
Mesencephalic Astrocyte-Derived Neurotrophic Factor (MANF): An Emerging Therapeutic Target for Neurodegenerative Disorders

Abstract
:1. Introduction
2. Structure-Activity Relationship of MANF
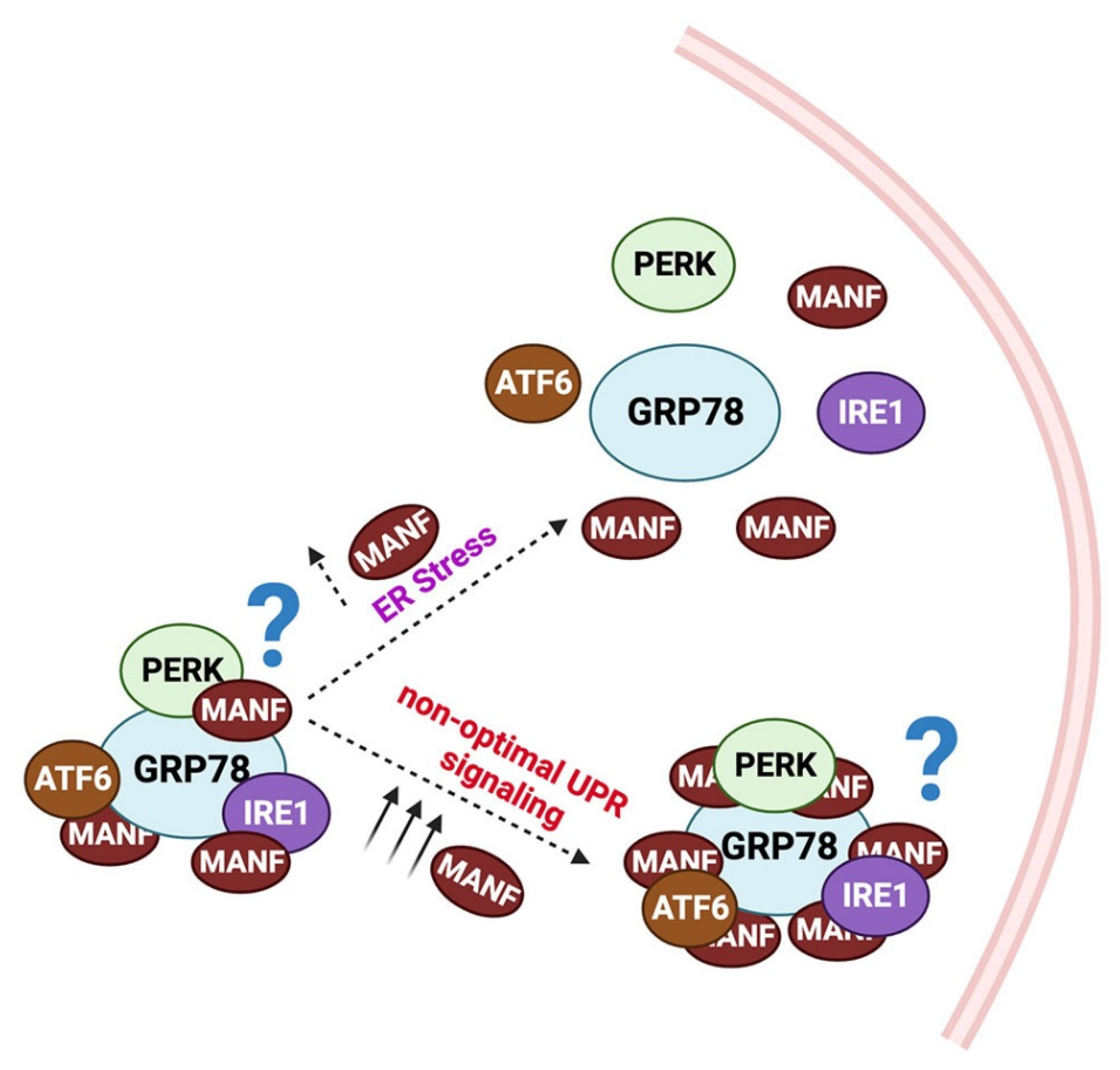
3. The Crosstalk between ER Stress, UPR, and MANF
4. MANF Signaling in Parkinson’s Disease (PD)
5. MANF Signaling in Alzheimer’s Disease (AD)
6. MANF Signaling in Stroke
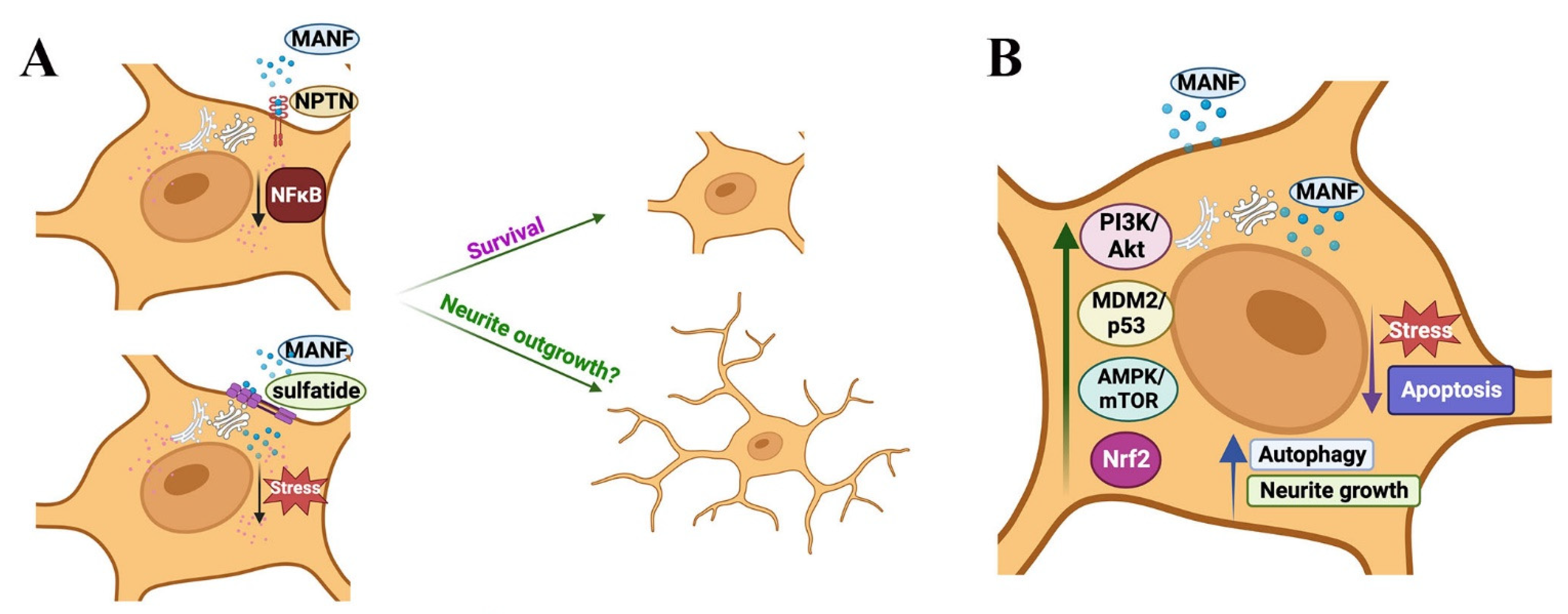
7. Mechanisms of MANF’s Actions Independent of the ER Stress-UPR Axis
8. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Hellman, M.; Arumae, U.; Yu, L.Y.; Lindholm, P.; Peranen, J.; Saarma, M.; Permi, P. Mesencephalic astrocyte-derived neurotrophic factor (MANF) has a unique mechanism to rescue apoptotic neurons. J. Biol. Chem. 2011, 286, 2675–2680. [Google Scholar] [CrossRef] [Green Version]
- Petrova, P.; Raibekas, A.; Pevsner, J.; Vigo, N.; Anafi, M.; Moore, M.K.; Peaire, A.E.; Shridhar, V.; Smith, D.I.; Kelly, J.; et al. MANF: A new mesencephalic, astrocyte-derived neurotrophic factor with selectivity for dopaminergic neurons. J. Mol. Neurosci. 2003, 20, 173–188. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, D.Y.; Chen, X.S.; Zhu, L.; Wan, L.H. MANF: A Novel Endoplasmic Reticulum Stress Response Protein-The Role in Neurological and Metabolic Disorders. Oxid. Med. Cell. Longev. 2021, 2021, 6467679. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, P.; Saarma, M. Novel CDNF/MANF family of neurotrophic factors. Dev. Neurobiol. 2010, 70, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Richman, C.; Rashid, S.; Prashar, S.; Mishra, R.; Selvaganapathy, P.R.; Gupta, B.P.C. elegans MANF Homolog Is Necessary for the Protection of Dopaminergic Neurons and ER Unfolded Protein Response. Front. Neurosci. 2018, 12, 544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palgi, M.; Lindstrom, R.; Peranen, J.; Piepponen, T.P.; Saarma, M.; Heino, T.I. Evidence that DmMANF is an invertebrate neurotrophic factor supporting dopaminergic neurons. Proc. Natl. Acad. Sci. USA 2009, 106, 2429–2434. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.C.; Sundvik, M.; Rozov, S.; Priyadarshini, M.; Panula, P. MANF regulates dopaminergic neuron development in larval zebrafish. Dev. Biol. 2012, 370, 237–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Ke, Z.; Alimov, A.; Xu, M.; Frank, J.A.; Fang, S.; Luo, J. Spatiotemporal expression of MANF in the developing rat brain. PLoS ONE 2014, 9, e90433. [Google Scholar] [CrossRef] [Green Version]
