

Electrophysiological Activity of Primary Cortical Neuron-Glia Mixed Cultures

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. MEA Design and Fabrication
2.2. Primary Cortical Culture
2.3. Immunocytochemistry
2.4. Image Analysis
2.5. Electrophysiological Recording and Analysis
2.6. Statistical Analysis
3. Results
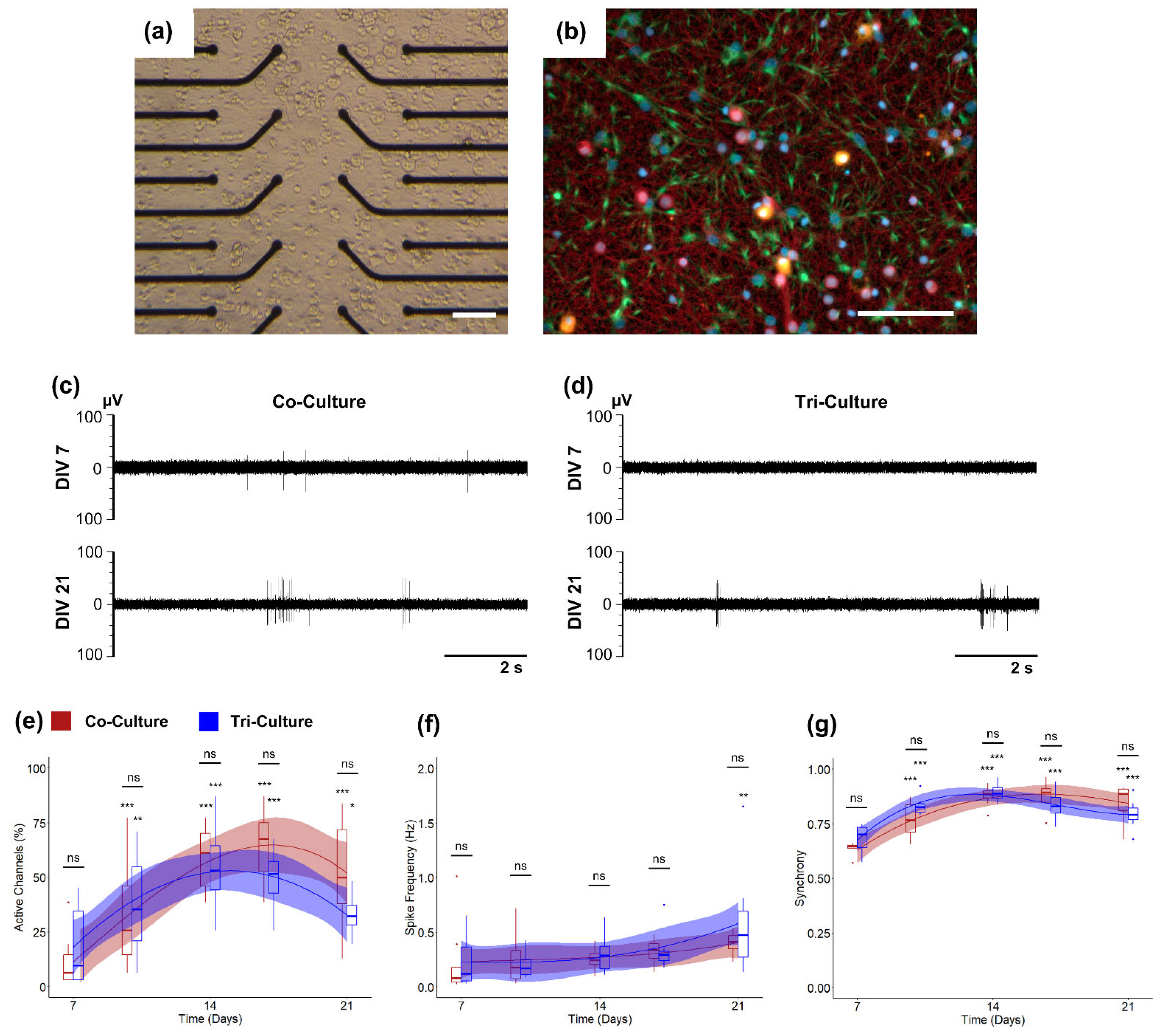
3.1. Electrophysiology
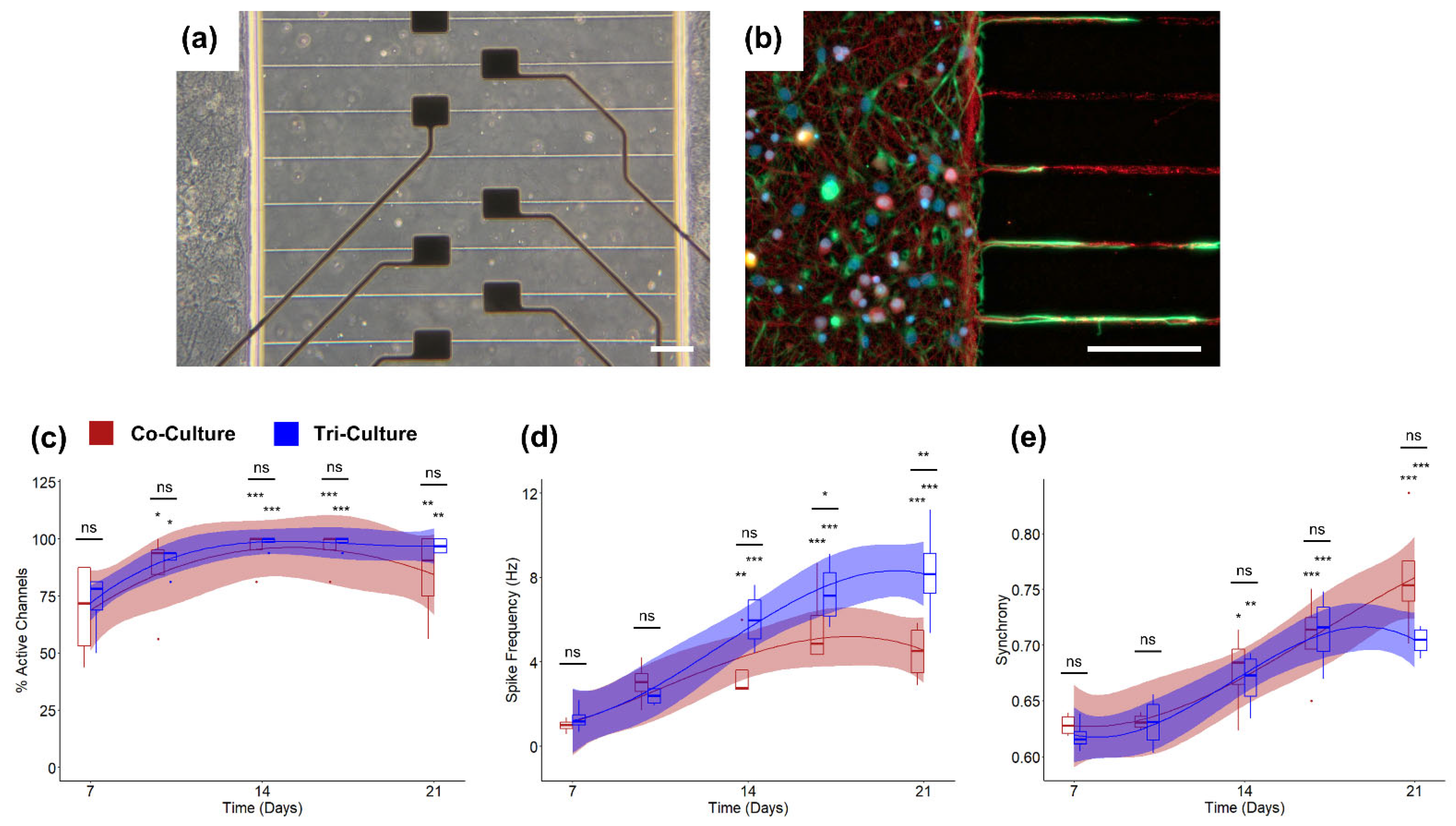
3.2. Incorporation into Microfluidic Platforms
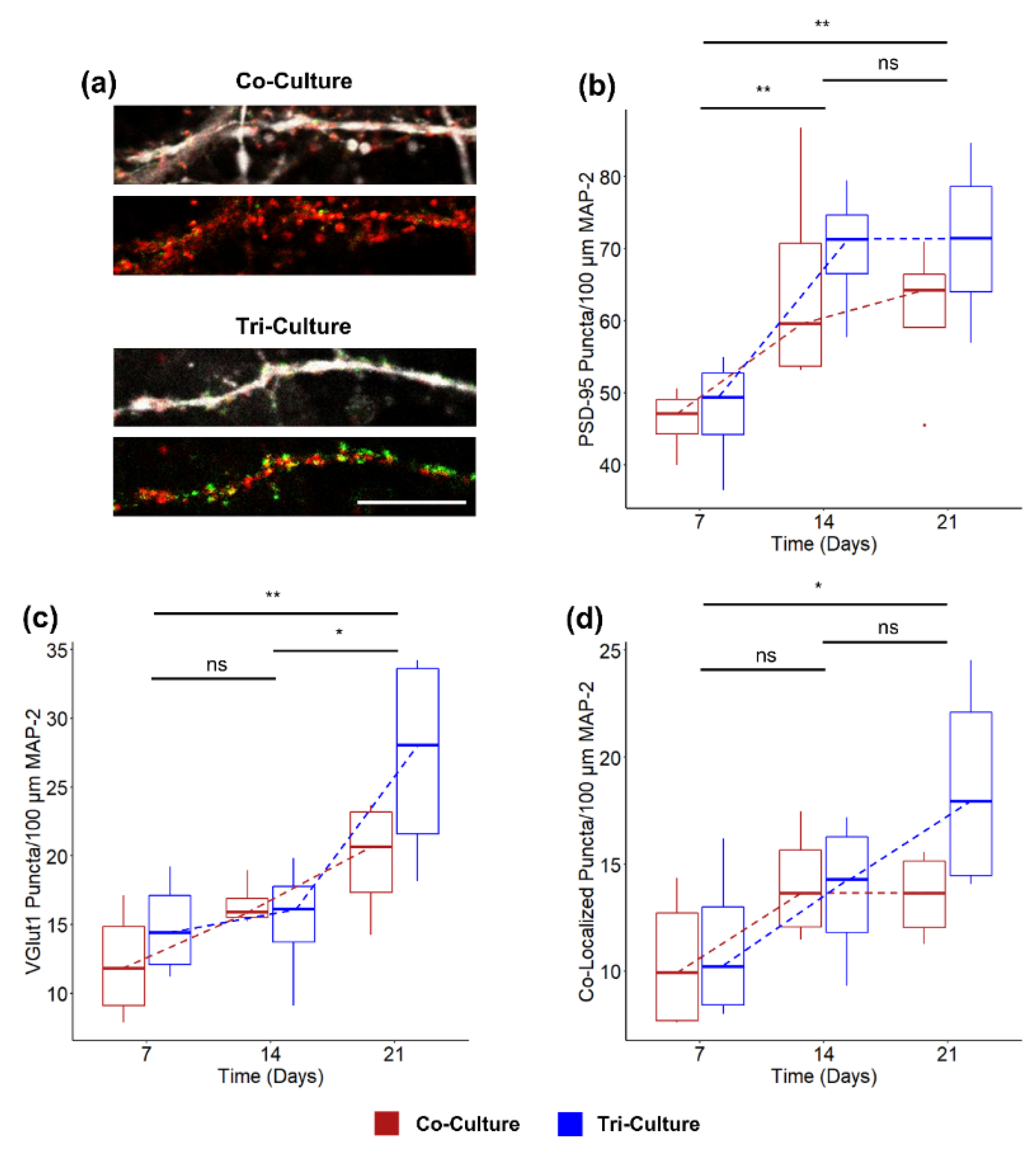
3.3. Synaptic Density
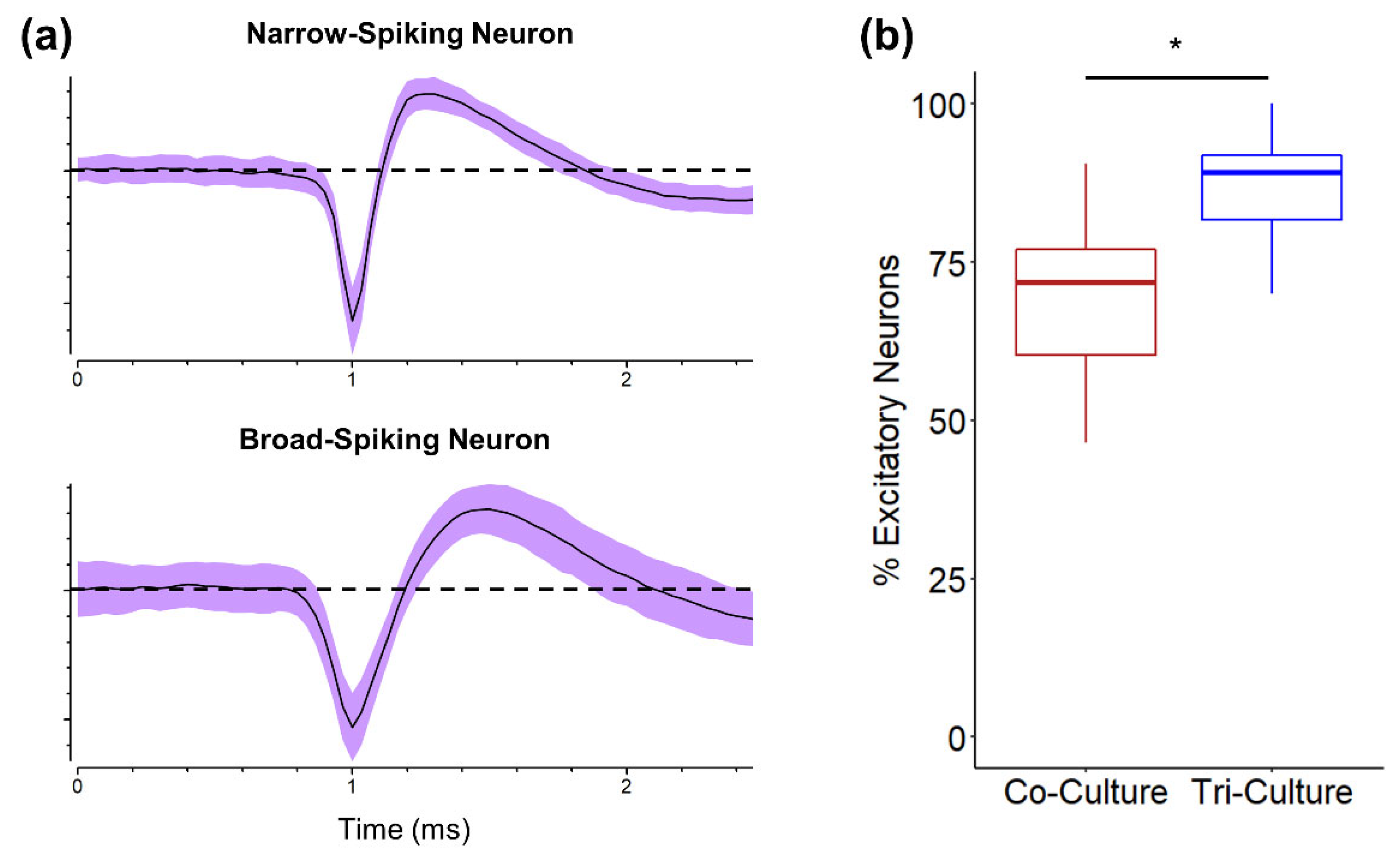
3.4. Excitatory vs. Inhibitory Neurons
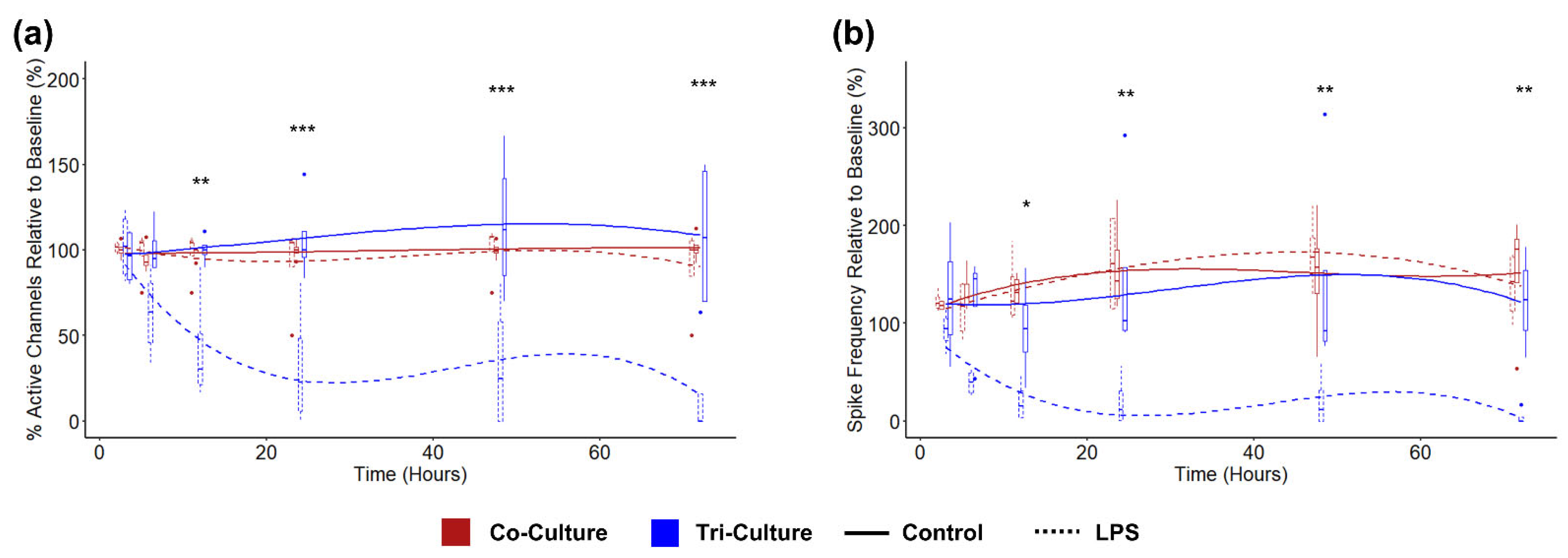
3.5. Influence of LPS Exposure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MEA | Microelectrode array |
| E/I ratio | Excitatory-to-inhibitory-neuron ratio |
| LPS | Lipopolysaccharide |
| CNS | Central nervous system |
| TLR | Toll-like receptor |
| DIV | Days in vitro |
| PDMS | Polydimethylsiloxane |
| PECVD | Plasma enhanced chemical vapor deposition |
| IL | Interleukin |
| TGF-β | Transforming growth factor beta |
| DPBS+ | Dulbecco’s phosphate buffered saline solution with calcium and magnesium |
| PFA | Paraformaldehyde |
| PSD-95 | Postsynaptic Density Protein 95 |
| VGlut1 | Vesicular Glutamate Transporter 1 |
| MAP2 | Microtubule-associated Protein 2 |
| GFAP | Glial fibrillary acidic protein |
| Iba-1 | Ionized Calcium-binding Adaptor Molecule 1 |
| DAPI | 4′,6-diamidino-2-phenylindole |
| SEM | Standard error of the mean |
| ANOVA | Analysis of variance |
| TLR4 | Toll-like Receptor 4 |
| PNN | Perineuronal nets |
References
- Naegele, M.; Martin, R. The Good and the Bad of Neuroinflammation in Multiple Sclerosis, 1st ed.; Handbook of Clinical Neurology; Elsevier B.V.: Amsterdam, The Netherlands, 2014. [Google Scholar] [CrossRef]
- Calsolaro, V.; Edison, P. Neuroinflammation in Alzheimer’s disease: Current evidence and future directions. Alzheimer’s Dement. 2016, 12, 719–732. [Google Scholar] [CrossRef] [PubMed]
- Hirsch, E.C.; Vyas, S.; Hunot, S. Neuroinflammation in Parkinson’s disease. Park. Relat. Disord. 2012, 18, 210–212. [Google Scholar] [CrossRef] [PubMed]
- Jayaraj, R.L.; Azimullah, S.; Beiram, R.; Jalal, F.Y.; Rosenberg, G.A. Neuroinflammation: Friend and foe for ischemic stroke. J. Neuroinflam. 2019, 16, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polikov, V.S.; Tresco, P.A.; Reichert, W.M. Response of brain tissue to chronically implanted neural electrodes. J. Neurosci. Methods 2005, 148, 1–18. [Google Scholar] [CrossRef]
- Li, Q.; Barres, B.A. Microglia and macrophages in brain homeostasis and disease. Nat. Rev. Immunol. 2017, 18, 225–242. [Google Scholar] [CrossRef]
- Simard, M.; Nedergaard, M. The neurobiology of glia in the context of water and ion homeostasis. Neuroscience 2004, 129, 877–896. [Google Scholar] [CrossRef]
