during spaceflight. Sci. Rep. 2019, 9, 19866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horie, K.; Sasanuma, H.; Kudo, T.; Fujita, S.-I.; Miyauchi, M.; Miyao, T.; Seki, T.; Akiyama, N.; Takakura, Y.; Shimbo, M.; et al. Down-regulation of gata1-dependent erythrocyte-related genes in the spleens of mice exposed to a space travel. Sci. Rep. 2019, 9, 7654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, Y.; Higashibata, A.; Matsunaga, Y.; Yonezawa, Y.; Kawano, T.; Higashitani, A.; Kuriyama, K.; Shimazu, T.; Tanaka, M.; Szewczyk, N.J.; et al. Genes down-regulated in spaceflight are involved in the control of longevity in caenorhabditis elegans. Sci. Rep. 2012, 2, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guéguinou, N.; Jeandel, J.; Kaminski, S.; Baatout, S.; Ghislin, S.; Frippiat, J.-P. Modulation of iberian ribbed newt complement component c3 by stressors similar to those encountered during a stay onboard the international space station. Int. J. Mol. Sci. 2019, 20, 1579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mader, T.H.; Gibson, C.R.; Pass, A.F.; Kramer, L.A.; Lee, A.G.; Fogarty, J.; Tarver, W.J.; Dervay, J.P.; Hamilton, D.R.; Sargsyan, A.; et al. Optic disc edema, globe flattening, choroidal folds, and hyperopic shifts observed in astronauts after long-duration space flight. Ophthalmology 2011, 118, 2058–2069. [Google Scholar] [CrossRef] [Green Version]
- Cialdai, F.; Bolognini, D.; Vignali, L.; Iannotti, N.; Cacchione, S.; Magi, A.; Balsamo, M.; Vukich, M.; Neri, G.; Donati, A.; et al. Effect of space flight on the behavior of human retinal pigment epithelial arpe-19 cells and evaluation of coenzyme q10 treatment. Cell. Mol. Life Sci. 2021, 78, 7795–7812. [Google Scholar] [CrossRef]
- Chakravarty, K.; Lai, S.-H.; Boyko, V.; Chen, R.; Dueck, S.; Berrios, D.C.; Fogle, H.; Marcu, O.; Timucin, L.; Reinsch, S.; et al. Genelab: Multi-Omics Investigation of Rodent Research-1 Bio-Banked Tissues. Available online: https://ntrs.nasa.gov/citations/20170000299 (accessed on 1 November 2022).
- Mao, X.W.; Sandberg, L.B.; Gridley, D.S.; Herrmann, E.C.; Zhang, G.; Raghavan, R.; Zubarev, R.A.; Zhang, B.; Stodieck, L.S.; Ferguson, V.L.; et al. Proteomic analysis of mouse brain subjected to spaceflight. Int. J. Mol. Sci. 2018, 20, 7. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.W.; Nishiyama, N.C.; Byrum, S.D.; Stanbouly, S.; Jones, T.; Drew, A.; Sridharan, V.; Boerma, M.; Tackett, A.J.; Zawieja, D.; et al. Characterization of mouse ocular response to a 35-day spaceflight mission: Evidence of blood-retinal barrier disruption and ocular adaptations. Sci. Rep. 2019, 9, 8215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overbey, E.G.; da Silveira, W.A.; Stanbouly, S.; Nishiyama, N.C.; Roque-Torres, G.D.; Pecaut, M.J.; Zawieja, D.C.; Wang, C.; Willey, J.S.; Delp, M.D.; et al. Spaceflight influences gene expression, photoreceptor integrity, and oxidative stress-related damage in the murine retina. Sci. Rep. 2019, 9, 13304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Stanbouly, S.; Nishiyama, N.C.; Chen, X.; Delp, M.D.; Qiu, H.; Mao, X.W.; Wang, C. Spaceflight decelerates the epigenetic clock orchestrated with a global alteration in DNA methylome and transcriptome in the mouse retina. Precis. Clin. Med. 2021, 4, 93–108. [Google Scholar] [CrossRef] [PubMed]
- Sharma, T.P.; Miller, R.C.S.; ShahulHameed, S.P.; Kubik, A.; Allen, N.; Altinok, S.; Oswalt, L.E.; Sanchez-Hodge, R.; Schisler, J.C.; Blaber, E.; et al. Spaceflight-associated mirnas alters expression of targeted genes in ocular and spinal tissue under simulated spaceflight conditions. Investig. Ophthalmol. Vis. Sci. 2022, 63, 1177-A0031. [Google Scholar]
- Laurie, S.S.; Vizzeri, G.; Taibbi, G.; Ferguson, C.R.; Hu, X.; Lee, S.M.C.; Ploutz-Snyder, R.; Smith, S.M.; Zwart, S.R.; Stenger, M.B. Effects of short-term mild hypercapnia during head-down tilt on intracranial pressure and ocular structures in healthy human subjects. Physiol. Rep. 2017, 5, e13302. [Google Scholar] [CrossRef]
- Zwart, S.R.; Laurie, S.S.; Chen, J.J.; Macias, B.R.; Lee, S.M.C.; Stenger, M.; Grantham, B.; Carey, K.; Young, M.; Smith, S.M. Association of genetics and b vitamin status with the magnitude of optic disc edema during 30-day strict head-down tilt bed rest. JAMA Ophthalmol. 2019, 137, 1195–1200. [Google Scholar] [CrossRef]
- Barravecchia, I.; De Cesari, C.; Forcato, M.; Scebba, F.; Pyankova, O.V.; Bridger, J.M.; Foster, H.A.; Signore, G.; Borghini, A.; Andreassi, M.; et al. Microgravity and space radiation inhibit autophagy in human capillary endothelial cells, through either opposite or synergistic effects on specific molecular pathways. Cell. Mol. Life Sci. 2021, 79, 28. [Google Scholar] [CrossRef]
- Versari, S.; Longinotti, G.; Barenghi, L.; Maier, J.A.; Bradamante, S. The challenging environment on board the international space station affects endothelial cell function by triggering oxidative stress through thioredoxin interacting protein overexpression: The esa-sphinx experiment. FASEB J. 2013, 27, 4466–4475. [Google Scholar] [CrossRef] [PubMed]
- Cazzaniga, A.; Locatelli, L.; Castiglioni, S.; Maier, J.A.M. The dynamic adaptation of primary human endothelial cells to simulated microgravity. FASEB J. 2019, 33, 5957–5966. [Google Scholar] [CrossRef]
- Liang, Y.; Wang, M.; Liu, Y.; Wang, C.; Takahashi, K.; Naruse, K. Meta-analysis-assisted detection of gravity-sensitive genes in human vascular endothelial cells. Front. Cell. Dev. Biol. 2021, 9, 689662. [Google Scholar] [CrossRef] [PubMed]
- Li, C.F.; Sun, J.X.; Gao, Y.; Shi, F.; Pan, Y.K.; Wang, Y.C.; Sun, X.Q. Clinorotation-induced autophagy via hdm2-p53-mtor pathway enhances cell migration in vascular endothelial cells. Cell. Death Dis. 2018, 9, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, Y.K.; Li, C.F.; Gao, Y.; Wang, Y.C.; Sun, X.Q. Effect of mir-27b-5p on apoptosis of human vascular endothelial cells induced by simulated microgravity. Apoptosis 2020, 25, 73–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, D.; Guo, Y.B.; Zhang, M.; Sun, Y.Q. The subsequent biological effects of simulated microgravity on endothelial cell growth in huvecs. Chin. J. Traumatol. 2018, 21, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Kasiviswanathan, D.; Chinnasamy Perumal, R.; Bhuvaneswari, S.; Kumar, P.; Sundaresan, L.; Philip, M.; Puthenpurackal Krishnankutty, S.; Chatterjee, S. Interactome of mirnas and transcriptome of human umbilical cord endothelial cells exposed to short-term simulated microgravity. NPJ Microgravity 2020, 6, 18. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Shi, Y.; Qiu, C.; Zhao, J.; Gong, Y.; Nie, C.; Wu, B.; Yang, Y.; Wang, F.; Luo, L. Effects of simulated microgravity on ultrastructure and apoptosis of choroidal vascular endothelial cells. Front. Physiol. 2020, 11, 577325. [Google Scholar] [CrossRef] [PubMed]
- Dittrich, A.; Grimm, D.; Sahana, J.; Bauer, J.; Krüger, M.; Infanger, M.; Magnusson, N.E. Key proteins involved in spheroid formation and angiogenesis in endothelial cells after long-term exposure to simulated microgravity. Cell. Physiol. Biochem. 2018, 45, 429–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krüger, M.; Pietsch, J.; Bauer, J.; Kopp, S.; Carvalho, D.T.O.; Baatout, S.; Moreels, M.; Melnik, D.; Wehland, M.; Egli, M.; et al. Growth of endothelial cells in space and in simulated microgravity—A comparison on the secretory level. Cell. Physiol. Biochem. 2019, 52, 1039–1060. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Wang, C.; Sun, S.; Zhang, C.; Lü, D.; Chen, Q.; Long, M. Microgravity-induced alterations of inflammation-related mechanotransduction in endothelial cells on board sj-10 satellite. Front. Physiol. 2018, 9, 1025. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Wang, Y.; Wang, H.; Pan, Q.; Zuo, R.; Bai, S.; Zhang, X.; Lee, W.Y.; Kang, Q.; Li, G. Conditioned media from endothelial progenitor cells cultured in simulated microgravity promote angiogenesis and bone fracture healing. Stem Cell Res. Ther. 2021, 12, 47. [Google Scholar] [CrossRef] [Green Version]
- Girn, H.R.; Orsi, N.M.; Homer-Vanniasinkam, S. An overview of cytokine interactions in atherosclerosis and implications for peripheral arterial disease. Vasc. Med. 2007, 12, 299–309. [Google Scholar] [CrossRef] [Green Version]
- Grosse, J.; Wehland, M.; Pietsch, J.; Schulz, H.; Saar, K.; Hübner, N.; Elles, C.; Bauer, J.; Abou-El-Ardat, K.; Baatout, S.; et al. Gravity-sensitive signaling drives 3-dimensional formation of multicellular thyroid cancer spheroids. FASEB J. 2012, 26, 5124–5140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janmaleki, M.; Pachenari, M.; Seyedpour, S.M.; Shahghadami, R.; Sanati-Nezhad, A. Impact of simulated microgravity on cytoskeleton and viscoelastic properties of endothelial cell. Sci. Rep. 2016, 6, 32418. [Google Scholar] [CrossRef]
- Rudimov, E.G.; Buravkova, L.B. Endothelial gravisensitivity: The role of cytoskeleton and adhesion molecules. Fiziol. Cheloveka 2016, 42, 116–123. [Google Scholar] [CrossRef] [Green Version]