- Glembotski, C.C. Functions for the cardiomyokine, MANF, in cardioprotection, hypertrophy and heart failure. J. Mol. Cell. Cardiol. 2011, 51, 512–517. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Lee, H.; Manson, S.R.; Lindahl, M.; Evans, B.; Miner, J.H.; Urano, F.; Chen, Y.M. Mesencephalic Astrocyte-Derived Neurotrophic Factor as a Urine Biomarker for Endoplasmic Reticulum Stress-Related Kidney Diseases. J. Am. Soc. Nephrol. 2016, 27, 2974–2982. [Google Scholar] [CrossRef] [Green Version]
- Palgi, M.; Greco, D.; Lindstrom, R.; Auvinen, P.; Heino, T.I. Gene expression analysis of Drosophilaa Manf mutants reveals perturbations in membrane traffic and major metabolic changes. BMC Genom. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindahl, M.; Danilova, T.; Palm, E.; Lindholm, P.; Võikar, V.; Hakonen, E.; Ustinov, J.; Andressoo, J.O.; Harvey, B.K.; Otonkoski, T.; et al. MANF is indispensable for the proliferation and survival of pancreatic β cells. Cell. Rep. 2014, 7, 366–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, M.; Vozdek, R.; Hnizda, A.; Jiang, C.; Wang, B.; Kuchar, L.; Li, T.; Zhang, Y.; Wood, C.; Feng, L.; et al. Conserved roles of C. elegans and human MANFs in sulfatide binding and cytoprotection. Nat. Commun. 2018, 9, 897. [Google Scholar] [CrossRef] [Green Version]
- Pakarinen, E.; Lindholm, P.; Saarma, M.; Lindahl, M. CDNF and MANF regulate ER stress in a tissue-specific manner. Cell. Mol. Life Sci. 2022, 79, 124. [Google Scholar] [CrossRef]
- Amsel, A.D.; Rathaus, M.; Kronman, N.; Cohen, H.Y. Regulation of the proapoptotic factor Bax by Ku70-dependent deubiquitylation. Proc. Natl. Acad. Sci. USA 2008, 105, 5117–5122. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Feng, L.; Wang, X.; Du, J.; Chen, Y.; Yang, W.; Zhou, C.; Cheng, L.; Shen, Y.; Fang, S.; et al. Mesencephalic astrocyte-derived neurotrophic factor is involved in inflammation by negatively regulating the NF-κB pathway. Sci. Rep. 2015, 5, 8133. [Google Scholar] [CrossRef] [Green Version]
- Parkash, V.; Lindholm, P.; Peranen, J.; Kalkkinen, N.; Oksanen, E.; Saarma, M.; Leppanen, V.M.; Goldman, A. The structure of the conserved neurotrophic factors MANF and CDNF explains why they are bifunctional. Protein Eng. Des. Sel. 2009, 22, 233–241. [Google Scholar] [CrossRef]
- Lohelaid, H.; Anttila, J.E.; Liew, H.K.; Tseng, K.Y.; Teppo, J.; Stratoulias, V.; Airavaara, M. UPR Responsive Genes Manf and Xbp1 in Stroke. Front. Cell. Neurosci. 2022, 16, 900725. [Google Scholar] [CrossRef] [PubMed]
