
Preimplantation Genetic Testing for Aneuploidy (PGT-A) Reveals High Levels of Chromosomal Errors in In Vivo-Derived Pig Embryos, with an Increased Incidence When Produced In Vitro
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. In Vivo Embryo Collection
2.3. In Vitro Embryo Production
2.4. Embryo Storage, Whole Genome Amplification and Genotyping
2.5. PGT-A Analysis
2.6. Statistical Analysis
2.7. Data Availability
3. Results
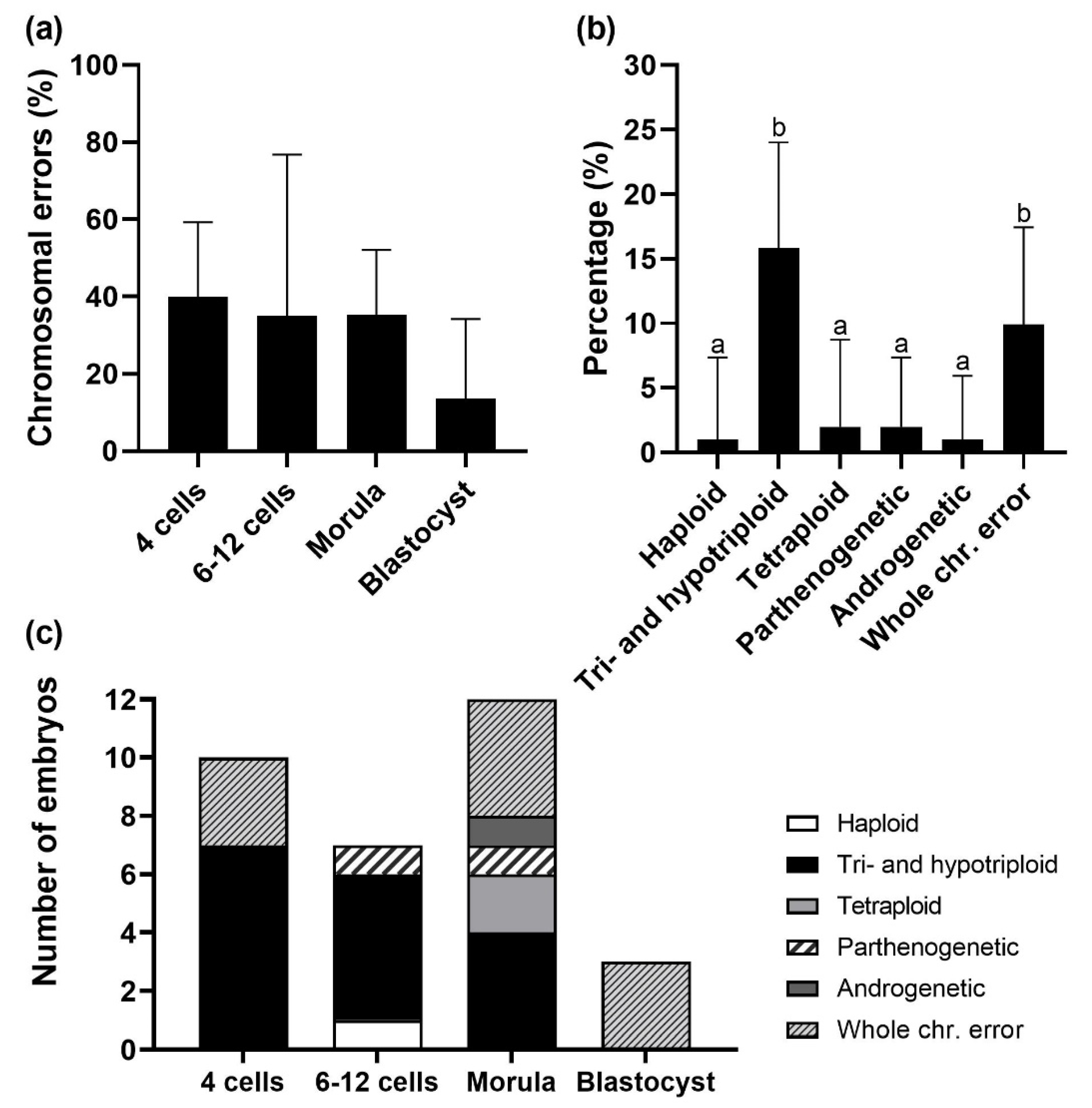
3.1. Chromosomal Errors and Aneuploidy Classes
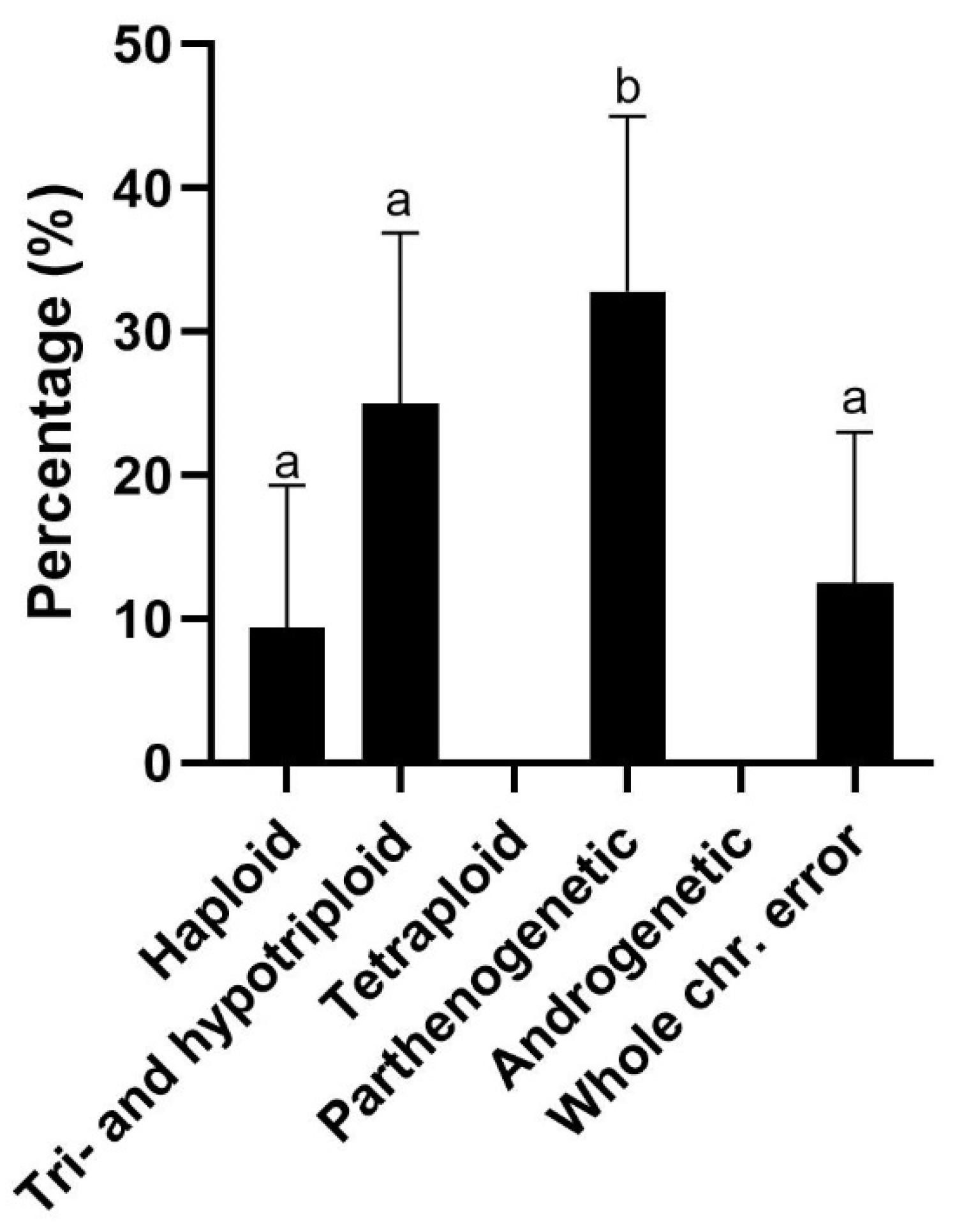
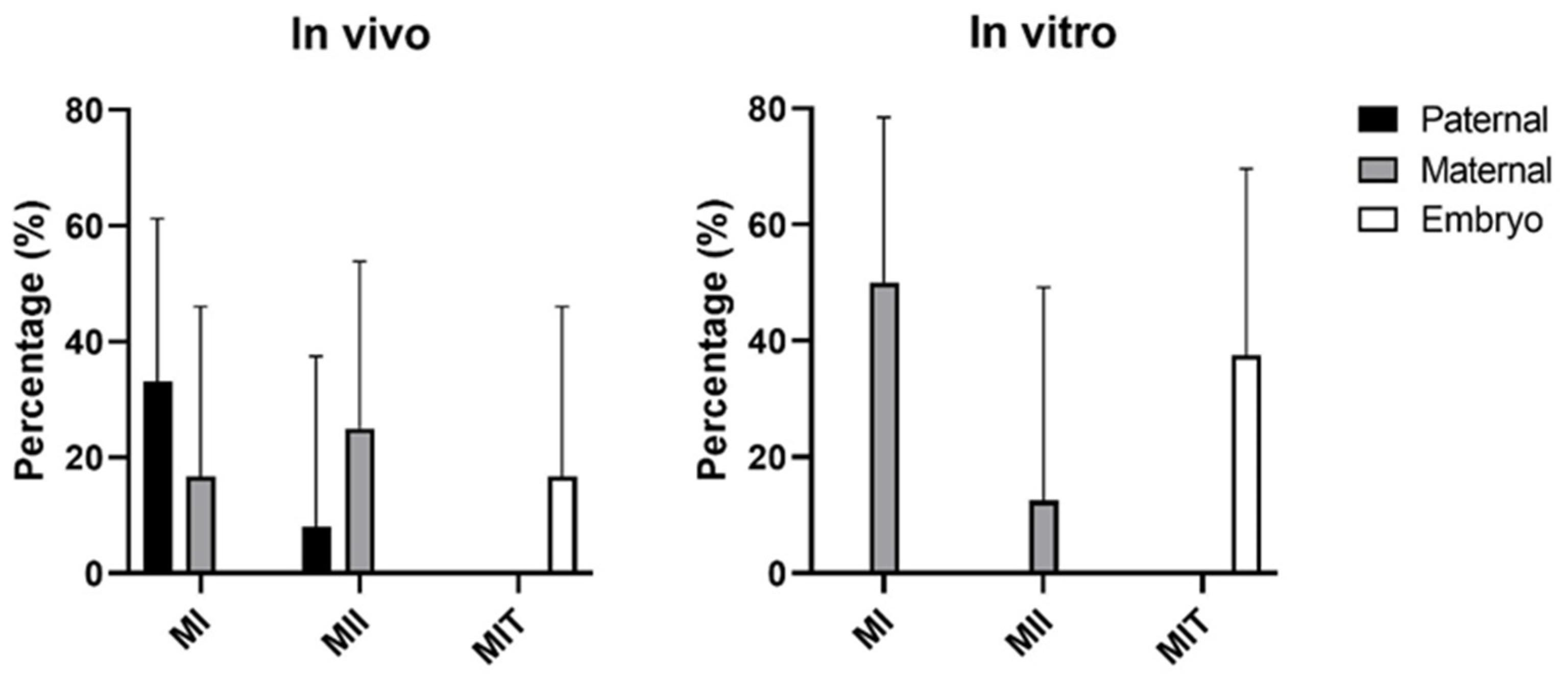
3.2. Origin of Chromosomal Errors
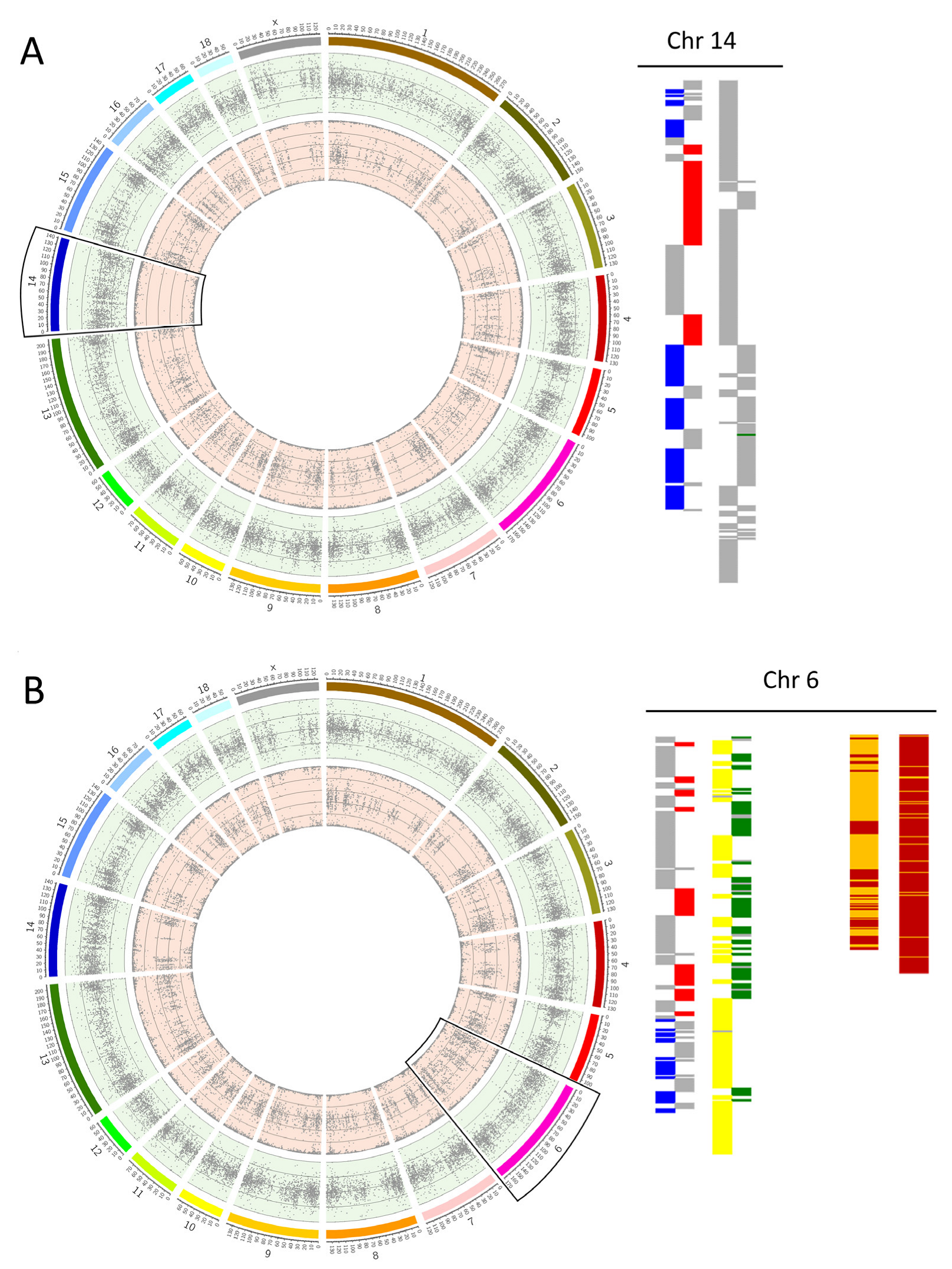
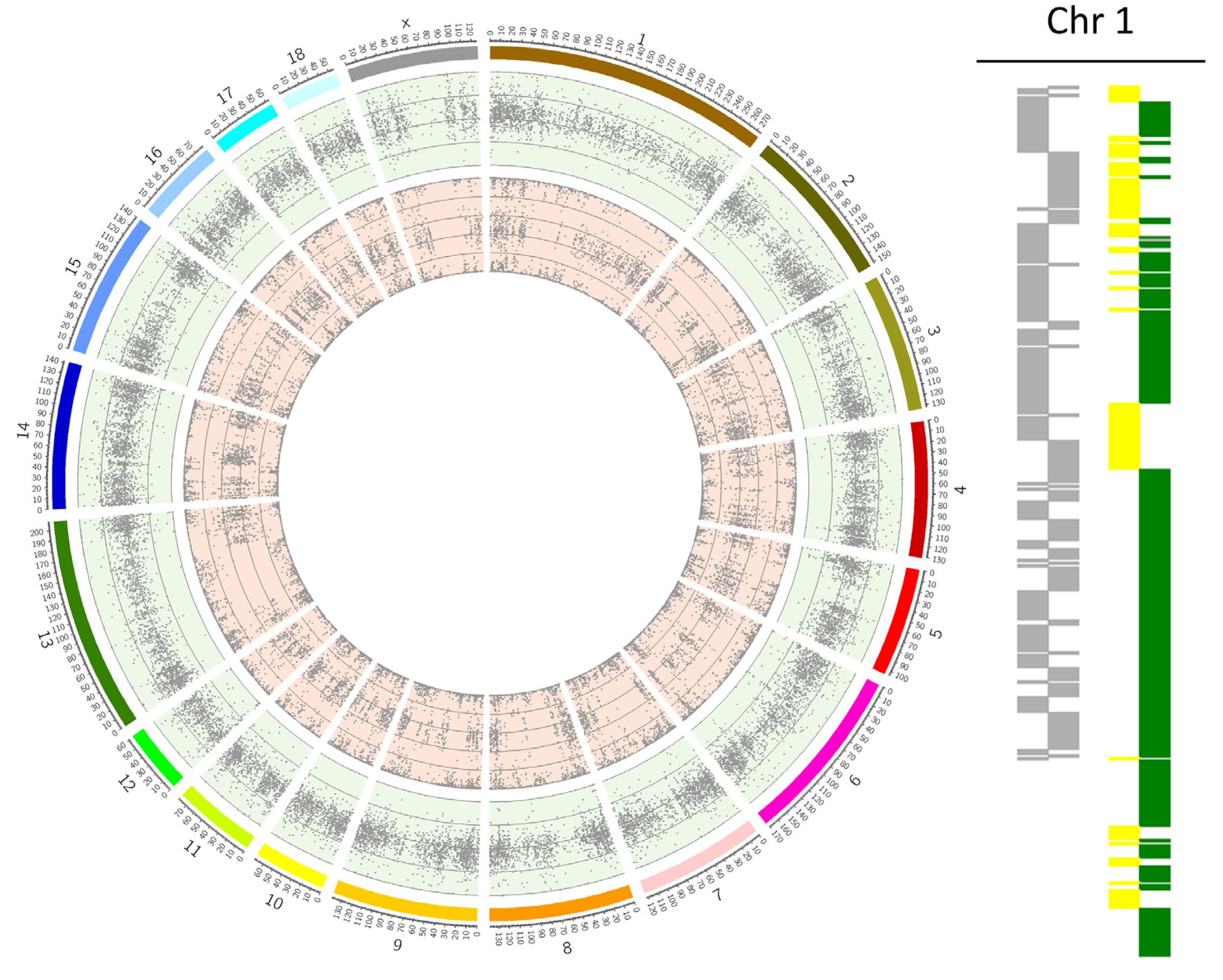
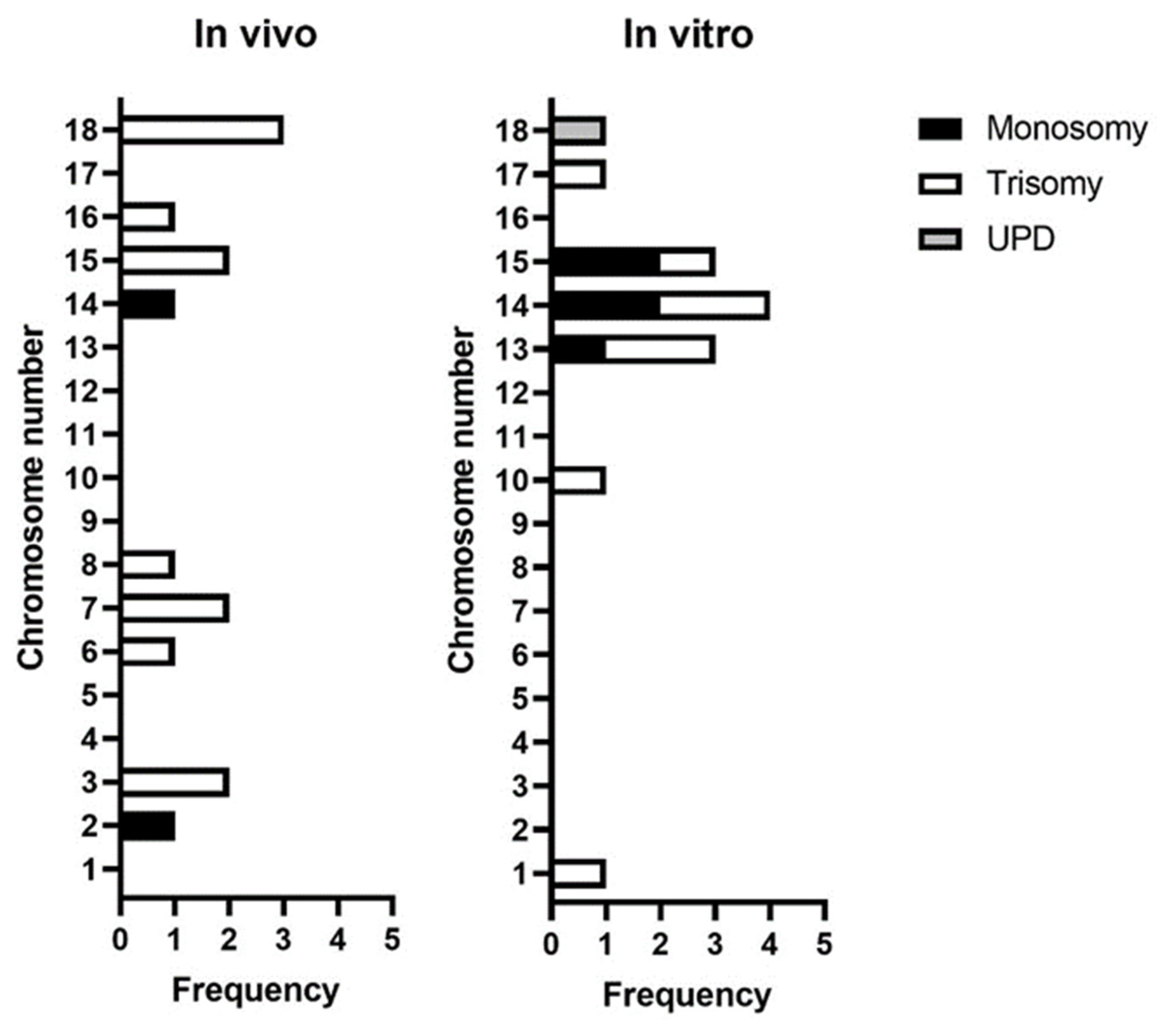
3.3. Incidence of Whole Chromosomal Errors by Chromosome
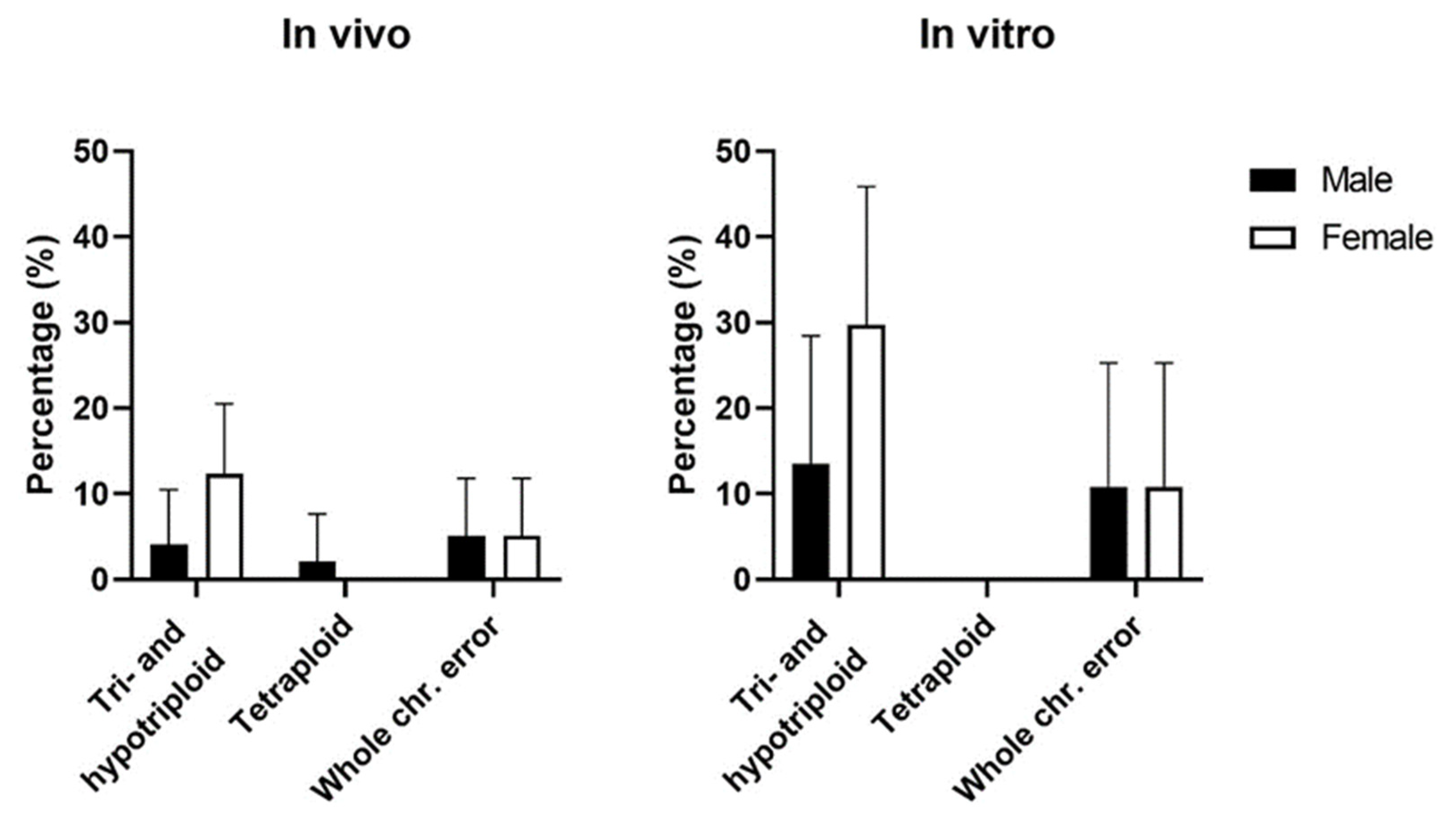
3.4. Chromosomal Errors by Sex
4. Discussion
4.1. In Vivo-Derived Embryos
4.2. IVP Blastocysts Show a High Prevalence of Parthenogenesis
4.3. A High Incidence of Triploidy in IVP Embryos
4.4. Origin of Chromosomal Errors
4.5. Acrocentric Chromosomes Are More Prone to Non-Disjunction
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Greco, E.; Litwicka, K.; Minasi, M.G.; Cursio, E.; Greco, P.F.; Barillari, P. Preimplantation Genetic Testing: Where We Are Today. Int. J. Mol. Sci. 2020, 21, 4381. [Google Scholar] [CrossRef]
- Viotti, M. Preimplantation Genetic Testing for Chromosomal Abnormalities: Aneuploidy, Mosaicism, and Structural Rearrangements. Genes 2020, 11, 602. [Google Scholar] [CrossRef]
- Mullaart, E.; Wells, D. Embryo Biopsies for Genomic Selection. In Animal Biotechnology 2; Niemann, H., Wrenzycki, C., Eds.; Springer: Cham, Switzerland, 2018; pp. 81–94. ISBN 978-3-319-92347-5. [Google Scholar]
- Mueller, M.L.; Van Eenennaam, A.L. Synergistic Power of Genomic Selection, Assisted Reproductive Technologies, and Gene Editing to Drive Genetic Improvement of Cattle. CABI Agric. Biosci. 2022, 3, 13. [Google Scholar] [CrossRef]