- Locatelli, L.; Cazzaniga, A.; De Palma, C.; Castiglioni, S.; Maier, J.A.M. Mitophagy contributes to endothelial adaptation to simulated microgravity. FASEB J. 2020, 34, 1833–1845. [Google Scholar] [CrossRef] [PubMed]
- Morbidelli, L.; Monici, M.; Marziliano, N.; Cogoli, A.; Fusi, F.; Waltenberger, J.; Ziche, M. Simulated hypogravity impairs the angiogenic response of endothelium by up-regulating apoptotic signals. Biochem. Biophys. Res. Commun. 2005, 334, 491–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, N.P.; Hui, T.T.; Ma, J.; Mei, Q.B. Effects of mir-503-5p on apoptosis of human pulmonary microvascular endothelial cells in simulated microgravity. J. Cell. Biochem. 2019, 120, 727–737. [Google Scholar] [CrossRef]
- Cogoli, A.; Tschopp, A. Lymphocyte reactivity during spaceflight. Immunol. Today 1985, 6, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Cogoli, A. Hematological and immunological changes during space flight. Acta Astronaut. 1981, 8, 995–1002. [Google Scholar] [CrossRef]
- Calcagno, G.; Jeandel, J.; Frippiat, J.-P.; Kaminski, S. Simulated Microgravity Disrupts Nuclear Factor kB Signaling and Impairs Murine Dendritic Cell Phenotype and Function. Int. J. Mol. Sci. 2023, 24, 1720. [Google Scholar] [CrossRef]
- Bisserier, M.; Shanmughapriya, S.; Rai, A.K.; Gonzalez, C.; Brojakowska, A.; Garikipati, V.N.S.; Madesh, M.; Mills, P.J.; Walsh, K.; Arakelyan, A.; et al. Cell-free mitochondrial DNA as a potential biomarker for astronauts’ health. J. Am. Heart Assoc. 2021, 10, e022055. [Google Scholar] [CrossRef] [PubMed]
- Brojakowska, A.; Kour, A.; Thel, M.C.; Park, E.; Bisserier, M.; Garikipati, V.N.S.; Hadri, L.; Mills, P.J.; Walsh, K.; Goukassian, D.A. Retrospective analysis of somatic mutations and clonal hematopoiesis in astronauts. Commun. Biol. 2022, 5, 828. [Google Scholar] [CrossRef]
- Bezdan, D.; Grigorev, K.; Meydan, C.; Pelissier Vatter, F.A.; Cioffi, M.; Rao, V.; MacKay, M.; Nakahira, K.; Burnham, P.; Afshinnekoo, E.; et al. Cell-free DNA (cfdna) and exosome profiling from a year-long human spaceflight reveals circulating biomarkers. iScience 2020, 23, 101844. [Google Scholar] [CrossRef]
- Gertz, M.L.; Chin, C.R.; Tomoiaga, D.; MacKay, M.; Chang, C.; Butler, D.; Afshinnekoo, E.; Bezdan, D.; Schmidt, M.A.; Mozsary, C.; et al. Multi-omic, single-cell, and biochemical profiles of astronauts guide pharmacological strategies for returning to gravity. Cell Rep. 2020, 33, 108429. [Google Scholar] [CrossRef]
- Schmidt, M.A.; Meydan, C.; Schmidt, C.M.; Afshinnekoo, E.; Mason, C.E. The nasa twins study: The effect of one year in space on long-chain fatty acid desaturases and elongases. Lifestyle Genom. 2020, 13, 107–121. [Google Scholar] [CrossRef] [Green Version]
- Thiel, C.S.; Christoffel, S.; Tauber, S.; Vahlensieck, C.; Zélicourt, D.; Layer, L.E.; Lauber, B.; Polzer, J.; Ullrich, O. Rapid cellular perception of gravitational forces in human jurkat t cells and transduction into gene expression regulation. Int. J. Mol. Sci. 2020, 21, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, J.; Thiel, C.S.; Tauber, S.; Stockmann, C.; Gassmann, M.; Ullrich, O. Expression of hypoxia-inducible factor 1α (hif-1α) and genes of related pathways in altered gravity. Int. J. Mol. Sci. 2019, 20, 436. [Google Scholar] [CrossRef] [PubMed]
- Vahlensieck, C.; Thiel, C.S.; Zhang, Y.; Huge, A.; Ullrich, O. Gravitational force-induced 3d chromosomal conformational changes are associated with rapid transcriptional response in human t cells. Int. J. Mol. Sci. 2021, 22, 9426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tauber, S.; Christoffel, S.; Thiel, C.S.; Ullrich, O. Transcriptional homeostasis of oxidative stress-related pathways in altered gravity. Int. J. Mol. Sci. 2018, 19, 2814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiel, C.S.; Hauschild, S.; Huge, A.; Tauber, S.; Lauber, B.A.; Polzer, J.; Paulsen, K.; Lier, H.; Engelmann, F.; Schmitz, B.; et al. Dynamic gene expression response to altered gravity in human t cells. Sci. Rep. 2017, 7, 5204. [Google Scholar] [CrossRef] [Green Version]
- Pecaut, M.J.; Mao, X.W.; Bellinger, D.L.; Jonscher, K.R.; Stodieck, L.S.; Ferguson, V.L.; Bateman, T.A.; Mohney, R.P.; Gridley, D.S. Is spaceflight-induced immune dysfunction linked to systemic changes in metabolism? PLoS ONE 2017, 12, e0174174. [Google Scholar] [CrossRef]
- ElGindi, M.; Sapudom, J.; Laws, P.; Garcia-Sabaté, A.; Daqaq, M.F.; Teo, J. 3d microenvironment attenuates simulated microgravity-mediated changes in t cell transcriptome. Cell. Mol. Life Sci. 2022, 79, 508. [Google Scholar] [CrossRef]
- Ludtka, C.; Moore, E.; Allen, J.B. The effects of simulated microgravity on macrophage phenotype. Biomedicines 2021, 9, 1205. [Google Scholar] [CrossRef]
- Spatz, J.M.; Fulford, M.H.; Tsai, A.; Gaudilliere, D.; Hedou, J.; Ganio, E.; Angst, M.; Aghaeepour, N.; Gaudilliere, B. Human immune system adaptations to simulated microgravity revealed by single-cell mass cytometry. Sci. Rep. 2021, 11, 11872. [Google Scholar] [CrossRef]
- Yuan, M.; Liu, H.; Zhou, S.; Zhou, X.; Huang, Y.E.; Hou, F.; Jiang, W. Integrative analysis of regulatory module reveals associations of microgravity with dysfunctions of multi-body systems and tumorigenesis. Int. J. Mol. Sci. 2020, 21, 7585. [Google Scholar] [CrossRef]
- Rinaldi, S.; Meloni, M.A.; Galleri, G.; Maioli, M.; Pigliaru, G.; Cugia, G.; Santaniello, S.; Castagna, A.; Fontani, V. Radio electric asymmetric conveyer (reac) technology to obviate loss of t cell responsiveness under simulated microgravity. PLoS ONE 2018, 13, e0200128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Villanueva, M.; Feiveson, A.H.; Krieger, S.; Kay Brinda, A.; von Scheven, G.; Bürkle, A.; Crucian, B.; Wu, H. Synergistic effects of weightlessness, isoproterenol, and radiation on DNA damage response and cytokine production in immune cells. Int. J. Mol. Sci. 2018, 19, 3689. [Google Scholar] [CrossRef] [PubMed]
- Paul, A.M.; Overbey, E.G.; da Silveira, W.A.; Szewczyk, N.; Nishiyama, N.C.; Pecaut, M.J.; Anand, S.; Galazka, J.M.; Mao, X.W. Immunological and hematological outcomes following protracted low dose/low dose rate ionizing radiation and simulated microgravity. Sci. Rep. 2021, 11, 11452. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Nie, L.; Xie, S.; Li, M.; Zhu, C.; Qiu, X.; Kuang, J.; Liu, C.; Lu, C.; Li, W.; et al. Attenuation of antiviral immune response caused by perturbation of trim25-mediated rig-i activation under simulated microgravity. Cell Rep. 2021, 34, 108600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fitzgerald, J.; Endicott, J.; Hansen, U.; Janowitz, C. Articular cartilage and sternal fibrocartilage respond differently to extended microgravity. NPJ Microgravity 2019, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Li, D.X.; Chee, R.K.W.; Kunze, M.; Mulet-Sierra, A.; Sommerfeldt, M.; Westover, L.; Graf, D.; Adesida, A.B. Mechanical unloading of engineered human meniscus models under simulated microgravity: A transcriptomic study. Sci. Data 2022, 9, 736. [Google Scholar] [CrossRef]
- Ma, Z.; Li, D.X.; Kunze, M.; Mulet-Sierra, A.; Westover, L.; Adesida, A.B. Engineered human meniscus in modeling sex differences of knee osteoarthritis in vitro. Front. Bioeng. Biotechnol. 2022, 10, 823679. [Google Scholar] [CrossRef] [Green Version]
- Wuest, S.L.; Caliò, M.; Wernas, T.; Tanner, S.; Giger-Lange, C.; Wyss, F.; Ille, F.; Gantenbein, B.; Egli, M. Influence of mechanical unloading on articular chondrocyte dedifferentiation. Int. J. Mol. Sci. 2018, 19, 1289. [Google Scholar] [CrossRef]
- Wehland, M.; Steinwerth, P.; Aleshcheva, G.; Sahana, J.; Hemmersbach, R.; Lützenberg, R.; Kopp, S.; Infanger, M.; Grimm, D. Tissue engineering of cartilage using a random positioning machine. Int. J. Mol. Sci. 2020, 21, 9596. [Google Scholar] [CrossRef] [PubMed]
- Henrich, M.; Ha, P.; Wang, Y.; Ting, K.; Stodieck, L.; Soo, C.; Adams, J.S.; Chun, R. Alternative splicing diversifies the skeletal muscle transcriptome during prolonged spaceflight. Skelet. Muscle 2022, 12, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambara, G.; Salanova, M.; Ciciliot, S.; Furlan, S.; Gutsmann, M.; Schiffl, G.; Ungethuem, U.; Volpe, P.; Gunga, H.-C.; Blottner, D. Gene expression profiling in slow-type calf soleus muscle of 30 days space-flown mice. PLoS ONE 2017, 12, e0169314. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, T.; Kudo, T.; Fujita, R.; Fujita, S.-i.; Tsubouchi, H.; Fuseya, S.; Suzuki, R.; Hamada, M.; Okada, R.; Muratani, M.; et al. Nuclear factor e2-related factor 2 (nrf2) deficiency accelerates fast fibre type transition in soleus muscle during space flight. Commun. Biol. 2021, 4, 787. [Google Scholar] [CrossRef] [PubMed]
- Okada, R.; Fujita, S.-I.; Suzuki, R.; Hayashi, T.; Tsubouchi, H.; Kato, C.; Sadaki, S.; Kanai, M.; Fuseya, S.; Inoue, Y.; et al. Transcriptome analysis of gravitational effects on mouse skeletal muscles under microgravity and artificial 1 g onboard environment. Sci. Rep. 2021, 11, a9168. [Google Scholar] [CrossRef]