- Matlik, K.; Yu, L.Y.; Eesmaa, A.; Hellman, M.; Lindholm, P.; Peranen, J.; Galli, E.; Anttila, J.; Saarma, M.; Permi, P.; et al. Role of two sequence motifs of mesencephalic astrocyte-derived neurotrophic factor in its survival-promoting activity. Cell Death Dis. 2015, 6, e2032. [Google Scholar] [CrossRef] [Green Version]
- Henderson, M.J.; Richie, C.T.; Airavaara, M.; Wang, Y.; Harvey, B.K. Mesencephalic astrocyte-derived neurotrophic factor (MANF) secretion and cell surface binding are modulated by KDEL receptors. J. Biol. Chem. 2013, 288, 4209–4225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Krieken, R.; Tsai, Y.L.; Carlos, A.J.; Ha, D.P.; Lee, A.S. ER residential chaperone GRP78 unconventionally relocalizes to the cell surface via endosomal transport. Cell. Mol. Life Sci. 2021, 78, 5179–5195. [Google Scholar] [CrossRef] [PubMed]
- Glembotski, C.C.; Thuerauf, D.J.; Huang, C.; Vekich, J.A.; Gottlieb, R.A.; Doroudgar, S. Mesencephalic astrocyte-derived neurotrophic factor protects the heart from ischemic damage and is selectively secreted upon sarco/endoplasmic reticulum calcium depletion. J. Biol. Chem. 2012, 287, 25893–25904. [Google Scholar] [CrossRef] [Green Version]
- Lindstrom, R.; Lindholm, P.; Kallijarvi, J.; Yu, L.Y.; Piepponen, T.P.; Arumae, U.; Saarma, M.; Heino, T.I. Characterization of the structural and functional determinants of MANF/CDNF in Drosophila in vivo model. PLoS ONE 2013, 8, e73928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voutilainen, M.H.; Back, S.; Porsti, E.; Toppinen, L.; Lindgren, L.; Lindholm, P.; Peranen, J.; Saarma, M.; Tuominen, R.K. Mesencephalic astrocyte-derived neurotrophic factor is neurorestorative in rat model of Parkinson’s disease. J. Neurosci. 2009, 29, 9651–9659. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.; Di, Z.; He, Y.; Wang, R.; Ma, Y.; Sun, R.; Li, J.; Wang, T.; Shen, Y.; Fang, S.; et al. Mesencephalic astrocyte-derived neurotrophic factor (MANF) protects against Aβ toxicity via attenuating Aβ-induced endoplasmic reticulum stress. J. Neuroinflamm. 2019, 16, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Airavaara, M.; Shen, H.; Kuo, C.C.; Peranen, J.; Saarma, M.; Hoffer, B.; Wang, Y. Mesencephalic astrocyte-derived neurotrophic factor reduces ischemic brain injury and promotes behavioral recovery in rats. J. Comp. Neurol. 2009, 515, 116–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danilova, T.; Lindahl, M. Emerging Roles for Mesencephalic Astrocyte-Derived Neurotrophic Factor (MANF) in Pancreatic Beta Cells and Diabetes. Front. Physiol. 2018, 9, 1457. [Google Scholar] [CrossRef]
