
Advances of Wnt Signalling Pathway in Colorectal Cancer
1
Marine College, Shandong University, Weihai 264200, China
2
Shandong Kelun Pharmaceutical Co., Ltd., Binzhou 256600, China
*
Author to whom correspondence should be addressed.
Cells 2023, 12(3), 447; https://doi.org/10.3390/cells12030447
Submission received: 18 December 2022
/
Revised: 25 January 2023
/
Accepted: 28 January 2023
/
Published: 30 January 2023
(This article belongs to the Special Issue From Mechanisms to Therapeutics: Wnt Signaling in Cancer)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Colorectal cancer (CRC) represents one of the most common cancers worldwide, with a high mortality rate despite the decreasing incidence and new diagnostic and therapeutic strategies. CRC arises from both epidemiologic and molecular backgrounds. In addition to hereditary factor and genetic mutations, the strongly varying incidence of CRC is closely linked to chronic inflammatory disorders of the intestine and terrible dietary habits. The Wnt signalling pathway is a complex regulatory network that is implicated in many CRC physiological processes, including cancer occurrence, development, prognosis, invasion, and metastasis. It is currently believed to include classical Wnt/β-catenin, Wnt/PCP, and Wnt/Ca2+. In this review, we summarise the recent mechanisms and potential regulators of the three branches of the Wnt signalling pathway in CRC.
1. Introduction
Wnt signalling is one of the key cascades regulating normal development and physiology and has also been tightly evolved to perform diverse functions in cancer [1,2]. Wnt signal transduction is divided into the Wnt/β-catenin or canonical Wnt signalling pathway, the planar cell polarity (Wnt/PCP) pathway, and the Wnt/Ca2+ signalling pathway [3]. Abnormal Wnt pathway components have been identified as drivers of cancer and as potential targets for cancer treatment, especially in colorectal cancer (CRC) [4]. CRC is the third most common cancer worldwide [5,6]. In recent years, the majority of CRCs have been sporadic and early-onset, which is largely attributed to a constellation of modifiable environmental risk factors (for example, obesity, physical inactivity, poor diet, alcohol consumption, and smoking) [7,8,9]. It is now widely accepted that the Wnt signalling pathway plays a pivotal role in tumour development, progression, invasion, and metastasis, and in drug resistance and the relapse of cancer. In this review, we determine the impact of the canonical and non-canonical Wnt signalling pathways in driving the occurrence, growth, invasion, and chemoresistance of CRC tumours, as well as their role in prognosis, to comprehensively understand the role enacted by the Wnt pathway in CRC [10].
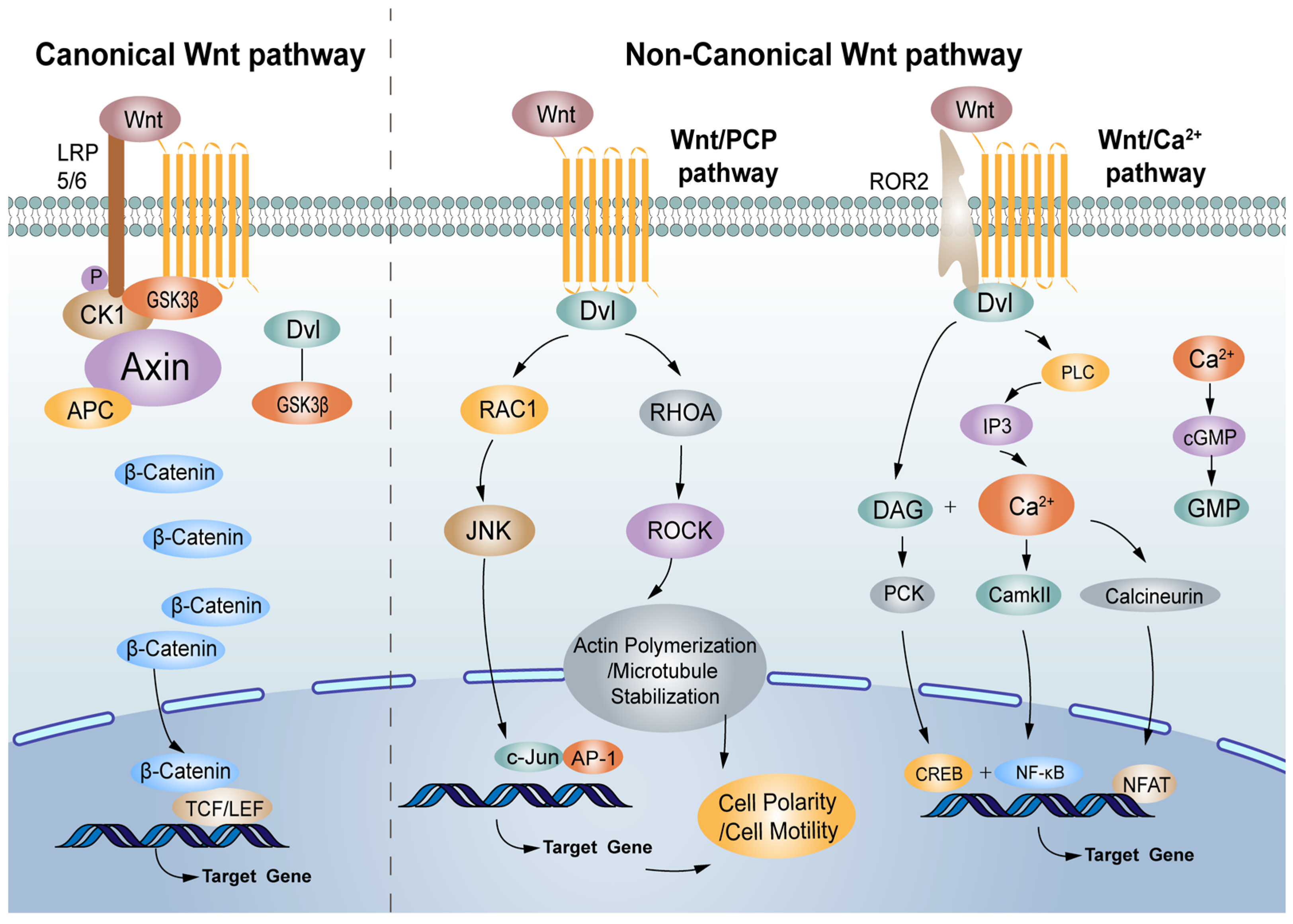
2. The Canonical Wnt/β-Catenin Pathway
The Wnt/β-catenin signalling pathway is associated with physiological processes and contributes to the diverse processes of some solid tumours and haematological malignancies, such as CRC [11,12]. Under normal physiological conditions, the transcription factor β-catenin, the crucial component of the Wnt signalling pathway, is destroyed by the β-catenin destruction complex formed by adenomatous polyposis (APC), casein kinase I (CK I), glycogen synthase kinase 3β (GSK3β) and Axin in which Axin is a scaffolding protein, and CK I and GSK3β phosphorylate β-catenin in turn [13,14,15]. Subsequently, the phosphorylated β-catenin is recognised and degraded by E3 ubiquitin ligase (β-TrCP) [16]. Low-density lipoprotein receptor-related protein 5/6 (LRP5/6), co-receptors for Wnt ligands, are associated with Frizzled (FZD) receptors. Once bound by Wnt, FZD/LRP complex activates the canonical signalling pathway. The Dishevelled (Dvl), a cytoplasmic protein that functions as upstream of β-catenin and GSK3β, is activated. Dvl is required for the phosphorylation of LRP5/6 PPPSPxS motifs. LRP5/6 are phosphorylated by several protein kinases including GSK3β and CK I, and recruits the scaffold protein Axin. Multimers of receptor-bound Dvl and Axin molecules support the formation of the LRP-FZD dimer. The loss of function of the destruction complex leads to the stabilization of β-catenin. As a result, β-catenin accumulates in the cytoplasm, undergoing nuclear translocation, and activates the transcription factor T cell factor (TCF)/lymph enhancer factor (LEF) family to initiate the expression of Wnt target genes [17,18,19,20].
3. The Non-Canonical Wnt Signalling Pathway
In addition to the Wnt/β-catenin signalling pathway, many researchers have indicated that the non-canonical Wnt signalling pathway also has a vital role, with growing links to cancer progression, exerting both oncogenic and tumour-suppressive effects [21]. The main non-canonical Wnt signalling pathways are the Wnt/PCP and Wnt/Ca2+ signalling pathways [22]. Wnt1, Wnt2, Wnt3, Wnt3a, Wnt8a, Wnt8b, Wnt10a, and Wnt10b are regarded as canonical Wnt proteins, while Wnt4, Wnt5a, Wnt5b, Wnt7a, Wnt7b, and Wnt11 belong to the non-canonical Wnt proteins, which initiate canonical and non-canonical pathways, respectively, by activating Frizzled receptors (Figure 1).
3.1. The Wnt/PCP Signalling Pathway
The polarization of many epithelia within the plane of the epithelium is referred to as planar cell polarity (PCP), and the associated molecular pathway can regulate cellular polarity in non-epithelial/epithelial contexts [23]. PCP is an evolutionarily conserved pathway that is regulated by Frizzled (FZD)/PCP core group, which includes the seven-pass transmembrane receptor Frizzled (FZD), the atypical cadherin Flamingo (Fmi/Celsr), the tetraspanin-like transmembrane protein Van Gogh (Vang or Stbm/Vangl), and the cytosolic components Dishevelled (Dsh/Dvl), Diego (Dgo/Ankrd6), and Prickle (Pk) [24]. Wnt5a-Frizzled binding needs to transduce signals with a co-receptor Ror2 and induces the formation of a Vangl2-Ror2 complex [25]. In addition, phosphorylated Vangl contributes to Vangl2-Ror2 localisation and/or function, which requires the activity of CK I and Dvl [26]. Downstream signalling cascades (RHO-ROCK (Rho-associated kinase) and RAC-JNK (c-Jun-N-terminal kinase)) in the Wnt/PCP pathway are activated [27]. Then, RHO, RAC, JNK, and ROCK activate a downstream target gene, which is involved in neuronal plasticity, regeneration, cell death, and the regulation of cellular senescence [28].
3.2. The Wnt/Ca2+ Signalling Pathway
More and more evidence indicates that the Wnt/Ca2+ signalling pathway influences cell fate decisions and cell migration [29]. Wnt-Fzd receptor ligand interaction along with the phosphorylation of co-receptor Ror by GSK3β activates phospholipase C, which derives inositol 1,4,5-triphosphate (IP3) and 1,2 diacylglycerol (DAG) from phospholipid phosphatidyl inositol 4,5-bisphosphate [30]. IP3 regulates the endoplasmic reticulum (ER) by releasing calcium ions, resulting in the activation of calcium calmodulin-dependent protein kinase II (CaMKII) and phosphatase calcinurin (Cn). Together, DAG and the calcium ions released from the ER activate protein kinase C (PKC) [31]. Both CaMKII and PKC activate various regulatory proteins (NF-κB and CREB), and Cn can activate the cytoplasmic protein nuclear factor associated with T cells (NFAT) [32]. NFAT, NF-κB, and CREB translocate to the nucleus and transcribe downstream regulatory genes [33]. The Wnt/FZD receptor ligand interaction may also activate phosphodiesterase 6 (PDE6) in a calcium-dependent manner, leading to a decrease in cyclic guanosine monophosphate (cGMP).
4. The Wnt Signalling Pathway in Occurrence of Colorectal Cancer (CRC)
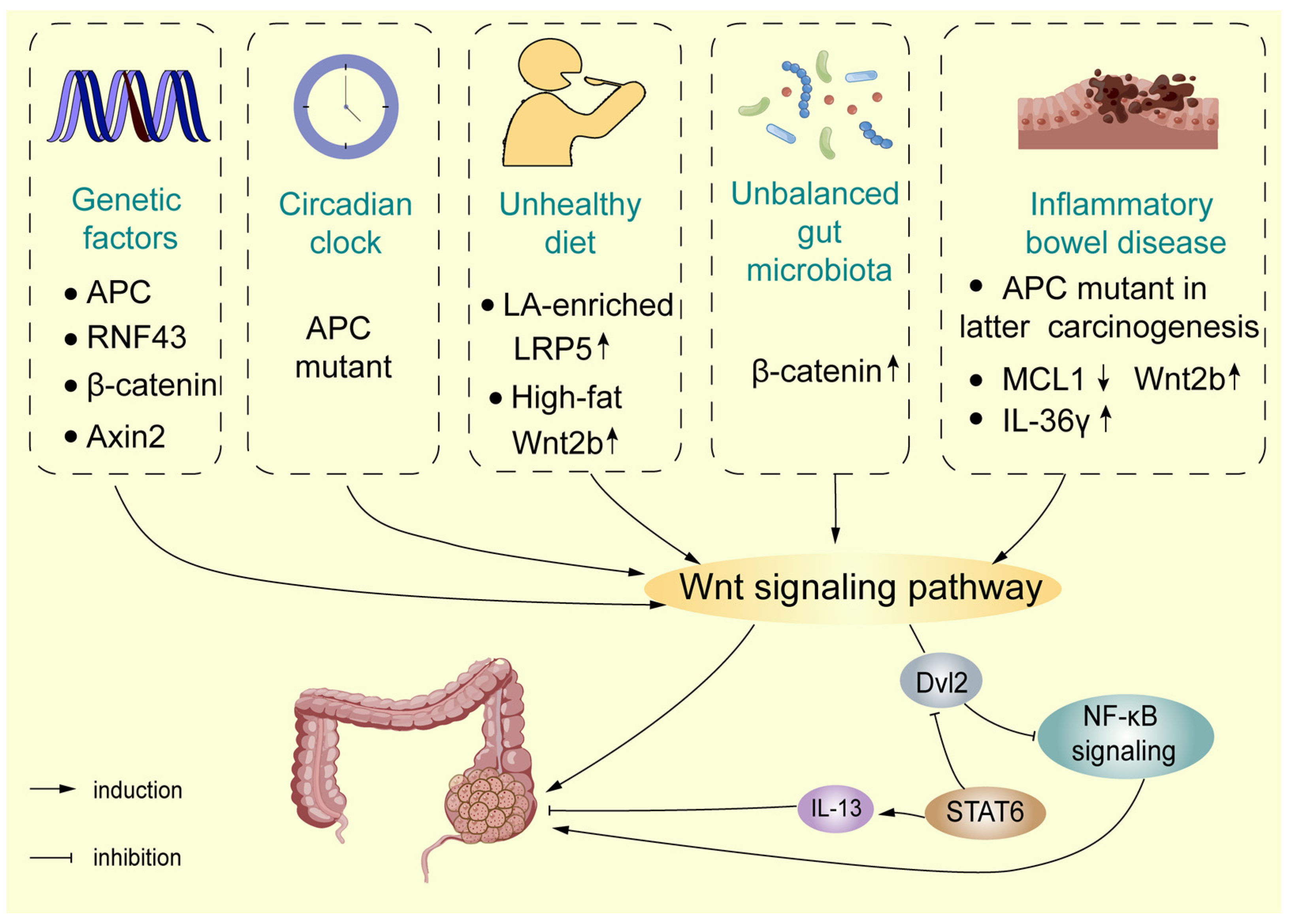
The Wnt signalling pathway is one of the most representative signalling pathways in CRC, playing an important role in the occurrence of CRC (Figure 2). According TCGA data, in colorectal cancer (CRC; n = 594), using Memorial Sloan Kettering Cancer Center (MSKCC), the largest fraction of sporadic tumours accumulates APC mutations (67%), followed by lower fractions of ring finger protein 43 (RNF43) (8%), β-catenin (6%), and Axin2 (5%) mutations according to frequency of mutation or deep deletion of the Wnt pathway genes [34]. In addition to the Wnt/β-catenin signalling pathway, many researchers have indicated that the non-canonical Wnt signalling pathway also has a vital role, with growing links to cancer progression, exerting both oncogenic and tumour-suppressive effects [21]. The main non-canonical Wnt signalling pathways are the Wnt/PCP and Wnt/Ca2+ signalling pathways [22].
