
Experimentally Created Magnetic Force in Microbiological Space and On-Earth Studies: Perspectives and Restrictions
 , ,
, , 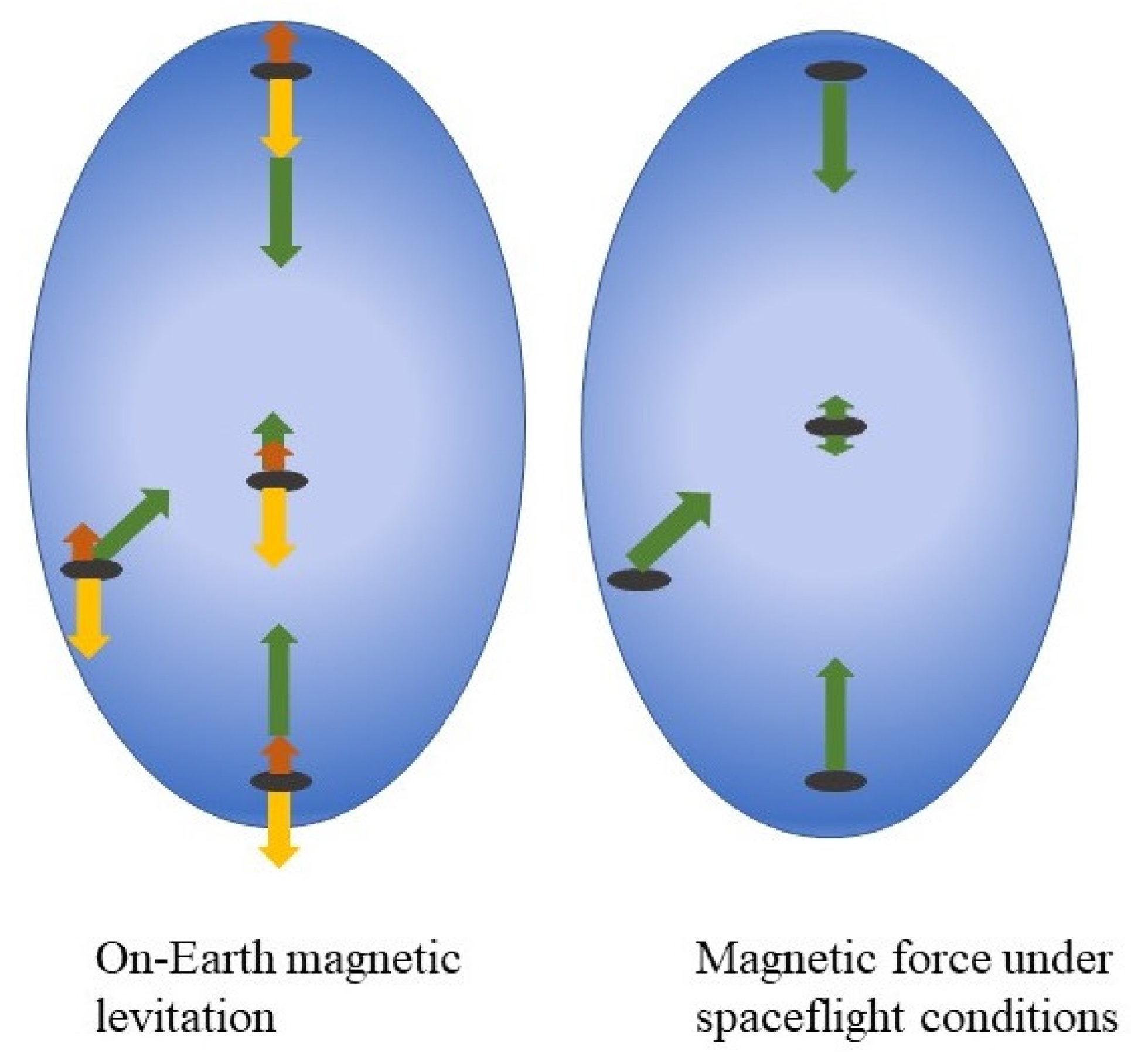{kind=link}
{kind=link}
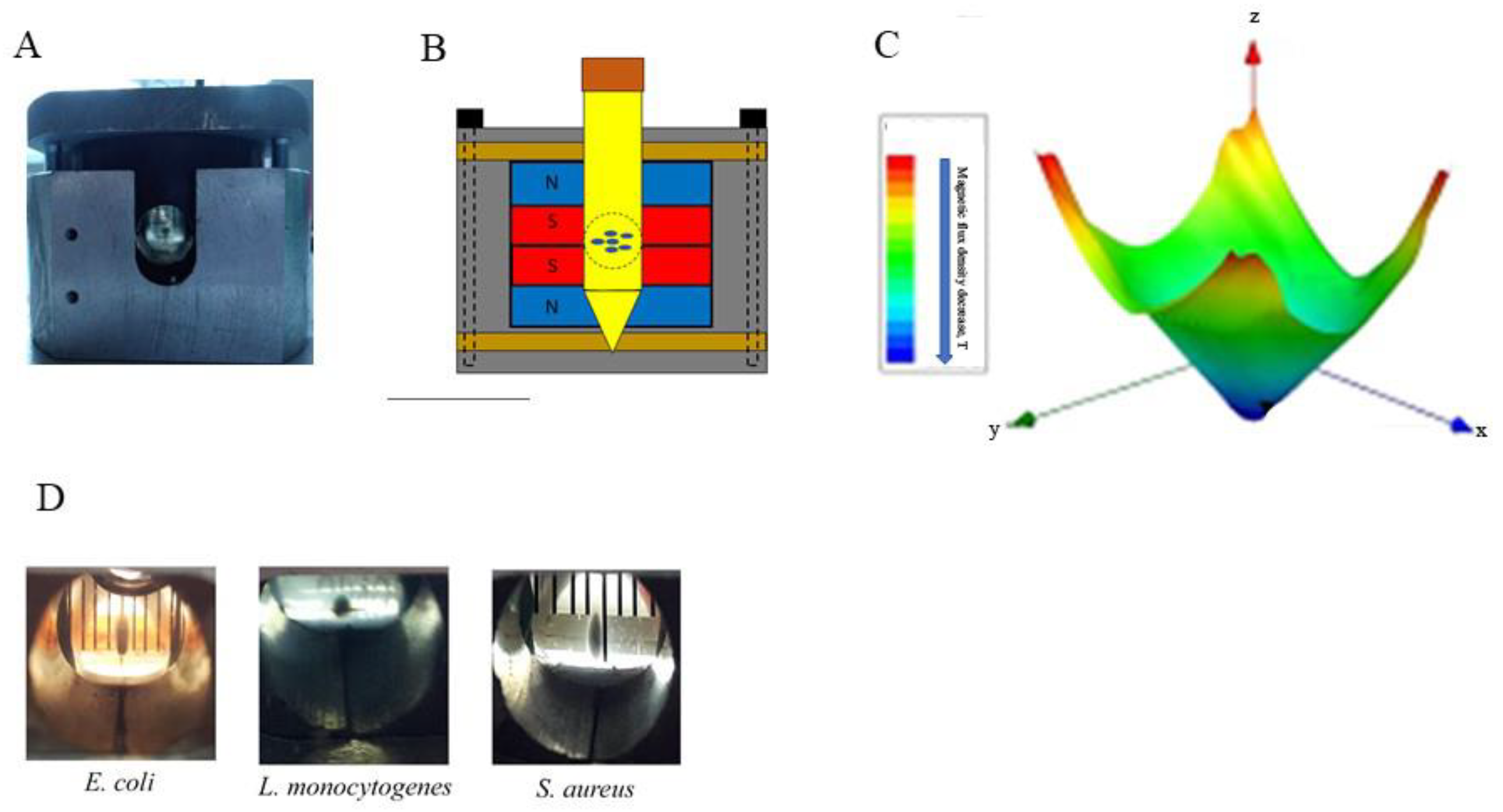{kind=link}
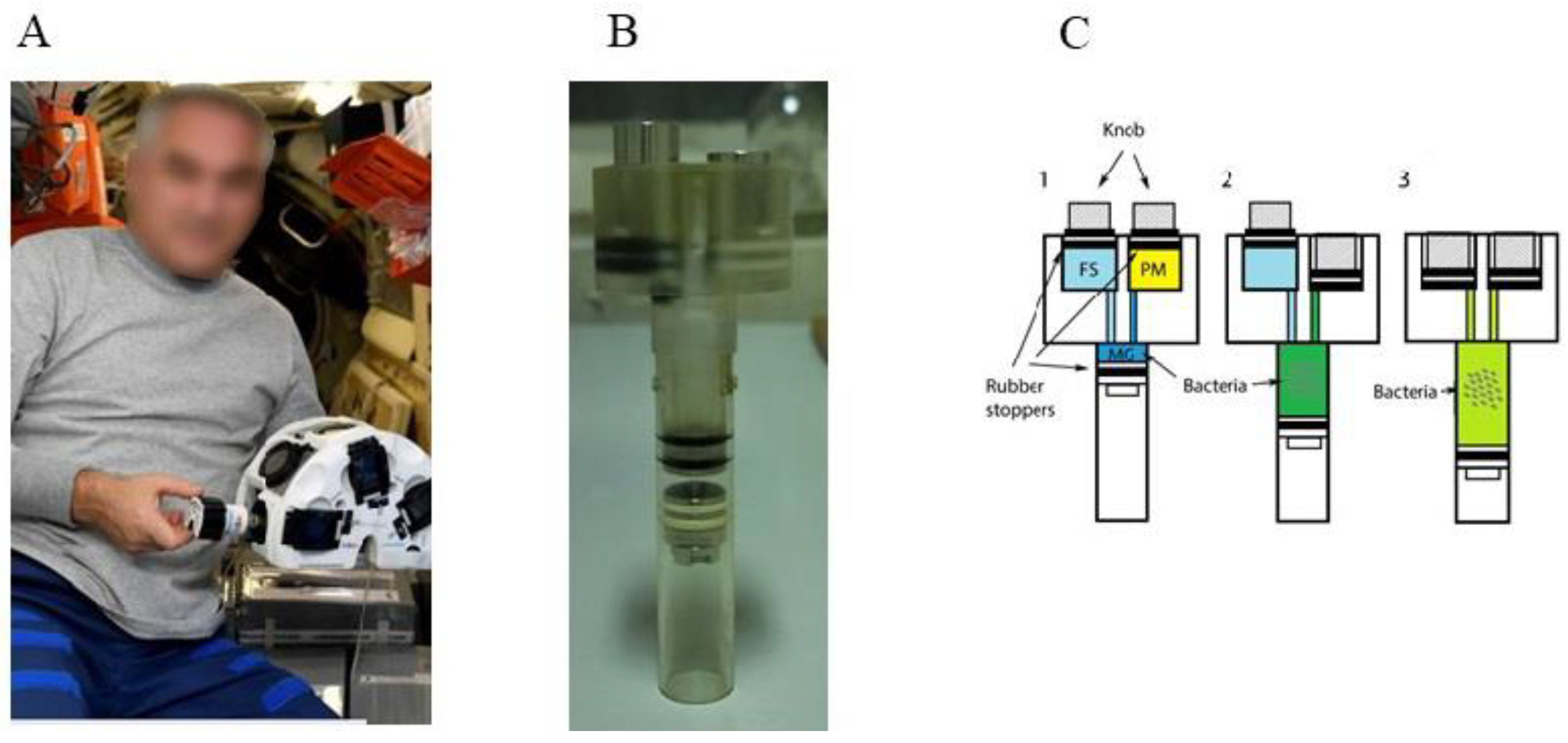{kind=link}
Abstract
:1. Introduction
2. Magnetic Force vs. Gravity
3. Superconducting Magnets for Magnetic Levitation of Living Objects
4. MagLev Systems Based on Permanent Magnets
5. Using the Magnetic Force for Cell Separation and Differentiation
6. Using a Magnetic Force for Cell Integration and Aggregation
7. Magnetic Force Application in Space Studies
8. Future Industrial Applications of the Magnetic Force in Microbiological Studies
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nickerson, C.A.; Ott, C.M.; Wilson, J.W.; Ramamurthy, R.; Pierson, D.L. Microbial Responses to Microgravity and Other Low-Shear Environments. Microbiol. Mol. Biol. Rev. 2004, 68, 345–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higginson, E.E.; Galen, J.E.; Levine, M.M.; Tennant, S.M. Microgravity as a biological tool to examine host-pathogen interactions and to guide development of therapeutics and preventatives that target pathogenic bacteria. Pathog. Dis. 2016, 74, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senatore, G.; Mastroleo, F.; Leys, N.; Mauriello, G. Effect of microgravity & space radiation on microbes. Future Microbiol. 2018, 13, 831–847. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Li, D.G.; Huang, Y.; Liu, C.T. Effects of spaceflight and simulated microgravity on microbial growth and secondary metabolism. Mil. Med. Res. 2018, 5, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bijlani, S.; Stephens, E.; Singh, N.K.; Venkateswaran, K.; Wang, C.C.C. Advances in space microbiology. iScience 2021, 24, 102395. [Google Scholar] [CrossRef] [PubMed]
- Klimarev, S.; Il’in, V.; Starkova, L. Choice of a method and a type of device for water decontamination and warming within physical-chemical life support systems. Av. Ekol. Med. 2008, 42, 3–14. [Google Scholar]
- Horneck, G.; Klaus, D.M.; Mancinelli, R.L. Space Microbiology. Microbiol. Mol. Biol. Rev. 2010, 74, 121–156. [Google Scholar] [CrossRef] [Green Version]
- Zea, L.; Larsen, M.; Estante, F.; Qvortrup, K.; Moeller, R.; de Oliveira, S.D.; Stodieck, L.; Klaus, D. Phenotypic changes exhibited by E. coli cultured in space. Front. Microbiol. 2017, 8, 1598. [Google Scholar] [CrossRef] [Green Version]
- Crabbé, A.; Nielsen-Preiss, S.M.; Woolley, C.M.; Barrila, J.; Buchanan, K.; McCracken, J.; Inglis, D.O.; Searles, S.C.; Nelman-Gonzalez, M.A.; Ott, C.M.; et al. Spaceflight enhances cell aggregation and random budding in Candida albicans. PLoS ONE 2013, 8, e80677. [Google Scholar] [CrossRef] [Green Version]
- Gasset, G.; Tixador, R.; Eche, B.; Lapchine, L.; Moatti, N.; Toorop, P.; Woldringh, C. Growth and division of Escherichia coli under microgravity conditions. Res. Microbiol. 1994, 145, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Tixador, R.; Gasset, G.; Eche, B.; Moatti, N.; Lapchine, L.; Woldringh, C.; Toorop, P.; Moatti, J.P.; Delmotte, F.; Tap, G. Behavior of bacteria and antibiotics under space conditions. Aviat. Space Environ. Med. 1994, 65, 551–556. [Google Scholar] [PubMed]