- Raudsepp, T.; Chowdhary, B.P. Chromosome Aberrations and Fertility Disorders in Domestic Animals. Annu. Rev. Anim. Biosci. 2016, 4, 15–43. [Google Scholar] [CrossRef] [PubMed]
- Harris, B.S.; Bishop, K.C.; Kuller, J.A.; Alkilany, S.; Price, T.M. Preimplantation Genetic Testing: A Review of Current Modalities. F S Rev. 2021, 2, 43–56. [Google Scholar] [CrossRef]
- Leader, B.; Lim, H.; Carabatsos, M.J.; Harrington, A.; Ecsedy, J.; Pellman, D.; Maas, R.; Leder, P. Formin-2, Polyploidy, Hypofertility and Positioning of the Meiotic Spindle in Mouse Oocytes. Nat. Cell Biol. 2002, 4, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Wasielak-Politowska, M.; Kordowitzki, P. Chromosome Segregation in the Oocyte: What Goes Wrong during Aging. Int. J. Mol. Sci. 2022, 23, 2880. [Google Scholar] [CrossRef]
- Silvestri, G.; Canedo-Ribeiro, C.; Serrano-Albal, M.; Labrecque, R.; Blondin, P.; Larmer, S.G.; Marras, G.; Tutt, D.A.R.; Handyside, A.H.; Farré, M.; et al. Preimplantation Genetic Testing for Aneuploidy Improves Live Birth Rates with In Vitro Produced Bovine Embryos: A Blind Retrospective Study. Cells 2021, 10, 2284. [Google Scholar] [CrossRef]
- Hornak, M.; Oracova, E.; Hulinska, P.; Urbankova, L.; Rubes, J. Aneuploidy Detection in Pigs Using Comparative Genomic Hybridization: From the Oocytes to Blastocysts. PLoS ONE 2012, 7, e30335. [Google Scholar] [CrossRef] [Green Version]
- Coppola, G.; Alexander, B.; Di Berardino, D.; St John, E.; Basrur, P.K.; King, W.A. Use of Cross-Species in-Situ Hybridization (ZOO-FISH) to Assess Chromosome Abnormalities in Day-6 in-Vivo- or in-Vitro-Produced Sheep Embryos. Chromosom. Res. 2007, 15, 399–408. [Google Scholar] [CrossRef] [Green Version]
- Hornak, M.; Jeseta, M.; Hanulakova, S.; Rubes, J. A High Incidence of Chromosome Abnormalities in Two-Cell Stage Porcine IVP Embryos. J. Appl. Genet. 2015, 56, 515–523. [Google Scholar] [CrossRef]
