
Isolated Peptide from Spider Venom Modulates Dendritic Cells In Vitro: A Possible Application in Oncoimmunotherapy for Glioblastoma
 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Ethical Considerations
2.2. Reagents and Phoneutria nigriventer Venom (PnV)
2.3. Venom Purification
2.4. Obtaining DCs
2.5. Culture of Human (NG97) and Murine (GL261) GB Cells and Obtaining Lysate
2.6. Treatment of DCs In Vitro
2.7. Cell Viability Test (MTT)
2.8. Detection of Surface Markers by Flow Cytometry
3. Measurement of Cytokine Production by Enzyme-Linked Immunosorbent Assay (ELISA)
3.1. Morphological Analysis
3.2. Statistical Analysis
4. Results
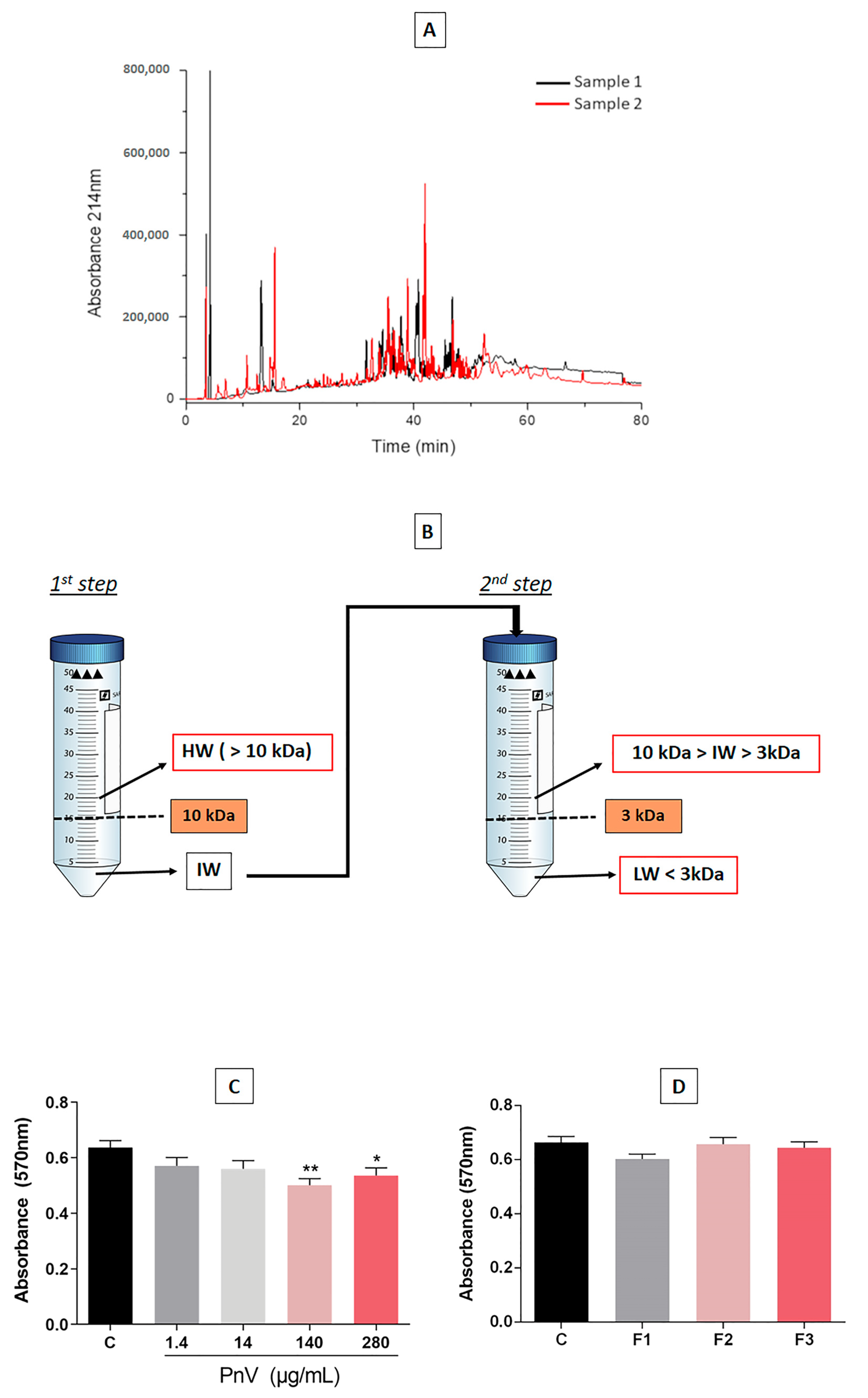
4.1. Reproducibility of the Venom, Purifications and Toxicity of Fractions
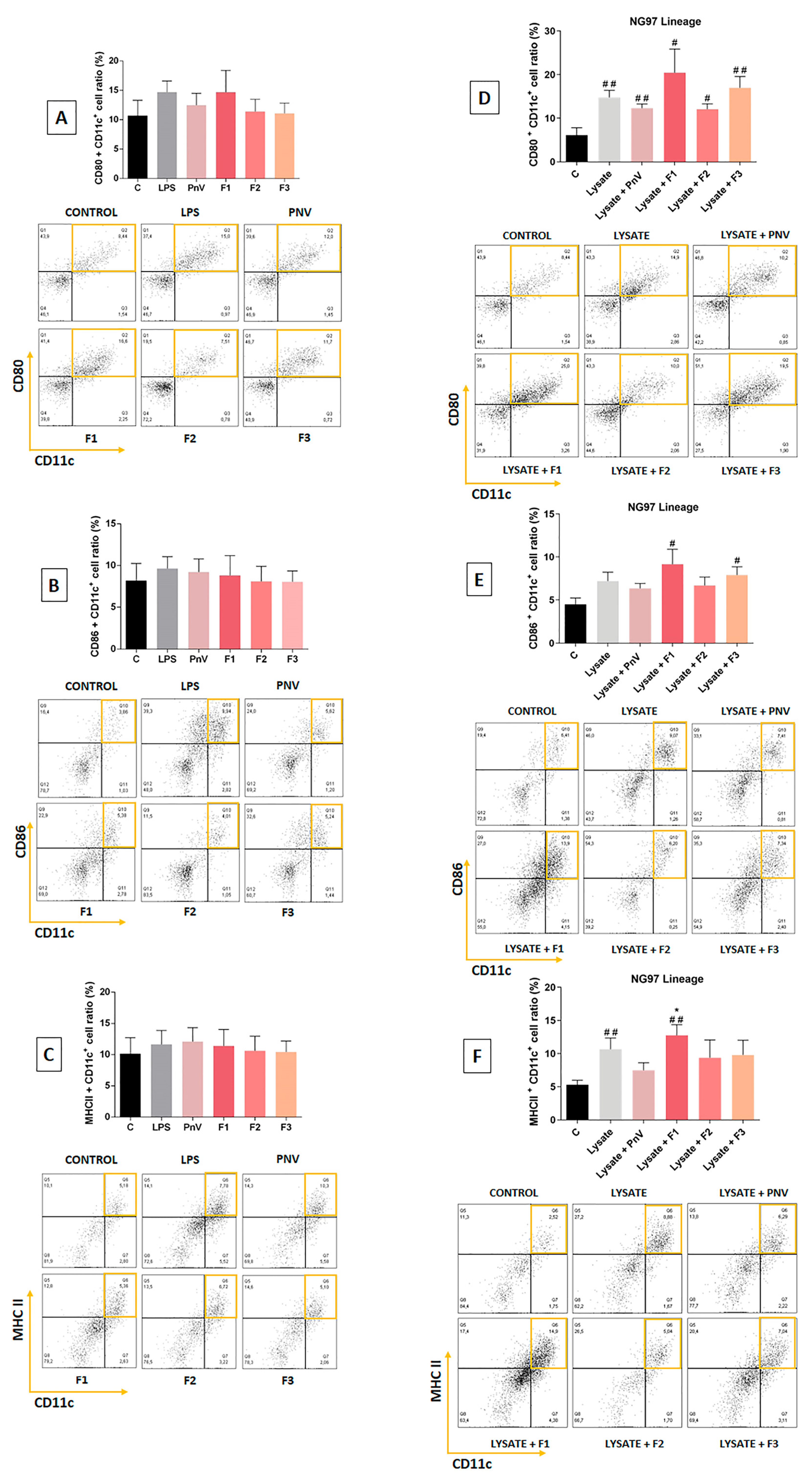
4.2. F1 Increased Surface Molecules of DCs When Used Concomitantly with the NG97 Cells Lysate
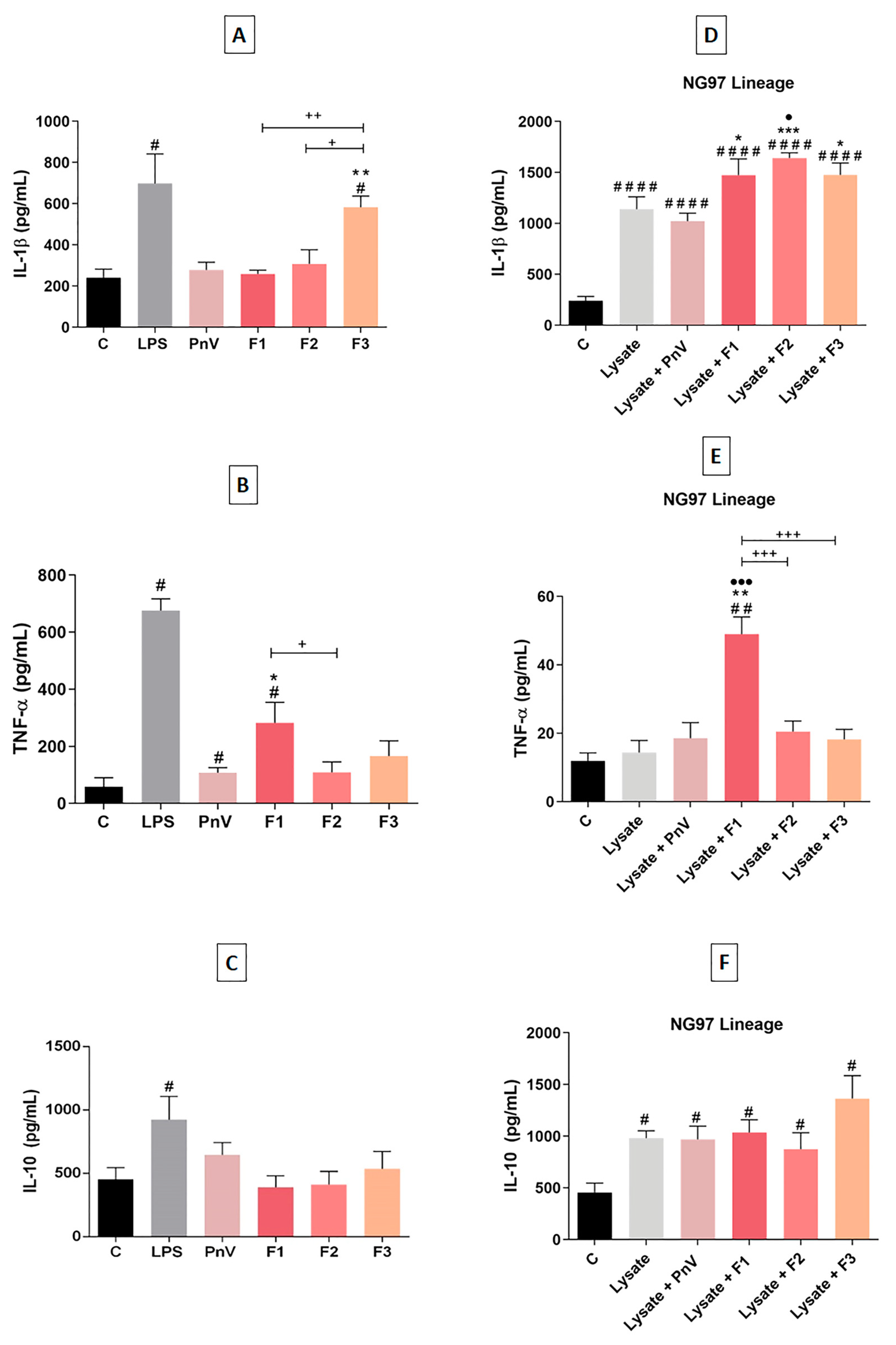
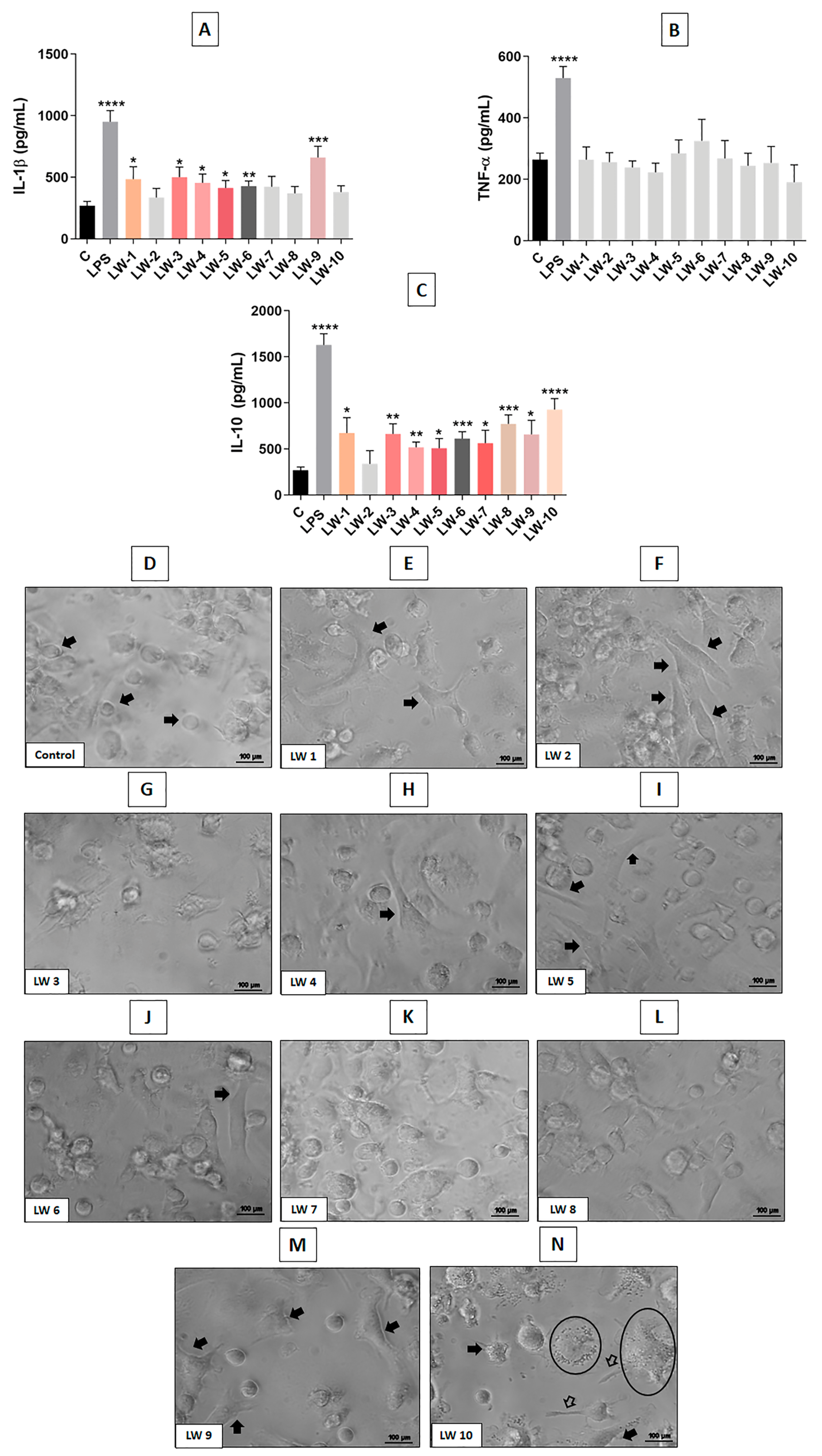
4.3. F1 and F2 Increased Cytokines Release When Used with Simultaneous NG97 Cells Lysate
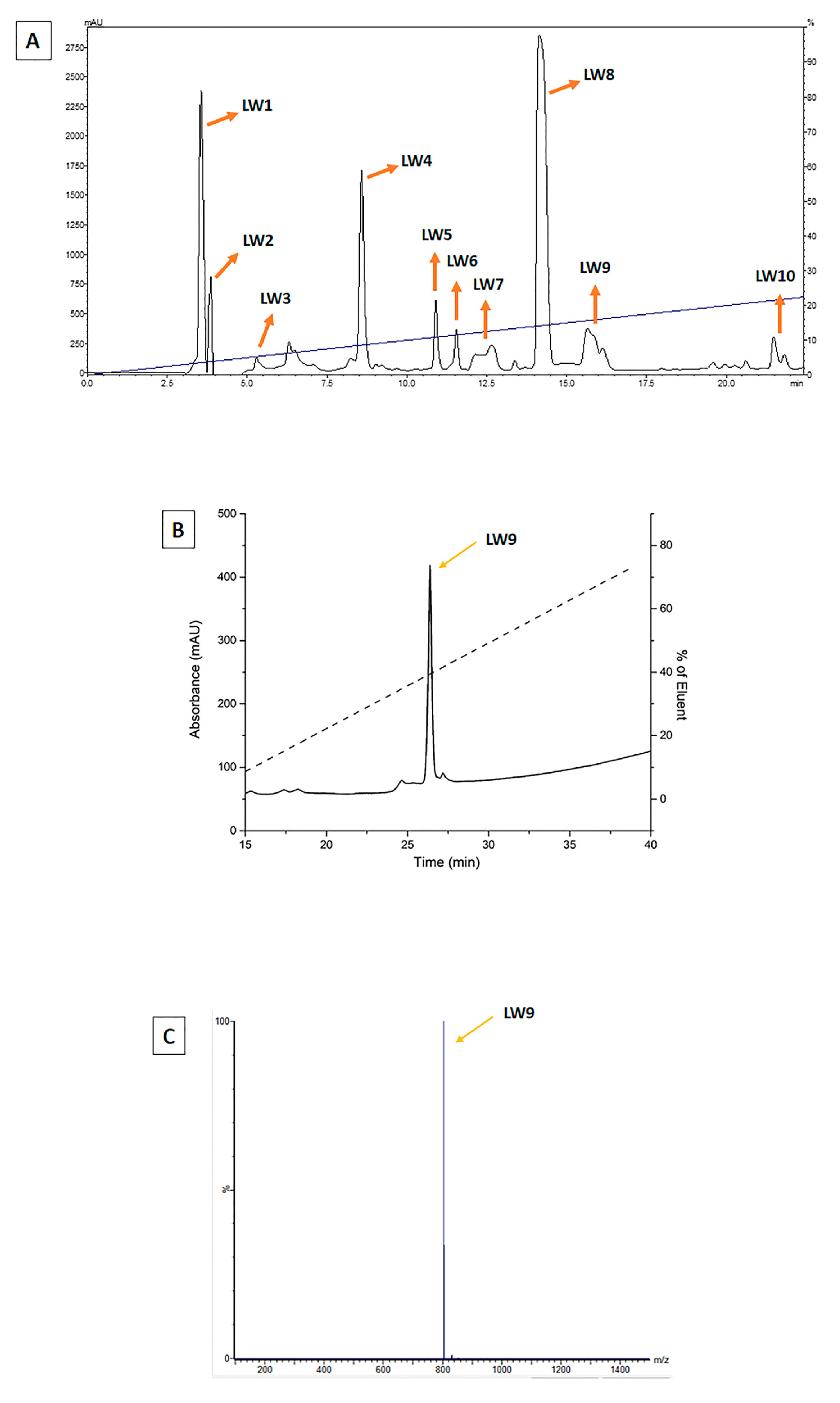
4.4. Evaluation and Selection of Molecules Purified from F1
5. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Balkwill, F.; Mantovani, A. Inflammation and cancer: Back to Virchow? Lancet 2001, 357, 539–545. [Google Scholar] [CrossRef] [PubMed]