- Uda, Y.; Spatz, J.M.; Hussein, A.; Garcia, J.H.; Lai, F.; Dedic, C.; Fulzele, K.; Dougherty, S.; Eberle, M.; Adamson, C.; et al. Global transcriptomic analysis of a murine osteocytic cell line subjected to spaceflight. FASEB J. 2021, 35, e21578. [Google Scholar] [CrossRef] [Green Version]
- Gambacurta, A.; Merlini, G.; Ruggiero, C.; Diedenhofen, G.; Battista, N.; Bari, M.; Balsamo, M.; Piccirillo, S.; Valentini, G.; Mascetti, G.; et al. Human osteogenic differentiation in space: Proteomic and epigenetic clues to better understand osteoporosis. Sci. Rep. 2019, 9, 8343. [Google Scholar] [CrossRef]
- Li, L.; Zhang, C.; Chen, J.-l.; Hong, F.-f.; Chen, P.; Wang, J.-f. Effects of simulated microgravity on the expression profiles of rna during osteogenic differentiation of human bone marrow mesenchymal stem cells. Cell Prolif. 2019, 52, e12539. [Google Scholar] [CrossRef] [Green Version]
- Mann, V.; Grimm, D.; Corydon, T.J.; Krüger, M.; Wehland, M.; Riwaldt, S.; Sahana, J.; Kopp, S.; Bauer, J.; Reseland, J.E.; et al. Changes in human foetal osteoblasts exposed to the random positioning machine and bone construct tissue engineering. Int. J. Mol. Sci. 2019, 20, 1357. [Google Scholar] [CrossRef] [PubMed]
- Braveboy-Wagner, J.; Lelkes, P.I. Impairment of 7f2 osteoblast function by simulated partial gravity in a random positioning machine. NPJ Microgravity 2022, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Zhang, Y.; Wei, S.; Zhang, X.; Guo, Y.; Han, B. Comprehensive circrna expression profile and function network in osteoblast-like cells under simulated microgravity. Gene 2021, 764, 145106. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Bauer, J.; Wise, P.; Krüger, M.; Simonsen, U.; Wehland, M.; Infanger, M.; Corydon, T.J. The role of sox family members in solid tumours and metastasis. Semin. Cancer Biol. 2020, 67, 122–153. [Google Scholar] [CrossRef] [PubMed]
- Cortés-Sánchez, J.L.; Callant, J.; Krüger, M.; Sahana, J.; Kraus, A.; Baselet, B.; Infanger, M.; Baatout, S.; Grimm, D. Cancer studies under space conditions: Finding answers abroad. Biomedicines 2022, 10, 25. [Google Scholar] [CrossRef]
- Drago-Ferrante, R.; Di Fiore, R.; Karouia, F.; Subbannayya, Y.; Das, S.; Aydogan Mathyk, B.; Arif, S.; Guevara-Cerdán, A.P.; Seylani, A.; Galsinh, A.S.; et al. Extraterrestrial gynecology: Could spaceflight increase the risk of developing cancer in female astronauts? An updated review. Int. J. Mol. Sci. 2022, 23, 7465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, R.J.; Bukhtiyarov, I.V.; Tikhonova, G.I.; Day, S.M.; Ushakov, I.B.; Gorchakova, T.Y.U. Contrapositive logic suggests space radiation not having a strong impact on mortality of us astronauts and soviet and russian cosmonauts. Sci. Rep. 2019, 9, 8583. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, R.; Little, M.P.; Day, S.; Charvat, J.; Blattnig, S.; Huff, J.; Patel, Z.S. Cancer incidence and mortality in the USA astronaut corps, 1959–2017. Occup. Environ. Med. 2021, 78, 869–875. [Google Scholar] [CrossRef] [Green Version]
- Clark, O.H. Controversies in familial thyroid cancer 2014. Ulus. Cerrahi Derg. 2014, 30, 62–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bongarzone, I.; Fugazzola, L.; Vigneri, P.; Mariani, L.; Mondellini, P.; Pacini, F.; Basolo, F.; Pinchera, A.; Pilotti, S.; Pierotti, M.A. Age-related activation of the tyrosine kinase receptor protooncogenes ret and ntrk1 in papillary thyroid carcinoma. J. Clin. Endocrinol. Metab. 1996, 81, 2006–2009. [Google Scholar] [CrossRef]
- American Cancer Society. Key Statistics for Thyroid Cancer. Available online: https://www.Cancer.Org/cancer/thyroid-cancer/about/key-statistics.Html (accessed on 28 November 2022).
- Plakhuta-Plakutina, G.I.; Kabitskii, E.N.; Dmitrieva, N.P.; Amirkhanian, E.A. Studies of the morphology of the thyroid gland and thyroid hormone levels in the blood of rats in experiments on “kosmos-1667” and “kosmos-1887”. Kosm. Biol. Aviakosm Med. 1990, 24, 25–27. [Google Scholar] [Green Version]
- Bauer, J.; Kopp, S.; Schlagberger, E.M.; Grosse, J.; Sahana, J.; Riwaldt, S.; Wehland, M.; Luetzenberg, R.; Infanger, M.; Grimm, D. Proteome analysis of human follicular thyroid cancer cells exposed to the random positioning machine. Int. J. Mol. Sci. 2017, 18, 546. [Google Scholar] [CrossRef]
- Bauer, J.; Grimm, D.; Gombocz, E. Semantic analysis of thyroid cancer cell proteins obtained from rare research opportunities. J. Biomed. Inform. 2017, 76, 138–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauer, J.; Wehland, M.; Infanger, M.; Grimm, D.; Gombocz, E. Semantic analysis of posttranslational modification of proteins accumulated in thyroid cancer cells exposed to simulated microgravity. Int. J. Mol. Sci. 2018, 19, 2257. [Google Scholar] [CrossRef] [PubMed]
- Hou, G.; Xu, B.; Bi, Y.; Wu, C.; Ru, B.; Sun, B.; Bai, X. Recent advances in research on aspartate beta-hydroxylase (asph) in pancreatic cancer: A brief update. Bosn. J. Basic. Med. Sci. 2018, 18, 297–304. [Google Scholar] [CrossRef] [Green Version]
- Kopp, S.; Kruger, M.; Feldmann, S.; Oltmann, H.; Schutte, A.; Schmitz, B.; Bauer, J.; Schulz, H.; Saar, K.; Huebner, N.; et al. Thyroid cancer cells in space during the texus-53 sounding rocket mission—The thyroid project. Sci. Rep. 2018, 8, 10355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopp, S.; Kruger, M.; Bauer, J.; Wehland, M.; Corydon, T.J.; Sahana, J.; Nassef, M.Z.; Melnik, D.; Bauer, T.J.; Schulz, H.; et al. Microgravity affects thyroid cancer cells during the texus-53 mission stronger than hypergravity. Int. J. Mol. Sci. 2018, 19, 4001. [Google Scholar] [CrossRef] [Green Version]
- Nassef, M.Z.; Kopp, S.; Wehland, M.; Melnik, D.; Sahana, J.; Krüger, M.; Corydon, T.J.; Oltmann, H.; Schmitz, B.; Schütte, A.; et al. Real microgravity influences the cytoskeleton and focal adhesions in human breast cancer cells. Int. J. Mol. Sci. 2019, 20, 3156. [Google Scholar] [CrossRef]
- Wise, P.M.; Neviani, P.; Riwaldt, S.; Corydon, T.J.; Wehland, M.; Braun, M.; Kruger, M.; Infanger, M.; Grimm, D. Changes in exosome release in thyroid cancer cells after prolonged exposure to real microgravity in space. Int. J. Mol. Sci. 2021, 22, 2132. [Google Scholar] [CrossRef]
- Melnik, D.; Kruger, M.; Schulz, H.; Kopp, S.; Wehland, M.; Bauer, J.; Baselet, B.; Vermeesen, R.; Baatout, S.; Corydon, T.J.; et al. The cellbox-2 mission to the international space station: Thyroid cancer cells in space. Int. J. Mol. Sci. 2021, 22, 8777. [Google Scholar] [CrossRef] [Green Version]
- Melnik, D.; Sahana, J.; Corydon, T.J.; Kopp, S.; Nassef, M.Z.; Wehland, M.; Infanger, M.; Grimm, D.; Kruger, M. Dexamethasone inhibits spheroid formation of thyroid cancer cells exposed to simulated microgravity. Cells 2020, 9, 367. [Google Scholar] [CrossRef] [PubMed]
- Wise, P.M.; Neviani, P.; Riwaldt, S.; Corydon, T.J.; Wehland, M.; Braun, M.; Krüger, M.; Infanger, M.; Grimm, D. Changes in exosomal mirna composition in thyroid cancer cells after prolonged exposure to real microgravity in space. Int. J. Mol. Sci. 2021, 22, 12841. [Google Scholar] [CrossRef]
- Melnik, D.; Cortés-Sánchez, J.L.; Sandt, V.; Kahlert, S.; Kopp, S.; Grimm, D.; Krüger, M. Dexamethasone selectively inhibits detachment of metastatic thyroid cancer cells during random positioning. Cancers 2023, 15, 1641. [Google Scholar] [CrossRef]
- Strube, F.; Infanger, M.; Wehland, M.; Delvinioti, X.; Romswinkel, A.; Dietz, C.; Kraus, A. Alteration of cytoskeleton morphology and gene expression in human breast cancer cells under simulated microgravity. Cell J. 2020, 22, 106–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopp, S.; Sahana, J.; Islam, T.; Petersen, A.G.; Bauer, J.; Corydon, T.J.; Schulz, H.; Saar, K.; Huebner, N.; Slumstrup, L.; et al. The role of nfκb in spheroid formation of human breast cancer cells cultured on the random positioning machine. Sci. Rep. 2018, 8, 921. [Google Scholar] [CrossRef] [PubMed]
- Sahana, J.; Corydon, T.J.; Wehland, M.; Krüger, M.; Kopp, S.; Melnik, D.; Kahlert, S.; Relja, B.; Infanger, M.; Grimm, D. Alterations of growth and focal adhesion molecules in human breast cancer cells exposed to the random positioning machine. Front. Cell. Dev. Biol. 2021, 9, 672098. [Google Scholar] [CrossRef] [Green Version]
- Nassef, M.Z.; Kopp, S.; Melnik, D.; Corydon, T.J.; Sahana, J.; Krüger, M.; Wehland, M.; Bauer, T.J.; Liemersdorf, C.; Hemmersbach, R.; et al. Short-term microgravity influences cell adhesion in human breast cancer cells. Int. J. Mol. Sci. 2019, 20, 5730. [Google Scholar] [CrossRef] [PubMed]
- Calvaruso, M.; Militello, C.; Minafra, L.; La Regina, V.; Torrisi, F.; Pucci, G.; Cammarata, F.P.; Bravata, V.; Forte, G.I.; Russo, G. Biological and mechanical characterization of the random positioning machine (rpm) for microgravity simulations. Life 2021, 11, 1190. [Google Scholar] [CrossRef]
- Sahana, J.; Cortés-Sánchez, J.L.; Sandt, V.; Melnik, D.; Corydon, T.J.; Schulz, H.; Cai, Z.; Evert, K.; Grimm, D.; Wehland, M. Long-term simulation of microgravity induces changes in gene expression in breast cancer cells. Int. J. Mol. Sci. 2023, 24, 1181. [Google Scholar] [CrossRef] [Green Version]