- Potter, S.M.; DeMarse, T.B. A New Approach to Neural Cell Culture for Long-Term Studies. J. Neurosci. Methods 2001, 110, 17–24. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0165027001004125 (accessed on 26 September 2022). [CrossRef]
- Kim, R.; Joo, S.; Jung, H.; Hong, N.; Nam, Y. Recent trends in microelectrode array technology for in vitro neural interface platform. Biomed. Eng. Lett. 2014, 4, 129–141. [Google Scholar] [CrossRef]
- Frank, C.L.; Brown, J.P.; Wallace, K.; Mundy, W.R.; Shafer, T.J. Developmental neurotoxicants disrupt activity in cortical networks on microelectrode arrays: Results of screening 86 compounds during neural network formation. Toxicol. Sci. 2017, 160, 121–135. [Google Scholar] [CrossRef]
- Shafer, T.J.; Brown, J.P.; Lynch, B.; Davila-Montero, S.; Wallace, K.; Friedman, K.P. Evaluation of chemical effects on network formation in cortical neurons grown on microelectrode arrays. Toxicol. Sci. 2019, 169, 436–455. [Google Scholar] [CrossRef]
- Johnstone, A.F.M.; Gross, G.W.; Weiss, D.G.; Schroeder, O.H.U.; Gramowski, A.; Shafer, T.J. Microelectrode arrays: A physiologically based neurotoxicity testing platform for the 21st century. Neurotoxicology 2010, 31, 331–350. [Google Scholar] [CrossRef]
- Gopal, K.V. Neurotoxic effects of mercury on auditory cortex networks growing on microelectrode arrays: A preliminary analysis. Neurotoxicol. Teratol. 2003, 25, 69–76. [Google Scholar] [CrossRef]
- Novellino, A.; Scelfo, B.; Palosaari, T.; Price, A.; Sobanski, T.; Shafer, T.J.; Johnstone, A.F.; Gross, G.W.; Gramowski, A.; Schroeder, O.; et al. Development of micro-electrode array based tests for neurotoxicity: Assessment of interlaboratory reproducibility with neuroactive chemicals. Front. Neuroeng. 2011, 4, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Charkhkar, H.; Meyyappan, S.; Matveeva, E.; Moll, J.R.; McHail, D.G.; Peixoto, N.; Cliff, R.O.; Pancrazio, J.J. Amyloid beta modulation of neuronal network activity in vitro. Brain Res. 2015, 1629, 1–9. [Google Scholar] [CrossRef]
- Gao, F.; Gao, K.; He, C.; Liu, M.; Wan, H.; Wang, P. Multi-site dynamic recording for Aβ oligomers-induced Alzheimer’s disease in vitro based on neuronal network chip. Biosens. Bioelectron. 2019, 133, 183–191. [Google Scholar] [CrossRef]
- Amin, H.; Nieus, T.; Lonardoni, D.; Maccione, A.; Berdondini, L. High-resolution bioelectrical imaging of Aβ-induced network dysfunction on CMOS-MEAs for neurotoxicity and rescue studies. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Belle, A.M.; Enright, H.A.; Sales, A.P.; Kulp, K.; Osburn, J.; Kuhn, E.A.; Fischer, N.O.; Wheeler, E.K. Evaluation of in vitro neuronal platforms as surrogates for in vivo whole brain systems. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Chapman, C.A.R.; Chen, H.; Stamou, M.; Biener, J.; Biener, M.M.; Lein, P.J.; Seker, E. Nanoporous gold as a neural interface coating: Effects of topography, surface chemistry, and feature size. ACS Appl. Mater. Interfaces 2015, 7, 7093–7100. [Google Scholar] [CrossRef] [Green Version]