- Pulendran, B. The varieties of immunological experience: Of pathogens, stress, and dendritic cells. Annu. Rev. Immunol. 2015, 33, 563–606. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Boltjes, A.; Van Wijk, F. Human dendritic cell functional specialization in steady-state and inflammation. Front. Immunol. 2014, 5, 131. [Google Scholar] [CrossRef] [Green Version]
- Steinman, R.M.; Banchereau, J. Taking dendritic cells into medicine. Nature 2007, 449, 419–426. [Google Scholar] [CrossRef]
- Dana, H.; Chalbatani, G.M.; Jalali, S.A.; Mirzaei, H.R.; Grupp, S.A.; Suarez, E.R.; Rapôso, C.; Webster, T.J. CAR-T cells: Early successes in blood cancer and challenges in solid tumors. Acta Pharm. Sin. B 2021, 5, 1129–1147. [Google Scholar] [CrossRef]
- Palucka, K.; Banchereau, J. Dendritic-cell-based therapeutic cancer vaccines. Immunity 2013, 39, 38–48. [Google Scholar] [CrossRef] [Green Version]
- Koido, S.; Homma, S.; Takahara, A.; Namiki, Y.; Tsukinaga, S.; Mitobe, J.; Odahara, S.; Yukawa, T.; Matsudaira, H.; Nagatsuma, K.; et al. Current immunotherapeutic approaches in pancreatic cancer. Clin. Dev. Immunol. 2011, 2011, 267539. [Google Scholar] [CrossRef]
- Van Brussel, I.; Berneman, Z.N.; Cools, N. Optimizing dendritic cell-based immunotherapy: Tackling the complexity of different arms of the immune system. Mediat. Inflamm. 2012, 2012, 690643. [Google Scholar] [CrossRef] [Green Version]
- Raposo, C. Scorpion and spider venoms in cancer treatment: State of the art, challenges, and perspectives. J. Clin. Transl. Res. 2017, 3, 233–249. [Google Scholar] [CrossRef]