4.1. The Wnt Signalling Pathway and Environmental Exposure
CRC is divided into sporadic and familial/hereditary cases, emphasising an aetiology with shared environmental factors and genes. Driver mutations in APC are representative of CRC subsets with distinct characteristics. Inactivating APC mutations causes familial adenomatous polyposis (FAP), which is a syndrome characterized by extensive intestinal polyp formation and a high risk of CRC development [35]. Moreover, a large number of APC mutations have also been found in sporadic CRCs [36]. In addition to individual and family genetic factors, the incidence of sporadic CRC (sCRC) and early colon cancer is increasingly common due to unhealthy lifestyles and environmental exposure [37].
4.1.1. Lifestyle
The causes of CRC tumorigenesis and progression are complicated and may include complex interactions. Apart from heredity factors, an unhealthy lifestyle is regarded as a potential risk factor associated with CRC. Diet influences a large part of the maintenance and function of the colon environment in mammals, and biological changes may be associated with the incidence of CRC [38,39]. A recent study indicated that linoleic acid (LA)-enriched diets promote chemically induced CRC via the Wnt/β-catenin signalling pathway. The 15-LOX-1 peroxidation of LA in phosphatidylinositol-3-phosphates (PI3P_LA) leads to the formation of PI3P_13-HODE, which decreases PI3P binding to SNX17 and LRP5 and inhibits the recycling of LRP5 from endosomes to the plasma membrane, thereby increasing LRP5 lysosomal degradation [40]. Moreover, the effect of a high-fat diet on the intestinal stem cell (ISC) niche is related to CRC risk: a high-fat diet resulted in the dysregulation of colonic mesenchymal stromal cells (MSCs), including increased cell numbers, increased bile acid (BA) receptor expression on colonic MSCs, the overexpression of Wnt2b, and the activation of CAF-like properties, which promoted crypt regeneration. In this way, BAs stimulate colonic MSC-induced Wnt2b production through the activation of FXR signalling [41].
Furthermore, a recent study found that the circadian clock was seemingly regarded as a risk factor impinging on intestinal physiology [42]. The molecular clock consists of the core transcription factors, circadian locomotor output cycles kaput (CLOCK), and aryl hydrocarbon receptor nuclear translocator-like protein 1 (Arntl encoding BMAL1), which regulate the clock gene expression programmes [43]. In human colorectal tumours, clock gene expression disorders cause abnormal cell cycle progression and regulate Wnt secretion in the intestinal epithelium [44]. Recently, researchers found the molecular pathways linking circadian disruption and the pathogenesis of CRC by developing a new tissue-specific genetically engineered mouse model (GEMM). The results demonstrated that the environmental disruption of the circadian clock accelerated the tumour burden in the APC mutant GEMM, as well as patient-derived organoids (PDOs), which highlights the importance of the circadian clock in the intestine. In addition, data demonstrated that disruption of intestinal Bmal1 accelerated the heterozygosity (LOH) of APC, and Wnt/β-catenin signalling was aberrantly activated, enhancing MYC-dependent glycolytic metabolism to accelerate CRC progression [45].
4.1.2. Pro-Tumorigenic Bacteria
The involvement of gut microbiota in CRC carcinogenesis has been extensively studied. Current evidence suggests that bacteria contribute to the development of CRC, partly by stimulating chronic inflammation [46,47]. Colonisation with anti-virulence factor A (AvrA) expressing Salmonella enterica significantly reduced several proteins of the Wnt family, including Wnt1, Wnt2, Wnt3, Wnt6, Wnt9a, and Wnt11 [48,49]. Furthermore, exposure to a short-term colibactin-producing (pks+) Escherichia coli infected healthy murine colon epithelial cells, which led to mutations in the Wnt signalling pathway in a study utilising organoids [50]. In terms of Fusobacterium nucleatum in CRC carcinogenesis via the Wnt signalling pathway, it has been reported that the F. nucleatum-associated CRC process was modulated by the phospholipid-binding protein Annexin A1, which was upregulated in cancerous cells by binding FadA, a F. nucleatum adhesion molecule, to E-cadherin, resulting in the activation of β-catenin-regulated transcription and an increased expression of transcription factors and the oncogenes c-Myc and Cyclin D1 [51,52]. Some researchers also believe that F. nucleatum activates the Wnt/β-catenin pathway by the upregulation of cyclin-dependent kinase 5 (Cdk5) to promote the proliferation and migration of cells [53]. Another study reported that F. nucleatum stably adheres to cancer stem cells (CSCs), likely by multiple interactions involving the tumour-associated Gal-GalNac disaccharide and the Fn-docking protein CEA-family cell adhesion molecule 1 (CEACAM-1), contributing to microbiota-driven colorectal carcinogenesis [54].
4.1.3. Gut Microbiota
The alteration of gut microbiota in all mucosal surfaces of the colon causes dysbiosis of the gut environment, which is involved in CRC tumorigenesis [55]. Intestinal microbiota activate the pathogen recognition receptors PRR9 to induce innate immune responses and activate the inflammatory response mediated by transcription factor IRF3 and NF-κB [56]. Recently, the author established the relationship between cytosolic IRF3 and Wnt/β-catenin signalling via AOM/DSS-induced chronic colonic inflammation mice models, demonstrating that IRF3 is a tumour suppressor, inhibiting Wnt signalling, instead of functioning as a transcription factor. In normal physiology, the site of a.a. 634–663 in the exact armadillo repeats (ARM) of β-catenin links IRF3-β-catenin interaction, which retains β-catenin in the cytoplasm and inhibits the nuclear import of β-catenin. Gut microbiota trigger IRF3 phosphorylation and the structural/interface change of activated IRF3, resulting in IRF3 failing to bind β-catenin or active-β-catenin (β-catenin-S33A), thus losing its capacity to retain β-catenin in the cytoplasm. The IRF3-β-catenin axis links the gut microbiota to the proliferation of the intestinal epithelium and the development of CRC [57].
4.2. The Wnt Signalling Pathway in Colitis-Associated Carcinoma
Apart from sporadic CRC, colitis-associated CRC (CAC) can arise and has caught the attention of the public. A growing number of studies show that intestinal inflammation drives the initiation of cancer and its development [58]. Inflammatory bowel disease (IBD), including Crohn’s disease (CD) and ulcerative colitis (UC), is a chronic, idiopathic, relapsing inflammatory disease of the gastrointestinal (GI) tract [59]. Chronic and long-term IBD changes are prone to turn to colonic carcinoma. Dysregulation of the Wnt/β-catenin pathway, such as the GSK3β and APC gene, plays a vital role in CAC progression [60,61,62].
Inflammation-related pathways are activated in the early stages of CAC, such as NF-κB signalling [63]. Importantly, Wnt/β-catenin signalling could be activated as a key regulator of the CRC transformation of CAC [64]. The differences in IBD-CRC formation concern the loss of APC and p53 function. Compared with p53, loss of function occurs early in CAC carcinogenesis, loss of APC function is less frequent and takes place later in the development of malignancy [65,66]. The author characterised IBD-associated tumorigenesis by multiple high-throughput approaches and compared the results with in-house data sets from sCRCs. The results demonstrated that Wnt signalling dominated the mesenchymal stroma-rich subtype instead of the canonical epithelial tumour subtype in IBD-CRCs, which may affect prognosis and treatment options. Negative Wnt regulators Axin2 and RNF43 were strongly downregulated in IBD-CRCs and chromosomal gains at HNF4A, a negative regulator of Wnt-induced epithelial–mesenchymal transition (EMT), were less, which compared to sCRCs, while, polymeric immunoglobulin receptor (PIGR) and Oncostatin M Receptor (OSMR) were dysregulated via epigenetic modifications., which is involved in mucosal immunity. Therefore, the mechanism of Wnt pathway dysregulation indicated that IBD-CRCs toward mesenchymal tumour subtype instead of canonical epithelial tumour subtype. Meanwhile, somatic mutations in APC and Kirsten rat sarcoma viral oncogene (KRAS) were less frequent in IBD-CRCs compared to sCRCs [67]. Recently, research found the mechanism responsible for the crosstalk between NF-κB and Wnt/β-catenin signalling. Dvl2, an adaptor protein of Wnt/β-catenin signalling, interacts with the C-terminus of tumour necrosis factor receptor 1 (TNFR1) and mediates TNFR1 endocytosis, leading to NF-κB signal inhibition and promoting CAC progression. In addition, STAT6 negatively regulates Dvl2 levels and restrains cancer cell colony formation by interleukin- 13 (IL-13) [68].
Intestinal mucosal injury is an important pathological change in individuals with IBD [69]. Normal cell turnover of IECs provides the foundation to guarantee intestinal epithelial homeostasis; excessive IEC death is a well-known clinical hallmark of IBD [70]. The Wnt/β-catenin signalling pathway is highly conserved during evolution and is involved in other signalling pathways, such as BMP signalling, to drive the orderly self-renewal and differentiation of ISCs and maintain intestinal homeostasis [71]. c-Myc and Cyclin D1, the target genes of the Wnt signalling pathway, promote the proliferation of intestinal epithelial cells (IECs), enhancing the activity of ISCs [72]. Recent study has indicated that IL-36γ upregulated extracellular matrix and cell-matrix adhesion molecules and positively correlated with beta-catenin levels facilitated by Wnt signalling. Moreover, the activation Wnt signalling is mitigated by IL-36Ra or IL-36γ neutralising antibody, in which IL-36γ and IL-36Ra played the critical roles of in gut inflammation and tumorigenesis through Wnt/β-catenin [73]. MCL1, a member of the pro-survival BCL2 family, has been identified as having an essential role in the preservation of intestinal homeostasis and the prevention of carcinoma [74]. IEC-specific MCL1 deficiency shares hallmark features with IBD and is independent of microbe-induced chronic inflammation. Further investigation into it functions revealed strong γH2AX positivity and GSEA (from RNASeq data), indicating that MCL1 deletions induce DNA damage and abnormal Wnt signalling activation, with a marked increase in Wnt2b expression levels in the stroma surrounding the intestinal crypts, which promotes a clear proliferation-enhancing effect [75].
Wnt/β-catenin signalling axis in regulatory T cells promotes IBD and colonic dysplasia. Treg cells are crucial in response to the inflammatory milieu, RORγt is one of the transcription factors for multiple Treg cell subpopulations are described that express additional helper T (TH) cell lineage [76,77]. In regulatory T (Treg) cells, excessive Wnt/β-catenin activation may impact their function due to that TCF1 is known the bona fide DNA-binding partner of β-catenin [78]. Foxp3 binding to gene enhancers made accessible by Foxo1 regulates a core regulatory programme of late differentiation of Treg cells to define Treg cells [79]. The overlap of Foxp3 binding with the transcription factor TCF1 impacts Treg cell function [80]. Recent findings indicated that TCF1 and Foxp3 together limited the expression of pro-inflammatory genes in Treg cells, which function was interfered by activation of β-catenin signalling interferes with this function, and activation of β-catenin signalling promoted the disease-associated RORγ t(+) T-reg phenotype [81].
Apart from individual gene mutations and familial hereditary factors, hazardous unhealthy lifestyle, such as diet and disorder circadian clock, pro-tumorigenic bacteria and imbalanced gut microbiota, may cause imbalance of Wnt signalling pathway and increase the rates of occurrence of CRC, especially sporadic and early-onset CRC. At the same time, the role of Wnt signalling pathway in colitis-associated carcinoma should not be overlooked. Wnt signalling pathway has been found to have differences between sCRC and colitis-associated carcinoma.
5. The Wnt Signalling Pathway in the Development of CRC
5.1. The Wnt Signalling Pathway in Cancer Stem Cells (CSCs)
CSCs with pluripotency and self-renewal properties are considered responsible for the progression and recurrence of the disease and for tumour resistance [82,83,84]. A rapid turnover rate maintains intestinal homeostasis by the asymmetric division and proliferation of SCs at the base of the intestinal crypts, migration, and differentiation of the SCs at intestinal villus [85]. Imbalances in the signalling between the microenvironment and ISCs play a key role in promoting the growth and progression of established CRCs [82]. The Wnt signalling pathways participate in CRC survival, proliferation, and self-renewal properties, and most of them are dysregulated in CSCs [86]. Canonical Wnt signalling causes the expansion of rapidly cycling CSCs and modulates both immune surveillance and immune tolerance, while non-canonical Wnt signalling supports the maintenance of slow cycling to promote epithelial–mesenchymal transition and immune evasion via crosstalk with TGFβ (transforming growth factor-β) signalling cascades [87]. Wnt-related cancer stemness features relating to proliferation/dormancy plasticity will be discussed here, while other cancer stemness features will be discussed in the following section.
ISC dynamics are influenced by many endogenous and exogenous stimuli, including, for example, diet and the presence of niche factors. Leucine-rich repeat-containing G-protein coupled receptor 5 (Lgr5), the first functional relation marker of ISC features, is the Wnt target gene. Lgr5+ cells, which constitute an important component of the epithelial stem cell niche, are capable of long-term self-renewal and possess multilineage differentiation potential. The key signalling pathway controlling the ISC function is the Wnt/β-catenin pathway, and the Wnt and R-spondin ligands are extensively secreted by the niche, reaching their highest levels in the bottom of the crypt [88,89,90,91]. Although the Hippo transcriptional coactivator YAP is considered oncogenic in many tissues, its roles in intestinal homeostasis and CRC remain controversial. A study confirmed that YAP reprogrammes Lgr5+ proliferating CSCs into Lgr5- dormant CSCs, even if an Apc mutation via CRISPR-Cas9 into tetO-YAP rtTA colon organoids present constitutively active Wnt signalling pathway [92].
Wnt pathways are significant in terms of orchestrating intestinal stem cell and differentiation cues. SOX9, a transcriptional activator, is a key early mediator of CRC due to it binding to enhancers that upregulate genes associated with Paneth and stem cell function. SOX9 directly activates PROM1 using a Wnt/TCF4-responsive intronic enhancer to impair intestinal differentiation and promote CRC growth by a PROM1-SOX9 positive feedback loop [93]. ISCs play a crucial role in renewing the intestinal epithelium and maintaining tissue homeostasis [94]. Lgr5+ cells serve ISCs for the homeostatic balance of the intestine and can be a key driver of CSCs in intestinal tumorigenesis [95]. Many investigations have revealed that the Wnt signalling pathway contributes to the process of ISCs and CSCs; for example, β-catenin in ISCs is activated to promote the development of CRC due to a deficiency of APC [96]. Recently, researchers found the machinery for β-catenin activation using in vivo and in vitro experiments: the kinase MST4 phosphorylates β-catenin at Thr40 resulting in that β-catenin negatively influenced phosphorylation at Ser33 by GSK3β mediation. Subsequently beta-catenin binding to and being degraded by beta-TrCP was hindered, which causes subsequent accumulation and full activation of β-catenin, leading to increased numbers of ISCs/CSCs and exacerbated tumorigenesis [97].
