
iPSC-Derived Striatal Medium Spiny Neurons from Patients with Multiple System Atrophy Show Hypoexcitability and Elevated α-Synuclein Release
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation and Cultivation of Human, Induced Pluripotent Stem Cells (iPSCs)
2.2. Trilineage Differentiation
2.3. Karyotyping
2.4. Differentiation into Striatal Medium Spiny Neurons (MSNs)
2.5. Immunocytochemistry
2.6. Analysis of Synaptic Density and Quantitative Image Analysis
2.7. Quantification of α-Synuclein Release by Dot Blot
2.8. Quantitative Real-Time PCR
2.9. Calcium Imaging
2.10. Electrophysiology
2.11. Statistical Analyses
3. Results
3.1. Generation of iPSC Lines from MSA-P Patients and Healthy Controls
3.2. Differentiation of iPSCs into Striatal Medium Spiny Neurons (MSNs)
3.3. Functional Properties of MSNs
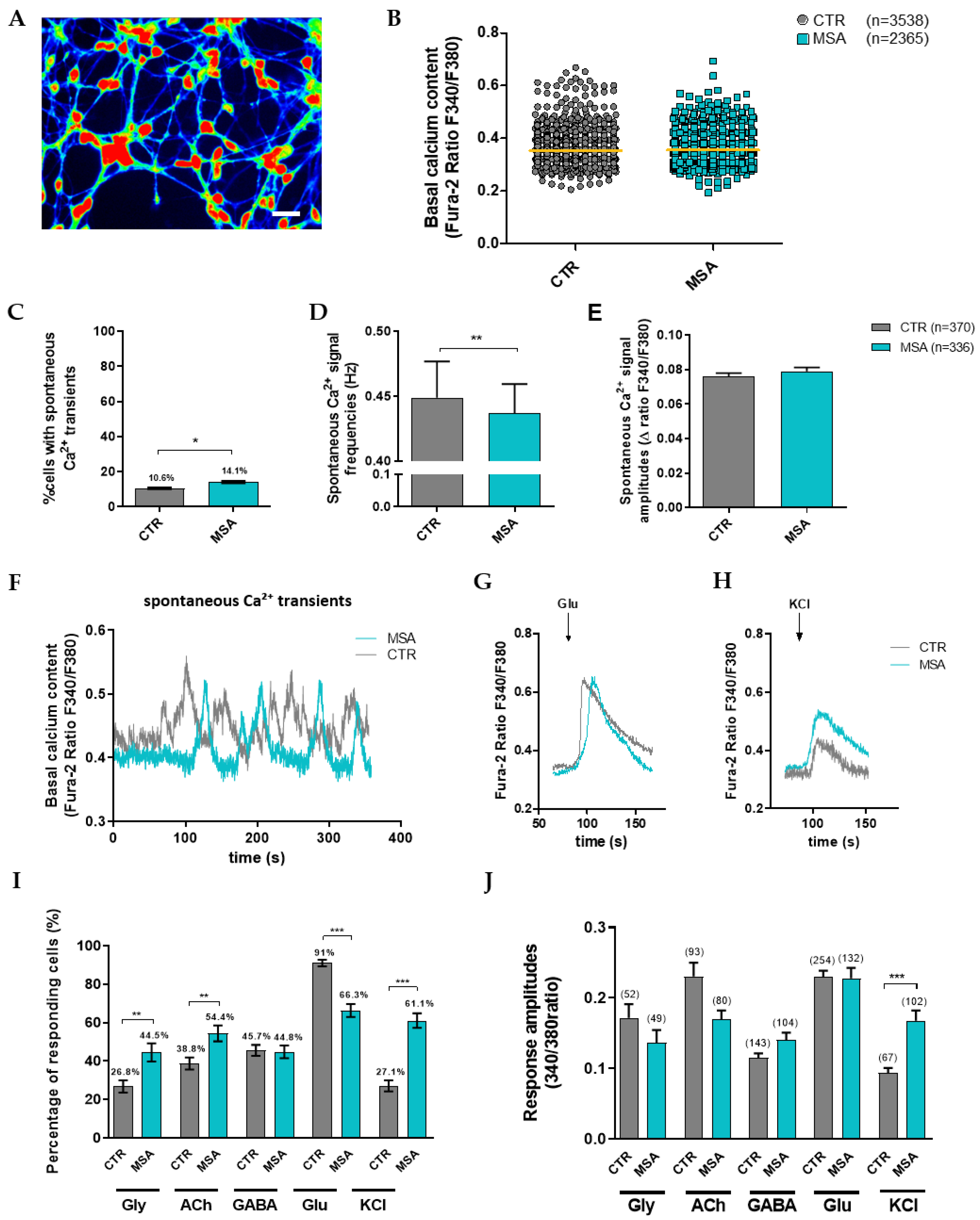
3.3.1. Calcium Signaling in Differentiated MSNs
3.3.2. Altered Responsiveness to Neurotransmitters and KCl in MSA-P MSNs
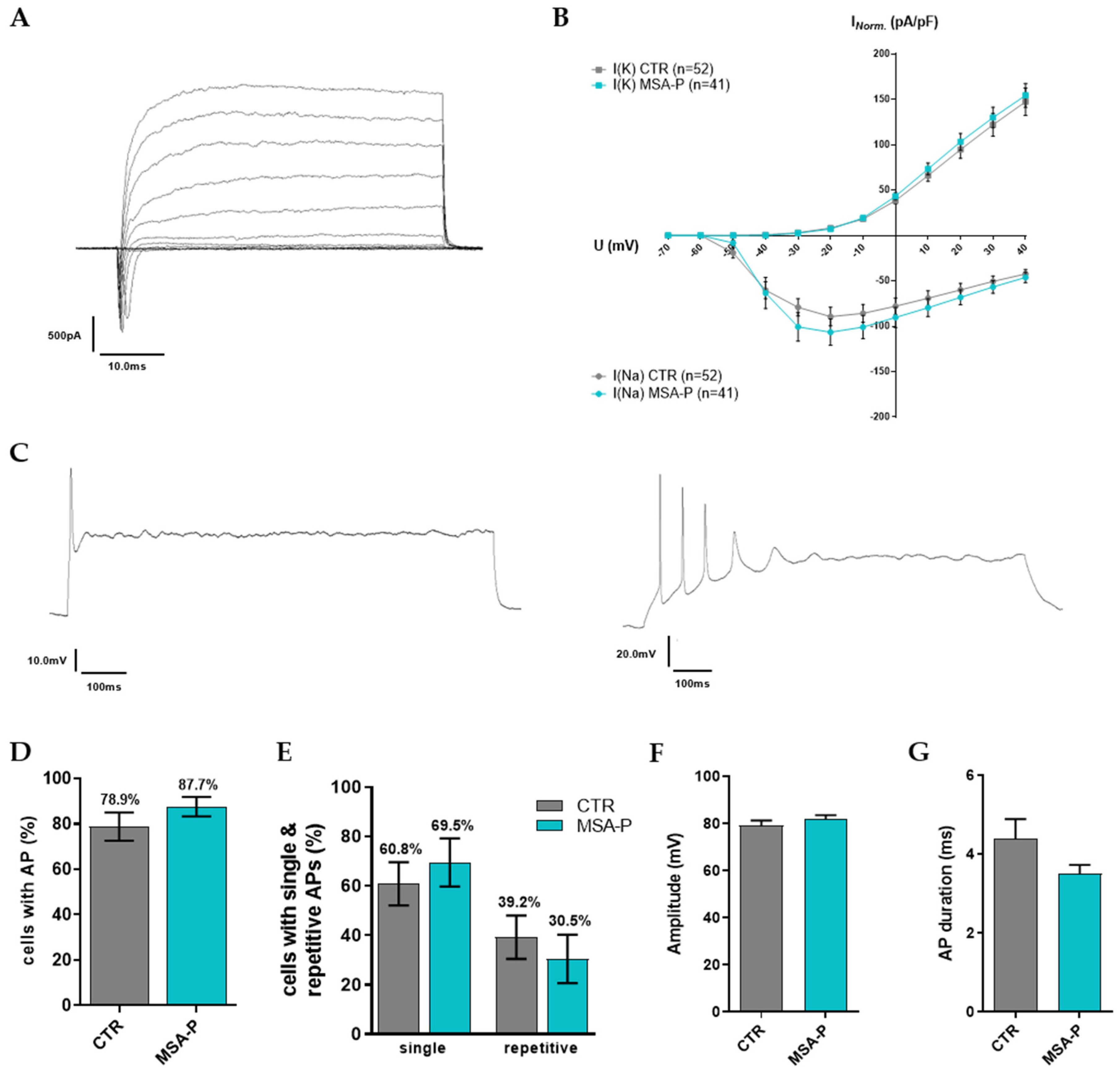