- Fajardo-Cavazos, P.; Leehan, J.D.; Nicholson, W.L. Alterations in the spectrum of spontaneous rifampicin-resistance mutations in the Bacillus subtilis rpoB gene after cultivation in the human spaceflight environment. Front. Microbiol. 2018, 9, 192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirumalai, M.R.; Karouia, F.; Tran, Q.; Stepanov, V.G.; Bruce, R.J.; Ott, C.M.; Pierson, D.L.; Fox, G.E. Evaluation of acquired antibiotic resistance in Escherichia coli exposed to long-term low-shear modeled microgravity and background antibiotic exposure. MBio 2019, 10, e02637-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, P.W. Impact of space flight on bacterial virulence and antibiotic susceptibility. Infect. Drug Resist. 2015, 8, 249–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Liu, F.; Wang, Q.; Ge, P.; Woo, P.C.Y.; Yan, J.; Zhao, Y.; Gao, G.F.; Liu, C.H.; Liu, C. Genomic and transcriptomic analysis of NDM-1 Klebsiella pneumoniae in spaceflight reveal mechanisms underlying environmental adaptability. Sci. Rep. 2014, 4, 6216. [Google Scholar] [CrossRef] [Green Version]
- Vukanti, R.; Model, M.A.; Leff, L.G. Effect of modeled reduced gravity conditions on bacterial morphology and physiology. BMC Microbiol. 2012, 12, 4. [Google Scholar] [CrossRef] [Green Version]
- Wilson, J.W.; Ott, C.M.; Höner Zu Bentrup, K.; Ramamurthy, R.; Quick, L.; Porwollik, S.; Cheng, P.; McClelland, M.; Tsaprailis, G.; Radabaugh, T.; et al. Space flight alters bacterial gene expression and virulence and reveals a role for global regulator Hfq. Proc. Natl. Acad. Sci. USA 2007, 104, 16299–16304. [Google Scholar] [CrossRef] [Green Version]
- Yim, J.; Cho, S.W.; Kim, B.; Park, S.; Han, Y.H.; Seo, S.W. Transcriptional profiling of the probiotic Escherichia Coli nissle 1917 strain under simulated microgravity. Int. J. Mol. Sci. 2020, 21, 2666. [Google Scholar] [CrossRef]
- Domnin, P.A.; Parfenov, V.A.; Kononikhin, A.S.; Petrov, S.V.; Shevlyagina, N.V.; Arkhipova, A.Y.; Koudan, E.V.; Nezhurina, E.K.; Brzhozovskiy, A.G.; Bugrova, A.E.; et al. Combined Impact of Magnetic Force and Spaceflight Conditions on Escherichia Coli Physiology. Int. J. Mol. Sci. 2022, 23, 1837. [Google Scholar] [CrossRef]
- Arunasri, K.; Adil, M.; Venu Charan, K.; Suvro, C.; Himabindu Reddy, S.; Shivaji, S. Effect of Simulated Microgravity on E. coli K12 MG1655 Growth and Gene Expression. PLoS ONE 2013, 8, e57860. [Google Scholar] [CrossRef] [Green Version]
- Morrison, M.D.; Fajardo-Cavazos, P.; Nicholson, W.L. Comparison of Bacillus subtilis transcriptome profiles from two separate missions to the International Space Station. NPJ Microgravity 2019, 5, 1. [Google Scholar] [CrossRef] [PubMed]
- Tixador, R.; Richoilley, G.; Gasset, G.; Planel, H.; Moatti, N.; Lapchine, L.; Enjalbert, L.; Raffin, J.; Bost, R.; Zaloguev, S.N.; et al. Preliminary results of cytos 2 experiment. Acta Astronaut. 1985, 12, 131–134. [Google Scholar] [CrossRef] [PubMed]
- Juergensmeyer, M.A.; Juergensmeyer, E.A.; Guikema, J.A. Long-term exposure to spaceflight conditions affects bacterial response to antibiotics. Microgravity Sci. Technol. 1999, 12, 41–47. [Google Scholar] [PubMed]
- Sharma, G.; Curtis, P.D. The Impacts of Microgravity on Bacterial Metabolism. Life 2022, 12, 774. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.B.; Klaus, D.; Todd, P. Effects of space flight, clinorotation, and centrifugation on the substrate utilization efficiency of E. coli. Microgravity Sci. Technol. 2002, 13, 24–29. [Google Scholar] [CrossRef]