- Xiang, G.; Pan, L.; Huang, L.; Yu, Z.; Song, X.; Cheng, J.; Xing, W.; Zhou, Y. Microelectrode array-based system for neuropharmacological applications with cortical neurons cultured in vitro. Biosens. Bioelectron. 2007, 22, 2478–2484. [Google Scholar] [CrossRef]
- James, C.; Spence, A.; Dowell-Mesfin, N.; Hussain, R.; Smith, K.; Craighead, H.; Isaacson, M.; Shain, W.; Turner, J. Extracellular recordings from patterned neuronal networks using planar microelectrode arrays. IEEE Trans. Biomed. Eng. 2004, 51, 1640–1648. [Google Scholar] [CrossRef]
- Nam, Y.; Wheeler, B.C.; Heuschkel, M.O. Neural recording and stimulation of dissociated hippocampal cultures using microfabricated three-dimensional tip electrode array. J. Neurosci. Methods 2006, 155, 296–299. [Google Scholar] [CrossRef] [PubMed]
- Chapman, C.A.R.; Wang, L.; Chen, H.; Garrison, J.; Lein, P.J.; Seker, E. Nanoporous Gold Biointerfaces: Modifying Nanostructure to Control Neural Cell Coverage and Enhance Electrophysiological Recording Performance. Adv. Funct. Mater. 2016, 27, 1604631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enright, H.A.; Lam, D.; Sebastian, A.; Sales, A.P.; Cadena, J.; Hum, N.R.; Osburn, J.J.; Peters, S.K.G.; Petkus, B.; Soscia, D.A.; et al. Functional and transcriptional characterization of complex neuronal co-cultures. Sci. Rep. 2020, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Napoli, A.; Obeid, I. Comparative Analysis of Human and Rodent Brain Primary Neuronal Culture Spontaneous Activity Using Micro-Electrode Array Technology. J. Cell Biochem. 2016, 117, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Odawara, A.; Saitoh, Y.; Alhebshi, A.; Gotoh, M.; Suzuki, I. Long-term electrophysiological activity and pharmacological response of a human induced pluripotent stem cell-derived neuron and astrocyte co-culture. Biochem. Biophys. Res. Commun. 2014, 443, 1176–1181. [Google Scholar] [CrossRef] [Green Version]
- Heikkilä, T.J.; Ylä-Outinen, L.; Tanskanen, J.M.; Lappalainen, R.S.; Skottman, H.; Suuronen, R.; Mikkonen, J.E.; Hyttinen, J.A.; Narkilahti, S. Human embryonic stem cell-derived neuronal cells form spontaneously active neuronal networks in vitro. Exp. Neurol. 2009, 218, 109–116. [Google Scholar] [CrossRef]
- Soscia, D.A.; Lam, D.; Tooker, A.C.; Enright, H.A.; Triplett, M.; Karande, P.; Peters, S.K.G.; Sales, A.P.; Wheeler, E.K.; Fischer, N.O. A flexible 3-dimensional microelectrode array for: In vitro brain models. Lab. Chip. R. Soc. Chem. 2020, 20, 901–911. [Google Scholar] [CrossRef] [Green Version]