- Hybel, T.E.; Dietrichs, D.; Sahana, J.; Corydon, T.J.; Nassef, M.Z.; Wehland, M.; Kruger, M.; Magnusson, N.E.; Bauer, J.; Utpatel, K.; et al. Simulated microgravity influences vegf, mapk, and pam signaling in prostate cancer cells. Int. J. Mol. Sci. 2020, 21, 1263. [Google Scholar] [CrossRef]
- Dietrichs, D.; Grimm, D.; Sahana, J.; Melnik, D.; Corydon, T.J.; Wehland, M.; Kruger, M.; Vermeesen, R.; Baselet, B.; Baatout, S.; et al. Three-dimensional growth of prostate cancer cells exposed to simulated microgravity. Front. Cell. Dev. Biol. 2022, 10, 841017. [Google Scholar] [CrossRef]
- Schulz, H.; Dietrichs, D.; Wehland, M.; Corydon, T.J.; Hemmersbach, R.; Liemersdorf, C.; Melnik, D.; Hubner, N.; Saar, K.; Infanger, M.; et al. In prostate cancer cells cytokines are early responders to gravitational changes occurring in parabolic flights. Int. J. Mol. Sci. 2022, 23, 7876. [Google Scholar] [CrossRef] [Green Version]
- Ahn, C.B.; Lee, J.H.; Han, D.G.; Kang, H.W.; Lee, S.H.; Lee, J.I.; Son, K.H.; Lee, J.W. Simulated microgravity with floating environment promotes migration of non-small cell lung cancers. Sci. Rep. 2019, 9, 14553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dietz, C.; Infanger, M.; Romswinkel, A.; Strube, F.; Kraus, A. Apoptosis induction and alteration of cell adherence in human lung cancer cells under simulated microgravity. Int. J. Mol. Sci. 2019, 20, 3601. [Google Scholar] [CrossRef] [PubMed]
- Baghoum, H.; Alahmed, H.; Hachim, M.; Senok, A.; Jalaleddine, N.; Al Heialy, S. Simulated microgravity influences immunity-related biomarkers in lung cancer. Int. J. Mol. Sci. 2022, 24, 155. [Google Scholar] [CrossRef] [PubMed]
- Arun, R.P.; Sivanesan, D.; Vidyasekar, P.; Verma, R.S. Pten/foxo3/akt pathway regulates cell death and mediates morphogenetic differentiation of colorectal cancer cells under simulated microgravity. Sci. Rep. 2017, 7, 5952. [Google Scholar] [CrossRef] [PubMed]
- Rembialkowska, N.; Baczynska, D.; Dubinska-Magiera, M.; Choromanska, A.; Biezunska-Kusiak, K.; Gajewska-Naryniecka, A.; Novickij, V.; Saczko, J.; Przystupski, D.; Kulbacka, J. Rccs bioreactor-based modeled microgravity affects gastric cancer cells and improves the chemotherapeutic effect. Membranes 2022, 12, 448. [Google Scholar] [CrossRef] [Green Version]
- Costantini, D.; Overi, D.; Casadei, L.; Cardinale, V.; Nevi, L.; Carpino, G.; Di Matteo, S.; Safarikia, S.; Valerio, M.; Melandro, F.; et al. Simulated microgravity promotes the formation of tridimensional cultures and stimulates pluripotency and a glycolytic metabolism in human hepatic and biliary tree stem/progenitor cells. Sci. Rep. 2019, 9, 5559. [Google Scholar] [CrossRef]
- Masini, M.A.; Bonetto, V.; Manfredi, M.; Pasto, A.; Barberis, E.; Timo, S.; Vanella, V.V.; Robotti, E.; Masetto, F.; Andreoli, F.; et al. Prolonged exposure to simulated microgravity promotes stemness impairing morphological, metabolic and migratory profile of pancreatic cancer cells: A comprehensive proteomic, lipidomic and transcriptomic analysis. Cell. Mol. Life Sci. 2022, 79, 226. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, K.; Eiermann, P.; Tsiockas, W.; Hemmersbach, R.; Gerzer, R. Differential regulation of cgmp signaling in human melanoma cells at altered gravity: Simulated microgravity down-regulates cancer-related gene expression and motility. Microgravity Sci. Technol. 2018, 30, 457–467. [Google Scholar] [CrossRef]
- Chung, Y.J.; Lee, J.I.; Chong, S.; Seok, J.W.; Park, S.J.; Jang, H.W.; Kim, S.W.; Chung, J.H. Anti-proliferative effect and action mechanism of dexamethasone in human medullary thyroid cancer cell line. Endocr. Res. 2011, 36, 149–157. [Google Scholar] [CrossRef]
- Kumar, K.; Moon, B.H.; Datta, K.; Fornace, A.J., Jr.; Suman, S. Simulated galactic cosmic radiation (gcr)-induced expression of spp1 coincide with mammary ductal cell proliferation and preneoplastic changes in apc(min/+) mouse. Life Sci. Space Res. 2023, 36, 116–122. [Google Scholar] [CrossRef]
- Chatziravdeli, V.; Katsaras, G.N.; Lambrou, G.I. Gene expression in osteoblasts and osteoclasts under microgravity conditions: A systematic review. Curr. Genom. 2019, 20, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Topal, U.; Zamur, C. Microgravity, stem cells, and cancer: A new hope for cancer treatment. Stem Cells Int. 2021, 2021, 5566872. [Google Scholar] [CrossRef] [PubMed]
- Sahana, J.; Nassef, M.Z.; Wehland, M.; Kopp, S.; Krüger, M.; Corydon, T.J.; Infanger, M.; Bauer, J.; Grimm, D. Decreased e-cadherin in mcf7 human breast cancer cells forming multicellular spheroids exposed to simulated microgravity. Proteomics 2018, 18, e1800015. [Google Scholar] [CrossRef] [PubMed]
- Wise, P.M.; Sahana, J.; Neviani, P.; Corydon, T.J.; Schulz, H.; Wehland, M.; Infanger, M.; Grimm, D. Prolonged exposure to simulated microgravity changes release of small extracellular vesicle in breast cancer cells. Int. J. Mol. Sci. 2022, 23, 16095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strube, F.; Infanger, M.; Dietz, C.; Romswinkel, A.; Kraus, A. Short-term effects of simulated microgravity on morphology and gene expression in human breast cancer cells. Physiol. Int. 2019, 106, 311–322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feger, B.J.; Thompson, J.W.; Dubois, L.G.; Kommaddi, R.P.; Foster, M.W.; Mishra, R.; Shenoy, S.K.; Shibata, Y.; Kidane, Y.H.; Moseley, M.A.; et al. Microgravity induces proteomics changes involved in endoplasmic reticulum stress and mitochondrial protection. Sci. Rep. 2016, 6, 34091. [Google Scholar] [CrossRef]
- Monti, N.; Masiello, M.G.; Proietti, S.; Catizone, A.; Ricci, G.; Harrath, A.H.; Alwasel, S.H.; Cucina, A.; Bizzarri, M. Survival pathways are differently affected by microgravity in normal and cancerous breast cells. Int. J. Mol. Sci. 2021, 22, 862. [Google Scholar] [CrossRef]
- American Cancer Society. Key Statistics for Prostate Cancer. Available online: https://www.Cancer.Org/cancer/prostate-cancer/about/key-statistics.Html (accessed on 28 November 2022).
- Twombly, R. Prostate modeling experiment success becomes part of legacy of shuttle astronauts. J. Natl. Cancer Inst. 2003, 95, 505–507. [Google Scholar] [CrossRef] [Green Version]
- Tai, S.; Sun, Y.; Squires, J.M.; Zhang, H.; Oh, W.K.; Liang, C.Z.; Huang, J. Pc3 is a cell line characteristic of prostatic small cell carcinoma. Prostate 2011, 71, 1668–1679. [Google Scholar] [CrossRef]
- Grimm, D.; Schulz, H.; Krüger, M.; Cortés-Sánchez, J.L.; Egli, M.; Kraus, A.; Sahana, J.; Corydon, T.J.; Hemmersbach, R.; Wise, P.M.; et al. The fight against cancer by microgravity: The multicellular spheroid as a metastasis model. Int. J. Mol. Sci. 2022, 23, 3073. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Degan, P.; Cortese, K.; Pulliero, A.; Bruno, S.; Gagliani, M.C.; Congiu, M.; Izzotti, A. Simulated microgravity effects on human adenocarcinoma alveolar epithelial cells: Characterization of morphological, functional, and epigenetic parameters. Int. J. Mol. Sci. 2021, 22, 6951. [Google Scholar] [CrossRef] [PubMed]
- Chung, J.H.; Ahn, C.B.; Son, K.H.; Yi, E.; Son, H.S.; Kim, H.S.; Lee, S.H. Simulated microgravity effects on nonsmall cell lung cancer cell proliferation and migration. Aerosp. Med. Hum. Perform. 2017, 88, 82–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisanu, M.E.; Noto, A.; De Vitis, C.; Masiello, M.G.; Coluccia, P.; Proietti, S.; Giovagnoli, M.R.; Ricci, A.; Giarnieri, E.; Cucina, A.; et al. Lung cancer stem cell lose their stemness default state after exposure to microgravity. Biomed. Res. Int. 2014, 2014, 470253. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Xu, P. Global colorectal cancer burden in 2020 and projections to 2040. Transl. Oncol. 2021, 14, 101174. [Google Scholar] [CrossRef]
- Grimm, D.; Bauer, J.; Kossmehl, P.; Shakibaei, M.; Schoberger, J.; Pickenhahn, H.; Schulze-Tanzil, G.; Vetter, R.; Eilles, C.; Paul, M.; et al. Simulated microgravity alters differentiation and increases apoptosis in human follicular thyroid carcinoma cells. FASEB J. 2002, 16, 604–606. [Google Scholar] [CrossRef] [Green Version]
- Masiello, M.G.; Cucina, A.; Proietti, S.; Palombo, A.; Coluccia, P.; D’Anselmi, F.; Dinicola, S.; Pasqualato, A.; Morini, V.; Bizzarri, M. Phenotypic switch induced by simulated microgravity on mda-mb-231 breast cancer cells. Biomed. Res. Int. 2014, 2014, 652434. [Google Scholar] [CrossRef]
- NIH. National Cancer Institute. In Cancer Stat Facts: Melanoma of the Skin. Available online: https://seer.Cancer.Gov/statfacts/html/melan.Html (accessed on 7 November 2022).[Green Version]
- Zhao, T.; Li, R.; Tan, X.; Zhang, J.; Fan, C.; Zhao, Q.; Deng, Y.; Xu, A.; Lukong, K.E.; Genth, H.; et al. Simulated microgravity reduces focal adhesions and alters cytoskeleton and nuclear positioning leading to enhanced apoptosis via suppressing fak/rhoa-mediated mtorc1/nf-kappab and erk1/2 pathways. Int. J. Mol. Sci. 2018, 19, 1994. [Google Scholar] [CrossRef] [Green Version]