- Mahadevappa, R.; Ma, R.; Kwok, H.F. Venom Peptides: Improving Specificity in Cancer Therapy. Trends Cancer 2017, 9, 611–614. [Google Scholar] [CrossRef] [PubMed]
- Wulff, H.; Christophersen, P.; Colussi, P.; Chandy, K.G.; Yarov-Yarovoy, V. Antibodies and venom peptides: New modalities for ion channels. Nat. Rev. Drug Discov. 2019, 18, 339–357. [Google Scholar] [CrossRef] [PubMed]
- Santos, N.B.; Bonfanti, A.P.; Rocha-E-Silva, T.A.A.D.; Silva, P.I.; Cruz-Höfling, M.A.; Verinaud, L.; Rapôso, C. Venom of the Phoneutria nigriventer spider alters the cell cycle, viability, and migration of cancer cells. J. Cell. Physiol. 2019, 234, 1398–1415. [Google Scholar] [CrossRef] [PubMed]
- Barreto, N.; Caballero, M.; Bonfanti, A.P.; de Mato, F.C.P.; Munhoz, J.; da Rocha-E-Silva, T.A.A.; Sutti, R.; Vitorino-Araujo, J.L.; Verinaud, L.; Rapôso, C. Spider venom components decrease glioblastoma cell migration and invasion through RhoA-ROCK and Na+/K+-ATPase β2: Potential molecular entities to treat invasive brain cancer. Cancer Cell Int. 2020, 20, 576. [Google Scholar] [CrossRef] [PubMed]
- Bonfanti, A.P.; Barreto, N.; Munhoz, J.; Caballero, M.; Cordeiro, G.; Rocha-E-Silva, T.; Sutti, R.; Moura, F.; Brunetto, S.; Ramos, C.D.; et al. Spider venom administration impairs glioblastoma growth and modulates immune response in a non-clinical model. Sci. Rep. 2020, 10, 5876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munhoz, J.; Peron, G.; Bonfanti, A.P.; Oliveira, J.; da Rocha-E-Silva, T.A.A.; Sutti, R.; Thomé, R.; Bombeiro, A.L.; Barreto, N.; Chalbatani, G.M.; et al. Components from spider venom activate macrophages against glioblastoma cells: New potential adjuvants for anticancer immunotherapy. J. Biochem. 2021, 170, 51–68. [Google Scholar] [CrossRef]