Interestingly, some findings were conflicting for TCF4, especially a potential tumour suppressive role of the gene in intestinal cancer cells or tumours. Pathological transformation of the gut epithelium is driven by stabilized β-catenin by activation of Wnt/β-catenin, in which TCF4 is the major nuclear mediator. TCF4 was completely knockout in the adult colon that resulted in the formation of aberrant crypt foci (ACF), the earliest neoplastic lesions during CRC initiation [98]. A recent study reported the importance of mouse TCF4, mainly in adult intestinal epithelium homeostasis and intestinal tumour initiation. In contrast, in human cells, the TCF4 function is substituted by other LEF/TCF family members [99].
5.2. The Wnt Signalling Pathway in the Tumour Microenvironment
The tumour microenvironment (TME) is primarily composed of fibroblasts, immune cells, blood vessels, and extracellular matrix (ECM). Intercellular communications between tumour cells and the TME are mainly established through paracrine signalling, in which the Wnt/β-catenin pathway could be mediated by external factors to affect CRC progression [100]. Extracellular vesicles (EV) can transport mutant β-catenin and activate the Wnt signalling pathway in the recipient cells, thus promoting CRC progression. A recent study confirmed that Wnt2 acts in an autocrine manner, generating morphogenetic changes in fibroblasts and contributing to the invasive and metastatic capacity of CRC-derived cells and EVs play a key role in activating Wnt signalling in CRC [101]. In the solid tumour TME, hypoxia is a common effect, which may be mediated by β-catenin. Hypoxic CRC cells have been found to secrete exosomes enriched with Wnt4 ligands, resulting in the activation of β-catenin signalling in normoxic CRC cells to stimulate prometastatic behaviours, such as cell migration and invasion [102,103]. It has also been reported that Wnt5a has both tumour promoting and suppressing functions in CRC [104,105]. Researchers found that Wnt5a was overexpressed in tumour stroma in vivo and in vitro, especially M2-like tumour-associated macrophages (TAMs), and Wnt5a induced M2 polarisation of TAMs by regulating CaKMII-ERK1/2-STAT3 pathway-mediated IL-10 secretion [105], enhancing tumour growth, invasion and metastasis of CRC, and recruited macrophages whose infiltration depended on CaMKII-ERK pathway-mediated CCL2 secretion [106].
5.3. The Wnt Signalling Pathway in Angiogenesis
Angiogenesis is one of the key mechanisms of tumour development and is critical for invasive tumour growth, metastasis [107]. It has long been suggested that the Wnt/FZD/β-catenin signalling pathway regulates the development of blood vessels in physiological and pathological conditions. Sprouting angiogenesis is an essential condition for tumour growth and metastasis, to access the blood system in order to maintain sufficient oxygen and nutrients [108,109]. Tumour angiogenesis is mediated by tumour cells, cancer-associated fibroblasts (CAFs), and immune cells in the tumour stroma, and by breaking the balance of pro- and anti-angiogenic factors [110]. A high expression of Wnt2 was found in stromal CAFs and in the autocrine activation of canonical Wnt signalling and enhanced fibroblast motility [111]. Subsequently, researchers found that Wnt2 derived from stromal CRC-CAFs enhances angiogenesis by increasing extracellular migration and invasion and by altering the CAF secretome towards pro-angiogenic factors and ECM remodelling signals [112]. Long non-coding RNA (lncRNA) GAS5 inhibited the activation of the Wnt/β-catenin signalling pathway, thereby suppressing the angiogenesis, invasion, and metastasis of CRC [113]. TRIM24 was highly expressed in CRC tissues compared to nonneoplastic adjacent tissues; the overexpression of TRIM24 upregulates the expression of vascular endothelial growth factor (VEGF), CCL2/5, and CSF-1 via Wnt/β-catenin signalling to stimulate angiogenesis and recruited TAMs, which promotes CRC tumour growth [114].
Wnt4, a member of the Wnt family, has been reported in wound healing, acute kidney injury, and angiogenesis and may play a role in CRC via the β-catenin- relevance Wnt pathway. A recent study found that Wnt4 is highly express in the serum of CRC patients and in tumour tissues. Wnt4 prompted the nuclear translocation of β-catenin and increased the expression of α-SMA and fibronectin in NFs, which triggered the transformation of NFs into CAFs and enhanced reinforced cell contraction and microsphere formation. Meanwhile, ANG2, a critical gene for angiogenesis, could be abnormally activated by the Wnt4/β-catenin/ANG2 pathway, resulting in enhanced angiogenesis in CRC. Wnt4 may also be secreted as exosomes from CRC cells, playing a crucial role in promoting tumour progression in the TME [115]. Exosomes are nano-sized membrane vesicles with diameters between 30–100 nm and are generated from endosomal compartment invaginations [116]. CRC cell-derived exosomes have important roles in tumour progression through effectively delivering microRNAs, mRNAs, and proteins [117,118,119]. The protein levels of Wnt4 were higher in hypoxic exosomes than in exosomes derived from normoxic cells. In hypoxia, Wnt4-loaded exosomes secreted by the tumour cells promoted angiogenesis through the proliferation and migration of ECs via activated Wnt/β-catenin signalling [103,120].
The Wnt signalling pathway plays crucial effect on cancer stem cells, tumour microenvironment, and angiogenesis to drive the development of CRC. The complex microenvironment components, such as EV and exosomes, also contribute to development of CRC via Wnt signalling pathway.
6. The Wnt Signalling Pathway in the Invasion and Metastasis of CRC
CRC is a heterogeneous disease and lethality is especially common in the subtype of CRC with high stromal infiltration. As metastasis is a main reason for cancer-related deaths, and an aberrant Wnt/β-catenin signalling pathway causes CRC invasion and metastasis, it is therefore significant to understand the effect of the Wnt signalling pathway in invasion and metastasis in CRC [121]. In the nucleus, together with TCF/LEF, β-catenin binds to the promoter region of a target gene, and activates its transcription such as Twist and Slug, as E-cadherin transcription factors, which regulate EMT [122,123].
During invasive and metastatic cancer development, both the signalling and gene expression of cells to neighbouring cells and the coordination between cell–cell adhesion is changed. β-catenin acts as an adhesion linker for cadherin transmembrane receptors to the cytoskeleton. L1CAM (L1) cell adhesion receptors, a downstream target oncogene of β-catenin-TCF, were detected as regulators of invasive and metastatic CRC cells [124,125,126]. RNF128 inhibited cell proliferation and metastasis of CRC cells via Wnt/β-catenin signalling-mediated deubiquitination [127]. SMYD2 overexpression promoted cell proliferation but also increased the metastatic ability of CRC and may activate the Wnt/β-catenin pathway, as SMYD2 overexpression suppresses the expression of adenomatous polyposis coli 2 (APC2), an inhibitor of the Wnt/β-catenin pathway. The low APC2 expression in CRC cells was due to the modification of SMYD2-mediated DNA methylation [128]. TM4SF1 promoted epithelial–mesenchymal transition (EMT) and cancer stemness via the Wnt/β-catenin/SOX2 pathway in CRC, which revealed that TM4SF1 modulated SOX2 expression in a Wnt/β-catenin activation-dependent manner [129]. Mechanistically, WDR74 decreased the phosphorylation of β-catenin and induced the nuclear accumulation of β-catenin, activating the Wnt/β-catenin signalling pathway in CRC cells [130]. B7-H4 overexpression promoted the proliferation and invasion of CRC cells, which may be actioned by Wnt signalling. FOXS1 could also function as an oncogene, promoting CRC cell proliferation, migration, invasion, and metastasis through the Wnt/β-catenin signalling pathway [131]. Overexpressed FOXO4 inhibited the migration and metastasis of CRC cells by enhancing the APC2/β-catenin axis, suggesting that FOXO4 is a potential therapeutic target of CRC [132]. FUBP1 is an oncogene that initiates the development of CSCs by activating Wnt/β-catenin signalling via direct binding to the promoter of Dvl1, resulting in increased pluripotent transcription factors, including c-Myc, NANOG, and SOX2 [133].
Metabolic reprogramming is a hallmark of cancer cells, supporting their growth and metastasis [134,135]. The metastasis of cancer cells to distant organs requires the ability to adapt to metabolic conditions distinct from those at the primary tumour site. C14orf159, a d-amino acid metabolising enzyme, is located in the mitochondrial matrix to maintain the mitochondrial membrane potential and has the function of reducing the expression of genes involved in CRC metastasis. In CRC, C14orf159 is involved in Wnt6 expression and β-catenin activation via regulation of the mitochondrial membrane potential [136,137,138].
EMT is a critical process triggered during tumour metastasis, by which tumour cells convert from epithelial cells to mesenchymal cells, losing cell polarity and the ability for cell adhesion, allowing cancer cells to easily spread into the lymphatic system and blood circulation, invade surrounding tissues and migrate to distant organs, primarily including the liver and lung [139,140,141]. Elevated secretion levels of Wnt7b were significantly associated with lymphatic and remote metastasis and predicted a poor outcome in patients with CRC. Researchers found that either a partial knockdown of Wnt7b or a blockade of the Wnt/β-catenin signalling pathway reversed the EMT process and inhibited the migration of CRC cells [142]. Neuroligin 1 (NLGN1) promoted APC localisation to the cell membrane, and co-immunoprecipitates with some isoforms of this protein stimulated β-catenin translocation to the nucleus, upregulating mesenchymal markers and Wnt target genes and inducing an “EMT phenotype” in CRC cell lines [143]. Claudin-7 (Cldn7), a tight junction protein, was recently reported to function as a candidate tumour suppressor gene in CRC. Cldn7 deficiency conferred stemness properties in CRC through SOX9-mediated Wnt/β-catenin signalling to promote cellular EMT [144]. Chemotherapeutic resistance is the key characteristic of CSCs and contributes to postoperative recurrence and metastasis. The activation of LRP5 could affect the stem-like properties of CRC cells by activating the canonical Wnt/β-catenin pathway. Activation of LRP5 induced the drug resistance of CRC cells to platinum-based drugs [145].
Wnt/PCP signalling components are frequently dysregulated in solid tumours, and aberrant pathway activation contributes to tumour cell migratory properties [146]. Wnt protein is secreted on the surface of exosomes [147]. Wnt1 expression on CRC cell–derived exosomes was considerable and exosomal Wnt1 substantially enhanced the capacity for CRC cell proliferation and migration by activating the factors RHO, JNK, both of the downstream of Wnt/PCP signalling pathway [148]. Wnt2 involved in invasive activity of CRC cells through Wnt/PCP signalling pathway coupled to GSK3β and c-Jun/AP-1 signalling [149]. Meanwhile, overexpression of Wnt5b also induced the proliferation, migration, and invasion of CRC cells by activating the non-canonical Wnt/JNK signalling [150]. Wnt11 signalling diminished the E-cadherin-mediated cell–cell contact signalling resulting from stimulates proliferation and promotes morphological transformation in the intestinal epithelial cells, via enhancing the activation of protein kinase C and Ca2+/CaMKII [151]. FZD7 initiates both canonical and noncanonical Wnt signalling pathways to control cell motility, metastasis, and invasion. Significantly, Wnt11/FZD7 receptors binding activated Wnt/PCP signalling pathway to promote CRC cell proliferation, migration, and invasion activities through phosphorylation of JNK and c-Jun [152,153,154].
The downstream genes involved EMT express by activating Wnt/β-catenin pathway. The Wnt/PCP signalling pathway plays key roles of in guiding cell polarity and tumour cell invasiveness.
7. The Wnt Signalling Pathway in CRC Prognosis
7.1. The Wnt Signalling Pathway in Chemotherapy Drug Resistance
Chemotherapy is common for the treatment of CRC; among the most used chemotherapeutic drugs, approved as first- and second-line adjuvants in CRC, are 5-fluorouracil (5-FU), irinotecan (CPT-11), oxaliplatin capecitabine, and leucovorin [155]. For high-level microsatellite instability/deficient mismatch repair metastatic CRC, chemotherapy is also used in combination with immune checkpoint inhibitors, such as bevacizumab or cetuximab [156]. However, the treatment of patients with advanced grades of metastatic disease is unsatisfactory, mainly due to chemoresistance. Membrane transporters including pump P-glycoprotein (P-gp), the ATP-binding cassette (ABC) superfamily, and the multidrug resistance-associated protein (MRP) subfamily, induce the ability of tumour cells to efflux drugs, which could pump out chemotherapeutic agents, resulting in drug resistance [157]. CSC-mediated multidrug resistance, an altered TME, and the functions of some noncoding RNAs are also associated with chemoresistance in CRC [158]. Meanwhile, it has increasingly been known that the Wnt signalling pathway contributes to cancer drug resistance [159].
CRC stem cells (CRC-SCs) have some unique features, such as a high level of heterogeneity and plasticity, and also participate in drug resistance and cancer recurrence. During the treatment of patients with CRC, the genetic contents of SCs and niche of SCs aberrantly activate Wnt signalling pathways [160], leading to the accruing of CRC-SCs that are resistant to chemotherapy and radiotherapy. Side population cells (SP), as a group of CSCs, have obvious worldwide regard for their association with tumorigenesis and drug resistance, including CRC [161]. The protein tyrosine phosphatase receptor type C (PTPRC) encoded CD45, which correlates with the stemness of CRC cells, was increased in remnant tumour tissues after expression in both 5-FU-treated and irradiated CRC cells. CD45 binds only to degradable β-catenin and not to active β-catenin, reducing the tyrosine phosphorylation levels of degradable β-catenin by colocalisation with only total and degradable β-catenin at the cellular membrane, which collectively enhances stemness and the therapy-resistant phenotype [162]. Forkhead Box M1 (FOXM1) also participated in the malignant behaviours of CRC in which the Wnt/β-catenin signalling pathway is involved. FOXM1 bound to Dvl2 and enhanced the nuclear translocation of Dvl2 and the Dvl2-mediated transcriptional activity of Wnt/β-catenin, known to induce metastasis and drug resistance [163]. RAI2, an independent poor prognostic marker, inhibited Wnt signalling by interacting with or downregulating CtBP2, resulting in the repression of stem cell-like properties and the increased chemosensitivity of CRC cells to oxaliplatin and fluorouracil [164]. LGR5, a marker of CSCs, is a target gene of the Wnt pathway [130]. LGR5-positive CSCs are chemotherapy-resistant, making the cells enter a static state to escape drug treatment [165]. Activation of the LRP5 gene promotes CSC-like phenotypes, including tumorigenicity and platinum-based drug resistance in CRC cells, through activation of the canonical Wnt/β-catenin and IL-6/STAT3 signalling pathways [145]. The drug efflux effect of ABC transporters induces chemical resistance in numerous solid tumours [166]. Studies have shown that Wnt/β-catenin signalling is closely related to the ABC transporter of CSCs [167]; for example, the pivotal role of the c-Myc-ABCB5 axis in 5-FU resistance in CRC cell [168]. SP cells showed more resistance to 5-FU and irinotecan than non-SP cells, as well as a higher activation of the Wnt signalling pathway; inhibiting β-catenin significantly decreased SP [169]. The author found that silencing the SNTB1 expression involved in the Wnt/β-catenin pathway resulted in the downregulation of several β-catenin downstream proteins, such as c-Myc and cell cycle modulator CCND1, and, importantly, decreased the percentage and clonogenicity of SP cells [170].