- Tan, X.; Xu, A.; Zhao, T.; Zhao, Q.; Zhang, J.; Fan, C.; Deng, Y.; Freywald, A.; Genth, H.; Xiang, J. Simulated microgravity inhibits cell focal adhesions leading to reduced melanoma cell proliferation and metastasis via fak/rhoa-regulated mtorc1 and ampk pathways. Sci. Rep. 2018, 8, 3769. [Google Scholar] [CrossRef]
- Liu, H.; Yao, Z.; Fu, Y.; Feng, J. Review of research into bioregenerative life support system(s) which can support humans living in space. Life Sci. Space Res. 2021, 31, 113–120. [Google Scholar] [CrossRef]
- Kawamoto, N.; Morita, M.T. Gravity sensing and responses in the coordination of the shoot gravitropic setpoint angle. New. Phytol. 2022, 236, 1637–1654. [Google Scholar] [CrossRef] [PubMed]
- Ferl, R.; Wheeler, R.; Levine, H.G.; Paul, A.L. Plants in space. Curr. Opin. Plant. Biol. 2002, 5, 258–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schüler, O.; Hemmersbach, R.; Böhmer, M. A bird’s-eye view of molecular changes in plant gravitropism using omics techniques. Front. Plant. Sci. 2015, 6, 1176. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Wang, L.; Zheng, H. Molecular basis to integrate microgravity signals into the photoperiodic flowering pathway in arabidopsis thaliana under spaceflight condition. Int. J. Mol. Sci. 2021, 23, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Y.; Xie, J.; Wang, L.; Zheng, H. Circumnutation and growth of inflorescence stems of arabidopsis thaliana in response to microgravity under different photoperiod conditions. Life 2020, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Karahara, I.; Suto, T.; Yamaguchi, T.; Yashiro, U.; Tamaoki, D.; Okamoto, E.; Yano, S.; Tanigaki, F.; Shimazu, T.; Kasahara, H.; et al. Vegetative and reproductive growth of arabidopsis under microgravity conditions in space. J. Plant. Res. 2020, 133, 571–585. [Google Scholar] [CrossRef]
- Link, B.M.; Durst, S.J.; Zhou, W.; Stankovic, B. Seed-to-seed growth of arabidopsis thaliana on the international space station. Adv. Space Res. 2003, 31, 2237–2243. [Google Scholar] [CrossRef] [Green Version]
- Link, B.M.; Busse, J.S.; Stankovic, B. Seed-to-seed-to-seed growth and development of arabidopsis in microgravity. Astrobiology 2014, 14, 866–875. [Google Scholar] [CrossRef]
- Musgrave, M.E.; Kuang, A. Plant reproductive development during spaceflight. Adv. Space Biol. Med. 2003, 9, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Califar, B.; Sng, N.J.; Zupanska, A.; Paul, A.L.; Ferl, R.J. Root skewing-associated genes impact the spaceflight response of arabidopsis thaliana. Front. Plant Sci. 2020, 11, 239. [Google Scholar] [CrossRef]
- Choi, W.G.; Barker, R.J.; Kim, S.H.; Swanson, S.J.; Gilroy, S. Variation in the transcriptome of different ecotypes of arabidopsis thaliana reveals signatures of oxidative stress in plant responses to spaceflight. Am. J. Bot. 2019, 106, 123–136. [Google Scholar] [CrossRef] [PubMed]
- Manian, V.; Orozco, J.; Gangapuram, H.; Janwa, H.; Agrinsoni, C. Network analysis of gene transcriptions of arabidopsis thaliana in spaceflight microgravity. Genes 2021, 12, 337. [Google Scholar] [CrossRef] [PubMed]
- Sheppard, J.; Land, E.S.; Toennisson, T.A.; Doherty, C.J.; Perera, I.Y. Uncovering transcriptional responses to fractional gravity in arabidopsis roots. Life 2021, 11, 1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herranz, R.; Vandenbrink, J.P.; Villacampa, A.; Manzano, A.; Poehlman, W.L.; Feltus, F.A.; Kiss, J.Z.; Medina, F.J. Rnaseq analysis of the response of arabidopsis thaliana to fractional gravity under blue-light stimulation during spaceflight. Front. Plant. Sci. 2019, 10, 1529. [Google Scholar] [CrossRef]
- Angelos, E.; Ko, D.K.; Zemelis-Durfee, S.; Brandizzi, F. Relevance of the unfolded protein response to spaceflight-induced transcriptional reprogramming in arabidopsis. Astrobiology 2021, 21, 367–380. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Soga, K.; Hoson, T.; Kotake, T.; Yamazaki, T.; Ishioka, N.; Shimazu, T.; Kamada, M. Microgravity affects the level of matrix polysaccharide 1,3:1,4-β-glucans in cell walls of rice shoots by increasing the expression level of a gene involved in their breakdown. Astrobiology 2020, 20, 820–829. [Google Scholar] [CrossRef]
- Jin, J.; Chen, H.; Cai, W. Transcriptomic analysis reveals the effects of microgravity on rice calli on board the chinese spaceship shenzhou 8. Microgravity Sci. Technol. 2018, 30, 807–816. [Google Scholar] [CrossRef]
- Xu, P.; Chen, H.; Hu, J.; Pang, X.; Jin, J.; Cai, W. Pectin methylesterase gene atpmepcra contributes to physiological adaptation to simulated and spaceflight microgravity in arabidopsis. iScience 2022, 25, 104331. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Sng, N.J.; LeFrois, C.E.; Paul, A.L.; Ferl, R.J. Epigenomics in an extraterrestrial environment: Organ-specific alteration of DNA methylation and gene expression elicited by spaceflight in arabidopsis thaliana. BMC Genom. 2019, 20, 205. [Google Scholar] [CrossRef]
- Xu, P.; Chen, H.; Hu, J.; Cai, W. Potential evidence for transgenerational epigenetic memory in arabidopsis thaliana following spaceflight. Commun. Biol. 2021, 4, 835. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Chen, H.; Jin, J.; Cai, W. Single-base resolution methylome analysis shows epigenetic changes in arabidopsis seedlings exposed to microgravity spaceflight conditions on board the sj-10 recoverable satellite. NPJ Microgravity 2018, 4, 12. [Google Scholar] [CrossRef]
- Jin, H.; Fu, X.; Yu, X.; Zhu, L.; Yang, Q.; Yuan, F. Genome-wide comparison of structural variations and transposon alterations in soybean cultivars induced by spaceflight. Int. J. Mol. Sci. 2022, 23, 13721. [Google Scholar] [CrossRef] [PubMed]
- Kruse, C.P.S.; Meyers, A.D.; Basu, P.; Hutchinson, S.; Luesse, D.R.; Wyatt, S.E. Spaceflight induces novel regulatory responses in arabidopsis seedling as revealed by combined proteomic and transcriptomic analyses. BMC Plant Biol. 2020, 20, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spartz, A.K.; Ren, H.; Park, M.Y.; Grandt, K.N.; Lee, S.H.; Murphy, A.S.; Sussman, M.R.; Overvoorde, P.J.; Gray, W.M. Saur inhibition of pp2c-d phosphatases activates plasma membrane h+-atpases to promote cell expansion in arabidopsis. Plant Cell 2014, 26, 2129–2142. [Google Scholar] [CrossRef] [PubMed]
- Vandenbrink, J.P.; Herranz, R.; Poehlman, W.L.; Alex Feltus, F.; Villacampa, A.; Ciska, M.; Javier Medina, F.; Kiss, J.Z. Rna-seq analyses of arabidopsis thaliana seedlings after exposure to blue-light phototropic stimuli in microgravity. Am. J. Bot. 2019, 106, 1466–1476. [Google Scholar] [CrossRef]
- Villacampa, A.; Ciska, M.; Manzano, A.; Vandenbrink, J.P.; Kiss, J.Z.; Herranz, R.; Medina, F.J. From spaceflight to mars g-levels: Adaptive response of a. Thaliana seedlings in a reduced gravity environment is enhanced by red-light photostimulation. Int. J. Mol. Sci. 2021, 22, 899. [Google Scholar] [CrossRef]
- da Silveira, W.A.; Fazelinia, H.; Rosenthal, S.B.; Laiakis, E.C.; Kim, M.S.; Meydan, C.; Kidane, Y.; Rathi, K.S.; Smith, S.M.; Stear, B.; et al. Comprehensive multi-omics analysis reveals mitochondrial stress as a central biological hub for spaceflight impact. Cell 2020, 183, 1185–1201. [Google Scholar] [CrossRef]
- Herranz, R.; Benguría, A.; Laván, D.A.; López-Vidriero, I.; Gasset, G.; Javier Medina, F.; van Loon, J.J.; Marco, R. Spaceflight-related suboptimal conditions can accentuate the altered gravity response of drosophila transcriptome. Mol. Ecol. 2010, 19, 4255–4264. [Google Scholar] [CrossRef] [Green Version]
- Beisel, N.S.; Noble, J.; Barbazuk, W.B.; Paul, A.L.; Ferl, R.J. Spaceflight-induced alternative splicing during seedling development in arabidopsis thaliana. NPJ Microgravity 2019, 5, 9. [Google Scholar] [CrossRef]
- Heyneke, E.; Hoefgen, R. Meeting the complexity of plant nutrient metabolism with multi-omics approaches. J. Exp. Bot. 2021, 72, 2261–2265. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.H.; Cai, X.; Vaughn, J.N.; von Arnim, A.G. On the functions of the h subunit of eukaryotic initiation factor 3 in late stages of translation initiation. Genome Biol. 2007, 8, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schepetilnikov, M.; Dimitrova, M.; Mancera-Martínez, E.; Geldreich, A.; Keller, M.; Ryabova, L.A. Tor and s6k1 promote translation reinitiation of uorf-containing mrnas via phosphorylation of eif3h. EMBO J. 2013, 32, 1087–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenbrink, J.P.; Kiss, J.Z. Space, the final frontier: A critical review of recent experiments performed in microgravity. Plant Sci. 2016, 243, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Ferranti, F.; Del Bianco, M.; Pacelli, C. Advantages and limitations of current microgravity platforms for space biology research. Appl. Sci. 2021, 11, 68. [Google Scholar] [CrossRef]






| Platforms | µg Duration | Residual Acceleration | Preparation Time | Cost Estimates [16] | Reference |
|---|---|---|---|---|---|
| Bremen Drop Tower: drop catapult | 4.74 s 9.3 s | <10−6 × g <10−6 × g | few months | ≈5 k€ | [17,18] |
| NASA Zero Gravity Research Facility | 5.18 s | <10−5 × g | few months | ≈5 k€ | [19,20] |