- Na, Y.R.; Jung, D.; Gu, G.J.; Seok, S.H. Grown Bone Marrow Derived Cells Are Composed of Phenotypically Different Dendritic Cells and Macrophages. Mol. Cells. 2016, 39, 734–741. [Google Scholar] [CrossRef] [Green Version]
- Machado, C.M.L.; Zorzeto, T.Q.; Bianco, J.E.R.; Rosa, R.G.; Genari, S.C.; Joazeiro, P.P.; Verinaud, L. Ultrastructural characterization of the new NG97ht human-derived glioma cell line using two different electron microscopy technical procedures. Microsc. Res. Tech. 2009, 72, 310–316. [Google Scholar] [CrossRef]
- Machado, C.M.; Ilkemori, R.Y.; Zorzeto, T.Q.; Nogueira, A.C.; Barbosa, S.D.; Savino, W.; Schenk, A.A.; Vassallo, J.; Henrich, J.K.; Boetcher-Luiz, F.; et al. Characterization of cells recovered from the xenotransplanted NG97 human-derived glioma cell line subcultured in a long-term in vitro. BMC Cancer 2008, 8, 291. [Google Scholar] [CrossRef] [Green Version]
- Schenka, A.A.; Machado, C.M.L.; Grippo, M.C.; Queiroz, L.S.; Schenka, N.G.M.; Chagas, C.A.; Verinaud, L.; Brousset, P.; Vassalo, J. Immunophenotypic and ultrastructural validation of a new human glioblastoma cell line. Cell. Mol. Neurobiol. 2005, 25, 929–941. [Google Scholar] [CrossRef]