The TME is a “complex network” of different cell types, various cytokine signalling molecules, and ECM components [171]. Wnt signalling pathway crosstalk between cancer cells and the surrounding supportive stromal cells in the TME promotes drug resistance [100]. CAFs are a key component of the TME and contribute to drug resistance, interacting with Wnt/β-catenin signalling [172,173,174]. Clear colocalisation between CAFs and tumour cells expressing nuclear β-catenin is observed in primary CRC samples. CAF-derived exosomes contribute to the secretion and carrying of Wnt ligands to activate Wnt/β-catenin signalling to regulate Wnt activity, resulting in the function of CSC characteristics being promoted [175]. Wnt16B and stable free radical polymerisation 2 (SFRP2) were produced by CAFs treated with chemotherapy, which synergistically activated the canonical Wnt pathway in cancer cells to reduce treatment sensitivity [176,177]. In hypoxia, hypoxia-inducible factor-1α/2α leads to the upregulation of the key Wnt coactivator BCL-9, and this crosstalk synergistically acts on the development of CRC resistance [178].
Metabolic reprogramming is one of the important hallmarks of drug resistance. Recent research found the effect of glucose metabolic reprogramming events of 5-FU resistance in CRC. The upregulation of HIF-1α, classical signalling the state of oxygen deficiency, promotes metabolic reprogramming that facilitates 5-FU resistance in CRC via reactive oxygen species (ROS) and the Wnt/β-catenin signalling pathway, instead of in the classical ways according to external oxygen concentrations [179]. Sec62 expression was upregulated by the m(6)A-mediated stabilisation of Sec62 mRNA, which restrains the sensitivity of CRC cells to chemotherapeutic drugs. Sec62 promoted the stemness of CRC cells through the activation of Wnt/β-catenin signalling. Mechanistically, Sec62 can bind to β-catenin and competitively disrupt the interaction between β-catenin and APC to inhibit the assembly of the β-catenin destruction complex, destroying the degradation of β-catenin [180]. Carnitine palmitoyltransferase 2 (CPT2) was decreased in CRC, and CPT2 downregulation could trigger stemness and oxaliplatin resistance in CRC via the activation of ROS/Wnt/β-catenin-induced glycolytic metabolism [181].
It has been increasingly recognised that long non-coding RNA (lncRNA, a non-coding regulatory RNA with greater than 200 nucleotides) regulates the Wnt/β-catenin pathway in multidrug-resistant CRC. Small nucleolar RNAs (snoRNAs) are a subset of nuclear non-coding (nc)RNAs with lengths of 60 to 300 nucleotides. The C/D box snoRNA SNORD1C was highly expressed in CRC to enhance cancer cell stemness and 5-FU resistance via the Wnt/β-catenin pathway, resulting in downregulation of c-Myc downstream, as well as other stem cell regulatory genes, such as CD44 and SOX2 [182].
7.2. The Wnt Signalling Pathway in Immunotherapy Drug Resistance
In recent times, immunotherapy has shown promising success in enhancing the treatment responses in some CRCs [183]. For example, nivolumab and pembmlizumab (Programmed cell death-1 blocking antibodies) have shown efficacy in the mismatch repair deficient high microsatellite instability (dMMR-MSI-H) subtype of metastatic CRC patients [183]. Immune checkpoint inhibitors such as nivolumab and pembrolizumab could reverse T cell dysfunction and apoptosis by inhibiting the effects of the interaction of programmed death-1 (PD-1) and its ligand PD-L1 to enhance T cell activation and cytotoxicity to tumour cells [184,185]. Moreover, chimeric antigen receptor-modified T (CAR-T) cells or cancer vaccines have also contributed to the development of immunotherapy for CRC [186]. However, only about 20% of patients respond to immune checkpoint blockade. Importantly, even if patients derived benefit from this treatment, the effect was not satisfactory due to developing resistance [187,188]. Preclinical and clinical models demonstrate that WNT/β-catenin activation leads to the suppression of CD8+ T-cell tumour infiltration and the evasion of immune elimination, which has been linked to decreased chemokine levels [189,190,191]. β-catenin could suppress the transcription of T-cell recruiting chemokine CCL4 from dendritic cells, which reduced the infiltration of CD8+ T cells within the TME in colon cancer [192]. Moreover, when WNT/β-catenin aberrantly activates, the recruitment of T cells to the TME is impaired, as a result of the failing recruitment of specific dendritic cells into the tumour leading to a lack of tumour-infiltrating T cells [193,194]. Mutations in components of the WNT/β-catenin signalling pathway induced downstream MYC expression and controlled PD-L1 and CD47 transcription, disfavouring the accumulation of tumour-associated T cells and macrophages [195]. Fermitin family member 3 (FERMT3) promoted the blockage of PD-L1 by FERMT3-mediated Wnt/β-catenin signalling to suppress CRC cell invasion, 5-FU resistance and NK cells-mediated tumour killing [196].
7.3. The Wnt Signalling Pathway as a Prognostic Indicator of CRC
The Wnt/β-catenin pathway plays an important role in advanced colorectal carcinoma [197]. Recently, a meta-analysis was performed to assess the prognostic value of β-catenin expression in CRC patients. High nuclear β-catenin expression was significantly associated with poorer disease-free survival (DFS), cancer-specific survival (CSS), and overall survival (OS) in patients with CRC whereas, low membranous β-catenin expression was associated with poor OS [198]. In univariate analysis, TCF4 expression turned out to be a negative prognostic factor being associated with shorter overall survival, whereas LEF1 expression as well as a LEF1/TCF4 ratio were positive prognostic factors and correlated with longer overall survival [199].
According to clinical, molecular, and pathologic features, mutations of F-box and WD repeat domain-containing 7 (FBXW7), a member of the Dickkopf (DKK) family, result in aberrant cytoplasmic and nuclear β-catenin accumulation, although at a lower rate than CTNNB1-mutated CRC [200]. S100A4, a Wnt/β-catenin target gene, is an established prognostic biomarker for CRC patient survival. The cross-regulation of S100A4 with Wnt pathway antagonist Dickkopf-1 (DKK1) resulted in S100A4 overexpression downregulating DKK1, while S100A4 knock-down increased DKK1. Significantly, S100A4 can be seen as the predominant factor in this feedback loop in Wnt signalling modulation. The combination of S100A4 and DKK1 improved the identification of CRC patients at high risk and improved the prognosis of overall and metastasis-free survival [201]. Overexpression of UBE2M in CRC specimens contributed to a decreased overall survival of patients and mediated 5-FU and oxaliplatin resistance in CRC cells via the Wnt/β-catenin signalling pathway [202]. Paired-like homeodomain transcription factor 2 (PITX2) could act as an effective biomarker in human cancer diagnosis and prognosis progression of CRC; PITX2 enhanced resistance to 5-FU in CRC, upregulating the Wnt/β-catenin axis [203].
It is known that the Wnt signalling pathway contributes to cancer drug resistance. Wnt signalling pathway could also interfere with immunotherapy. Moreover, prognostic factors associated with Wnt signalling are also noteworthy.
8. Challenges and Prospects
A large number of aspects of the Wnt pathway have been studied in CRC, the roles of therapeutic targets for CRC have been previously established. However, these extensive studies suggest that the regulation of Wnt signalling is more complicated than previously hypothesized, and thus various information still needs to be further clarified. Some colorectal tumours exhibit a reverse correlation between Wnt signalling levels and patient survival, due to the variability in signalling levels and driver mutations across different tissues [204], as well as the complexity of components involved in this pathway, including β-catenin, Axin, APC, GSK3β, Dvl, CK I, and the TCF/LEF family, in addition Wnt signalling play important role in homeostasis, which cause obstacles in developing Wnt-suppressing therapy [205]. To date, there are no FDA-approved Wnt signalling inhibitors in clinical use.
Admittedly, with the development of targeted drugs and combination therapy strategies, many efforts have been made over the years to targeted intervention of Wnt signal transduction in CRC. In addition, several inhibitors have entered clinical studies [206]. For example, OMP-131R10 has been shown to inhibit the growth and metastasis of advanced CRC in phase I clinical trials [207].
CRC heterogeneity may render the difficulty to improve the rates of patients surviving. Thus, further insight into Wnt signalling pathway may enable deeper understanding of CRC to solve the clinical problems of tumour metastasis, recurrence and chemoresistance. Therefore, further studies may focus on the following: (i) Identifying the connection between Wnt signalling pathway and immunotherapy may be a promising therapeutic strategy for CRC. (ii) Researching the feasibility of certain multitarget therapeutic strategies concurrently may improve treatment. (iii) Understanding the crosstalk among oncogenes may cope with the complexity of CRC. (iv) Finding more information about components of Wnt signalling may provide opportunities to research the highly selective inhibitor for CRC to avoid unnecessary side effects.
9. Conclusions
The Wnt/β-catenin pathway affects almost all physiological and biochemical process. The Wnt1 and Wnt2 usually involved in Wnt/PCP signalling pathway, which plays key roles in guiding cell polarity and tumour cell invasiveness. Wnt5 and Wnt11 can have both oncogenic and tumour-suppressive functions. Their tumour-suppressing activities are mainly attributed to the activation of Wnt/Ca2+ signalling that may inhibit the Wnt/β-catenin signalling. Over the years, the molecular mechanisms and functional effects of the Wnt signalling pathway have been accumulatively studied at all stages of CRC (Figure 3). However, improving the cure rate and extended survival of CRC remains a challenge. Nevertheless, owing to the complexity of CRC, numerous details remain to be uncovered with regard to its connections to the Wnt signalling pathway, which would provide clearer guidance in individualised treatment and promote the development of specific inhibitors or drug combinations and prognosis markers to improve anticancer efficacy. It is hoped that immunotherapy can eradicate cancer. For CRC, the treatment response of patients remains unsatisfactory due to the differences between individuals.
Funding
This work was supported by grants from the National Natural Science Foundation of China (No. 82273992 and 82073870).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Steinhart, Z.; Angers, S. Wnt Signaling in Development and Tissue Homeostasis. Development 2018, 145, dev146589. [Google Scholar] [CrossRef] [Green Version]
- Taciak, B.; Pruszynska, I.; Kiraga, L.; Bialasek, M.; Krol, M. Wnt Signaling Pathway in Development and Cancer. J. Physiol. Pharmacol. 2018, 69, 185–195. [Google Scholar] [CrossRef]
- Nie, X.; Liu, H.; Liu, L.; Wang, Y.-D.; Chen, W.-D. Emerging Roles of Wnt Ligands in Human Colorectal Cancer. Front. Oncol. 2020, 10, 1341. [Google Scholar] [CrossRef] [PubMed]
- Parichha, A.; Suresh, V.; Chatterjee, M.; Kshirsagar, A.; Ben-Reuven, L.; Olender, T.; Taketo, M.M.; Radosevic, V.; Bobic-Rasonja, M.; Trnski, S.; et al. Constitutive Activation of Canonical Wnt Signaling Disrupts Choroid Plexus Epithelial Fate. Nat. Commun. 2022, 13, 633. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.; Arnold, M.; Gini, A.; Lorenzoni, V.; Cabasag, C.J.; Laversanne, M.; Vignat, J.; Ferlay, J.; Murphy, N.; Bray, F. Global Burden of Colorectal Cancer in 2020 and 2040: Incidence and Mortality Estimates from GLOBOCAN. Gut 2022, 72, 338–344. [Google Scholar] [CrossRef]
- Xi, Y.; Xu, P. Global Colorectal Cancer Burden in 2020 and Projections to 2040. Transl. Oncol. 2021, 14, 101174. [Google Scholar] [CrossRef]
- Ochs-Balcom, H.M.; Kanth, P.; Cannon-Albright, L.A. Early-Onset Colorectal Cancer Risk Extends to Second- and Third-Degree Relatives. Cancer Epidemiol. 2021, 73, 101973. [Google Scholar] [CrossRef]
- Gausman, V.; Dornblaser, D.; Anand, S.; Hayes, R.B.; O’Connell, K.; Du, M.; Liang, P.S. Risk Factors Associated with Early-Onset Colorectal Cancer. Clin. Gastroenterol. Hepatol. 2020, 18, 2752–2759. [Google Scholar] [CrossRef]