- Szegezdi, E.; Logue, S.E.; Gorman, A.M.; Samali, A. Mediators of endoplasmic reticulum stress-induced apoptosis. EMBO Rep. 2006, 7, 880–885. [Google Scholar] [CrossRef] [Green Version]
- Montaser, H.; Patel, K.A.; Balboa, D.; Ibrahim, H.; Lithovius, V.; Naatanen, A.; Chandra, V.; Demir, K.; Acar, S.; Ben-Omran, T.; et al. Loss of MANF Causes Childhood-Onset Syndromic Diabetes Due to Increased Endoplasmic Reticulum Stress. Diabetes 2021, 70, 1006–1018. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, J.; Jiang, M.; Cai, Q.; Fang, J.; Jin, L. MANF improves the MPP(+)/MPTP-induced Parkinson’s disease via improvement of mitochondrial function and inhibition of oxidative stress. Am. J. Transl. Res. 2018, 10, 1284–1294. [Google Scholar]
- Inagi, R.; Ishimoto, Y.; Nangaku, M. Proteostasis in endoplasmic reticulum—New mechanisms in kidney disease. Nat. Rev. Nephrol. 2014, 10, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Hetz, C.; Zhang, K.; Kaufman, R.J. Mechanisms, regulation and functions of the unfolded protein response. Nat. Rev. Mol. Cell. Biol. 2020, 21, 421–438. [Google Scholar] [CrossRef]
- Papaioannou, A.; Chevet, E. Driving Cancer Tumorigenesis and Metastasis Through UPR Signaling. Curr. Top. Microbiol. Immunol. 2018, 414, 159–192. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.J.; Kopp, M.C.; Larburu, N.; Nowak, P.R.; Ali, M.M.U. Structure and Molecular Mechanism of ER Stress Signaling by the Unfolded Protein Response Signal Activator IRE1. Front. Mol. Biosci. 2019, 6, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufman, R.J.; Back, S.H.; Song, B.; Han, J.; Hassler, J. The unfolded protein response is required to maintain the integrity of the endoplasmic reticulum, prevent oxidative stress and preserve differentiation in β-cells. Diabetes Obes. Metab. 2010, 12 (Suppl. S2), 99–107. [Google Scholar] [CrossRef] [Green Version]
- Tousson-Abouelazm, N.; Papillon, J.; Guillemette, J.; Cybulsky, A.V. Urinary ERdj3 and mesencephalic astrocyte-derived neutrophic factor identify endoplasmic reticulum stress in glomerular disease. Lab. Investig. 2020, 100, 945–958. [Google Scholar] [CrossRef]