| Einstein-Elevator: drop lift/drop | 2 s 4 s | <10−6 × g <10−6 × g | n/a | n/a | [21] |
| Parabolic flight | 22 s | <10−2 × g | months | ≈125 k€ | [22] |
| Sounding rocket | 6 min, 13 min | <10−4 × g | years | >400 k€ | [23] |
| New Shepard | 3 min | <5 × 10−3 × g | years | n/a | [24] |
| Satellites, taxi flights | days to weeks | <10−5 × g | years | n/a | [25,26,27] |
| ISS, Tiangong | months to years | >10−6 × g | years | ≈1–5 M€ | [28] |
| Organisms and Conditions | Observations | Reference |
| U937 Cells (RCCS) and infection with Escherichia coli, mice (HU), gut bacteria, and investigation of intestinal immunity to Citrobacter rodentium | Suppressed MAPK pathway under s-µg in U937 cells. Downregulation of 25 transcripts and upregulation of 11 transcripts of the MAPK signaling pathway. In mice: suppression in the production of TNF and IL-6 in colon and spleen. Higher sensitivity against Citrobacter rodentium due to suppressed innate immune response. | [86] |
| Human colonic epithelial cell line, HT-29 (ATCC, HTB-38) infected with Salmonella typhimurium | Differences in gene expression between space- and ground-infected cells (among others, TNF signaling, NF-κB signaling, and cellular response to lipopolysaccharides). | [69] |
| HU mice, investigation of the gut biome | Dysbiosis followed after HU, which could be mitigated with 4-PBA, indicating that unloading leads to cellular stress. | [60] |
| Lactobacillus reuteri in RWV | These important probiotic bacteria respond with increased expression of stress genes. | [58] |
| Wheat and endophytic bacteria under s-µg | Change in root metabolism and secretion of metabolites. Change in bacteria composition of the rhizosphere. | [61] |
| Yersina pestis, HARV Cultivation | 218 differentially regulated genes. Increased biofilm formation and reduced virulence (downregulation of virulence-associated genes). | [70] |
| Serratia marcescens and Drosophila melanogaster, HARV cultivation | Increased virulence of Serratia marcescens against Drosophila melanogaster. | [74] |
| Drosophila melanogaster on the ISS | Flies reared on the ISS showed smaller and weaker hearts, reduced sarcomeric and ECM gene expression, upregulated expression of proteasome subunit genes, and increased number of proteasomes. | [87] |
| Listeria monocytogenes (LSMMG) | Downregulation of virulence genes. Decreased heat and acid resistance, but increased cold resistance (upregulation of cold-stress genes). | [75] |
| Bacillus subtilis, Spaceflight | Increased mutation rate of rpoB. | [76] |
| Escherichia coli long-term HARV cultivation | Genome sequencing revealed an increased mutation rate, which resulted in the acquirement of antibiotic resistance. | [77] |
| Acinetobacter pittii and Staphylococcus aureus HARV cultivation | Increased rate of horizontal gene transfer. | [71] |
| Vibrio fischeri HARV cultivation | Impaired membrane stability, increased lipopolysaccharide production. | [81] |
| Knufia chersonesos (colorless and pigmented strain) HARV cultivation | Changes in the secretome, transcriptome, and proteome between 1g samples as well among the two strains. No increased stress response in the colorless strain. | [82] |
| Cryptococcus neoformans (melanized and colorless strain) on the ISS | Higher survival rate of melanized strains. Melanin acts as an antioxidant. | [83] |
| Eprymna scolopes HARV co-cultivation with Vibrio fischeri | Complex network of extrinsic/intrinsic apoptosis genes revealed; earlier and stronger expression of caspases. | [88] |
| Caenorhabditis elegans on the ISS | Downregulation of metabolism and cytoskeletal genes. | [89] |
| Expression levels are regulated epigenetically. | [90] | |
| Caenorhabditis elegans on the RCCS | Global upregulation of genes related to oxidative stress. Local overexpression prevented dysfunction but not increased lumen and permeability of the intestines. | [91] |
| Expressional adaptations of the insulin signaling pathway in the intestines. | [92] | |
| Mus musculus on the ISS | No estrous cycle discontinuation. No difference in the expression of genes related to steroidogenesis or mitochondrial cholesterol uptake. | [93] |
| Thymi of ISS mice were smaller, with downregulated genes related to cell cycle control and chromosome organization. | [94] | |
| Erythrocyte production-related genes in the spleen were downregulated. No influence on gene expression in lymph nodes. | [95] |
| Cell Type | Gene Regulation | Microgravity | Reference |
|---|---|---|---|
| Human microvascular endothelial cell line HMEC-1 | Differentially regulated Hallmark collection gene sets: Up: HALLMARK_ADIPOGENESIS, HALLMARK_ ALLOGRAFT_REJECTION, HALLMARK_ ANDROGEN_RESPONSE, HALLMARK_BILE_ACID_ METABOLISM, HALLMARK_CHOLESTEROL_ HOMEOSTASIS, HALLMARK_COAGULATION, HALLMARK_E2F_TARGETS, HALLMARK_FATTY_ACID_METABOLISM, HALLMARK_G2M_CHECKPOINT, HALLMARK_ MITOTIC_SPINDLE, HALLMARK_MTORC1_SIGNALING, HALLMARK_MYC_TARGETS_V1, HALLMARK_ OXIDATIVE_PHOSPHORYLATION, HALLMARK_ PEROXISOME, HALLMARK_PI3K_AKT_MTOR_ SIGNALING, HALLMARK_PROTEIN_SECRETION, HALLMARK_REACTIVE_OXIGEN_SPECIES_PATHWAY, HALLMARK_SPERMATOGENESIS, HALLMARK_UV_ RESPONSE_DN Down: HALLMARK_ESTROGEN_RESPONSE_EARLY, HALLMARK_ESTROGEN_RESPONSE_LATE, HALLMARK_HEDGEHOG_SIGNALING, HALLMARK_HYPOXIA, HALLMARK_IL2_STAT5_ SIGNALING, HALLMARK_KRAS_SIGNALING_DN, HALLMARK_MYOGENESIS, HALLMARK_NOTCH_ SIGNALING, HALLMARK_P53_PATHWAY, HALLMARK_TNFA_SIGNALING_VIA_NFKB, HALLMARK_UV_RESPONSE_UP, HALLMARK_WNT_BETA_CATENIN_SIGNALING | r-µg on the ISS for 160 h | [108] |
| HUVECs | 585 genes significantly upregulated (top 5: TXNIP, MIR15A, ANAPC1, TP53INP1, ID1); 438 genes significantly downregulated (top 5: HSPA1A and -B, HSP90AA2, ATF3, CLCA2, RYBP) | r-µg on the ISS for 10 d | [109] |
| HUVECs | Early upregulation of HSPA1A only after 4 days, string upregulation of TXNIP after 10 days | s-µg on the RWV for 4 and 10 d | [110] |
| HUVECs | Downregulation of SLCO2A1 | s-µg on a clinostat for 7 d | [111] |
| HUVECs | No significant difference in TP53 gene expression | s-µg on a clinostat for 24, 48, and 73 h | [112] |
| HUVECs | Upregulated miRNAs: hsa-miR-628-3p, hsa-miR-3195, hsa-miR-3687, hsa-miR-1257, hsa-miR-3614-5p | s-µg on a clinostat for 48 h | [113] |
| HUVECs | Upregulation of miR-22 after 72 h, concurrent downregulation of SRF and LAMC1 | s-µg on a HARV for 48, 72 and 96 h | [114] |
| HUVECs | 1870 differentially expressed miRNAs; hsa-mir-496, hsa-mir-151a, hsa-miR-296-3p, hsa-mir-148a, hsa-miR-365b-5p, hsamiR-3687, hsa-mir-454, hsa-miR-155-5p, and hsa-miR-145-5p involved in cell adhesion, angiogenesis, cell cycle, JAK-STAT signaling, MAPK signaling, nitric oxide signaling, VEGF signaling, and wound healing pathways | s-µg on a 3D-clinostat for 2 h | [115] |
| Human CVECs | Upregulation of BAX, CASP3, and CYP2D6 after 1 and 3 days Downregulation of BCL2 after 1 and 3 days | s-µg on an RCCS for 1 and 3 d | [116] |
| EA.hy926 | Upregulation of CXCL8 and FN1 in spheroids | s-µg on an RPM for 35 d | [117] |
| EA.hy926 | Differential regulation of TIMP1, IL6, CXCL8, CCL2, B2M | r-µg on the ISS for 12 d | [118] |
| EA.hy926 | Top 20 upregulated genes in EA.hy926 exosomes: ACTB, PGK1, HSPA8, RPL7A, FTH1, GAPDH, HSP90AB1, RPL37A, TPI1, FABP4, S100A6, TOT1, TUBA1B, RPS4X, RPL5, HNRNPK, RPS3A, RPS2, HNRNPA1 | r-µg on the SJ-10 Recoverable Scientific Satellite for 3 and 10 d | [119] |
| Human peripheral blood-derived endothelial progenitor cells | Increase in HIF1A and NOS3 after 12 and 24 h, then decrease after 48 h | s-µg on a 3D-clinostat for 24 h | [120] |
| Cell Type | Gene Regulation | Microgravity | Reference |
|---|---|---|---|
| Mouse splenic dendritic cells and mouse Flt-3L-differentiated BMDCs | Downregulation of IL6, IL1B, IL12B, and CXCL10 | s-µg on an RPM for 24 h | [130] |
| Human blood plasma and peripheral blood mononuclear cells from astronauts | Significant increase in cell-free mitochondrial DNA with high variability (between 2- and 355-fold); IL6, IL8, SOD1, SOD2, GPX1, NOX4, GADD45, CAT1, DNA-PK, and PARP1 elevated in at least 1 of the postflight time points | r-µg on the ISS for 5–13 d | [131] |
| Whole blood samples from astronauts | The most commonly mutated gene was TP53 (7 variants) followed by DNTM3A (6 variants) accounting for 38% of mutations detected | r-µg on the ISS (median 12 d) | [132] |
| Human plasma from two male monozygotic twins, one on Earth, one in space (NASA Twins study) | Significant increase in the proportion of cell-free mitochondrial DNA over time | r-µg on the ISS for 340 d | [133] |
| Humans blood from two male monozygotic twins, one on Earth, one in space (NASA Twins study) | IL-6, TNFR1, and TNFR2 non-canonical NF-kB pathways were affected | r-µg on the ISS for 340 d | [134] |
| Human blood from two male monozygotic twins, one on Earth, one in space (NASA Twins study) | No persistent alteration of long-chain fatty acid desaturase and elongase gene expression associated with 1 year in space | r-µg on the ISS for 340 d | [135] |
| Human Jurkat T cells | 5 µg-sensitive transcript clusters: G3BP1, KPNB1, NUDT3, POMK, SFT2D2 | r-µg on a sounding rocket for 5 min and s-µg on a clinostat for 5 min | [136] |