- Machado, C.M.; Schenka, A.; Vassalo, J.; Tamashiro, W.M.; Gonçalves, E.M.; Genari, S.C.; Verinaud, L. Morphological characterization of a human glioma cell l ine. Cancer Cell Int. 2005, 5, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rainone, V.; Martelli, C.; Ottobrini, L.; Biasin, M.; Texido, G.; Degrassi, A.; Borelli, M.; Lucignani, G.; Trabattoni, D.; Clerici, M. Immunological Characterization of Whole Tumour Lysate-Loaded Dendritic Cells for Cancer Immunotherapy. PLoS ONE 2016, 11, e0146622. [Google Scholar] [CrossRef] [PubMed]
- Nagayama, H.; Sato, K.; Morishita, M.; Uchimaru, K.; Oyaizu, N.; Inazawa, T.; Yamasaki, T.; Enomoto, M.; Nakaoka, T.; Nakamura, T.; et al. Results of a phase I clinical study using autologous tumour lysate-pulsed monocyte-derived mature dendritic cell vaccinations for stage IV malignant melanoma patients combined with low dose interleukin-2. Melanoma Res. 2003, 13, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Trefzer, U.; Herberth, G.; Wohlan, K.; Milling, A.; Thiemann, M.; Sharav, T.; Sparbier, K.; Sterry, W.; Walden, P.T. Tumour-dendritic hybrid cell vaccination for the treatment of patients with malignant melanoma: Immunological effects and clinical results. Vaccine 2005, 23, 2367–2373. [Google Scholar] [CrossRef] [PubMed]
- Ridolfi, R.; Petrini, M.; Fiammenghi, L.; Stefanelli, M.; Ridolfi, L.; Ridolfi, R.; Ballardini, M.; Migliori, G.; Riccobon, A. Improved overall survival in dendritic cell vaccination-induced immunoreactive subgroup of advanced melanoma patients. J. Transl. Med. 2006, 4, 36. [Google Scholar] [CrossRef] [Green Version]