- Aran, V.; Victorino, A.P.; Thuler, L.C.; Ferreira, C.G. Colorectal Cancer: Epidemiology, Disease Mechanisms and Interventions to Reduce Onset and Mortality. Clin. Colorectal Cancer 2016, 15, 195–203. [Google Scholar] [CrossRef]
- Koni, M.; Pinnaro, V.; Brizzi, M.F. The Wnt Signalling Pathway: A Tailored Target in Cancer. Int. J. Mol. Sci. 2020, 21, 7697. [Google Scholar] [CrossRef]
- Clevers, H.; Nusse, R. Wnt/Beta-Catenin Signaling and Disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, X.; Xu, X.; Chen, D.; Zhao, F.; Wang, W. Therapeutic Potential of Targeting the Wnt/Beta-Catenin Signaling Pathway in Colorectal Cancer. Biomed. Pharmacother. 2019, 110, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Li, V.S.W.; Ng, S.S.; Boersema, P.J.; Low, T.Y.; Karthaus, W.R.; Gerlach, J.P.; Mohammed, S.; Heck, A.J.R.; Maurice, M.M.; Mahmoudi, T.; et al. Wnt Signaling through Inhibition of Beta-Catenin Degradation in an Intact Axin1 Complex. Cell 2012, 149, 1245–1256. [Google Scholar] [CrossRef] [Green Version]
- Mantilla, C.; Suárez Mellado, I.; Duque Jaramillo, A.; Navas, M.C. β-catenin signaling mechanisms and its role in carcinogenesis. CES Med. 2015, 29, 109–127. [Google Scholar]
- Kim, N.-G.; Xu, C.; Gumbiner, B.M. Identification of Targets of the Wnt Pathway Destruction Complex in Addition to Beta-Catenin. Proc. Natl. Acad. Sci. USA 2009, 106, 5165–5170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latres, E.; Chiaur, D.S.; Pagano, M. The Human F Box Protein Beta-Trcp Associates with the Cul1/Skp1 Complex and Regulates the Stability of Beta-Catenin. Oncogene 1999, 18, 849–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Semenov, M.; Tamai, K.; Zeng, X. LDL Receptor-Related Proteins 5 and 6 in Wnt/Beta-Catenin Signaling: Arrows Point the Way. Development 2004, 131, 1663–1677. [Google Scholar] [CrossRef] [Green Version]
- Kishida, S.; Yamamoto, H.; Hino, S.; Ikeda, S.; Kishida, M.; Kikuchi, A. DIX Domains of Dvl and Axin Are Necessary for Protein Interactions and Their Ability to Regulate β-Catenin Stability. Mol. Cell. Biol. 1999, 19, 4414. [Google Scholar] [CrossRef] [Green Version]
- Cadigan, K.M.; Waterman, M.L. TCF/LEFs and Wnt Signaling in the Nucleus. Cold Spring Harb. Perspect. Biol. 2012, 4, a007906. [Google Scholar] [CrossRef] [Green Version]
- Kikuchi, A.; Kishida, S.; Yamamoto, H. Regulation of Wnt Signaling by Protein-Protein Interaction and Post-Translational Modifications. Exp. Mol. Med. 2006, 38, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Chen, Z.; Tang, Y.; Xiao, Q. The Involvement of Noncanonical Wnt Signaling in Cancers. Biomed. Pharmacother. 2021, 133, 110946. [Google Scholar] [CrossRef] [PubMed]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt Signaling in Cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, C.A.; VanderVorst, K.; Carraway, K.L. Vangl as a Master Scaffold for Wnt/Planar Cell Polarity Signaling in Development and Disease. Front. Cell Dev. Biol. 2022, 10, 887100. [Google Scholar] [CrossRef] [PubMed]
- Devenport, D. The Cell Biology of Planar Cell Polarity. J. Cell Biol. 2014, 207, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Gao, B.; Song, H.; Bishop, K.; Elliot, G.; Garrett, L.; English, M.A.; Andre, P.; Robinson, J.; Sood, R.; Minami, Y.; et al. Wnt Signaling Gradients Establish Planar Cell Polarity by Inducing Vangl2 Phosphorylation through Ror2. Dev. Cell 2011, 20, 163–176. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Garrett, L.; Feng, D.; Elliott, G.; Liu, X.; Wang, N.; Wong, Y.M.; Choi, N.T.; Yang, Y.; Gao, B. Wnt-Induced Vangl2 Phosphorylation Is Dose-Dependently Required for Planar Cell Polarity in Mammalian Development. Cell Res. 2017, 27, 1466–1484. [Google Scholar] [CrossRef] [Green Version]
- Schlessinger, K.; Hall, A.; Tolwinski, N. Wnt Signaling Pathways Meet Rho GTPases. Genes Dev. 2009, 23, 265–277. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Feng, D.; Gao, B. An Overview of Potential Therapeutic Agents Targeting WNT/PCP Signaling. Handb. Exp. Pharmacol. 2021, 269, 175–213. [Google Scholar] [CrossRef]
- Rogers, S.; Scholpp, S. Vertebrate Wnt5a-At the Crossroads of Cellular Signalling. Semin. Cell Dev. Biol. 2022, 125, 3–10. [Google Scholar] [CrossRef]
- Kühl, M.; Sheldahl, L.C.; Park, M.; Miller, J.R.; Moon, R.T. The Wnt/Ca2+ Pathway: A New Vertebrate Wnt Signaling Pathway Takes Shape. Trends Genet. 2000, 16, 279–283. [Google Scholar] [CrossRef]
- Kühl, M. The WNT/Calcium Pathway: Biochemical Mediators, Tools and Future Requirements. Front. Biosci. J. Virtual Libr. 2004, 9, 967–974. [Google Scholar] [CrossRef] [PubMed]
- De, A. Wnt/Ca2+ Signaling Pathway: A Brief Overview. Acta Biochim. Biophys. Sin. 2011, 43, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Vuong, L.T.; Mlodzik, M. Different Strategies by Distinct Wnt-Signaling Pathways in Activating a Nuclear Transcriptional Response. Curr. Top. Dev. Biol. 2022, 149, 59–89. [Google Scholar] [CrossRef] [PubMed]
- Correction: The CBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov. 2012, 2, 960. [CrossRef] [Green Version]
- Joslyn, G.; Carlson, M.; Thliveris, A.; Albertsen, H.; Gelbert, L.; Samowitz, W.; Groden, J.; Stevens, J.; Spirio, L.; Robertson, M. Identification of Deletion Mutations and Three New Genes at the Familial Polyposis Locus. Cell 1991, 66, 601–613. [Google Scholar] [CrossRef]
- Segditsas, S.; Tomlinson, I. Colorectal Cancer and Genetic Alterations in the Wnt Pathway. Oncogene 2006, 25, 7531–7537. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Chan, A.T.; Sun, J. Influence of the Gut Microbiome, Diet, and Environment on Risk of Colorectal Cancer. Gastroenterology 2020, 158, 322–340. [Google Scholar] [CrossRef]
- Afrin, S.; Giampieri, F.; Gasparrini, M.; Forbes-Hernandez, T.Y.; Cianciosi, D.; Reboredo-Rodriguez, P.; Zhang, J.; Manna, P.P.; Daglia, M.; Atanasov, A.G.; et al. Dietary Phytochemicals in Colorectal Cancer Prevention and Treatment: A Focus on the Molecular Mechanisms Involved. Biotechnol. Adv. 2020, 38, 107322. [Google Scholar] [CrossRef]
- Seiwert, N.; Heylmann, D.; Hasselwander, S.; Fahrer, J. Mechanism of Colorectal Carcinogenesis Triggered by Heme Iron from Red Meat. Biochim. Biophys. Acta-Rev. Cancer 2020, 1873, 188334. [Google Scholar] [CrossRef]
- Liu, F.; Zuo, X.; Liu, Y.; Deguchi, Y.; Moussalli, M.J.; Chen, W.; Yang, P.; Wei, B.; Tan, L.; Lorenzi, P.L.; et al. Suppression of Membranous LRP5 Recycling, Wnt/Beta-Catenin Signaling, and Colon Carcinogenesis by 15-LOX-1 Peroxidation of Linoleic Acid in PI3P. Cell Rep. 2020, 32, 108049. [Google Scholar] [CrossRef]
- Kim, T.-Y.; Kim, S.; Kim, Y.; Lee, Y.-S.; Lee, S.; Lee, S.-H.; Kweon, M.-N. A High-Fat Diet Activates the BAs-FXR Axis and Triggers Cancer-Associated Fibroblast Properties in the Colon. Cell. Mol. Gastroenterol. Hepatol. 2022, 13, 1141–1159. [Google Scholar] [CrossRef] [PubMed]
- Rijo-Ferreira, F.; Takahashi, J.S. Genomics of Circadian Rhythms in Health and Disease. Genome Med. 2019, 11, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koike, N.; Yoo, S.-H.; Huang, H.-C.; Kumar, V.; Lee, C.; Kim, T.-K.; Takahashi, J.S. Transcriptional Architecture and Chromatin Landscape of the Core Circadian Clock in Mammals. Science 2012, 338, 349–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altman, B.J.; Hsieh, A.L.; Sengupta, A.; Krishnanaiah, S.Y.; Stine, Z.E.; Walton, Z.E.; Gouw, A.M.; Venkataraman, A.; Li, B.; Goraksha-Hicks, P.; et al. MYC Disrupts the Circadian Clock and Metabolism in Cancer Cells. Cell Metab. 2015, 22, 1009–1019. [Google Scholar] [CrossRef] [Green Version]
- Chun, S.K.; Fortin, B.M.; Fellows, R.C.; Habowski, A.N.; Verlande, A.; Song, W.A.; Mahieu, A.L.; Lefebvre, A.E.Y.T.; Sterrenberg, J.N.; Velez, L.M.; et al. Disruption of the Circadian Clock Drives Apc Loss of Heterozygosity to Accelerate Colorectal Cancer. Sci. Adv. 2022, 8, eabo2389. [Google Scholar] [CrossRef] [PubMed]
- Bennedsen, A.L.B.; Furbo, S.; Bjarnsholt, T.; Raskov, H.; Gogenur, I.; Kvich, L. The Gut Microbiota Can Orchestrate the Signaling Pathways in Colorectal Cancer. Apmis 2022, 130, 121–139. [Google Scholar] [CrossRef]
- Cai, J.; Sun, L.; Gonzalez, F.J. Gut Microbiota-Derived Bile Acids in Intestinal Immunity, Inflammation, and Tumorigenesis. Cell Host Microbe 2022, 30, 289–300. [Google Scholar] [CrossRef]
- Liu, X.; Lu, R.; Wu, S.; Sun, J. Salmonella Regulation of Intestinal Stem Cells through the Wnt/Beta-Catenin Pathway. FEBS Lett. 2010, 584, 911–916. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Lu, R.; Fu, X.; Dan, Z.; Zhang, Y.-G.; Chang, X.; Liu, Q.; Xia, Y.; Liu, X.; Sun, J. Novel Regulatory Roles of Wnt1 in Infection-Associated Colorectal Cancer. Neoplasia 2018, 20, 499–509. [Google Scholar] [CrossRef]
- Iftekhar, A.; Berger, H.; Bouznad, N.; Heuberger, J.; Boccellato, F.; Dobrindt, U.; Hermeking, H.; Sigal, M.; Meyer, T.F. Genomic Aberrations after Short-Term Exposure to Colibactin-Producing E. Coli Transform Primary Colon Epithelial Cells. Nat. Commun. 2021, 12, 1003. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Baik, J.E.; Lagana, S.M.; Han, R.P.; Raab, W.J.; Sahoo, D.; Dalerba, P.; Wang, T.C.; Han, Y.W. Fusobacterium Nucleatum Promotes Colorectal Cancer by Inducing Wnt/β-Catenin Modulator Annexin A1. EMBO Rep. 2019, 20, e47638. [Google Scholar] [CrossRef] [PubMed]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium Nucleatum Promotes Colorectal Carcinogenesis by Modulating E-Cadherin/β-Catenin Signaling via Its FadA Adhesin. Cell Host Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Huang, J.; Yu, T.; Fang, X.; Lou, L.; Xin, S.; Ji, L.; Jiang, F.; Lou, Y. Fusobacterium Nucleatum Promotes the Progression of Colorectal Cancer Through Cdk5-Activated Wnt/Beta-Catenin Signaling. Front. Microbiol. 2021, 11, 545251. [Google Scholar] [CrossRef]
- Cavallucci, V.; Palucci, I.; Fidaleo, M.; Mercuri, A.; Masi, L.; Emoli, V.; Bianchetti, G.; Fiori, M.E.; Bachrach, G.; Scaldaferri, F.; et al. Proinflammatory and Cancer-Promoting Pathobiont Fusobacterium Nucleatum Directly Targets Colorectal Cancer Stem Cells. Biomolecules 2022, 12, 1256. [Google Scholar] [CrossRef]
- Akimoto, N.; Ugai, T.; Zhong, R.; Hamada, T.; Fujiyoshi, K.; Giannakis, M.; Wu, K.; Cao, Y.; Ng, K.; Ogino, S. Rising Incidence of Early-Onset Colorectal Cancer-a Call to Action. Nat. Rev. Clin. Oncol. 2021, 18, 230–243. [Google Scholar] [CrossRef] [PubMed]
- Dejea, C.M.; Fathi, P.; Craig, J.M.; Boleij, A.; Taddese, R.; Geis, A.L.; Wu, X.; Shields, C.E.D.; Hechenbleikner, E.M.; Huso, D.L.; et al. Patients with Familial Adenomatous Polyposis Harbor Colonic Biofilms Containing Tumorigenic Bacteria. Science 2018, 359, 592–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, M.; Wang, X.; Sun, J.; Lin, W.; Chen, L.; Liu, S.; Wu, X.; Shi, L.; Xu, P.; Cai, X.; et al. IRF3 Prevents Colorectal Tumorigenesis via Inhibiting the Nuclear Translocation of Beta-Catenin. Nat. Commun. 2020, 11, 5762. [Google Scholar] [CrossRef]
- Fantini, M.C.; Guadagni, I. From Inflammation to Colitis-Associated Colorectal Cancer in Inflammatory Bowel Disease: Pathogenesis and Impact of Current Therapies. Dig. Liver Dis. 2021, 53, 558–565. [Google Scholar] [CrossRef]
- Kaplan, G.G. The Global Burden of IBD: From 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 720–727. [Google Scholar] [CrossRef]
- Foersch, S.; Neurath, M.F. Colitis-Associated Neoplasia: Molecular Basis and Clinical Translation. Cell. Mol. Life Sci. 2014, 71, 3523–3535. [Google Scholar] [CrossRef]
- Cooper, H.S.; Everley, L.; Chang, W.C.; Pfeiffer, G.; Lee, B.; Murthy, S.; Clapper, M.L. The Role of Mutant Apc in the Development of Dysplasia and Cancer in the Mouse Model of Dextran Sulfate Sodium-Induced Colitis. Gastroenterology 2001, 121, 1407–1416. [Google Scholar] [CrossRef]
- Nagao-Kitamoto, H.; Kitamoto, S.; Kamada, N. Inflammatory Bowel Disease and Carcinogenesis. Cancer Metastasis Rev. 2022, 41, 301–316. [Google Scholar] [CrossRef] [PubMed]
- Ben-Neriah, Y.; Karin, M. Inflammation Meets Cancer, with NF-Kappa B as the Matchmaker. Nat. Immunol. 2011, 12, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Lasry, A.; Zinger, A.; Ben-Neriah, Y. Inflammatory Networks Underlying Colorectal Cancer. Nat. Immunol. 2016, 17, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Dyson, J.K.; Rutter, M.D. Colorectal Cancer in Inflammatory Bowel Disease: What Is the Real Magnitude of the Risk? World J. Gastroenterol. 2012, 18, 3839–3848. [Google Scholar] [CrossRef]