| Human cell line U937 and human Jurkat T cells | HIF1A was differently regulated in early response to altered gravity, quick adaption of HIF-1α-dependent gene expression (only IL1B continuously downregulated under µg and SERPINE1, PDK1, and SLC2A3 continuously upregulated under hyper-g) | r-µg on a parabolic flight for 20 s and r-µg on a sounding rocket for 5 min | [137] |
| Human Jurkat T cells | Gene expression upregulation in chromosome 18 and downregulation in chromosome 19 for all alterations of r-µg conditions | r-µg on a parabolic flight for 20 s, r-µg on a sounding rocket for 5 min, and s-µg on a clinostat for 5 min | [138] |
| Human myelomonocytic U937 cells and human Jurkat T cells | U937 cells: differentially regulated transcripts during parabolic flight and sounding rocket missions: CYBA, PTGS1, PXDN, and ALOX12, GCLM, GSR, MSRA, MT3, OXSR1, PRDX4, PRNP, PTGS2, SELS, SOD1, respectively. Jurkat T cells: no response of oxidative stress-related transcripts to altered gravity | r-µg on a parabolic flight for 20 s, r-µg on a sounding rocket for 5 min | [139] |
| Human Jurkat T cells | 5 transcripts found to be gravity-regulated in both independent experiments: ATP6V1A, ATP6V1D, IGHD3-3, IGHD3-10, LINC00837 | r-µg on a parabolic flight for 20 s, r-µg on a sounding rocket for 5 min | [140] |
| Mouse hematopoietic progenitor cells | Downregulation of genes of the RAS/ERK/NFκB signaling pathways | r-µg on the Tianzhou-1 cargo ship for 12 days and r-µg on the SJ-10 recoverable satellite for 12 d | [67] |
| Male C57BL/6 J mice | Downregulation of GATA1 and Tal1, downregulation of GATA and Tal1-mediated transcripts | r-µg on the ISS for 35 d | [95] |
| Female C57BL/6J mice | Selected genes involved in glycogen metabolism: Foxo1, Gbe1, Gys2, Ppp1cb, Ppp1ca, Gsk3b, Pck1 upregulated; Pygl downregulated | r-µg on Space Shuttle mission STS-135 for 13 d | [141] |
| Human Jurkat T cells | Resting T cells: FLT1, OTULIN, GPBP1L1, TP53BP1, STK38, PDS5A, PIK3R4, ABCC5, BRWD3, HSPH1, BRD8, NAPB, SLC5A3 upregulated; CXCR3, RBM3, C19orf60, APOBEC2C, SNHG11, SNHG17, ASPSCR1, SFXN2, DCAF16 downregulated Activated T cells: KDM58 upregulated; RBM3, HES1, CXCR3, TUBA4A, HCST, ARID5A, LIMD2, YRDC, LY9, APOBEC3C, LANCL2, RNASEH1, STT3B, PHF7 downregulated | s-µg on an RPM for 24 h | [142] |
| Murine macrophage RAW 264.7 cells | M0 phenotype: Cd86, Actb upregulated, Arg1 downregulated M1 phenotype: Cd86, Mrc1, Actb upregulated M2 phenotype: Cd86, Mrc1, Arg1, Actb upregulated | s-µg on an RWV for 3 d | [143] |
| Human peripheral blood mononuclear cells | s-mg decreased effects of 1.5 h-long ConA/anti-CD28 stimulation on IL2RA, TNFA, CD69, and CCL4 | s-µg on a HARV for 18 h | [144] |
| Human peripheral blood lymphocytes | 230 dysregulated transcription factor and microRNA feed-forward loops were found in µg including immune, cardiovascular, endocrine, nervous, and skeletal system subnetworks | s-µg on an RWV for 24 h | [145] |
| Human peripheral blood mononuclear cells | The Radio Electric Asymmetric Conveyer technology could increase IL2 and IL2R gene expression under s-µg | s-µg on an RPM for 2, 4 and 12 h | [146] |
| Human peripheral blood mononuclear cells | BAX, CASP3, PCNA, LIG4, and MDM2 were positively correlated with all cytokines, expressions of AKT1, TP53, PARP1, OGG1, and APXE1 were negatively correlated with these cytokines | s-µg on an RWV for 24 h (+10μM (-)-isoproterenol hydrochloride and/or 0.8 or 2 Gy radiation) | [147] |
| Female C57BL/6J mice | Top 5 upregulated: Slc22a4, Wt1os, 1700063H04Rik, Eral1, Zfp341 Top 4 downregulated: Tmem161b, Prrc1, Kbtbd8, Gm33989, Vps39 | s-µg by hindlimb unloading + 0.04 Gy irradiation | [148] |
| Danio rerio embryos | Both RLR and TLR signaling pathways were enriched with upregulated genes upon poly(I:C) stimulation and enriched with downregulated genes under s-µg conditions (trim25, nlrx1, traf6, and traf2a and traf6, tlr7, respectively) | s-µg on an RCCS for 24 h | [149] |
| Tissue/Cell Type | Gene Regulation | Microgravity | Reference |
|---|---|---|---|
| Mouse articular cartilage | Upregulated: Igkv6-20, Eif3m, Stfa2, Ms4a3, Gstm2, Gm10417, Top2a, Rbm3, Ifitm6, Igkv4-91 Downregulated: Omd, Olfr1437, Gm6432, Fmod, Gm10673, Ogn, C1s, Olfr118, Olfr1454, Serpina1b, Olfr764, Gsn, Ccdc80, Slc35e3, Olfr948, Clu, Dcn, Olfr347, Olfr1014, Gm21428, Ect2l, Dpt, Prg4, Prelp, Olfr338, Angptl7, Col10a1, Retnla, Myoc, Thbs4, Pcolce2, Cyp2e1, Cxcl13, Clec3a, Comp, Ecrg4, Cytl1 | r-µg spaceflight BION, 30 d | [150] |
| Mouse sternal cartilage | Upregulated: Klhl38, Acot2, Fbxo32, Nr1d1, Trim63, Htra4, Lox, Prg4, Slc43a1, Lmod2, Aldoc, Slc39a8, Etv5, n-R5s88, Chac1, Tango2, Pdk4, Chi3l1, Impdh2, Zfp600, Sesn1, Myf6, Cfhr2, Tsen15, Omd, Inmt, Sgcg, Gm5886, Tacc2, Ankrd1 Downregulated: Taf1d, Hba-a2, Mki67, Slc4a1, Atp6v0d2, Sfrp2, Car1, Nr4a1, Mpo, Svs3b, Hist1h3f, Retnlg, Hist1h3a, Top2a, Hist1h3d, Hist1h3i, Hist1h2ab, Alpl, Hist2h3c2, Hbb-bs, Hbb-bt, Col1a1, Acp5, Hp, Car2, Igkv4-55, Gypa, Ltf, Igkv1-117, Ibsp, Mmp13, Mmp9, S100a9, S100a8, Ngp | ||
| Meniscus constructs from healthy human meniscus fibrochondrocytes (male and female donors) | Upregulation: BMP8A, CD36, COL10A1, COL9A3, FGF1, IBSP, IHH, MMP10, PHOSPHO1, S100A1, SPP1 Only in a subpopulation of high-responding (related to COL10A1 expression) cells from female donors | s-µg on an RCCS for 3 weeks | [151] |
| Tissue-engineered menisci from human female and male meniscus fibrochondrocytes | Top 10 Upregulated: IGFBP1, OLFML2A, NET O 1, ADAMTS14. PCSK9, VSTM2L, BMPER, NTM, HMOX1, CAPG Top 10 Downregulated: LEP, R3HDML, APLN, STC1, NGF, TGFA, VEGFA, DSCAML1, ADAMTSL2, PDE4C | s-µg on an RCCS for 3 weeks | [152] |
| Primary bovine chondrocytes | Downregulation: TRPC1 (5- to 10-fold) | s-µg on the RPM for 6 and 8 d | [153] |
| Human articular chondrocytes | Upregulation of IL6, RUNX2, RUNX3, SPP1, SOX6, SOX9, and MMP13 | s-µg on the RPM for 24 h | [154] |
| Muscle Type | Gene Regulation | Microgravity | Reference |
|---|---|---|---|
| Mouse gastrocnemius and quadriceps muscles | DEGs in both muscles: Atp5g2, Peg3, Mid1, Agpat2, Cyfip2, Pgk1, Rpl37a, Nav2, Rpl24, Hspb7, Fabp3, Agbl1, Bdh1, Ccnb1ip1, H2afz | r-µg on the ISS for 9 weeks | [155] |
| Mouse soleus and extensor digitorum longus muscles | DEGs in both muscles: Orm1, Myo5a, Hp, Adipoq, Lcn2, Snhg5, Acp5, Snhg1, Cdkal1, Cfd, Cdkn1a, Myf6, Eif4ebp1, Synj2, Sesn1, Pnmt, Snhg1, Npr3, Sorbs1, Aspn, Ung, Mafb, Tfrc, Dbp | r-µg on a BION-MQ-capsule for 30 d | [156] |
| Mouse soleus muscle | A total of 1130 differentially expressed genes were found between wild-type and Nrf2 knock-out mice; top 10 enriched pathways: mitochondrial dysfunction, oxidative phosphorylation, sirtuin signaling pathway, remodelling of epithelial adherens junctions, ILK signaling, epithelial adherens junction signaling, integrin signaling, germ cell–sertoli cell junction signaling, actin cytoskeleton signaling, EIF2 signaling | r-µg on the ISS for 31 d | [157] |
| Mouse soleus muscle | Identification of voltage-dependent calcium channel gamma-1 subunit (Cacng1) as a novel candidate gene for muscle atrophy | r-µg on the ISS for 35 d | [158] |
| Cell Type | Gene Regulation | Microgravity | Reference |
|---|---|---|---|
| Mouse Ocy454 | Top 5: 2 d upregulated: Cxcl14, Mmp13, C2cd4a, Cdc25c, Gdf7 2 d downregulated: Fabp3, Ptprz1, Sparcl1, Slc12a2, Ogn 4 d upregulated: Scgb1b12, Acan, Taar5, Pappa2, Cfap20dc 4 d downregulated: Slc12a2, Tfrc, Ajuba, Idi1, Dmp1 6 d upregulated: Car9, Egln3, Serping1, Tmem45a, Selenbp1 6 d downregulated: Cth, Ctps, Pinx1, Ddx21, Ostn | r-µg on the ISS for 2, 4, and 6 d | [159] |
| Human blood-derived stem cells | Methylation at histone H3 sites H3K4me3, H3K27me2/3, H3K79me2/3, and H3K9me2/3 mediates cellular reprogramming that drives gene expression | r-µg on the ISS for 72 h | [160] |
| Murine osteoblasts and osteoclasts | Downregulation: Atf4, RunX2, Osterix | r-µg on the ISS for 14 d | [26] |
| Human bone marrow mesenchymal stem cells | Downregulation: CDKN3, MCM5, CCNB1, CDK1, RUNX2, ALPL, BMP2, and COL1A1 Upregulation: RUNX2, ALPL, BMP2, and COL1A1 | s-µg on an RPM for 2, 7, and 14 d | [161] |
| Human fetal osteoblast cells | Differential gene expression of TGFB1, BMP2, SOX9, ACTB, TUBB, VIM, LAMA1, COL1A1, SPP1, and FN1 | s-µg on an RPM for 7 and 14 d | [162] |
| 7F2 murine pre-osteoblasts | Downregulation: Alpl, Run, and on after 6 days, no difference between g-levels | s-µg (“full”, 0.16 g (Moon), and 0.38 g (Mars)) on an RPM for 2, 4, and 6 d | [163] |
| MC3T3-E1 mouse pre-osteoblasts | Identification of 3 core circRNAs (circ_014154, circ_010383, and circ_012460) and six core mRNAs (Alpl, Bg1ap, Col1a1, Omd, Ogn, and Bmp-4) involved in osteogenic differentiation under µg | s-µg on an RCCS for 72 h | [164] |
| Organism | Gene Regulation | Tissue | Microgravity | Reference |
|---|---|---|---|---|