- Liau, L.M.; Ashkan, K.; Tran, D.D.; Campian, J.L.; Trusheim, J.E.; Cobbs, C.S.; Heth, A.J.; Salacz, M.; Taylor, S.; D’andre, S.D.; et al. First results on survival from a large Phase 3 clinical trial of an autologous dendritic cell vaccine in newly diagnosed glioblastoma. J. Transl. Med. 2018, 16, 142. [Google Scholar] [CrossRef] [Green Version]
- Herbert, G.S.; Vreeland, T.J.; Clifton, G.T.; Greene, J.M.; Jackson, D.O.; Hardin, M.O.; Hale, D.F.; Berry, J.S.; Nichol, P.; Yin, S.; et al. Initial phase I/IIa trial results of an autologours tumor lysate, particle-loaded, dendritic cell (TLPLDC) vaccine in patients with solid tumors. Vaccine 2018, 36, 3247–3253. [Google Scholar] [CrossRef]
- Rodríguez-Ruiz, M.E.; Perez-Gracia, J.L.; Rodríguez, I.; Alfaro, C.; Oñate, C.; Pérez, G.; Gil-Bazo, I.; Benito, A.; Inogés, S.; López-Diaz de Cerio, A.; et al. Combined immunotherapy emcompassing intratumoral poly-ICLC, dendritic-cell vaccination and radiotherapy in advanced cancer patients. Ann. Oncol. 2018, 29, 1312–1319. [Google Scholar] [CrossRef]
- Kheirkhah, A.; Darabad, R.R.; Cruzat, A.; Hairasouliha, A.R.; Witkin, D.; Wong, N.; Dana, R.; Hamrah, P. Corneal epithelial immune dendritic cell alterations in subtypes of dry eye disease: A pilot in vivo confocal microscopic study. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7179–7185. [Google Scholar] [CrossRef]