- Sousa-Victor, P.; Neves, J.; Cedron-Craft, W.; Ventura, P.B.; Liao, C.Y.; Riley, R.R.; Soifer, I.; van Bruggen, N.; Kolumam, G.A.; Villeda, S.A.; et al. MANF regulates metabolic and immune homeostasis in ageing and protects against liver damage. Nat. Metab. 2019, 1, 276–290. [Google Scholar] [CrossRef]
- Dernoncourt, A.; Sauzay, C.; Schmidt, J.; Louandre, C.; Gomila, C.; Duhaut, P.; Herpe, Y.E.; Saidak, Z.; Galmiche, A.; Salle, V. Analysis of Mesencephalic Astrocyte-derived Neurotrophic Factor in Multiple Myeloma. Anticancer Res. 2021, 41, 4305–4312. [Google Scholar] [CrossRef]
- Mizobuchi, N.; Hoseki, J.; Kubota, H.; Toyokuni, S.; Nozaki, J.; Naitoh, M.; Koizumi, A.; Nagata, K. ARMET is a soluble ER protein induced by the unfolded protein response via ERSE-II element. Cell. Struct. Funct. 2007, 32, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Apostolou, A.; Shen, Y.; Liang, Y.; Luo, J.; Fang, S. Armet, a UPR-upregulated protein, inhibits cell proliferation and ER stress-induced cell death. Exp. Cell Res. 2008, 314, 2454–2467. [Google Scholar] [CrossRef]
- Wang, Y.; Wen, W.; Li, H.; Clementino, M.; Xu, H.; Xu, M.; Ma, M.; Frank, J.; Luo, J. MANF is neuroprotective against ethanol-induced neurodegeneration through ameliorating ER stress. Neurobiol. Dis. 2021, 148, 105216. [Google Scholar] [CrossRef] [PubMed]
- Lindahl, M.; Saarma, M.; Lindholm, P. Unconventional neurotrophic factors CDNF and MANF: Structure, physiological functions and therapeutic potential. Neurobiol. Dis. 2017, 97, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Yan, Y.; Rato, C.; Rohland, L.; Preissler, S.; Ron, D. MANF antagonizes nucleotide exchange by the endoplasmic reticulum chaperone BiP. Nat. Commun. 2019, 10, 541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maetzler, W.; Liepelt, I.; Berg, D. Progression of Parkinson’s disease in the clinical phase: Potential markers. Lancet Neurol. 2009, 8, 1158–1171. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.; Kolaski, E.; Babcock, D.T. Vexed mutations promote degeneration of dopaminergic neurons through excessive activation of the innate immune response. NPJ Parkinsons Dis. 2022, 8, 147. [Google Scholar] [CrossRef]
- Zhang, Z.; Shen, Y.; Luo, H.; Zhang, F.; Peng, D.; Jing, L.; Wu, Y.; Xia, X.; Song, Y.; Li, W.; et al. MANF protects dopamine neurons and locomotion defects from a human α-synuclein induced Parkinson’s disease model in C. elegans by regulating ER stress and autophagy pathways. Exp. Neurol. 2018, 308, 59–71. [Google Scholar] [CrossRef]