| Arabidopsis thaliana (Col-0) | Upregulated: DNA replication, DNA repair, far red/red light response, photosynthesis, secondary metabolite biosynthesis Downregulated: Abscisic acid, response to stress, amino acid catabolism, sucrose starvation, absence of light, ribosome biogenesis, translation Notes: Unfolded protein response seems to play a minor role in spaceflight adaptation. | Whole seedlings, 14 days old | BRIC hardware on the ISS, 14 days of µg. (GC) | [241] |
| Arabidopsis thaliana (Col-0 and Ws-0) | Differential alternative splicing in response to spaceflight in 48 genes in Ws-0 and 27 genes in Col-0. | Roots, 4 and 8 days old | VEGGIE system on the ISS, up to 8 days of µg. (GC) | [255] |
| Arabidopsis thaliana (Col-0 and Ws-0) | Col-0 vs. spr1: Less DEGs in spr1 than in Col-0 WT during spaceflight. Enriched categories: Response to salicylic acid, carbon fixation, RuBisCo genes, apoplast-localized genes Ws-0 vs. sku5: 2–5 times more DEGs in sku5 than in Ws-0 WT during spaceflight Enriched categories: Abscisic acid signaling, response to stress, seed development, post-embryonic development | Roots, 4 and 8 days old | VEGGIE system on the ISS, up to 8 days of µg. (GC) | [236] |
| Arabidopsis thaliana (Col-0, Ler-0, Ws-2, Cvi-0) | Upregulation across all ecotypes: HSP101, HSP70, HSP23.5, HSP81.1, AT2G32120.2 Downregulation across all ecotypes: AT5G44417.1, RCI3, AT1G64370.1, AtPrx22 Notes: Enrichment of 21 upregulated heat shock protein/factor transcripts and 14 downregulated peroxidase transcripts across some but not all ecotypes show characteristics of oxidative stress. | Whole seedlings, 8 days old | BRIC hardware on the ISS, 8 days of µg. (GC) | [237] |
| Arabidopsis thaliana (Ler-0) | Enriched GO categories per g-level: µg—response to light, photosynthesis 0.1g—response to stress, chemicals 0.3g—cell wall, membrane, plastid Decrease in DEGs with higher g-levels Changes in expression along the g-gradient: Mitochondria, plastid, cell wall, and cell membrane-related processes. F-box/RMI-like/FDB-like domain family genes (AT5G44980, AT5G56370, AT5G56380, AT5G42460) Notes: Highest number of DEGs in 0.1 g condition with mostly stress-related pathways. Authors suggest competition of weak phototropism with weak gravitropism leading to stress. | Whole seedlings, 6 days old | EMCS hardware on the ISS, 6 days of µg, 0.1g, 0.3g, 0.5g, 0.8g, and 1g. Blue light stimulus for the last 48 h. (FC) | [240] |
| Arabidopsis thaliana (Col-0) | Enriched categories: Cell wall organization (transcripts downregulated, proteins upregulated) Redox homeostasis and ROS signaling (increased transcript regulation, no protein regulation) Protein degradation (only transcripts) Auxin-related genes (PIN, LAX, IAA, ARF, TIR1-family) Post-transcriptional regulation (upregulation of DCP5 and AGO4) Plastid translation, import to stroma, targeting to chloroplast, plastid organization, chlorophyll biosynthetic process (only transcripts) Notes: Large discrepancy between transcript changes and protein changes in plastids, suggesting plastid dysregulation under µg. | Whole seedlings, 3 days old | BRIC hardware on the ISS, 3 days of µg. (GC) | [249] |
| Arabidopsis thaliana (Col-0 and Ws-0) | Network analysis reveals 5 strongly regulated hub genes of the cell wall-modifying XTH-family. | Whole seedlings and root tips, 11–12 days old | ABRS hardware on the ISS, up to 12 days of µg. (GC) | [238] |
| Arabidopsis thaliana (Col-0) | Identification of 101 genes with dosage-dependent response to g-level. Enriched categories: Transcription factors (20 genes) Chaperones (9 genes) Defense response (12 genes) Cell wall associated (16 genes) Root development (8 genes) Auxin-related (3 genes) | Roots, 5 days old | EMCS hardware on the ISS, 5 days of µg, 0.53g, 0.65g, and 0.88g. (FC) | [239] |
| Arabidopsis thaliana (Ler-0) | Upregulated categories: Ribosome synthesis Oxidative phosphorylation Downregulated categories: Photosynthesis/antenna proteins Porphyrin and chlorophyll metabolism Protein processing in ER Starch and sucrose metabolism Carotenoid biosynthesis | Whole seedlings, 6 days old | EMCS hardware on the ISS, 6 days of µg and 1g. Blue light stimulus for the last 48 h. (FC) | [251] |
| Arabidopsis thaliana (Ler-0) | Enriched categories and pathways: Transcription factors (WRKY, ERF, NAC, MYB) Plastidic transcription (enriched only in µg) Auxin (GH3 and SAUR genes, activated in µg, inhibited at 0.3g + darkness) Cytokinins (activated in µg, inhibited at 0.3g) ABA (activated in all conditions except µg + darkness) Brassinosteroid (inhibited by upregulation of BRI1 suppressor 1) Jasmonic acid (upregulation of repressors) Notes: Hormone pathways are more affected by g-level than light conditions. At 0.3g, red light can reverse auxin pathway inhibition. Stress-related pathways are more activated at 0.3g than in µg. | Whole seedlings, 6 days old | EMCS hardware on the ISS, 6 days of µg and 0.3g. Red light or darkness for the last 48 h. (GC) | [252] |
| Arabidopsis thaliana (Col-0) | Upregulated categories: Metabolic process, response to stress, temperature and wounding, protein stabilization Downregulated categories: Response to stimulus, reproductive developmental process, regulation of the metabolic process, circadian rhythm, gibberellins, mRNA processes Notes: Flowering genes (53) were mostly downregulated under µg, affecting the flowering hub genes FT and SOC1. Delay of flowering time in µg by 20 days. | Rosette leaves, 48 days old | Plant culture box hardware on the Chinese spacelab TG-2, 48 days of µg. (GC) | [230] |
| Arabidopsis thaliana (Col-0) | Altered gene methylation in DNA methylation-associated genes, cell-wall modification genes, hormone signaling-related genes and transposable elements. Induction of transposable elements by unstable methylations in response to spaceflight. | Whole seedlings, 8.5 days old | Cultivation units on the recoverable satellite SJ-10, 60 h of µg after 6 days of pre-culturing on the ground. (GC) | [247] |
| Arabidopsis thaliana (Col-0) | Altered gene methylation and expression in the first offspring generation after spaceflight in the abscisic acid-activated pathway, protein phosphorylation, and nitrate signaling pathway. Partial retention of phenotypic differences and differentially methylated regions in the following generation. Notes: Could not confirm methylation changes in parent generation from previous study. | Whole seedlings, 11 days old, multiple generations | Cultivation units on the recoverable satellite SJ-10, 11 days of µg after 6 days of pre-culturing on the ground. (GC) | [246] |
| Arabidopsis thaliana (Col-0) | Altered gene methylation and expression of pectin methylesterase regulator gene AtPMEPCRA in parent and offspring generation after spaceflight. | Whole seedlings, 11 days old, multiple generations | Cultivation units on the recoverable satellite SJ-10, 11 days of µg after 6 days of pre-culturing on the ground. (GC) | [244] |
| Arabidopsis thaliana (Ws-0) | Portion of differentially expressed genes, that are also differentially methylated: Leaves: 143 of 743 Roots: 21 of 75 Increased methylation in CHG and CHH contexts in leaves during spaceflight. Notes: Nearly half of all differentially expressed genes in leaves were reactive oxygen signaling-related. | Leaves and roots, 11 days old | VEGGIE system on the ISS, 11 days of µg. (GC) | [245] |
| Oryza sativa | Differential regulation of 8 cell-wall-related transcripts and 3 aquaporins. | Calli, 17.5 days old | SIMBOX hardware on the Shenzhou 8, 324 h of µg and 1g. (FC + GC) | [243] |
| Oryza sativa (Koshihikari) | Upregulation of endo-1,3:1,3-β-glucanase (OsEGL1) under µg, leading to a reduced 1,3:1,3-β-glucan content in the cell wall. | Shoots, 4–5 days old | CBEF hardware on the ISS, 99 h, 127 h and 136 h of µg and 1g. (FC) | [242] |
| Glycine max (T75 and Z9) | Increased activity of transposable elements and genomic plasticity after space mutation. Enriched gene categories affected by genomic restructuring: Stress response and stimulus—278 genes Cell wall-related—27 genes Auxin metabolism and transport—26 genes General signal transduction—77 genes | Leaves, 15 days old, 6 years of breeding after space mutation | Space mutation for 15 days of µg aboard recoverable satellite Shijian 8. (GC) | [248] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corydon, T.J.; Schulz, H.; Richter, P.; Strauch, S.M.; Böhmer, M.; Ricciardi, D.A.; Wehland, M.; Krüger, M.; Erzinger, G.S.; Lebert, M.; et al. Current Knowledge about the Impact of Microgravity on Gene Regulation. Cells 2023, 12, 1043. https://doi.org/10.3390/cells12071043
Corydon TJ, Schulz H, Richter P, Strauch SM, Böhmer M, Ricciardi DA, Wehland M, Krüger M, Erzinger GS, Lebert M, et al. Current Knowledge about the Impact of Microgravity on Gene Regulation. Cells. 2023; 12(7):1043. https://doi.org/10.3390/cells12071043
Chicago/Turabian StyleCorydon, Thomas J., Herbert Schulz, Peter Richter, Sebastian M. Strauch, Maik Böhmer, Dario A. Ricciardi, Markus Wehland, Marcus Krüger, Gilmar S. Erzinger, Michael Lebert, and et al. 2023. "Current Knowledge about the Impact of Microgravity on Gene Regulation" Cells 12, no. 7: 1043. https://doi.org/10.3390/cells12071043