- Rambags, B.P.B.; Krijtenburg, P.J.; Van Drie, H.F.; Lazzari, G.; Galli, C.; Pearson, P.L.; Colenbrander, B.; Stout, T.A.E. Numerical Chromosomal Abnormalities in Equine Embryos Produced in Vivo and in Vitro. Mol. Reprod. Dev. 2005, 72, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Tšuiko, O.; Catteeuw, M.; Esteki, M.Z.; Destouni, A.; Pascottini, O.B.; Besenfelder, U.; Havlicek, V.; Smits, K.; Kurg, A.; Salumets, A.; et al. Genome Stability of Bovine in Vivo-Conceived Cleavage-Stage Embryos Is Higher Compared to in Vitro-Produced Embryos. Hum. Reprod. 2017, 32, 2348–2357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coticchio, G.; Barrie, A.; Lagalla, C.; Borini, A.; Fishel, S.; Griffin, D.; Campbell, A. Plasticity of the Human Preimplantation Embryo: Developmental Dogmas, Variations on Themes and Self-Correction. Hum. Reprod. Update 2021, 27, 848–865. [Google Scholar] [CrossRef] [PubMed]
- Jochems, R.; Gaustad, A.H.; Styrishave, B.; Zak, L.J.; Oskam, I.C.; Grindflek, E.; Myromslien, F.D.; Kommisrud, E.; Krogenæs, A.K. Follicular Fluid Steroid Hormones and in Vitro Embryo Development in Duroc and Landrace Pigs. Theriogenology 2022, 190, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, K.; Suzuki, C.; Onishi, A. Defined System for In Vitro Production of Porcine Embryos Using a Single Basic Medium. J. Reprod. Dev. 2008, 54, 208–213. [Google Scholar] [CrossRef] [Green Version]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An Information Aesthetic for Comparative Genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [Green Version]
- Attiyeh, E.F.; Diskin, S.J.; Attiyeh, M.A.; Mossé, Y.P.; Hou, C.; Jackson, E.M.; Kim, C.; Glessner, J.; Hakonarson, H.; Biegel, J.A.; et al. Genomic Copy Number Determination in Cancer Cells from Single Nucleotide Polymorphism Microarrays Based on Quantitative Genotyping Corrected for Aneuploidy. Genome Res. 2009, 19, 276–283. [Google Scholar] [CrossRef] [Green Version]