- Herranz, R.; Anken, R.; Boonstra, J.; Braun, M.; Christianen, P.C.M.; De Geest, M.; Hauslage, J.; Hilbig, R.; Hill, R.J.A.; Lebert, M.; et al. Ground-based facilities for simulation of microgravity: Organism-specific recommendations for their use, and recommended terminology. Astrobiology 2013, 13, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Kiss, J.Z.; Wolverton, C.; Wyatt, S.E.; Hasenstein, K.H.; van Loon, J.J.W.A. Comparison of Microgravity Analogs to Spaceflight in Studies of Plant Growth and Development. Front. Plant Sci. 2019, 10, 1577. [Google Scholar] [CrossRef] [Green Version]
- Valles, J.M.; Guevorkian, K. Low gravity on earth by magnetic levitation of biological material. J. Gravit. Physiol. 2002, 9, 11–14. [Google Scholar]
- Berry, M.V.; Geim, A.K. Of flying frogs and levitrons. Eur. J. Phys. 1997, 18, 307–313. [Google Scholar] [CrossRef]
- Ashkarran, A.A.; Mahmoudi, M. Magnetic Levitation Systems for Disease Diagnostics. Trends Biotechnol. 2021, 39, 311–321. [Google Scholar] [CrossRef]
- Brooks, J.S.; Reavis, J.A.; Medwood, R.A.; Stalcup, T.F.; Meisel, M.W.; Steinberg, E.; Arnowitz, L.; Stover, C.C.; Perenboom, J.A.A.J. New opportunities in science, materials, and biological systems in the low-gravity (magnetic levitation) environment (invited). J. Appl. Phys. 2000, 87, 6194. [Google Scholar] [CrossRef] [Green Version]
- Panina, L.V.; Gurevich, A.; Beklemisheva, A.; Omelyanchik, A.; Levada, K.; Rodionova, V. Spatial Manipulation of Particles and Cells at Micro-and Nanoscale via Magnetic Forces. Cells 2022, 11, 950. [Google Scholar] [CrossRef] [PubMed]
- Khalil, I.S.M.; Klingner, A.; Misra, S. Theory of electromagnetics: Soft-magnetic bodies. In Mathematical Modelling of Swiming Soft Microrobots; Academic Press: Cambridge, MA, USA, 2021; pp. 43–59. [Google Scholar] [CrossRef]
- Gaeta, M.; Cavallaro, M.; Vinci, S.L.; Mormina, E.; Blandino, A.; Marino, M.A.; Granata, F.; Tessitore, A.; Galletta, K.; D’Angelo, T.; et al. Magnetism of materials: Theory and practice in magnetic resonance imaging. Insights Imaging 2021, 12, 179. [Google Scholar] [CrossRef] [PubMed]
- Yamato, M.; Kimura, T. Magnetic Processing of Diamagnetic Materials. Polymers 2020, 12, 1491. [Google Scholar] [CrossRef] [PubMed]
- Guevorkian, K.; Valles, J.M. Swimming Paramecium in magnetically simulated enhanced, reduced, and inverted gravity environments. Proc. Natl. Acad. Sci. USA 2006, 103, 13051–13056. [Google Scholar] [CrossRef] [Green Version]
- Barber-Zucker, S.; Zarivach, R. A Look into the Biochemistry of Magnetosome Biosynthesis in Magnetotactic Bacteria. ACS Chem. Biol. 2017, 12, 13–22. [Google Scholar] [CrossRef]
- Goswami, P.; He, K.; Li, J.; Pan, Y.; Roberts, A.P.; Lin, W. Magnetotactic bacteria and magnetofossils: Ecology, evolution and environmental implications. NPJ Biofilms Microb. 2022, 8, 43. [Google Scholar] [CrossRef]
- Meister, M. Physical limits to magnetogenetics. Elife 2016, 5, e17210. [Google Scholar] [CrossRef]
- Schenck, J.F. Physical interactions of static magnetic fields with living tissues. Prog. Biophys. Mol. Biol. 2005, 87, 185–204. [Google Scholar] [CrossRef]
- Letuta, U.G.; Berdinskiy, V.L. Biological effects of static magnetic fields and zinc isotopes on E. coli bacteria. Bioelectromagnetics 2019, 40, 62–73. [Google Scholar] [CrossRef] [Green Version]