- Triantafillidis, J.K.; Nasioulas, G.; Kosmidis, P.A. Colorectal Cancer and Inflammatory Bowel Disease: Epidemiology, Risk Factors, Mechanisms of Carcinogenesis and Prevention Strategies. Anticancer Res. 2009, 29, 2727–2737. [Google Scholar]
- Rajamaki, K.; Taira, A.; Katainen, R.; Valimaki, N.; Kuosmanen, A.; Plaketti, R.-M.; Seppala, T.T.; Ahtiainen, M.; Wirta, E.-V.; Vartiainen, E.; et al. Genetic and Epigenetic Characteristics of Inflammatory Bowel Disease-Associated Colorectal Cancer. Gastroenterology 2021, 161, 592–607. [Google Scholar] [CrossRef]
- Tang, F.; Cao, F.; Lu, C.; He, X.; Weng, L.; Sun, L. Dvl2 Facilitates the Coordination of NF-Kappa B and Wnt Signaling to Promote Colitis-Associated Colorectal Progression. Cancer Sci. 2022, 113, 565–575. [Google Scholar] [CrossRef]
- Zheng, L.; Duan, S.-L.; Wen, X.-L.; Dai, Y.-C. Molecular Regulation after Mucosal Injury and Regeneration in Ulcerative Colitis. Front. Mol. Biosci. 2022, 9, 1015049. [Google Scholar] [CrossRef]
- Guenther, C.; Neumann, H.; Neurath, M.F.; Becker, C. Apoptosis, Necrosis and Necroptosis: Cell Death Regulation in the Intestinal Epithelium. Gut 2013, 62, 1062–1071. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Cui, D.; Wu, Q.-N.; Zhao, Q.; Chen, Z.-H.; Li, L.; Birchmeier, W.; Yu, Y.; Tao, R. Targeting Wnt/β-Catenin Signaling by TET1/FOXO4 Inhibits Metastatic Spreading and Self-Renewal of Cancer Stem Cells in Gastric Cancer. Cancers 2022, 14, 3232. [Google Scholar] [CrossRef]
- Sahebdel, F.; Parvaneh Tafreshi, A.; Arefian, E.; Roussa, E.; Nadri, S.; Zeynali, B. Wnt/β-Catenin Signaling Pathway Is Involved in Early Dopaminergic Differentiation of Trabecular Meshwork-Derived Mesenchymal Stem Cells. J. Cell. Biochem. 2022, 123, 1120–1129. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Dong, H.; Wang, P.; Xu, Z.; Xian, J.; Chen, J.; Wu, H.; Lou, Y.; Lin, D.; Zhong, B. IL-36 γ and IL-36Ra Reciprocally Regulate Colon Inflammation and Tumorigenesis by Modulating the Cell–Matrix Adhesion Network and Wnt Signaling. Adv. Sci. 2022, 9, 2103035. [Google Scholar] [CrossRef] [PubMed]
- Boege, Y.; Malehmir, M.; Healy, M.E.; Bettermann, K.; Lorentzen, A.; Vucur, M.; Ahuja, A.K.; Bohm, F.; Mertens, J.C.; Shimizu, Y.; et al. A Dual Role of Caspase-8 in Triggering and Sensing Proliferation-Associated DNA Damage, a Key Determinant of Liver Cancer Development. Cancer Cell 2017, 32, 342–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Healy, M.E.; Boege, Y.; Hodder, M.C.; Bohm, F.; Malehmir, M.; Scherr, A.-L.; Jetzer, J.; Chan, L.K.; Parrotta, R.; Jacobs, K.; et al. MCL1 Is Required for Maintenance of Intestinal Homeostasis and Prevention of Carcinogenesis in Mice. Gastroenterology 2020, 159, 183–199. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Komai, K.; Mise-Omata, S.; Iizuka-Koga, M.; Noguchi, Y.; Kondo, T.; Sakai, R.; Matsuo, K.; Nakayama, T.; Yoshie, O.; et al. Brain Regulatory T Cells Suppress Astrogliosis and Potentiate Neurological Recovery. Nature 2019, 565, 246–250. [Google Scholar] [CrossRef] [PubMed]
- Sefik, E.; Geva-Zatorsky, N.; Oh, S.; Konnikova, L.; Zemmour, D.; McGuire, A.M.; Burzyn, D.; Ortiz-Lopez, A.; Lobera, M.; Yang, J.; et al. Individual Intestinal Symbionts Induce a Distinct Population of RORγ + Regulatory T Cells. Science 2015, 349, 993–997. [Google Scholar] [CrossRef] [Green Version]
- van Loosdregt, J.; Fleskens, V.; Tiemessen, M.M.; Mokry, M.; van Boxtel, R.; Meerding, J.; Pals, C.E.G.M.; Kurek, D.; Baert, M.R.M.; Delemarre, E.M.; et al. Canonical Wnt Signaling Negatively Modulates Regulatory T Cell Function. Immunity 2013, 39, 298–310. [Google Scholar] [CrossRef] [Green Version]
- Samstein, R.M.; Arvey, A.; Josefowicz, S.Z.; Peng, X.; Reynolds, A.; Sandstrom, R.; Neph, S.; Sabo, P.; Kim, J.M.; Liao, W.; et al. Foxp3 Exploits a Pre-Existent Enhancer Landscape for Regulatory T Cell Lineage Specification. Cell 2012, 151, 153–166. [Google Scholar] [CrossRef] [Green Version]
- Xing, S.; Gai, K.; Li, X.; Shao, P.; Zeng, Z.; Zhao, X.; Zhao, X.; Chen, X.; Paradee, W.J.; Meyerholz, D.K.; et al. Tcf1 and Lef1 Are Required for the Immunosuppressive Function of Regulatory T Cells. J. Exp. Med. 2019, 216, 847–866. [Google Scholar] [CrossRef]
- Quandt, J.; Arnovitz, S.; Haghi, L.; Woehlk, J.; Mohsin, A.; Okoreeh, M.; Mathur, P.S.; Emmanuel, A.O.; Osman, A.; Krishnan, M.; et al. Wnt–β-Catenin Activation Epigenetically Reprograms Treg Cells in Inflammatory Bowel Disease and Dysplastic Progression. Nat. Immunol. 2021, 22, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, R.; van Driel, M.S.; Vermeulen, L.; van Neerven, S.M. Intestinal Stem Cell Dynamics in Homeostasis and Cancer. Trends Cancer 2022, 8, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Pashirzad, M.; Sathyapalan, T.; Sheikh, A.; Kesharwani, P.; Sahebkar, A. Cancer Stem Cells: An Overview of the Pathophysiological and Prognostic Roles in Colorectal Cancer. Process Biochem. 2022, 115, 19–29. [Google Scholar] [CrossRef]
- Batlle, E.; Clevers, H. Cancer Stem Cells Revisited. Nat. Med. 2017, 23, 1124–1134. [Google Scholar] [CrossRef] [PubMed]
- Ricci-Vitiani, L.; Fabrizi, E.; Palio, E.; De Maria, R. Colon Cancer Stem Cells. J. Mol. Med. 2009, 87, 1097. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.R.; Santos, L.d.S.; Dias, R.B.; Quadros, C.A.; Bezerra, D.P. Emerging Agents That Target Signaling Pathways to Eradicate Colorectal Cancer Stem Cells. Cancer Commun. 2021, 41, 1275–1313. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Katoh, M. WNT Signaling and Cancer Stemness. Essays Biochem. 2022, 66, 319–331. [Google Scholar] [CrossRef]
- Sato, T.; van Es, J.H.; Snippert, H.J.; Stange, D.E.; Vries, R.G.; van den Born, M.; Barker, N.; Shroyer, N.F.; van de Wetering, M.; Clevers, H. Paneth Cells Constitute the Niche for Lgr5 Stem Cells in Intestinal Crypts. Nature 2011, 469, 415–418. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, N.; Kraiczy, J.; Shivdasani, R.A. Cellular and Molecular Architecture of the Intestinal Stem Cell Niche. Nat. Cell Biol. 2020, 22, 1033–1041. [Google Scholar] [CrossRef]
- van Neerven, S.M.; Vermeulen, L. The Interplay between Intrinsic and Extrinsic Wnt Signaling in Controlling Intestinal Transformation. Differentiation 2019, 108, 17–23. [Google Scholar] [CrossRef]
- Nusse, R.; Clevers, H. Wnt/β-Catenin Signaling, Disease, and Emerging Therapeutic Modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, P.; Xiol, J.; Dill, M.T.; Yuan, W.-C.; Panero, R.; Roper, J.; Osorio, F.G.; Maglic, D.; Li, Q.; Gurung, B.; et al. Regenerative Reprogramming of the Intestinal Stem Cell State via Hippo Signaling Suppresses Metastatic Colorectal Cancer. Cell Stem Cell 2020, 27, 590–604.e9. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Duronio, G.N.; Yang, Y.; Bala, P.; Hebbar, P.; Spisak, S.; Sahgal, P.; Singh, H.; Zhang, Y.; Xie, Y.; et al. An Enhancer-Driven Stem Cell-Like Program Mediated by SOX9 Blocks Intestinal Differentiation in Colorectal Cancer. Gastroenterology 2022, 162, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Tomic, G.; Morrissey, E.; Kozar, S.; Ben-Moshe, S.; Hoyle, A.; Azzarelli, R.; Kemp, R.; Chilamakuri, C.S.R.; Itzkovitz, S.; Philpott, A.; et al. Phospho-Regulation of ATOH1 Is Required for Plasticity of Secretory Progenitors and Tissue Regeneration. Cell Stem Cell 2018, 23, 436–443.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sousa e Melo, F.; Kurtova, A.V.; Harnoss, J.M.; Kljavin, N.; Hung, J.; Anderson, J.E.; Storm, E.E.; Modrusan, Z.; Koeppen, H.; Dijkgraaf, G.J.P.; et al. A Distinct Role for Lgr5(+) Stem Cells in Primary and Metastatic Colon Cancer. Nature 2017, 543, 676–680. [Google Scholar] [CrossRef]
- Sansom, O.J.; Meniel, V.S.; Muncan, V.; Phesse, T.J.; Wilkins, J.A.; Reed, K.R.; Vass, J.K.; Athineos, D.; Clevers, H.; Clarke, A.R. Myc Deletion Rescues Apc Deficiency in the Small Intestine. Nature 2007, 446, 676–679. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Lin, M.; Dong, C.; Tang, Y.; An, L.; Ju, J.; Wen, F.; Chen, F.; Wang, M.; Wang, W.; et al. An MST4-p Beta-Catenin(Thr40) Signaling Axis Controls Intestinal Stem Cell and Tumorigenesis. Adv. Sci. 2021, 8, 2004850. [Google Scholar] [CrossRef]
- Angus-Hill, M.L.; Elbert, K.M.; Hidalgo, J.; Capecchi, M.R. T-Cell Factor 4 Functions as a Tumor Suppressor Whose Disruption Modulates Colon Cell Proliferation and Tumorigenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 4914–4919. [Google Scholar] [CrossRef] [Green Version]
- Hrckulak, D.; Janeckova, L.; Lanikova, L.; Kriz, V.; Horazna, M.; Babosova, O.; Vojtechova, M.; Galuskova, K.; Sloncova, E.; Korinek, V. Wnt Effector TCF4 Is Dispensable for Wnt Signaling in Human Cancer Cells. Genes 2018, 9, 439. [Google Scholar] [CrossRef] [Green Version]
- Novoa Diaz, M.B.; Martin, M.J.; Gentili, C. Tumor Microenvironment Involvement in Colorectal Cancer Progression via Wnt/Beta-Catenin Pathway: Providing Understanding of the Complex Mechanisms of Chemoresistance. World J. Gastroenterol. 2022, 28, 3027–3046. [Google Scholar] [CrossRef]
- Kalra, H.; Gangoda, L.; Fonseka, P.; Chitti, S.V.; Liem, M.; Keerthikumar, S.; Samuel, M.; Boukouris, S.; Al Saffar, H.; Collins, C.; et al. Extracellular Vesicles Containing Oncogenic Mutant β-Catenin Activate Wnt Signalling Pathway in the Recipient Cells. J. Extracell. Vesicles 2019, 8, 1690217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, H.-J.; Jang, G.-B.; Lee, H.-Y.; Park, S.-R.; Kim, J.-Y.; Nam, J.-S.; Hong, I.-S. The Wnt/β-Catenin Signaling/Id2 Cascade Mediates the Effects of Hypoxia on the Hierarchy of Colorectal-Cancer Stem Cells. Sci. Rep. 2016, 6, 22966. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Yang, M.; Li, Y.; Yang, F.; Feng, Y. Exosomes Derived from Hypoxic Colorectal Cancer Cells Transfer Wnt4 to Normoxic Cells to Elicit a Prometastatic Phenotype. Int. J. Biol. Sci. 2018, 14, 2094–2102. [Google Scholar] [CrossRef]
- Cheng, R.; Sun, B.; Liu, Z.; Zhao, X.; Qi, L.; Li, Y.; Gu, Q. Wnt5a Suppresses Colon Cancer by Inhibiting Cell Proliferation and Epithelial-Mesenchymal Transition. J. Cell. Physiol. 2014, 229, 1908–1917. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, C.; Wang, S.; Shi, D.; Wei, C.; Song, J.; Lin, X.; Dou, R.; Bai, J.; Xiang, Z.; et al. Wnt5a-Induced M2 Polarization of Tumor-Associated Macrophages via IL-10 Promotes Colorectal Cancer Progression. Cell Commun. Signal. 2020, 18, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Q.; Song, J.; Pan, Y.; Shi, D.; Yang, C.; Wang, S.; Xiong, B. Wnt5a/CaMKII/ERK/CCL2 Axis Is Required for Tumor-Associated Macrophages to Promote Colorectal Cancer Progression. Int. J. Biol. Sci. 2020, 16, 1023–1034. [Google Scholar] [CrossRef]
- Kasprzak, A. Angiogenesis-Related Functions of Wnt Signaling in Colorectal Carcinogenesis. Cancers 2020, 12, 3601. [Google Scholar] [CrossRef]
- Baeriswyl, V.; Christofori, G. The Angiogenic Switch in Carcinogenesis. Semin. Cancer Biol. 2009, 19, 329–337. [Google Scholar] [CrossRef]
- Hillen, F.; Griffioen, A.W. Tumour Vascularization: Sprouting Angiogenesis and Beyond. Cancer Metastasis Rev. 2007, 26, 489–502. [Google Scholar] [CrossRef] [Green Version]
- Watnick, R.S. The Role of the Tumor Microenvironment in Regulating Angiogenesis. Cold Spring Harb. Perspect. Med. 2012, 2, a006676. [Google Scholar] [CrossRef]
- Kramer, N.; Schmöllerl, J.; Unger, C.; Nivarthi, H.; Rudisch, A.; Unterleuthner, D.; Scherzer, M.; Riedl, A.; Artaker, M.; Crncec, I.; et al. Autocrine WNT2 Signaling in Fibroblasts Promotes Colorectal Cancer Progression. Oncogene 2017, 36, 5460–5472. [Google Scholar] [CrossRef] [PubMed]
- Unterleuthner, D.; Neuhold, P.; Schwarz, K.; Janker, L.; Neuditschko, B.; Nivarthi, H.; Crncec, I.; Kramer, N.; Unger, C.; Hengstschlaeger, M.; et al. Cancer-Associated Fibroblast-Derived WNT2 Increases Tumor Angiogenesis in Colon Cancer. Angiogenesis 2020, 23, 159–177. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Shu, H.; Zhang, L.; Xiong, J. Long Noncoding RNA GAS5 Inhibits Angiogenesis and Metastasis of Colorectal Cancer through the Wnt/Β-catenin Signaling Pathway. J. Cell. Biochem. 2019, 120, 6937–6951. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Zhao, H.; Qu, B.; Chu, X.; Xin, X.; Zhang, Q.; Li, W.; Yang, S. TRIM24 Promotes Colorectal Cancer Cell Progression via the Wnt/Beta-Catenin Signaling Pathway Activation. Am. J. Transl. Res. 2022, 14, 831–848. [Google Scholar] [PubMed]