- Handyside, A.H.; Harton, G.L.; Mariani, B.; Thornhill, A.R.; Affara, N.; Shaw, M.A.; Griffin, D.K. Karyomapping: A Universal Method for Genome Wide Analysis of Genetic Disease Based on Mapping Crossovers between Parental Haplotypes. J. Med. Genet. 2010, 47, 651–658. [Google Scholar] [CrossRef] [Green Version]
- Gabriel, A.S.; Hassold, T.J.; Thornhill, A.R.; Affara, N.A.; Handyside, A.H.; Griffin, D.K. An Algorithm for Determining the Origin of Trisomy and the Positions of Chiasmata from SNP Genotype Data. Chromosom. Res. 2011, 19, 155–163. [Google Scholar] [CrossRef]
- Martinez, E.A.; Cuello, C.; Parrilla, I.; Rodriguez-Martinez, H.; Roca, J.; Vazquez, J.L.; Vazquez, J.M.; Gil, M.A. Design, Development, and Application of a Non-Surgical Deep Uterine Embryo Transfer Technique in Pigs. Anim. Front. 2013, 3, 40–47. [Google Scholar] [CrossRef] [Green Version]
- Martinez, E.A.; Martinez, C.A.; Nohalez, A.; Sanchez-Osorio, J.; Vazquez, J.M.; Roca, J.; Parrilla, I.; Gil, M.A.; Cuello, C. Nonsurgical Deep Uterine Transfer of Vitrified, in Vivo-Derived, Porcine Embryos Is as Effective as the Default Surgical Approach. Sci. Rep. 2015, 5, 10587. [Google Scholar] [CrossRef] [Green Version]
- Hornak, M.; Hulinska, P.; Musilova, P.; Kubickova, S.; Rubes, J. Investigation of Chromosome Aneuploidies in Early Porcine Embryos Using Comparative Genomic Hybridization. Cytogenet. Genome Res. 2009, 126, 210–216. [Google Scholar] [CrossRef]
- McCauley, T.C.; Mazza, M.R.; Didion, B.A.; Mao, J.; Wu, G.; Coppola, G.; Coppola, G.F.; Di Berardino, D.; Day, B.N. Chromosomal Abnormalities in Day-6, in Vitro-Produced Pig Embryos. Theriogenology 2003, 60, 1569–1580. [Google Scholar] [CrossRef]
- Tutt, D.A.R.; Silvestri, G.; Serrano-Albal, M.; Simmons, R.J.; Kwong, W.Y.; Guven-Ates, G.; Canedo-Ribeiro, C.; Labrecque, R.; Blondin, P.; Handyside, A.H.; et al. Analysis of Bovine Blastocysts Indicates Ovarian Stimulation Does Not Induce Chromosome Errors, nor Discordance between Inner-Cell Mass and Trophectoderm Lineages. Theriogenology 2021, 161, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Pauerova, T.; Radonova, L.; Kovacovicova, K.; Novakova, L.; Skultety, M.; Anger, M. Aneuploidy during the Onset of Mouse Embryo Development. Reproduction 2020, 160, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Griffin, D.K.; Brezina, P.R.; Tobler, K.; Zhao, Y.; Silvestri, G.; Mccoy, R.C.; Anchan, R.; Benner, A.; Kearns, W.G. The Human Embryonic Genome Is Karyotypically Complex, with Chromosomally Abnormal Cells Preferentially Located Away from the Developing Fetus. Hum. Reprod. 2023, 38, 180–188. [Google Scholar] [CrossRef]
- Zudova, D.; Rezacova, O.; Kubickova, S.; Rubes, J. Aneuploidy Detection in Porcine Embryos Using Fluorescence in Situ Hybridization. Cytogenet. Genome Res. 2003, 102, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Diskin, M.G.; Murphy, J.J.; Sreenan, J.M. Embryo Survival in Dairy Cows Managed under Pastoral Conditions. Anim. Reprod. Sci. 2006, 96, 297–311. [Google Scholar] [CrossRef]
- Hassold, T.; Hall, H.; Hunt, P. The Origin of Human Aneuploidy: Where We Have Been, Where We Are Going. Hum. Mol. Genet. 2007, 16, 203–208. [Google Scholar] [CrossRef]
- Turner, K.J.; Silvestri, G.; Black, D.H.; Dobson, G.; Smith, C.; Handyside, A.H.; Sinclair, K.D.; Griffin, D.K. Karyomapping for Simultaneous Genomic Evaluation and Aneuploidy Screening of Preimplantation Bovine Embryos: The First Live-Born Calves. Theriogenology 2019, 125, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.S.; Park, M.R.; Lee, H.S.; Kwak, T.U.; Son, H.Y.; Kang, J.K.; Lee, J.W.; Lee, K.; Park, E.W.; Hwang, S. Developmental and Degenerative Characterization of Porcine Parthenogenetic Fetuses during Early Pregnancy. Animals 2020, 10, 622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagutina, I.; Lazzari, G.; Duchi, R.; Galli, C. Developmental Potential of Bovine Androgenetic and Parthenogenetic Embryos: A Comparative Study. Biol. Reprod. 2004, 70, 400–405. [Google Scholar] [CrossRef]