- Letuta, U.G.; Avdeeva, E.I. Magnetic-dependent ATP pool in Escherichia coli. Dokl. Biochem. Biophys. 2017, 474, 196–199. [Google Scholar] [CrossRef] [PubMed]
- Dijkstra, C.E.; Larkin, O.J.; Anthony, P.; Davey, M.R.; Eaves, L.; Rees, C.E.D.; Hill, R.J.A. Diamagnetic levitation enhances growth of liquid bacterial cultures by increasing oxygen availability. J. R. Soc. Interface 2011, 8, 334–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valles, J.M.; Maris, H.J.; Seidel, G.M.; Tang, J.; Yao, W. Magnetic levitation-based Martian and Lunar gravity simulator. Adv. Space Res. 2005, 36, 114–118. [Google Scholar] [CrossRef]
- Valles, J.M.; Lin, K.; Denegre, J.M.; Mowry, K.L. Stable magnetic field gradient levitation of Xenopus laevis: Toward low-gravity simulation. Biophys. J. 1997, 73, 1130–1133. [Google Scholar] [CrossRef] [Green Version]
- Manzano, A.I.; van Loon, J.J.W.A.; Christianen, P.C.M.; Gonzalez-Rubio, J.M.; Medina, F.J.; Herranz, R. Gravitational and magnetic field variations synergize to cause subtle variations in the global transcriptional state of Arabidopsis in vitro callus cultures. BMC Genom. 2012, 13, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanavandi, H.; Guo, W. A magnetic levitation based low-gravity simulator with an unprecedented large functional volume. NPJ microgravity 2021, 7, 40. [Google Scholar] [CrossRef]
- Tsuchiya, K.; Nakamura, K.; Okuno, K.; Ano, T.; Shoda, M. Effect of homogeneous and inhomogeneous high magnetic fields on the growth of Escherichia coli. J. Ferment. Bioeng. 1996, 81, 343–346. [Google Scholar] [CrossRef]
- Tsuchiya, K.; Okuno, K.; Ano, T.; Tanaka, K.; Takahashi, H.; Shoda, M. High magnetic field enhances stationary phase-specific transcription activity of Escherichia coli. Bioelectrochem. Bioenerg. 1999, 48, 383–387. [Google Scholar] [CrossRef]
- Liu, M.; Gao, H.; Shang, P.; Zhou, X.; Ashforth, E.; Zhuo, Y.; Chen, D.; Ren, B.; Liu, Z.; Zhang, L. Magnetic field is the dominant factor to induce the response of Streptomyces avermitilis in altered gravity simulated by diamagnetic levitation. PLoS ONE 2011, 6, e24697. [Google Scholar] [CrossRef]
- Zablotskii, V.; Polyakova, T.; Lunov, O.; Dejneka, A. How a High-Gradient Magnetic Field Could Affect Cell Life. Sci. Rep. 2016, 6, 37407. [Google Scholar] [CrossRef] [Green Version]
- Domnin, P.; Arkhipov, A.; Petrov, S.; Sysolyatina, E.; Parfenov, V.; Karalkin, P.; Mukhachev, A.; Gusarov, A.; Moisenovich, M.; Khesuani, Y.; et al. An In Vitro Model of Nonattached Biofilm-Like Bacterial Aggregates Based on Magnetic Levitation. Appl. Environ. Microbiol. 2020, 86, e01074-20. [Google Scholar] [CrossRef] [PubMed]
- Herranz, R.; Larkin, O.J.; Dijkstra, C.E.; Hill, R.J.A.; Anthony, P.; Davey, M.R.; Eaves, L.; van Loon, J.J.W.A.; Medina, F.J.; Marco, R. Microgravity simulation by diamagnetic levitation: Effects of a strong gradient magnetic field on the transcriptional profile of Drosophila melanogaster. BMC Genom. 2012, 13, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zhang, N.; Di, J.; Zhao, W.; Shi, G.; Xie, R.; Hu, B.; Yang, H. Analysis of the effects of magnetic levitation to simulate microgravity environment on the Arp2/3 complex pathway in macrophage. J. Biol. Phys. 2021, 47, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Go, G.; Yoo, A.; Kang, B.; Choi, E.; Park, J.O.; Kim, C.S. Wearable Fixation Device for a Magnetically Controllable Therapeutic Agent Carrier: Application to Cartilage Repair. Pharmaceutics 2020, 12, 593. [Google Scholar] [CrossRef] [PubMed]