- Cordero-Llana, Ó.; Houghton, B.C.; Rinaldi, F.; Taylor, H.; Yáñez-Muñoz, R.J.; Uney, J.B.; Wong, L.F.; Caldwell, M.A. Enhanced efficacy of the CDNF/MANF family by combined intranigral overexpression in the 6-OHDA rat model of Parkinson’s disease. Mol. Ther. 2015, 23, 244–254. [Google Scholar] [CrossRef] [Green Version]
- Hao, F.; Yang, C.; Chen, S.S.; Wang, Y.Y.; Zhou, W.; Hao, Q.; Lu, T.; Hoffer, B.; Zhao, L.R.; Duan, W.M.; et al. Long-term protective effects of AAV9-mesencephalic astrocyte-derived neurotrophic factor gene transfer in parkinsonian rats. Exp. Neurol. 2017, 291, 120–133. [Google Scholar] [CrossRef]
- Li, Q.M.; Li, X.; Su, S.Q.; Wang, Y.T.; Xu, T.; Zha, X.Q.; Pan, L.H.; Shang, Z.Z.; Zhang, F.Y.; Luo, J.P. Dendrobine inhibits dopaminergic neuron apoptosis via MANF-mediated ER stress suppression in MPTP/MPP(+)-induced Parkinson’s disease models. Phytomedicine 2022, 102, 154193. [Google Scholar] [CrossRef]
- Galli, E.; Planken, A.; Kadastik-Eerme, L.; Saarma, M.; Taba, P.; Lindholm, P. Increased Serum Levels of Mesencephalic Astrocyte-Derived Neurotrophic Factor in Subjects with Parkinson’s Disease. Front. Neurosci. 2019, 13, 929. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.C.; Qi, X.H.; Fang, H.; Zhou, K.Q.; Wang, Q.S.; Chen, G.H. Increased MANF Expression in the Inferior Temporal Gyrus in Patients With Alzheimer Disease. Front. Aging Neurosci. 2021, 13, 639318. [Google Scholar] [CrossRef] [PubMed]
- GBD 2019 Stroke Collaborators. Global, regional, and national burden of stroke and its risk factors, 1990–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet Neurol. 2021, 20, 795–820. [Google Scholar] [CrossRef] [PubMed]
- Campbell, B.C.V.; De Silva, D.A.; Macleod, M.R.; Coutts, S.B.; Schwamm, L.H.; Davis, S.M.; Donnan, G.A. Ischaemic stroke. Nat. Rev. Dis. Primers 2019, 5, 70. [Google Scholar] [CrossRef]
- Li, T.; Xu, W.; Gao, L.; Guan, G.; Zhang, Z.; He, P.; Xu, H.; Fan, L.; Yan, F.; Chen, G. Mesencephalic astrocyte-derived neurotrophic factor affords neuroprotection to early brain injury induced by subarachnoid hemorrhage via activating Akt-dependent prosurvival pathway and defending blood-brain barrier integrity. FASEB J. 2019, 33, 1727–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matlik, K.; Anttila, J.E.; Kuan-Yin, T.; Smolander, O.P.; Pakarinen, E.; Lehtonen, L.; Abo-Ramadan, U.; Lindholm, P.; Zheng, C.; Harvey, B.; et al. Poststroke delivery of MANF promotes functional recovery in rats. Sci. Adv. 2018, 4, eaap8957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belayev, L.; Hong, S.H.; Freitas, R.S.; Menghani, H.; Marcell, S.J.; Khoutorova, L.; Mukherjee, P.K.; Reid, M.M.; Oria, R.B.; Bazan, N.G. DHA modulates MANF and TREM2 abundance, enhances neurogenesis, reduces infarct size, and improves neurological function after experimental ischemic stroke. CNS Neurosci. Ther. 2020, 26, 1155–1167. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Deng, J.; Zhang, X.; Sun, H.; Jia, G.; Li, J.; Zhang, K.; Wan, C.; Wang, L.; Yan, L.J.; et al. Effects of mesencephalic astrocyte-derived neurotrophic factor on cerebral angiogenesis in a rat model of cerebral ischemia. Neurosci. Lett. 2020, 715, 134657. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, A.; Duraikannu, A.; Zochodne, D.W. Releasing ‘brakes’ to nerve regeneration: Intrinsic molecular targets. Eur. J. Neurosci. 2016, 43, 297–308. [Google Scholar] [CrossRef]
- Zhang, J.; Tong, W.; Sun, H.; Jiang, M.; Shen, Y.; Liu, Y.; Gu, H.; Guo, J.; Fang, J.; Jin, L. Nrf2-mediated neuroprotection by MANF against 6-OHDA-induced cell damage via PI3K/AKT/GSK3β pathway. Exp. Gerontol. 2017, 100, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Li, F.; Zhang, H.; Ji, C.; Shu, Q.; Wang, C.; Ni, H.; Zhu, Y.; Wang, S. Mesencephalic astrocyte-derived neurotrophic factor restores blood-brain barrier integrity of aged mice after ischaemic stroke/reperfusion through anti-inflammation via TLR4/MyD88/NF-kappaB pathway. J. Drug Target 2022, 30, 430–441. [Google Scholar] [CrossRef]
- Zhang, J.X.; Tong, W.F.; Jiang, M.; Zhou, K.G.; Xiang, X.R.; He, Y.J.; Zhang, Z.Y.; Guan, Q.; Jin, L.J. MANF Inhibits α-Synuclein Accumulation through Activation of Autophagic Pathways. Oxid. Med. Cell. Longev. 2022, 2022, 7925686. [Google Scholar] [CrossRef]
- Dobson, R.; Giovannoni, G. Multiple sclerosis—A review. Eur. J. Neurol. 2019, 26, 27–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, J.; Koppinen, T.K.; Voutilainen, M.H. MANF Is Neuroprotective in Early Stages of EAE, and Elevated in Spinal White Matter by Treatment With Dexamethasone. Front. Cell. Neurosci. 2021, 15, 640084. [Google Scholar] [CrossRef]
- Lindholm, P.; Peranen, J.; Andressoo, J.O.; Kalkkinen, N.; Kokaia, Z.; Lindvall, O.; Timmusk, T.; Saarma, M. MANF is widely expressed in mammalian tissues and differently regulated after ischemic and epileptic insults in rodent brain. Mol. Cell. Neurosci. 2008, 39, 356–371. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Wang, Y.; Li, H.; Xu, H.; Xu, M.; Frank, J.A.; Ma, M.; Luo, J. Mesencephalic Astrocyte-Derived Neurotrophic Factor (MANF) Regulates Neurite Outgrowth Through the Activation of Akt/mTOR and Erk/mTOR Signaling Pathways. Front. Mol. Neurosci. 2020, 13, 560020. [Google Scholar] [CrossRef]