- Kawarasaki, T.; Otake, M.; Tsuchiya, S.; Shibata, M.; Matsumoto, K.; Isobe, N. Co-Transfer of Parthenogenotes and Single Porcine Embryos Leads to Full-Term Development of the Embryos. Anim. Reprod. Sci. 2009, 112, 8–21. [Google Scholar] [CrossRef]
- Cuello, C.; Gil, M.A.; Almiñana, C.; Sanchez-Osorio, J.; Parrilla, I.; Caballero, I.; Vazquez, J.M.; Roca, J.; Rodriguez-Martinez, H.; Martinez, E.A. Vitrification of in Vitro Cultured Porcine Two-to-Four Cell Embryos. Theriogenology 2007, 68, 258–264. [Google Scholar] [CrossRef]
- Xu, X.; Seth, P.C.; Harbison, D.S.; Cheung, A.P.; Foxcroft, G.R. Semen Dilution for Assessment of Boar Ejaculate Quality in Pig IVM and IVF Systems. Theriogenology 1996, 46, 1325–1337. [Google Scholar] [CrossRef]
- Suzuki, H.; Saito, Y.; Kagawa, N.; Yang, X. In Vitro Fertilization and Polyspermy in the Pig: Factors Affecting Fertilization Rates and Cytoskeletal Reorganization of the Oocyte. Microsc. Res. Tech. 2003, 61, 327–334. [Google Scholar] [CrossRef] [Green Version]
- Jochems, R.; Gaustad, A.H.; Zak, L.J.; Grindflek, E.; Zeremichael, T.T.; Oskam, I.C.; Myromslien, F.D.; Kommisrud, E.; Krogenæs, A.K. Effect of Two ‘Progressively Motile Sperm—Oocyte’ Ratios on Porcine in Vitro Fertilization and Embryo Development. Zygote 2022, 30, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Sosnowski, J.; Waroczyk, M.; Switonski, M. Chromosome Abnormalities in Secondary Pig Oocytes Matured in Vitro. Theriogenology 2003, 60, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Ulloa Ulloa, C.M.; Yoshizawa, M.; Komoriya, E.; Mitsui, A.; Nagai, T.; Kikuchi, K. The Blastocyst Production Rate and Incidence of Chromosomal Abnormalities by Developmental Stage in in Vitro Produced Porcine Embryos. J. Reprod. Dev. 2008, 54, 22–29. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.T.; Quang, T.; Somfai, D.T.; Thi, N.; Nguyen, M.; Linh, V.; Xuan, B.; Junko, N. Selection Based on Morphological Features of Porcine Embryos Produced by in Vitro Fertilization: Timing of Early Cleavages and the Effect of Polyspermy. Anim. Sci. J. 2020, 91, e13401. [Google Scholar] [CrossRef] [PubMed]