- Mirica, K.A.; Ilievski, F.; Ellerbee, A.K.; Shevkoplyas, S.S.; Whitesides, G.M. Using Magnetic Levitation for Three Dimensional Self-Assembly. Adv. Mater. 2011, 23, 4134–4140. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, A.B.; Yang, D.; Yu, H.D.; Nemiroski, A.; Tricard, S.; Ellerbee, A.K.; Soh, S.; Whitesides, G.M. Noncontact orientation of objects in three-dimensional space using magnetic levitation. Proc. Natl. Acad. Sci. USA 2014, 111, 12980–12985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durmus, N.G.; Tekin, H.C.; Guven, S.; Sridhar, K.; Yildiz, A.A.; Calibasi, G.; Ghiran, I.; Davis, R.W.; Steinmetz, L.M.; Demirci, U. Magnetic levitation of single cells. Proc. Natl. Acad. Sci. USA 2015, 112, E3661–E3668. [Google Scholar] [CrossRef] [Green Version]
- Parfenov, V.A.; Koudan, E.V.; Bulanova, E.A.; Karalkin, P.A.; Das Pereira, F.; Norkin, N.E.; Knyazeva, A.D.; Gryadunova, A.A.; Petrov, O.F.; Vasiliev, M.M.; et al. Scaffold-free, label-free and nozzle-free biofabrication technology using magnetic levitational assembly. Biofabrication 2018, 10, 034104. [Google Scholar] [CrossRef]
- Parfenov, V.A.; Petrov, S.V.; Pereira, F.D.A.S.; Levin, A.A.; Koudan, E.V.; Nezhurina, E.K.; Karalkin, P.A.; Vasiliev, M.M.; Petrov, O.F.; Komlev, V.S.; et al. Scaffold-free, Label-free, and Nozzle-free Magnetic Levitational Bioassembler for Rapid Formative Biofabrication of 3D Tissues and Organs. Int. J. Bioprint. 2020, 6, 110–119. [Google Scholar] [CrossRef]
- Do, Q.N.; Lenkinski, R.E.; Tircso, G.; Kovacs, Z. How the Chemical Properties of GBCAs Influence Their Safety Profiles In Vivo. Molecules 2021, 27, 58. [Google Scholar] [CrossRef]
- Anderhalten, L.; Silva, R.V.; Morr, A.; Wang, S.; Smorodchenko, A.; Saatz, J.; Traub, H.; Mueller, S.; Boehm-Sturm, P.; Rodriguez-Sillke, Y.; et al. Different Impact of Gadopentetate and Gadobutrol on Inflammation-Promoted Retention and Toxicity of Gadolinium Within the Mouse Brain. Investig. Radiol. 2022, 57, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Erdoğan, M.A.; Apaydin, M.; Armagan, G.; Taskiran, D. Evaluation of toxicity of gadolinium-based contrast agents on neuronal cells. Acta Radiol. 2021, 62, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Tisa, L.S.; Adler, J. Calcium ions are involved in Escherichia coli chemotaxis. Proc. Natl. Acad. Sci. USA 1992, 89, 11804–11808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orans, J.; Johnson, M.D.L.; Coggan, K.A.; Sperlazza, J.R.; Heiniger, R.W.; Wolfgang, M.C.; Redinbo, M.R. Crystal structure analysis reveals Pseudomonas PilY1 as an essential calcium-dependent regulator of bacterial surface motility. Proc. Natl. Acad. Sci. USA 2010, 107, 1065–1070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, H.K.; Shrivastava, S.; Sharma, R. A novel calcium uptake transporter of uncharacterized P-type ATPase family supplies calcium for cell surface integrity in Mycobacterium smegmatis. Mbio 2017, 8, e01388-17. [Google Scholar] [CrossRef] [Green Version]
- Nemiroski, A.; Soh, S.; Kwok, S.W.; Yu, H.D.; Whitesides, G.M. Tilted Magnetic Levitation Enables Measurement of the Complete Range of Densities of Materials with Low Magnetic Permeability. J. Am. Chem. Soc. 2016, 138, 1252–1257. [Google Scholar] [CrossRef] [Green Version]