- Shin, H.; Jeong, S.; Lee, J.-H.; Sun, W.; Choi, N.; Cho, I.-J. 3D high-density microelectrode array with optical stimulation and drug delivery for investigating neural circuit dynamics. Nat. Commun. 2021, 12, 1–18. [Google Scholar] [CrossRef]
- Dauth, S.; Maoz, B.M.; Sheehy, S.P.; Hemphill, M.A.; Murty, T.; Macedonia, M.K.; Greer, A.M.; Budnik, B.; Parker, K.K. Neurons derived from different brain regions are inherently different in vitro: A novel multiregional brain-on-a-chip. J. Neurophysiol. 2017, 117, 1320–1341. [Google Scholar] [CrossRef]
- Goshi, N.; Girardi, G.; Souza, F.D.C.; Gardner, A.; Lein, P.J.; Seker, E. Influence of microchannel geometry on device performance and electrophysiological recording fidelity during long-term studies of connected neural populations. Lab. Chip 2022, 22, 3961–3975. [Google Scholar] [CrossRef]
- Goshi, N.; Morgan, R.K.; Lein, P.J.; Seker, E. A primary neural cell culture model to study neuron, astrocyte, and microglia interactions in neuroinflammation. J. Neuroinflamm. 2020, 17, 1–16. [Google Scholar] [CrossRef]
- Goshi, N.; Kim, H.; Seker, E. Primary Cortical Cell Tri-Culture-Based Screening of Neuroinflammatory Response in Toll-like Receptor Activation. Biomedicines 2022, 10, 2122. [Google Scholar] [CrossRef]
- Guttenplan, K.A.; Liddelow, S.A. Astrocytes and Microglia: Models and Tools. J. Exp. Med. 2018, 216, 71–83. [Google Scholar] [CrossRef]
- Taylor, A.M.; Rhee, S.W.; Tu, C.H.; Cribbs, D.H.; Cotman, C.W.; Jeon, N.L. Microfluidic multicompartment device for neuroscience research. Langmuir 2003, 19, 1551–1556. [Google Scholar] [CrossRef] [Green Version]
- Bohlen, C.J.; Bennett, F.C.; Tucker, A.F.; Collins, H.Y.; Mulinyawe, S.B.; Barres, B.A. Diverse Requirements for Microglial Survival, Specification, and Function Revealed by Defined-Medium Cultures. Neuron 2017, 94, 759–773.e8. [Google Scholar] [CrossRef] [Green Version]
- Wayman, G.A.; Bose, D.D.; Yang, D.; Lesiak, A.; Bruun, D.; Impey, S.; Ledoux, V.; Pessah, I.N.; Lein, P.J. PCB-95 modulates the calcium-dependent signaling pathway responsible for activity-dependent dendritic growth. Environ. Health Perspect. 2012, 120, 1003–1009. [Google Scholar] [CrossRef] [Green Version]
- Kreuz, T.; Chicharro, D.; Houghton, C.; Andrzejak, R.G.; Mormann, F. Monitoring spike train synchrony. J. Neurophysiol. 2013, 109, 1457–1472. [Google Scholar] [CrossRef] [Green Version]
- Mulansky, M.; Kreuz, T. PySpike—A Python library for analyzing spike train synchrony. Softwarex 2016, 5, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, P.; Cotterill, E.; Morton, A.; Grant, S.G.; Eglen, S.J. Quantitative differences in developmental profiles of spontaneous activity in cortical and hippocampal cultures. Neural Dev. 2015, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sethi, S.; Keil, K.P.; Lein, P.J. 3,3’-Dichlorobiphenyl (PCB 11) Promotes Dendritic Arborization in Primary Rat Cortical Neurons via a CREB-Dependent Mechanism. Arch. Toxicol. 2018, 92, 3337–3345. [Google Scholar] [CrossRef] [Green Version]