- Funahashi, H. Polyspermic Penetration in Porcine IVM–IVF Systems. Reprod. Fertil. Dev. 2003, 15, 167. [Google Scholar] [CrossRef] [PubMed]
- Franasiak, J.M.; Forman, E.J.; Hong, K.H.; Werner, M.D.; Upham, K.M.; Treff, N.R.; Scott, R.T. Aneuploidy across Individual Chromosomes at the Embryonic Level in Trophectoderm Biopsies: Changes with Patient Age and Chromosome Structure. J. Assist. Reprod. Genet. 2014, 31, 1501–1509. [Google Scholar] [CrossRef] [Green Version]
- Gruhn, J.R.; Zielinska, A.P.; Shukla, V.; Blanshard, R.; Cimadomo, D.; Nikiforov, D.; Chan, A.C.; Louise, J.; Vogel, I.; Scarica, C.; et al. Chromosome Errors in Human Eggs Shape Natural Fertility over Reproductive Lifespan. Science 2020, 365, 1466–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ljunger, E.; Cnattingius, S.; Lundin, C.; Annerén, G. Chromosomal Anomalies in First-Trimester Miscarriages. Acta Obstet. Gynecol. Scand. 2005, 84, 1103–1107. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sow | Animal | Collection | Embryos | Embryo Stages | CLs | CR |
|---|---|---|---|---|---|---|
| 1 | Primiparous | Day 4 | 24 | 4–8 cells (17 × 4 cell) | 26 | 92% |
| 2 | Primiparous | Day 4 | 15 | 3–6 cells (equal distr.) | 22 | 68% |
| 3 | Nulliparous | Day 5 | 38 | Compacted morulae | 50 | 76% |
| 4 | Nulliparous | Day 5 | 20 | Compacted morulae | 25 | 80% |
| 5 | Primiparous | Day 6 | 26 | Expanded blastocysts | - | - |
| Aneuploidy Classes | S1 | S2 | S3 | S4 | S5 | S6 | S7 | S8 | S9 | S10 | Tot |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Euploid | 2 | 2 | 1 | 2 | 3 | 3 | 13 | ||||
| Haploid | 2 | 1 | 2 | 1 | 6 | ||||||
| Tri- and hypotriploid | 3 | 3 | 3 | 3 | 1 | 1 | 2 | 16 | |||
| Parthenogenetic | 4 | 13 | 4 | 21 | |||||||
| Whole chr. error | 1 | 3 | 1 | 1 | 1 | 1 | 8 | ||||
| Total | 4 | 10 | 7 | 2 | 4 | 9 | 3 | 5 | 13 | 7 | 64 |
| Aneuploidy Classes | Overall | Paternal | Maternal | Embryo | ||||