- Ge, S.; Whitesides, G.M. “Axial” Magnetic Levitation Using Ring Magnets Enables Simple Density-Based Analysis, Separation, and Manipulation. Anal. Chem. 2018, 90, 12239–12245. [Google Scholar] [CrossRef]
- Ge, S.; Wang, Y.; Deshler, N.J.; Preston, D.J.; Whitesides, G.M. High-Throughput Density Measurement Using Magnetic Levitation. J. Am. Chem. Soc. 2018, 140, 7510–7518. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, P.; Tang, D.; Xia, N.; Zhang, X.; Nie, J.; Gu, F.; Zhou, H.; Fu, J. Axial magnetic levitation: A high-sensitive and maneuverable density-based analysis device. Sens. Actuators B Chem. 2020, 304, 127362. [Google Scholar] [CrossRef]
- Parfenov, V.A.; Mironov, V.A.; Van Kampen, K.A.; Karalkin, P.A.; Koudan, E.V.; Pereira, F.D.A.S.; Petrov, S.V.; Nezhurina, E.K.; Petrov, O.F.; Myasnikov, M.I.; et al. Scaffold-free and label-free biofabrication technology using levitational assembly in a high magnetic field. Biofabrication 2020, 12, 045022. [Google Scholar] [CrossRef]
- Zhang, Z. The flocculation mechanism and treatment of oily wastewater by flocculation. Water Sci. Technol. 2017, 76, 2630–2637. [Google Scholar] [CrossRef] [PubMed]
- Heidari, M. Role of Natural Flocculation in Eliminating Toxic Metals. Arch. Environ. Contam. Toxicol. 2019, 76, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Salehizadeh, H.; Yan, N.; Farnood, R. Recent advances in polysaccharide bio-based flocculants. Biotechnol. Adv. 2018, 36, 92–119. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, D.; Foubert, I.; Muylaert, K. Flocculation as a low-cost method for harvesting microalgae for bulk biomass production. Trends Biotechnol. 2013, 31, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Walch, H.; von der Kammer, F.; Hofmann, T. Freshwater suspended particulate matter—Key components and processes in floc formation and dynamics. Water Res. 2022, 220, 118655. [Google Scholar] [CrossRef]
- Alhede, M.; Kragh, K.N.; Qvortrup, K.; Allesen-Holm, M.; van Gennip, M.; Christensen, L.D.; Jensen, P.Ø.; Nielsen, A.K.; Parsek, M.; Wozniak, D.; et al. Phenotypes of non-attached pseudomonas aeruginosa aggregates resemble surface attached biofilm. PLoS ONE 2011, 6, e27943. [Google Scholar] [CrossRef]
- Tremblay, Y.D.N.; Labrie, J.; Chénier, S.; Jacques, M. Actinobacillus pleuropneumoniae grows as aggregates in the lung of pigs: Is it time to refine our in vitro biofilm assays? Microb. Biotechnol. 2017, 10, 756–760. [Google Scholar] [CrossRef]
- Anderson, G.G.; Palermo, J.J.; Schilling, J.D.; Roth, R.; Heuser, J.; Hultgren, S.J. Intracellular bacterial biofilm-like pods in urinary tract infections. Science 2003, 301, 105–107. [Google Scholar] [CrossRef] [Green Version]
- Demirdjian, S.; Sanchez, H.; Hopkins, D.; Berwin, B. Motility-Independent Formation of Antibiotic-Tolerant Pseudomonas aeruginosa Aggregates. Appl. Environ. Microbiol. 2019, 85, e00844-19. [Google Scholar] [CrossRef] [Green Version]
- Parfenov, V.A.; Khesuani, Y.D.; Petrov, S.V.; Karalkin, P.A.; Koudan, E.V.; Nezhurina, E.K.; Pereira, F.D.A.S.; Krokhmal, A.A.; Gryadunova, A.A.; Bulanova, E.A.; et al. Magnetic levitational bioassembly of 3D tissue construct in space. Sci. Adv. 2020, 6, eaba4174. [Google Scholar] [CrossRef]