- Toivanen, M.; Pelkonen, A.; Mäkinen, M.; Ylä-Outinen, L.; Sukki, L.; Kallio, P.; Ristola, M.; Narkilahti, S. Optimised PDMS tunnel devices on MEAs increase the probability of detecting electrical activity from human stem cell-derived neuronal networks. Front Neurosci. 2017, 11, 606. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Riss, M.; Buitrago, J.O.; Claverol-tintur, E. Biophysics of microchannel-enabled neuron-electrode interfaces. Neural Eng. 2012, 9, 26010. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Dissing-Olesen, L.; Stevens, B. New insights on the role of microglia in synaptic pruning in health and disease. Curr. Opin. Neurobiol. 2016, 36, 128–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crapser, J.D.; Arreola, M.A.; Tsourmas, K.I.; Green, K.N. Microglia as hackers of the matrix: Sculpting synapses and the extracellular space. Cell Mol. Immunol. 2021, 18, 2472–2488. [Google Scholar] [CrossRef]
- Andoh, M.; Koyama, R. Microglia regulate synaptic development and plasticity. Dev. Neurobiol. 2021, 81, 568–590. [Google Scholar] [CrossRef]
- Flavell, S.W.; Cowan, C.W.; Kim, T.-K.; Greer, P.L.; Lin, Y.; Paradis, S.; Griffith, E.C.; Hu, L.S.; Chen, C.; Greenberg, M.E. Activity-dependent regulation of MEF2 transcription factors suppresses excitatory synapse number. Science 2006, 311, 1008–1012. [Google Scholar] [CrossRef] [Green Version]
- Verstraelen, P.; Barriga, G.G.-D.; Verschuuren, M.; Asselbergh, B.; Nuydens, R.; Larsen, P.H.; Timmermans, J.-P.; De Vos, W.H. Systematic Quantification of Synapses in Primary Neuronal Culture. iScience 2020, 23, 101542. [Google Scholar] [CrossRef]
- Mitchell, J.F.; Sundberg, K.A.; Reynolds, J.H. Differential Attention-Dependent Response Modulation across Cell Classes in Macaque Visual Area V4. Neuron 2007, 55, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Ke, W.; Guang, J.; Chen, G.; Yin, L.; Deng, S.; He, Q.; Liu, Y.; He, T.; Zheng, R.; et al. Firing frequency maxima of fast-spiking neurons in human, monkey, and mouse neocortex. Front. Cell Neurosci. 2016, 10, 239. [Google Scholar] [CrossRef] [Green Version]
- Robbins, A.A.; Fox, S.E.; Holmes, G.L.; Scott, R.C.; Barry, J.M. Short duration waveforms recorded extracellularly from freely moving rats are representative of axonal activity. Front. Neural Circuits 2013, 7, 181. [Google Scholar] [CrossRef] [Green Version]
- Sahara, S.; Yanagawa, Y.; O’Leary, D.D.M.; Stevens, C.F. The fraction of cortical GABAergic neurons is constant from near the start of cortical neurogenesis to adulthood. J. Neurosci. 2012, 32, 4755–4761. [Google Scholar] [CrossRef] [Green Version]