|---|---|---|---|---|---|---|---|---|
| IVD | IVP | IVD | IVP | IVD | IVP | IVD | IVP | |
| Haploid | 1 | 6 | 1 | 6 | ||||
| Tri- and hypotriploid | 16 | 16 | 12 | 12 | 4 | 4 | ||
| Tetraploidy | 2 | * | * | |||||
| Parthenogenetic | 2 | 21 | 2 | 21 | ||||
| Androgenetic | 1 | 1 | ||||||
| Whole chr. error | 14 | 14 | 5 | 1 | 7 | 10 | 2 | 2 |
| Monosomy | 2 | 5 | 2 | 5 | ||||
| Trisomy | 12 | 8 | 5 | 5 | 5 | 2 | 3 | |
| Uniparental disomy | 1 | 1 | ||||||
| Total | 36 | 57 | 19 | 13 | 14 | 41 | 2 | 3 |
| Sex | n (%) | % Chromosomal Errors | |
|---|---|---|---|
| In vivo | Male | 41 (42.3%) | 26.8% (11/41) |
| Female | 56 (57.7%) | 30.4% (17/56) | |
| In vitro | Male | 18 (48.6%) | 50.0% (9/18) |
| Female | 19 (51.4%) | 78.9% (15/19) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jochems, R.; Canedo-Ribeiro, C.; Silvestri, G.; Derks, M.F.L.; Hamland, H.; Zak, L.J.; Knol, E.F.; Handyside, A.H.; Grindflek, E.; Griffin, D.K. Preimplantation Genetic Testing for Aneuploidy (PGT-A) Reveals High Levels of Chromosomal Errors in In Vivo-Derived Pig Embryos, with an Increased Incidence When Produced In Vitro. Cells 2023, 12, 790. https://doi.org/10.3390/cells12050790
Jochems R, Canedo-Ribeiro C, Silvestri G, Derks MFL, Hamland H, Zak LJ, Knol EF, Handyside AH, Grindflek E, Griffin DK. Preimplantation Genetic Testing for Aneuploidy (PGT-A) Reveals High Levels of Chromosomal Errors in In Vivo-Derived Pig Embryos, with an Increased Incidence When Produced In Vitro. Cells. 2023; 12(5):790. https://doi.org/10.3390/cells12050790
Chicago/Turabian StyleJochems, Reina, Carla Canedo-Ribeiro, Giuseppe Silvestri, Martijn F. L. Derks, Hanne Hamland, Louisa J. Zak, Egbert F. Knol, Alan H. Handyside, Eli Grindflek, and Darren K. Griffin. 2023. "Preimplantation Genetic Testing for Aneuploidy (PGT-A) Reveals High Levels of Chromosomal Errors in In Vivo-Derived Pig Embryos, with an Increased Incidence When Produced In Vitro" Cells 12, no. 5: 790. https://doi.org/10.3390/cells12050790