- Acres, J.M.; Youngapelian, M.J.; Nadeau, J. The influence of spaceflight and simulated microgravity on bacterial motility and chemotaxis. NPJ Microgravity 2021, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Klaus, D.; Simske, S.; Todd, P.; Stodieck, L. Investigation of space flight effects on Escherichia coil and a proposed model of underlying physical mechanisms. Microbiology 1997, 143, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Crabbé, A.; De Boever, P.; Van Houdt, R.; Moors, H.; Mergeay, M.; Cornelis, P. Use of the rotating wall vessel technology to study the effect of shear stress on growth behaviour of Pseudomonas aeruginosa PA01. Environ. Microbiol. 2008, 10, 2098–2110. [Google Scholar] [CrossRef] [PubMed]
- Castro, S.L.; Nelman-Gonzalez, M.; Nickerson, C.A.; Ott, C.M. Induction of attachment-independent biofilm formation and repression of hfq expression by low-fluid-shear culture of Staphylococcus aureus. Appl. Environ. Microbiol. 2011, 77, 6368–6378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kjaergaard, K.; Schembri, M.A.; Hasman, H.; Klemm, P. Antigen 43 from Escherichia coli induces inter- and intraspecies cell aggregation and changes in colony morphology of Pseudomonas fluorescens. J. Bacteriol. 2000, 182, 4789–4796. [Google Scholar] [CrossRef] [Green Version]
- Maloy, S.R.; Bohlander, M.; Nunn, W.D. Elevated levels of glyoxylate shunt enzymes in Escherichia coli strains constitutive for fatty acid degradation. J. Bacteriol. 1980, 143, 720–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renilla, S.; Bernal, V.; Fuhrer, T.; Castaño-Cerezo, S.; Pastor, J.M.; Iborra, J.L.; Sauer, U.; Cánovas, M. Acetate scavenging activity in Escherichia coli: Interplay of acetyl-CoA synthetase and the PEP-glyoxylate cycle in chemostat cultures. Appl. Microbiol. Biotechnol. 2012, 93, 2109–2124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Bryant, D.A. Biochemical validation of the glyoxylate cycle in the cyanobacterium Chlorogloeopsis fritschii strain PCC 9212. J. Biol. Chem. 2015, 290, 14019–14030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaval, K.G.; Garsin, D.A. Ethanolamine utilization in bacteria. MBio 2018, 9, e00066-18. [Google Scholar] [CrossRef] [Green Version]
- Garsin, D.A. Ethanolamine utilization in bacterial pathogens: Roles and regulation. Nat. Rev. Microbiol. 2010, 8, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.M.; Go, J.; Yoon, M.Y.; Park, Y.; Kim, S.C.; Yong, D.E.; Yoon, S.S. Vitamin B 12-Mediated restoration of defective anaerobic growth leads to reduced biofilm formation in Pseudomonas aeruginosa. Infect. Immun. 2012, 80, 1639–1649. [Google Scholar] [CrossRef] [PubMed]
- Chang, G.W.; Chang, J.T. Evidence for the B12-dependent enzyme ethanolamine deaminase in Salmonella. Nature 1975, 254, 150–151. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, C.M.; Turner, J.M. Microbial metabolism of amino alcohols. Purification and properties of coenzyme B12-dependent ethanolamine ammonia-lyase of Escherichia coli. Biochem. J. 1978, 175, 555–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fajardo-Cavazos, P.; Nicholson, W.L. Mechanotransduction in Prokaryotes: A Possible Mechanism of Spaceflight Adaptation. Life 2021, 11, 33. [Google Scholar] [CrossRef]
- Zeikus, J.G.; Jain, M.K.; Elankovan, P. Biotechnology of succinic acid production and markets for derived industrial products. Appl. Microbiol. Biotechnol. 1999, 51, 545–552. [Google Scholar] [CrossRef]