- Jung, H.; Lee, S.Y.; Lim, S.; Choi, H.R.; Choi, Y.; Kim, M.; Kim, S.; Lee, Y.; Han, K.H.; Chung, W.-S.; et al. Anti-inflammatory clearance of amyloid-β by a chimeric Gas6 fusion protein. Nat. Med. 2022, 28, 1802–1812. [Google Scholar] [CrossRef]
- Jones, E.V.; Cook, D.; Murai, K.K. A Neuron-Astrocyte Co-Culture System to Investigate Astrocyte-Secreted Factors in Mouse Neuronal Development. In Astrocytes; Humana Press: Totoma, NJ, USA, 2012; pp. 341–352. ISBN 9781617794520. [Google Scholar]
- Ozog, M.A.; Siushansian, R.; Naus, C.C.G. Blocked Gap Junctional Coupling Increases Glutamate-Induced Neurotoxicity in Neuron-Astrocyte Co-Cultures. J. Neuropathol. Exp. Neurol. 2002, 61, 132–141. [Google Scholar] [CrossRef] [Green Version]
- Miyamoto, A.; Wake, H.; Ishikawa, A.W.; Eto, K.; Shibata, K.; Murakoshi, H.; Koizumi, S.; Moorhouse, A.J.; Yoshimura, Y.; Nabekura, J. Microglia contact induces synapse formation in developing somatosensory cortex. Nat. Commun. 2016, 7, 12540. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Dissing-Olesen, L.; MacVicar, B.A.; Stevens, B. Microglia: Dynamic Mediators of Synapse Development and Plasticity. Trends Immunol. 2015, 36, 605–613. [Google Scholar] [CrossRef] [Green Version]
- Spittau, B. Aging microglia-phenotypes, functions and implications for age-related neurodegenerative diseases. Front. Aging Neurosci. 2017, 9, 194. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Ma, N.; Yu, B.; Zhang, W.; Wan, J. Transcriptomic profiling of microglia and astrocytes throughout aging. J. Neuroinflam. 2020, 17, 97. [Google Scholar] [CrossRef] [Green Version]
- Luchena, C.; Zuazo-Ibarra, J.; Valero, J.; Matute, C.; Alberdi, E.; Capetillo-Zarate, E. A Neuron, Microglia, and Astrocyte Triple Co-culture Model to Study Alzheimer’s Disease. Front Aging Neurosci. 2022, 14, 271. [Google Scholar] [CrossRef]
- Phadke, L.; Lau, D.H.W.; Aghaizu, N.D.; Ibarra, S.; Navarron, C.M.; Granat, L.; Magno, L.; Whiting, P.; Jolly, S. A primary rodent triculture model to investigate the role of glia-neuron crosstalk in regulation of neuronal activity. Front. Aging Neurosci. 2022, 14, 1–16. [Google Scholar] [CrossRef]
- Von Bartheld, C.S.; Bahney, J.; Herculano-houzel, S. The Search for True Numbers of Neurons and Glial Cells in the Human Brain: A Review of 150 Years of Cell Counting. J. Comp. Neurol. 2016, 524, 3865–3895. [Google Scholar] [CrossRef] [Green Version]
- Pan, L.; Alagapan, S.; Franca, E.; Demarse, T.; Brewer, G.J.; Wheeler, B.C. Large extracellular spikes recordable from axons in microtunnels. IEEE Trans. Neural Syst. Rehabil. Eng. 2014, 22, 453–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biffi, E.; Regalia, G.; Menegon, A.; Ferrigno, G.; Pedrocchi, A. The influence of neuronal density and maturation on network activity of hippocampal cell cultures: A methodological study. PLoS ONE 2013, 8, e83899. [Google Scholar] [CrossRef] [PubMed]