- Hubo, M.; Trinschek, B.; Kryczanowsky, F.; Tuettenberg, A.; Steinbrink, K.; Jonuleit, H. Costimulatory molecules on immunogenic versus tolerogenic human dendritic cells. Front. Immunol. 2013, 4, 82. [Google Scholar] [CrossRef] [Green Version]
- Hussein, M.R. Dendritic cells and melanoma tumorigenesis: An insight. Cancer Biol. Ther. 2005, 4, 501–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, N.C.; Lee, J.H.; Chung, K.H.; Kwak, Y.S.; Lim, D.S. Dendritic cell-based immunoteraphy for solid tumors. Transl. Oncol. 2018, 11, 686–690. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Tan, J.; Li, X.; Li, H.; Wu, W.; Wu, Y.; Zhang, J.; Gu, L. Myeloid and plasmacytoid dendritic cell combined vaccines loaded with heat-treated tumor cell lysates enhance antitumor activity in murine lung cancer. Oncol. Lett. 2021, 21, 1. [Google Scholar] [CrossRef] [PubMed]
- Tagliamonte, M.; Petrizzo, A.; Tornesello, M.L.; Buonaguro, F.M.; Buonaguro, L. Antigen-specific vaccines for cancer treatment. Hum. Vaccin. Immunother. 2014, 10, 3332–3346. [Google Scholar] [CrossRef] [PubMed]
- Schuler, G.; Schuler-Thurner, B.; Steinman, R.M. The use of dendritic cells in cancer immunotherapy. Curr. Opin. Immunol. 2003, 15, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Mocellin, S.; Mandruzzato, S.; Bronte, V.; Lise, M.; Nitti, D. Part1 I: Vaccines for solid tumours. Lancet Oncol. 2004, 5, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Audiger, C.; Rahman, M.J.; Yun, T.J.; Tarbell, K.V.; Lesage, S. The Importance of Dendritic Cells in Maintaining Immune Tolerance. J. Immunol. 2017, 198, 2223–2231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dranoff, G. Cytokines in cancer pathogenesis and cancer therapy. Nat. Rev. Cancer 2004, 4, 11–12. [Google Scholar] [CrossRef]