- Birajdar, M.S.; Joo, H.; Koh, W.G.; Park, H. Natural bio-based monomers for biomedical applications: A review. Biomater. Res. 2021, 25, 8. [Google Scholar] [CrossRef]
- Pham, J.V.; Yilma, M.A.; Feliz, A.; Majid, M.T.; Maffetone, N.; Walker, J.R.; Kim, E.; Cho, H.J.; Reynolds, J.M.; Song, M.C.; et al. A review of the microbial production of bioactive natural products and biologics. Front. Microbiol. 2019, 10, 4881–4905. [Google Scholar] [CrossRef] [Green Version]
- Yin, X.; Li, J.; Shin, H.-D.; Du, G.; Liu, L.; Chen, J. Metabolic engineering in the biotechnological production of organic acids in the tricarboxylic acid cycle of microorganisms: Advances and prospects. Biotechnol. Adv. 2015, 33, 830–841. [Google Scholar] [CrossRef]
- Vuoristo, K.S.; Mars, A.E.; Sanders, J.P.M.; Eggink, G.; Weusthuis, R.A. Metabolic Engineering of TCA Cycle for Production of Chemicals. Trends Biotechnol. 2016, 34, 191–197. [Google Scholar] [CrossRef]
- Liu, J.; Wu, X.; Yao, M.; Xiao, W.; Zha, J. Chassis engineering for microbial production of chemicals: From natural microbes to synthetic organisms. Curr. Opin. Biotechnol. 2020, 66, 105–112. [Google Scholar] [CrossRef]
- Huangfu, J.; Zhang, G.; Li, J.; Li, C. Advances in engineered microorganisms for improving metabolic conversion via microgravity effects. Bioeng. 2015, 6, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.J.; Song, H.; Lee, S.Y. Genome-based metabolic engineering of Mannheimia succiniciproducens for succinic acid production. Appl. Environ. Microbiol. 2006, 72, 1939–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nghiem, N.; Davison, B.; Suttle, B.; Richardson, G. Production of succinic acid by Anaerobiospirillum succiniciproducens. Appl. Biochem. Biotechnol. 1997, 63–65, 565–576. [Google Scholar] [CrossRef] [PubMed]
- Louasté, B.; Eloutassi, N. Succinic acid production from whey and lactose by Actinobacillus succinogenes 130Z in batch fermentation. Biotechnol. Rep. 2020, 27, e00481. [Google Scholar] [CrossRef] [PubMed]
- Bretz, K.; Kabasci, S. Feed-control development for succinic acid production with Anaerobiospirillum succiniciproducens. Biotechnol. Bioeng. 2012, 109, 1187–1192. [Google Scholar] [CrossRef]
- Vemuri, G.N.; Eiteman, M.A.; Altman, E. Succinate production in dual-phase Escherichia coli fermentations depends on the time of transition from aerobic to anaerobic conditions. J. Ind. Microbiol. Biotechnol. 2002, 28, 325–332. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ermolaeva, S.A.; Parfenov, V.A.; Karalkin, P.A.; Khesuani, Y.D.; Domnin, P.A. Experimentally Created Magnetic Force in Microbiological Space and On-Earth Studies: Perspectives and Restrictions. Cells 2023, 12, 338. https://doi.org/10.3390/cells12020338
Ermolaeva SA, Parfenov VA, Karalkin PA, Khesuani YD, Domnin PA. Experimentally Created Magnetic Force in Microbiological Space and On-Earth Studies: Perspectives and Restrictions. Cells. 2023; 12(2):338. https://doi.org/10.3390/cells12020338
Chicago/Turabian StyleErmolaeva, Svetlana A., Vladislav A. Parfenov, Pavel A. Karalkin, Yusef D. Khesuani, and Pavel A. Domnin. 2023. "Experimentally Created Magnetic Force in Microbiological Space and On-Earth Studies: Perspectives and Restrictions" Cells 12, no. 2: 338. https://doi.org/10.3390/cells12020338