- Wagenaar, D.A.; Pine, J.; Potter, S.M. An extremely rich repertoire of bursting patterns during the development of cortical cultures. BMC Neurosci. 2006, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.H.; Park, E.; You, B.; Jung, Y.; Park, A.R.; Park, S.G.; Lee, J.R. Neuronal synapse formation induced by microglia and interleukin 10. PLoS ONE 2013, 8, e81218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wake, H.; Moorhouse, A.J.; Jinno, S.; Kohsaka, S.; Nabekura, J. Resting microglia directly monitor the functional state of synapses in vivo and determine the fate of ischemic terminals. J. Neurosci. 2009, 29, 3974–3980. [Google Scholar] [CrossRef] [Green Version]
- Tremblay, M.Ě.; Lowery, R.L.; Majewska, A.K. Microglial interactions with synapses are modulated by visual experience. PLoS Biol. 2010, 8, e1000527. [Google Scholar] [CrossRef] [Green Version]
- Perez-Catalan, N.A.; Doe, C.Q.; Ackerman, S.D. The role of astrocyte-mediated plasticity in neural circuit development and function. Neural Dev. 2021, 16, 1. [Google Scholar] [CrossRef]
- Turko, P.; Groberman, K.; Browa, F.; Cobb, S.; Vida, I. Differential dependence of GABAergic and glutamatergic neurons on glia for the establishment of synaptic transmission. Cereb. Cortex 2019, 29, 1230–1243. [Google Scholar] [CrossRef] [Green Version]
- Miyata, S.; Nishimura, Y.; Hayashi, N.; Oohira, A. Construction of perineuronal net-like structure by cortical neurons in culture. Neuroscience 2005, 136, 95–104. [Google Scholar] [CrossRef]
- Paulk, A.C.; Kfir, Y.; Khanna, A.R.; Mustroph, M.L.; Trautmann, E.M.; Soper, D.J.; Stavisky, S.D.; Welkenhuysen, M.; Dutta, B.; Shenoy, K.V.; et al. Large-Scale Neural Recordings with Single Neuron Resolution Using Neuropixels Probes in Human Cortex. Nat. Neurosci. 2022, 25, 252–263. [Google Scholar] [CrossRef]
- Vigneswaran, G.; Kraskov, A.; Lemon, R.N. Large Identified Pyramidal Cells in Macaque Motor and Premotor Cortex Exhibit “Thin Spikes”: Implications for Cell Type Classification. J. Neurosci. 2011, 31, 14235–14242. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, Y.; Si, Q.; Kataoka, K. Lipopolysaccharide-induced microglial activation in culture: Temporal profiles of morphological change and release of cytokines and nitric oxide. Neurosci. Res. 1999, 35, 95–100. [Google Scholar] [CrossRef]
- Lehnardt, S.; Lachance, C.; Patrizi, S.; Lefebvre, S.; Follett, P.L.; Jensen, F.E.; Rosenberg, P.A.; Volpe, J.J.; Vartanian, T. The Toll-Like Receptor TLR4 Is Necessary for Lipopolysaccharide- Induced Oligodendrocyte Injury in the CNS. J. Neurosci. 2002, 22, 2478–2486. [Google Scholar] [CrossRef] [Green Version]
- Kothur, K.; Wienholt, L.; Brilot, F.; Dale, R.C. CSF cytokines/chemokines as biomarkers in neuroinflammatory CNS disorders: A systematic review. Cytokine 2016, 77, 227–237. [Google Scholar] [CrossRef]