- Bedoui, S.; Heath, W.R.; Mueller, S.N. CD4(+) T-cell help amplifies innate signals for primary CD8(+) T-cell immunity. Immunol. Rev. 2016, 272, 52–64. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Mato, F.C.; Barreto, N.; Cordeiro, G.; Munhoz, J.; Bonfanti, A.P.; da Rocha-e-Silva, T.A.A.; Sutti, R.; Cruz, P.B.M.; Sanches, L.R.; Bombeiro, A.L.; et al. Isolated Peptide from Spider Venom Modulates Dendritic Cells In Vitro: A Possible Application in Oncoimmunotherapy for Glioblastoma. Cells 2023, 12, 1023. https://doi.org/10.3390/cells12071023
de Mato FC, Barreto N, Cordeiro G, Munhoz J, Bonfanti AP, da Rocha-e-Silva TAA, Sutti R, Cruz PBM, Sanches LR, Bombeiro AL, et al. Isolated Peptide from Spider Venom Modulates Dendritic Cells In Vitro: A Possible Application in Oncoimmunotherapy for Glioblastoma. Cells. 2023; 12(7):1023. https://doi.org/10.3390/cells12071023
Chicago/Turabian Stylede Mato, Felipe Cezar, Natália Barreto, Gabriel Cordeiro, Jaqueline Munhoz, Amanda Pires Bonfanti, Thomaz A. A. da Rocha-e-Silva, Rafael Sutti, Priscilla B. M. Cruz, Livia R. Sanches, André Luis Bombeiro, and et al. 2023. "Isolated Peptide from Spider Venom Modulates Dendritic Cells In Vitro: A Possible Application in Oncoimmunotherapy for Glioblastoma" Cells 12, no. 7: 1023. https://doi.org/10.3390/cells12071023