- Yang, D.; Li, Q.; Shang, R.; Yao, L.; Wu, L.; Zhang, M.; Zhang, L.; Xu, M.; Lu, Z.; Zhou, J.; et al. WNT4 Secreted by Tumor Tissues Promotes Tumor Progression in Colorectal Cancer by Activation of the Wnt/Beta-Catenin Signalling Pathway. J. Exp. Clin. Cancer Res. 2020, 39, 251. [Google Scholar] [CrossRef]
- EL Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular Vesicles: Biology and Emerging Therapeutic Opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Yamada, N.; Kuranaga, Y.; Kumazaki, M.; Shinohara, H.; Taniguchi, K.; Akao, Y. Colorectal Cancer Cell-Derived Extracellular Vesicles Induce Phenotypic Alteration of T Cells into Tumor-Growth Supporting Cells with Transforming Growth Factor-Β1-Mediated Suppression. Oncotarget 2016, 7, 27033–27043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, B.S.; Cho, J.-H.; Kim, H.; Choi, E.-J.; Rho, S.; Kim, J.; Kim, J.H.; Choi, D.-S.; Kim, Y.-K.; Hwang, D.; et al. Colorectal Cancer Cell-Derived Microvesicles Are Enriched in Cell Cycle-Related MRNAs That Promote Proliferation of Endothelial Cells. BMC Genom. 2009, 10, 556. [Google Scholar] [CrossRef] [Green Version]
- Ji, H.; Greening, D.W.; Barnes, T.W.; Lim, J.W.; Tauro, B.J.; Rai, A.; Xu, R.; Adda, C.; Mathivanan, S.; Zhao, W.; et al. Proteome Profiling of Exosomes Derived from Human Primary and Metastatic Colorectal Cancer Cells Reveal Differential Expression of Key Metastatic Factors and Signal Transduction Components. Proteomics 2013, 13, 1672–1686. [Google Scholar] [CrossRef]
- Huang, Z.; Feng, Y. Exosomes Derived from Hypoxic Colorectal Cancer Cells Promote Angiogenesis Through Wnt4-Induced β-Catenin Signaling in Endothelial Cells. Oncol. Res. 2017, 25, 651–661. [Google Scholar] [CrossRef]
- Winkler, J.; Abisoye-Ogunniyan, A.; Metcalf, K.J.; Werb, Z. Concepts of Extracellular Matrix Remodelling in Tumour Progression and Metastasis. Nat. Commun. 2020, 11, 5120. [Google Scholar] [CrossRef] [PubMed]
- Vu, T.; Datta, P. Regulation of EMT in Colorectal Cancer: A Culprit in Metastasis. Cancers 2017, 9, 171. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Lai, Q.; He, C.; Fang, Y.; Yan, Q.; Zhang, Y.; Wang, X.; Gu, C.; Wang, Y.; Ye, L.; et al. RUNX1 Promotes Tumour Metastasis by Activating the Wnt/β-Catenin Signalling Pathway and EMT in Colorectal Cancer. J. Exp. Clin. Cancer Res. 2019, 38, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCrea, P.D.; Gottardi, C.J. Beyond β-Catenin: Prospects for a Larger Catenin Network in the Nucleus. Nat. Rev. Mol. Cell Biol. 2016, 17, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Basu, S.; Cheriyamundath, S.; Ben-Ze’ev, A. Cell–Cell Adhesion: Linking Wnt/β-Catenin Signaling with Partial EMT and Stemness Traits in Tumorigenesis. F1000Research 2018, 7, 1488. [Google Scholar] [CrossRef] [Green Version]
- Valenta, T.; Hausmann, G.; Basler, K. The Many Faces and Functions of β-Catenin. EMBO J. 2012, 31, 2714–2736. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Gan, Y.; Zou, R.; Sha, H.; Lu, Y.; Zhang, Y.; Feng, J. RNF128 Suppresses the Malignancy of Colorectal Cancer Cells via Inhibition of Wnt/Beta-Catenin Signaling. Am. J. Transl. Res. 2021, 13, 13567–13578. [Google Scholar]
- Meng, F.; Liu, X.; Lin, C.; Xu, L.; Liu, J.; Zhang, P.; Zhang, X.; Song, J.; Yan, Y.; Ren, Z.; et al. SMYD2 Suppresses APC2 Expression to Activate the Wnt/Beta-Catenin Pathway and Promotes Epithelial-Mesenchymal Transition in Colorectal Cancer. Am. J. Cancer Res. 2020, 10, 997–1011. [Google Scholar]
- Tang, Q.; Chen, J.; Di, Z.; Yuan, W.; Zhou, Z.; Liu, Z.; Han, S.; Liu, Y.; Ying, G.; Shu, X.; et al. TM4SF1 Promotes EMT and Cancer Stemness via the Wnt/Beta-Catenin/SOX2 Pathway in Colorectal Cancer. J. Exp. Clin. Cancer Res. 2020, 39, 232. [Google Scholar] [CrossRef]
- Cai, Z.; Mei, Y.; Jiang, X.; Shi, X. WDR74 Promotes Proliferation and Metastasis in Colorectal Cancer Cells through Regulating the Wnt/Beta-Catenin Signaling Pathway. Open Life Sci. 2021, 16, 920–929. [Google Scholar] [CrossRef]
- Zhang, L.; Ren, C.; Yang, Z.; Gong, L.; Wang, C.; Feng, M.; Guan, W. Forkhead Box S1 Mediates Epithelial-Mesenchymal Transition through the Wnt/Beta-Catenin Signaling Pathway to Regulate Colorectal Cancer Progression. J. Transl. Med. 2022, 20, 327. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, L.; Xu, X.; Han, P.; Wu, J.; Tian, X.; Li, M. FOXO4 Inhibits the Migration and Metastasis of Colorectal Cancer by Regulating the APC2/Beta-Catenin Axis. Front. Cell Dev. Biol. 2021, 9, 659731. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Gao, T.; Xie, J.; Huang, Z.; Zhang, X.; Yang, F.; Qi, W.; Yang, Z.; Zhou, T.; Gao, G.; et al. FUBP1 Promotes Colorectal Cancer Stemness and Metastasis via DVL1-Mediated Activation of Wnt/Beta-Catenin Signaling. Mol. Oncol. 2021, 15, 3490–3512. [Google Scholar] [CrossRef]
- Faubert, B.; Solmonson, A.; DeBerardinis, R.J. Metabolic Reprogramming and Cancer Progression. Science 2020, 368, eaaw5473. [Google Scholar] [CrossRef]
- Martinez-Reyes, I.; Chandel, N.S. Cancer Metabolism: Looking Forward. Nat. Rev. Cancer 2021, 21, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, K.; Oi, R.; Okuzaki, D.; Motooka, D.; Shinohara, M.; Nojima, S.; Morii, E. Mitochondrial Matrix Protein C14orf159 Attenuates Colorectal Cancer Metastasis by Suppressing Wnt/Beta-Catenin Signalling. Br. J. Cancer 2021, 125, 1699–1711. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, K.; Nojima, S.; Tahara, S.; Kurashige, M.; Kawasaki, K.; Hori, Y.; Taniguchi, M.; Umakoshi, Y.; Okuzaki, D.; Wada, N.; et al. Serine Racemase Enhances Growth of Colorectal Cancer by Producing Pyruvate from Serine. Nat. Metab. 2020, 2, 81–96. [Google Scholar] [CrossRef]
- Zhu, Y.-M.; Li, Q.; Gao, X.-Z.; Meng, X.; Sun, L.-L.; Shi, Y.; Lu, E.-T.; Zhang, Y. C14orf159 Suppresses Gastric Cancer Cells’ Invasion and Proliferation by Inactivating ERK Signaling. Cancer Manag. Res. 2019, 11, 1717–1723. [Google Scholar] [CrossRef] [Green Version]
- Bakir, B.; Chiarella, A.M.; Pitarresi, J.R.; Rustgi, A.K. EMT, MET, Plasticity, and Tumor Metastasis. Trends Cell Biol. 2020, 30, 764–776. [Google Scholar] [CrossRef]
- Bimonte, S.; Barbieri, A.; Leongito, M.; Piccirillo, M.; Giudice, A.; Pivonello, C.; De Angelis, C.; Granata, V.; Palaia, R.; Izzo, F. Curcumin AntiCancer Studies in Pancreatic Cancer. Nutrients 2016, 8, 433. [Google Scholar] [CrossRef] [Green Version]
- Dongre, A.; Weinberg, R.A. New Insights into the Mechanisms of Epithelial-Mesenchymal Transition and Implications for Cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Li, Q.; Liu, Y.; Zhang, H.; Wang, Q.; Chen, Y.; Shi, X.; Li, J.; Zhang, H.; Zhang, Y.; et al. Activation of WNT7b Autocrine Eases Metastasis of Colorectal Cancer via Epithelial to Mesenchymal Transition and Predicts Poor Prognosis. BMC Cancer 2021, 21, 180. [Google Scholar] [CrossRef] [PubMed]
- Pergolizzi, M.; Bizzozero, L.; Maione, F.; Maldi, E.; Isella, C.; Macagno, M.; Mariella, E.; Bardelli, A.; Medico, E.; Marchio, C.; et al. The Neuronal Protein Neuroligin 1 Promotes Colorectal Cancer Progression by Modulating the APC/Beta-Catenin Pathway. J. Exp. Clin. Cancer Res. 2022, 41, 266. [Google Scholar] [CrossRef]
- Xu, C.; Ding, Y.; Wang, K.; Hao, M.; Li, H.; Ding, L. Claudin-7 Deficiency Promotes Stemness Properties in Colorectal Cancer through Sox9-Mediated Wnt/Beta-Catenin Signalling. J. Transl. Med. 2021, 19, 311. [Google Scholar] [CrossRef]
- Nie, X.; Liu, H.; Ye, W.; Wei, X.; Fan, L.; Ma, H.; Li, L.; Xue, W.; Qi, W.; Wang, Y.-D.; et al. LRP5 Promotes Cancer Stem Cell Traits and Chemoresistance in Colorectal Cancer. J. Cell. Mol. Med. 2022, 26, 1095–1112. [Google Scholar] [CrossRef] [PubMed]
- VanderVorst, K.; Dreyer, C.A.; Konopelski, S.E.; Lee, H.; Ho, H.-Y.H.; Carraway, K.L. Wnt/PCP Signaling Contribution to Carcinoma Collective Cell Migration and Metastasis. Cancer Res. 2019, 79, 1719–1729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, J.C.; Chaudhary, V.; Bartscherer, K.; Boutros, M. Active Wnt Proteins Are Secreted on Exosomes. Nat. Cell Biol. 2012, 14, 1036–1045. [Google Scholar] [CrossRef]
- Wang, F.-W.; Cao, C.-H.; Han, K.; Zhao, Y.-X.; Cai, M.-Y.; Xiang, Z.-C.; Zhang, J.-X.; Chen, J.-W.; Zhong, L.-P.; Huang, Y.; et al. APC-Activated Long Noncoding RNA Inhibits Colorectal Carcinoma Pathogenesis through Reduction of Exosome Production. J. Clin. Investig. 2019, 129, 727–743. [Google Scholar] [CrossRef] [Green Version]
- Le Floch, N.; Rivat, C.; De Wever, O.; Bruyneel, E.; Mareel, M.; Dale, T.; Gespach, C. The Proinvasive Activity of Wnt-2 Is Mediated through a Noncanonical Wnt Pathway Coupled to GSK-3β and C- Jun/AP-1 Signaling. FASEB J. 2005, 19, 144–146. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, L.; Jin, Y.; Lin, Y.; Cao, Y.; Zheng, C. Overexpression of WNT5B Promotes COLO 205 Cell Migration and Invasion through the JNK Signaling Pathway. Oncol. Rep. 2016, 36, 23–30. [Google Scholar] [CrossRef] [Green Version]
- Ouko, L.; Ziegler, T.R.; Gu, L.H.; Eisenberg, L.M.; Yang, V.M. Wnt11 Signaling Promotes Proliferation, Transformation, and Migration of IEC6 Intestinal Epithelial Cells. J. Biol. Chem. 2004, 279, 26707–26715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, K.; Hazama, S.; Mitomori, S.; Nishioka, M.; Suehiro, Y.; Hirata, H.; Oka, M.; Imai, K.; Dahiya, R.; Hinoda, Y. Down-Regulation of Frizzled-7 Expression Decreases Survival, Invasion and Metastatic Capabilities of Colon Cancer Cells. Br. J. Cancer 2009, 101, 1374–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, K.; Hiura, M.; Suehiro, Y.; Hazama, S.; Hirata, H.; Oka, M.; Imai, K.; Dahiya, R.; Hinoda, Y. Frizzled-7 as a Potential Therapeutic Target in Colorectal Cancer. Neoplasia 2008, 10, 697–705. [Google Scholar] [CrossRef] [Green Version]
- Nishioka, M.; Ueno, K.; Hazama, S.; Okada, T.; Sakai, K.; Suehiro, Y.; Okayama, N.; Hirata, H.; Oka, M.; Imai, K.; et al. Possible Involvement of Wnt11 in Colorectal Cancer Progression. Mol. Carcinog. 2013, 52, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Parsons, S.; Maldonado, E.B.; Prasad, V. Comparison of Drugs Used for Adjuvant and Metastatic Therapy of Colon, Breast, and Non-Small Cell Lung Cancers. JAMA Netw. Open 2020, 3, e202488. [Google Scholar] [CrossRef] [Green Version]
- Hirano, H.; Takashima, A.; Hamaguchi, T.; Shida, D.; Kanemitsu, Y. Current Status and Perspectives of Immune Checkpoint Inhibitors for Colorectal Cancer. Jpn. J. Clin. Oncol. 2021, 51, 10–19. [Google Scholar] [CrossRef]
- Zhu, G.-X.; Gao, D.; Shao, Z.-Z.; Chen, L.; Ding, W.-J.; Yu, Q.-F. Wnt/Beta-Catenin Signaling: Causes and Treatment Targets of Drug Resistance in Colorectal Cancer. Mol. Med. Rep. 2021, 23, 105. [Google Scholar] [CrossRef]
- Das, P.K.; Islam, F.; Lam, A.K. The Roles of Cancer Stem Cells and Therapy Resistance in Colorectal Carcinoma. Cells 2020, 9, 1392. [Google Scholar] [CrossRef]
- Yeoh, Y.; Low, T.Y.; Abu, N.; Lee, P.Y. Regulation of Signal Transduction Pathways in Colorectal Cancer: Implications for Therapeutic Resistance. PeerJ 2021, 9, e12338. [Google Scholar] [CrossRef]
- Liu, Y.-S.; Hsu, H.-C.; Tseng, K.-C.; Chen, H.-C.; Chen, S.-J. Lgr5 Promotes Cancer Stemness and Confers Chemoresistance through ABCB1 in Colorectal Cancer. Biomed. Pharmacother. 2013, 67, 791–799. [Google Scholar] [CrossRef]
- Liu, H.; Pan, Y.; Guo, G.; Hu, X.; Huang, K.; Zhang, X. Isolation of Side Population Cells and Detection of ABCG2 from SW480. Chin. J. Cancer Res. 2007, 19, 238–243. [Google Scholar] [CrossRef]
- Park, S.-Y.; Kim, J.-Y.; Jang, G.-B.; Choi, J.-H.; Kim, J.-H.; Lee, C.-J.; Lee, S.; Baek, J.-H.; Park, K.-K.; Kim, J.-M.; et al. Aberrant Activation of the CD45-Wnt Signaling Axis Promotes Stemness and Therapy Resistance in Colorectal Cancer Cells. Theranostics 2021, 11, 8755–8770. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Jiang, H.; Li, W.; Chen, L.; Zhu, W.; Xian, Y.; Han, Z.; Yin, L.; Liu, Y.; Wang, Y.; et al. FOXM1/DVL2/Snail Axis Drives Metastasis and Chemoresistance of Colorectal Cancer. Aging 2020, 12, 24424–24440. [Google Scholar] [CrossRef]
- Zhang, W.; Kong, L.; Zhu, H.; Sun, D.; Han, Q.; Yan, B.; Cui, Z.; Zhang, W.; Zhang, S.; Kang, X.; et al. Retinoic Acid-Induced 2 (RAI2) Is a Novel Antagonist of Wnt/Beta-Catenin Signaling Pathway and Potential Biomarker of Chemosensitivity in Colorectal Cancer. Front. Oncol. 2022, 12, 805290. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Yamada-Okabe, H.; Suzuki, M.; Natori, O.; Kato, A.; Matsubara, K.; Chen, Y.J.; Yamazaki, M.; Funahashi, S.; Yoshida, K.; et al. LGR5-Positive Colon Cancer Stem Cells Interconvert with Drug-Resistant LGR5-Negative Cells and Are Capable of Tumor Reconstitution. Stem Cells 2012, 30, 2631–2644. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Settleman, J. EMT, Cancer Stem Cells and Drug Resistance: An Emerging Axis of Evil in the War on Cancer. Oncogene 2010, 29, 4741–4751. [Google Scholar] [CrossRef] [Green Version]