- Nimmervoll, B.; White, R.; Yang, J.W.; An, S.; Henn, C.; Sun, J.J.; Luhmann, H.J. LPS-induced microglial secretion of TNFα increases activity-dependent neuronal apoptosis in the neonatal cerebral cortex. Cereb. Cortex 2013, 23, 1742–1755. [Google Scholar] [CrossRef]
- Wang, X.; Chen, S.; Ma, G.; Ye, M.; Lu, G. Involvement of proinflammatory factors, apoptosis, caspase-3 activation and Ca2+ disturbance in microglia activation-mediated dopaminergic cell degeneration. Mech. Ageing Dev. 2005, 126, 1241–1254. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Guttenplan, K.A.; Clarke, L.E.; Bennett, F.C.; Bohlen, C.J.; Schirmer, L.; Bennett, M.L.; Münch, A.E.; Chung, W.-S.; Peterson, T.C.; et al. Neurotoxic Reactive Astrocytes are Induced by Activated Microglia. Nature 2017, 541, 481–487. Available online: http://www.nature.com/doifinder/10.1038/nature21029 (accessed on 3 March 2020). [CrossRef] [Green Version]
- Masuda, T.; Sankowski, R.; Staszewski, O.; Prinz, M. Microglia Heterogeneity in the Single-Cell Era. Cell Rep. 2020, 30, 1271–1281. [Google Scholar] [CrossRef]
- Dimou, L.; Gallo, V. NG2-Glia and Their Functions in the Central Nervous System. Glia 2015, 63, 1429–1451. [Google Scholar] [CrossRef] [Green Version]
- Kassmann, C.M.; Lappe-Siefke, C.; Baes, M.; Brügger, B.; Mildner, A.; Werner, H.B.; Natt, O.; Michaelis, T.; Prinz, M.; Frahm, J.; et al. Axonal Loss and Neuroinflammation Caused by Peroxisome-Deficient Oligodendrocytes. Nat. Genet. 2007, 39, 969–976. [Google Scholar] [CrossRef] [Green Version]
- Diniz, L.P.; Matias, I.C.P.; Garcia, M.N.; Gomes, F.C.A. Astrocytic Control of Neural Circuit Formation: Highlights on TGF-Beta Signaling. Neurochem. Int. 2014, 78, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Pfrieger, F.W. Cholesterol Homeostasis and Function in Neurons of the Central Nervous System. Cell. Mol. Life Sci. 2003, 60, 1158–1171. [Google Scholar] [CrossRef] [PubMed]
- Henrich-Noack, P.; Prehn, J.H.; Krieglstein, J. Neuroprotective Effects of TGF-Beta 1. J. Neural Transm. Suppl. 1994, 43, 33–45. [Google Scholar] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goshi, N.; Kim, H.; Girardi, G.; Gardner, A.; Seker, E. Electrophysiological Activity of Primary Cortical Neuron-Glia Mixed Cultures. Cells 2023, 12, 821. https://doi.org/10.3390/cells12050821
Goshi N, Kim H, Girardi G, Gardner A, Seker E. Electrophysiological Activity of Primary Cortical Neuron-Glia Mixed Cultures. Cells. 2023; 12(5):821. https://doi.org/10.3390/cells12050821
Chicago/Turabian StyleGoshi, Noah, Hyehyun Kim, Gregory Girardi, Alexander Gardner, and Erkin Seker. 2023. "Electrophysiological Activity of Primary Cortical Neuron-Glia Mixed Cultures" Cells 12, no. 5: 821. https://doi.org/10.3390/cells12050821