- Tseng, K.Y.; Danilova, T.; Domanskyi, A.; Saarma, M.; Lindahl, M.; Airavaara, M. MANF Is Essential for Neurite Extension and Neuronal Migration in the Developing Cortex. eNeuro 2017, 4, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Yagi, T.; Asada, R.; Kanekura, K.; Eesmaa, A.; Lindahl, M.; Saarma, M.; Urano, F. Neuroplastin Modulates Anti-inflammatory Effects of MANF. iScience 2020, 23, 101810. [Google Scholar] [CrossRef]
- Christie, K.J.; Krishnan, A.; Martinez, J.A.; Purdy, K.; Singh, B.; Eaton, S.; Zochodne, D. Enhancing adult nerve regeneration through the knockdown of retinoblastoma protein. Nat. Commun. 2014, 5, 3670. [Google Scholar] [CrossRef] [Green Version]
- Krishnan, A.; Dwivedi, S.; Chandrasekhar, A.; Areti, A.; Zochodne, D.W. In vitro priming response in dorsal root ganglia partially mimics injury-driven pre-conditioning response and reprograms neurons for enhanced outgrowth. Mol. Cell. Neurosci. 2021, 110, 103573. [Google Scholar] [CrossRef] [PubMed]
- Bautista, M.; Katselis, G.S.; Chowdhury, B.; Chumala, P.; Mahendra, R.; Desai, P.; Hall, J.; Kalyaanamoorthy, S.; Krishnan, A. Comparative Proteomics Analysis of Growth-Primed Adult Dorsal Root Ganglia Reveals Key Molecular Mediators for Peripheral Nerve Regeneration. eNeuro 2023, 10, 1–15. [Google Scholar] [CrossRef]
- Krishnan, A. Neuregulin-1 type I: A hidden power within Schwann cells for triggering peripheral nerve remyelination. Sci. Signal 2013, 6, jc1. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| MANF in PD | MANF in AD | MANF in STROKE |
|---|---|---|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sivakumar, B.; Krishnan, A. Mesencephalic Astrocyte-Derived Neurotrophic Factor (MANF): An Emerging Therapeutic Target for Neurodegenerative Disorders. Cells 2023, 12, 1032. https://doi.org/10.3390/cells12071032
Sivakumar B, Krishnan A. Mesencephalic Astrocyte-Derived Neurotrophic Factor (MANF): An Emerging Therapeutic Target for Neurodegenerative Disorders. Cells. 2023; 12(7):1032. https://doi.org/10.3390/cells12071032
Chicago/Turabian StyleSivakumar, Bhadrapriya, and Anand Krishnan. 2023. "Mesencephalic Astrocyte-Derived Neurotrophic Factor (MANF): An Emerging Therapeutic Target for Neurodegenerative Disorders" Cells 12, no. 7: 1032. https://doi.org/10.3390/cells12071032