- Zinzi, L.; Contino, M.; Cantore, M.; Capparelli, E.; Leopoldo, M.; Colabufo, N.A. ABC Transporters in CSCs Membranes as a Novel Target for Treating Tumor Relapse. Front. Pharmacol. 2014, 5, 163. [Google Scholar] [CrossRef]
- Kugimiya, N.; Nishimoto, A.; Hosoyama, T.; Ueno, K.; Enoki, T.; Li, T.-S.; Hamano, K. The C-MYC-ABCB5 Axis Plays a Pivotal Role in 5-Fluorouracilresistance in Human Colon Cancer Cells. J. Cell. Mol. Med. 2015, 19, 1569–1581. [Google Scholar] [CrossRef] [Green Version]
- Chikazawa, N.; Tanaka, H.; Tasaka, T.; Nakamura, M.; Tanaka, M.; Onishi, H.; Katano, M. Inhibition of Wnt Signaling Pathway Decreases Chemotherapy-Resistant Side-Population Colon Cancer Cells. Anticancer Res. 2010, 30, 2041–2048. [Google Scholar]
- Liang, Y.; Wang, B.; Chen, S.; Ye, Z.; Chai, X.; Li, R.; Li, X.; Kong, G.; Li, Y.; Zhang, X.; et al. Beta-1 Syntrophin (SNTB1) Regulates Colorectal Cancer Progression and Stemness via Regulation of the Wnt/Beta-Catenin Signaling Pathway. Ann. Transl. Med. 2021, 9, 1016. [Google Scholar] [CrossRef]
- Catalano, V.; Turdo, A.; Di Franco, S.; Dieli, F.; Todaro, M.; Stassi, G. Tumor and Its Microenvironment: A Synergistic Interplay. Semin. Cancer Biol. 2013, 23, 522–532. [Google Scholar] [CrossRef]
- Sleeman, J.P. The Metastatic Niche and Stromal Progression. Cancer Metastasis Rev. 2012, 31, 429–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mierke, C.T. The Matrix Environmental and Cell Mechanical Properties Regulate Cell Migration and Contribute to the Invasive Phenotype of Cancer Cells. Rep. Prog. Phys. 2019, 82, 064602. [Google Scholar] [CrossRef] [PubMed]
- Czekay, R.-P.; Cheon, D.-J.; Samarakoon, R.; Kutz, S.M.; Higgins, P.J. Cancer-Associated Fibroblasts: Mechanisms of Tumor Progression and Novel Therapeutic Targets. Cancers 2022, 14, 1231. [Google Scholar] [CrossRef]
- Hu, Y.-B.; Yan, C.; Mu, L.; Mil, Y.-L.; Zhao, H.; Hu, H.; Li, X.-L.; Tao, D.-D.; Wu, Y.-Q.; Gong, J.-P.; et al. Exosomal Wnt-Induced Dedifferentiation of Colorectal Cancer Cells Contributes to Chemotherapy Resistance. Oncogene 2019, 38, 1951–1965. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Campisi, J.; Higano, C.; Beer, T.M.; Porter, P.; Coleman, I.; True, L.; Nelson, P.S. Treatment-Induced Damage to the Tumor Microenvironment Promotes Prostate Cancer Therapy Resistance through WNT16B. Nat. Med. 2012, 18, 1359–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Zhu, D.; Chen, F.; Qian, M.; Wei, H.; Chen, W.; Xu, J. SFRP2 Augments WNT16B Signaling to Promote Therapeutic Resistance in the Damaged Tumor Microenvironment. Oncogene 2016, 35, 4321–4334. [Google Scholar] [CrossRef]
- Takada, K.; Zhu, D.; Bird, G.H.; Sukhdeo, K.; Zhao, J.-J.; Mani, M.; Lemieux, M.; Carrasco, D.E.; Ryan, J.; Horst, D.; et al. Targeted Disruption of the BCL9/Beta-Catenin Complex Inhibits Oncogenic Wnt Signaling. Sci. Transl. Med. 2012, 4, 148ra117. [Google Scholar] [CrossRef] [Green Version]
- Dong, S.; Liang, S.; Cheng, Z.; Zhang, X.; Luo, L.; Li, L.; Zhang, W.; Li, S.; Xu, Q.; Zhong, M.; et al. ROS/PI3K/Akt and Wnt/Beta-Catenin Signalings Activate HIF-1 Alpha-Induced Metabolic Reprogramming to Impart 5-Fluorouracil Resistance in Colorectal Cancer. J. Exp. Clin. Cancer Res. 2022, 41, 15. [Google Scholar] [CrossRef]
- Liu, X.; Su, K.; Sun, X.; Jiang, Y.; Wang, L.; Hu, C.; Zhang, C.; Lu, M.; Du, X.; Xing, B. Sec62 Promotes Stemness and Chemoresistance of Human Colorectal Cancer through Activating Wnt/Beta-Catenin Pathway. J. Exp. Clin. Cancer Res. 2021, 40, 132. [Google Scholar] [CrossRef]
- Li, H.; Chen, J.; Liu, J.; Lai, Y.; Huang, S.; Zheng, L.; Fan, N. CPT2 Downregulation Triggers Stemness and Oxaliplatin Resistance in Colorectal Cancer via the ROS/Wnt/-Beta Catenin-Induced Glycolytic Metabolism. Exp. Cell Res. 2021, 409, 112892. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, C.; Wang, G.; Chen, J.; Ju, S.; Huang, J.; Wang, X. SNORD1C Maintains Stemness and 5-FU Resistance by Activation of Wnt Signaling Pathway in Colorectal Cancer. Cell Death Discov. 2022, 8, 200. [Google Scholar] [CrossRef] [PubMed]
- Kishore, C.; Bhadra, P. Current Advancements and Future Perspectives of Immunotherapy in Colorectal Cancer Research. Eur. J. Pharmacol. 2021, 893, 173819. [Google Scholar] [CrossRef]
- Yaghoubi, N.; Soltani, A.; Ghazvini, K.; Hassanian, S.M.; Hashemy, S.I. PD-1/PD-L1 Blockade as a Novel Treatment for Colorectal Cancer. Biomed. Pharmacother. 2019, 110, 312–318. [Google Scholar] [CrossRef] [PubMed]
- Payandeh, Z.; Khalili, S.; Somi, M.H.; Mard-Soltani, M.; Baghbanzadeh, A.; Hajiasgharzadeh, K.; Samadi, N.; Baradaran, B. PD-1/PD-L1-Dependent Immune Response in Colorectal Cancer. J. Cell. Physiol. 2020, 235, 5461–5475. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Jin, L.; Chen, P.; Li, D.; Gao, W.; Dong, G. Colorectal Cancer Immunotherapy-Recent Progress and Future Directions. Cancer Lett. 2022, 545, 215816. [Google Scholar] [CrossRef] [PubMed]
- Fountzilas, E.; Kurzrock, R.; Vo, H.H.; Tsimberidou, A.-M. Wedding of Molecular Alterations and Immune Checkpoint Blockade: Genomics as a Matchmaker. J. Natl. Cancer Inst. 2021, 113, 1634–1647. [Google Scholar] [CrossRef]
- Kim, C.W.; Chon, H.J.; Kim, C. Combination Immunotherapies to Overcome Intrinsic Resistance to Checkpoint Blockade in Microsatellite Stable Colorectal Cancer. Cancers 2021, 13, 4906. [Google Scholar] [CrossRef]
- Ganesh, S.; Shui, X.; Craig, K.P.; Park, J.; Wang, W.; Brown, B.D.; Abrams, M.T. RNAi-Mediated β-Catenin Inhibition Promotes T Cell Infiltration and Antitumor Activity in Combination with Immune Checkpoint Blockade. Mol. Ther. J. Am. Soc. Gene Ther. 2018, 26, 2567–2579. [Google Scholar] [CrossRef]
- Spranger, S.; Dai, D.; Horton, B.; Gajewski, T.F. Tumor-Residing Batf3 Dendritic Cells Are Required for Effector T Cell Trafficking and Adoptive T Cell Therapy. Cancer Cell 2017, 31, 711–723.e4. [Google Scholar] [CrossRef] [Green Version]
- Berraondo, P.; Ochoa, M.C.; Olivera, I.; Melero, I. Immune Desertic Landscapes in Hepatocellular Carcinoma Shaped by β-Catenin Activation. Cancer Discov. 2019, 9, 1003–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, J.; Yu, X.; Xue, L.; Ge, X.; Zhao, W.; Peng, W. Intrinsic β-Catenin Signaling Suppresses CD8+ T-Cell Infiltration in Colorectal Cancer. Biomed. Pharmacother. Biomed. Pharmacother. 2019, 115, 108921. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, J.A.; Sweis, R.F.; Bao, R.; Luke, J.J. T Cell–Inflamed versus Non-T Cell–Inflamed Tumors: A Conceptual Framework for Cancer Immunotherapy Drug Development and Combination Therapy Selection. Cancer Immunol. Res. 2018, 6, 990–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Tian, T.; Kalland, K.-H.; Ke, X.; Qu, Y. Targeting Wnt/β-Catenin Signaling for Cancer Immunotherapy. Trends Pharmacol. Sci. 2018, 39, 648–658. [Google Scholar] [CrossRef]
- Pai, S.G.; Carneiro, B.A.; Mota, J.M.; Costa, R.; Leite, C.A.; Barroso-Sousa, R.; Kaplan, J.B.; Chae, Y.K.; Giles, F.J. Wnt/Beta-Catenin Pathway: Modulating Anticancer Immune Response. J. Hematol. Oncol. 2017, 10, 1–12. [Google Scholar] [CrossRef]
- Tang, Y.; Nan, N.; Gui, C.; Zhou, X.; Jiang, W.; Zhou, X. Blockage of PD-L1 by FERMT3-Mediated Wnt/Beta-Catenin Signalling Regulates Chemoresistance and Immune Evasion of Colorectal Cancer Cells. Clin. Exp. Pharmacol. Physiol. 2022, 49, 988–997. [Google Scholar] [CrossRef]
- Stanczak, A.; Stec, R.; Bodnar, L.; Olszewski, W.; Cichowicz, M.; Kozlowski, W.; Szczylik, C.; Pietrucha, T.; Wieczorek, M.; Lamparska-Przybysz, M. Prognostic Significance of Wnt-1, β-Catenin and E-Cadherin Expression in Advanced Colorectal Carcinoma. Pathol. Oncol. Res. 2011, 17, 955–963. [Google Scholar] [CrossRef] [Green Version]
- Matly, A.; Quinn, J.A.; McMillan, D.C.; Park, J.H.; Edwards, J. The Relationship between β-Catenin and Patient Survival in Colorectal Cancer Systematic Review and Meta-Analysis. Crit. Rev. Oncol./Hematol. 2021, 163, 103337. [Google Scholar] [CrossRef]
- Kriegl, L.; Horst, D.; Reiche, J.A.; Engel, J.; Kirchner, T.; Jung, A. LEF-1 and TCF4 Expression Correlate Inversely with Survival in Colorectal Cancer. J. Transl. Med. 2010, 8, 123. [Google Scholar] [CrossRef] [Green Version]
- Escobar, D.; Bushara, O.; Sun, L.; Liao, J.; Yang, G.-Y. Clinicopathologic Characteristics of FBXW7-Mutated Colorectal Adenocarcinoma and Association with Aberrant Beta-Catenin Localization. Hum. Pathol. 2022, 119, 51–58. [Google Scholar] [CrossRef]
- Dahlmann, M.; Monks, A.; Harris, E.D.; Kobelt, D.; Osterland, M.; Khaireddine, F.; Herrmann, P.; Kemmner, W.; Burock, S.; Walther, W.; et al. Combination of Wnt/Beta-Catenin Targets S100A4 and DKK1 Improves Prognosis of Human Colorectal Cancer. Cancers 2022, 14, 37. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lv, G.; Xu, B.; Jiang, B. Overexpression of UBE2M through Wnt/Beta-Catenin Signaling Is Associated with Poor Prognosis and Chemotherapy Resistance in Colorectal Cancer. Transl. Cancer Res. 2020, 9, 5614–5625. [Google Scholar] [CrossRef] [PubMed]
- Shu, Y.; He, C.; Xiong, Y.; Lan, J.; Song, R. Paired-like Homeodomain Transcription Factor 2 Strengthens Chemoresistance in Colorectal Cancer by Activating the Wnt/Beta-Catenin Axis. Neoplasma 2021, 68, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Guinney, J.; Dienstmann, R.; Wang, X.; de Reyniès, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The Consensus Molecular Subtypes of Colorectal Cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- Caspi, M.; Wittenstein, A.; Kazelnik, M.; Shor-Nareznoy, Y.; Rosin-Arbesfeld, R. Therapeutic Targeting of the Oncogenic Wnt Signaling Pathway for Treating Colorectal Cancer and Other Colonic Disorders. Adv. Drug Deliv. Rev. 2021, 169, 118–136. [Google Scholar] [CrossRef] [PubMed]
- Lenz, H.; Kahn, M. Safely Targeting Cancer Stem Cells via Selective Catenin Coactivator Antagonism. Cancer Sci. 2014, 105, 1087–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bendell, J.; Eckhardt, G.S.; Hochster, H.S.; Morris, V.K.; Strickler, J.; Kapoun, A.M.; Wang, M.; Xu, L.; McGuire, K.; Dupont, J.; et al. Initial Results from a Phase 1a/b Study of OMP-131R10, a First-in-Class Anti-RSPO3 Antibody, in Advanced Solid Tumors and Previously Treated Metastatic Colorectal Cancer (CRC). Eur. J. Cancer 2016, 69, S29–S30. [Google Scholar] [CrossRef]
Figure 1.
The canonical and non-canonical Wnt signalling pathway.

Figure 2.
The Wnt signaling pathway in occurrence of colorectal cancer. Genetic factors, and environmental factors, such as unhealthy lifestyle, and imbalanced gut microbiota may induce colorectal cancer (CRC) by Wnt signaling pathway. Moreover, aberrant Wnt signaling pathway also play a crucial role colitis-associated carcinoma.
Figure 2.
The Wnt signaling pathway in occurrence of colorectal cancer. Genetic factors, and environmental factors, such as unhealthy lifestyle, and imbalanced gut microbiota may induce colorectal cancer (CRC) by Wnt signaling pathway. Moreover, aberrant Wnt signaling pathway also play a crucial role colitis-associated carcinoma.

Figure 3.
The Wnt signalling pathway in the biochemical process of colorectal cancer (CRC). The aberrant Wnt pathway drives the cancer stem cells, the tumour microenvironment, angiogenesis, invasion, metastasis, and drug resistance in CRC.
Figure 3.
The Wnt signalling pathway in the biochemical process of colorectal cancer (CRC). The aberrant Wnt pathway drives the cancer stem cells, the tumour microenvironment, angiogenesis, invasion, metastasis, and drug resistance in CRC.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhu, Y.; Li, X. Advances of Wnt Signalling Pathway in Colorectal Cancer. Cells 2023, 12, 447. https://doi.org/10.3390/cells12030447
AMA Style
Zhu Y, Li X. Advances of Wnt Signalling Pathway in Colorectal Cancer. Cells. 2023; 12(3):447. https://doi.org/10.3390/cells12030447
Chicago/Turabian StyleZhu, Yaoyao, and Xia Li. 2023. "Advances of Wnt Signalling Pathway in Colorectal Cancer" Cells 12, no. 3: 447. https://doi.org/10.3390/cells12030447
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.