3.3.3. Characteristics of Voltage-Gated ion Channels and Evoked Action Potentials
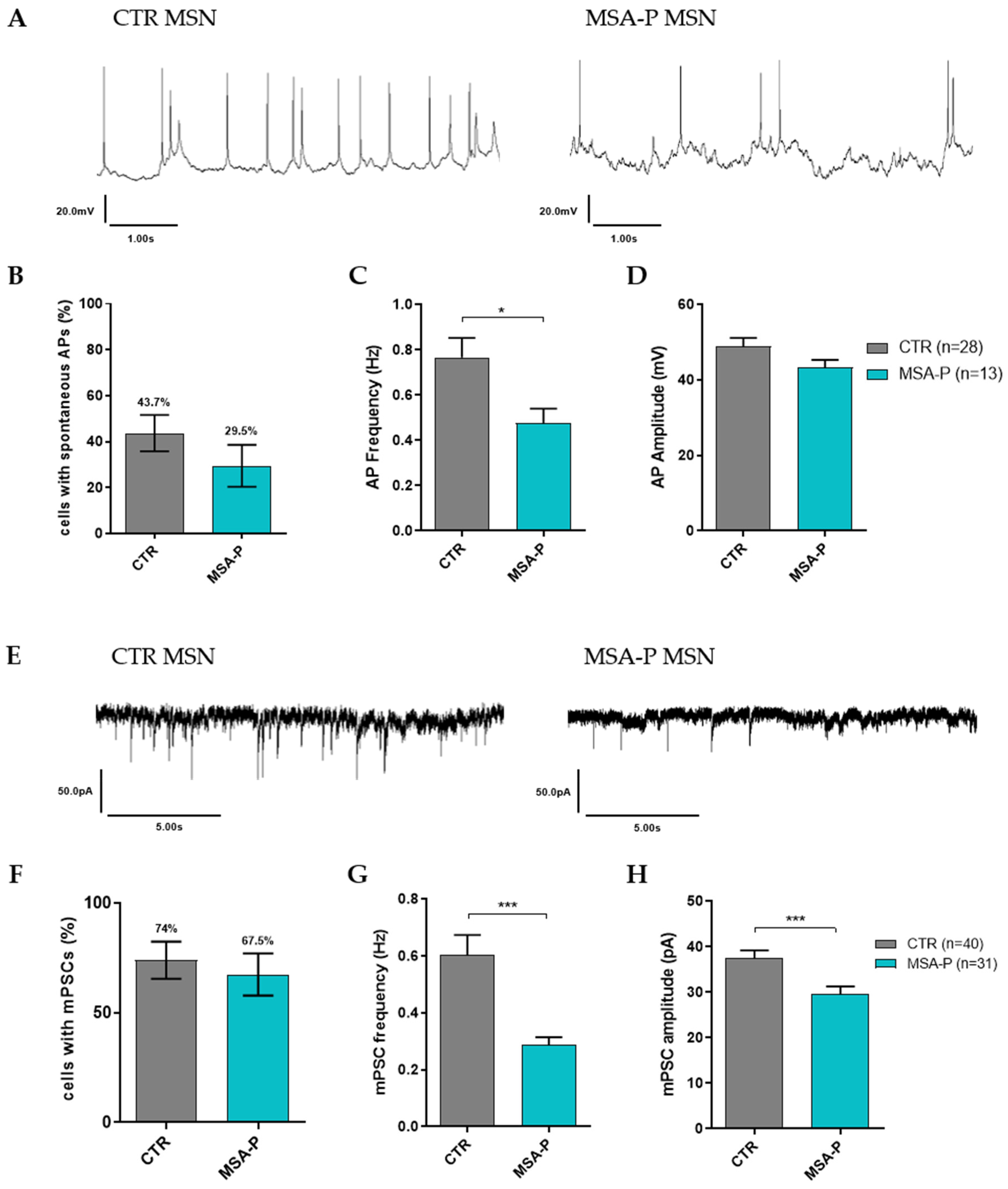
3.3.4. MSA-P MSNs Exhibit Reduced Spontaneous AP Frequency and Synaptic Activity
3.4. Expression of GABAA and GABAB Receptor Subunits, ATP-Dependent K+ Channels (Sulfonylurea Receptors), and Voltage-Gated Ca2+ Channels
3.4.1. Altered Expression of GABAA Receptor Subunits
3.4.2. Reduced Expression of GABAB Receptor Subunit 2
3.4.3. Comparable Expression of Voltage-Gated Ca2+ Channel Subtypes
3.5. α-Synuclein Characteristics in MSNs
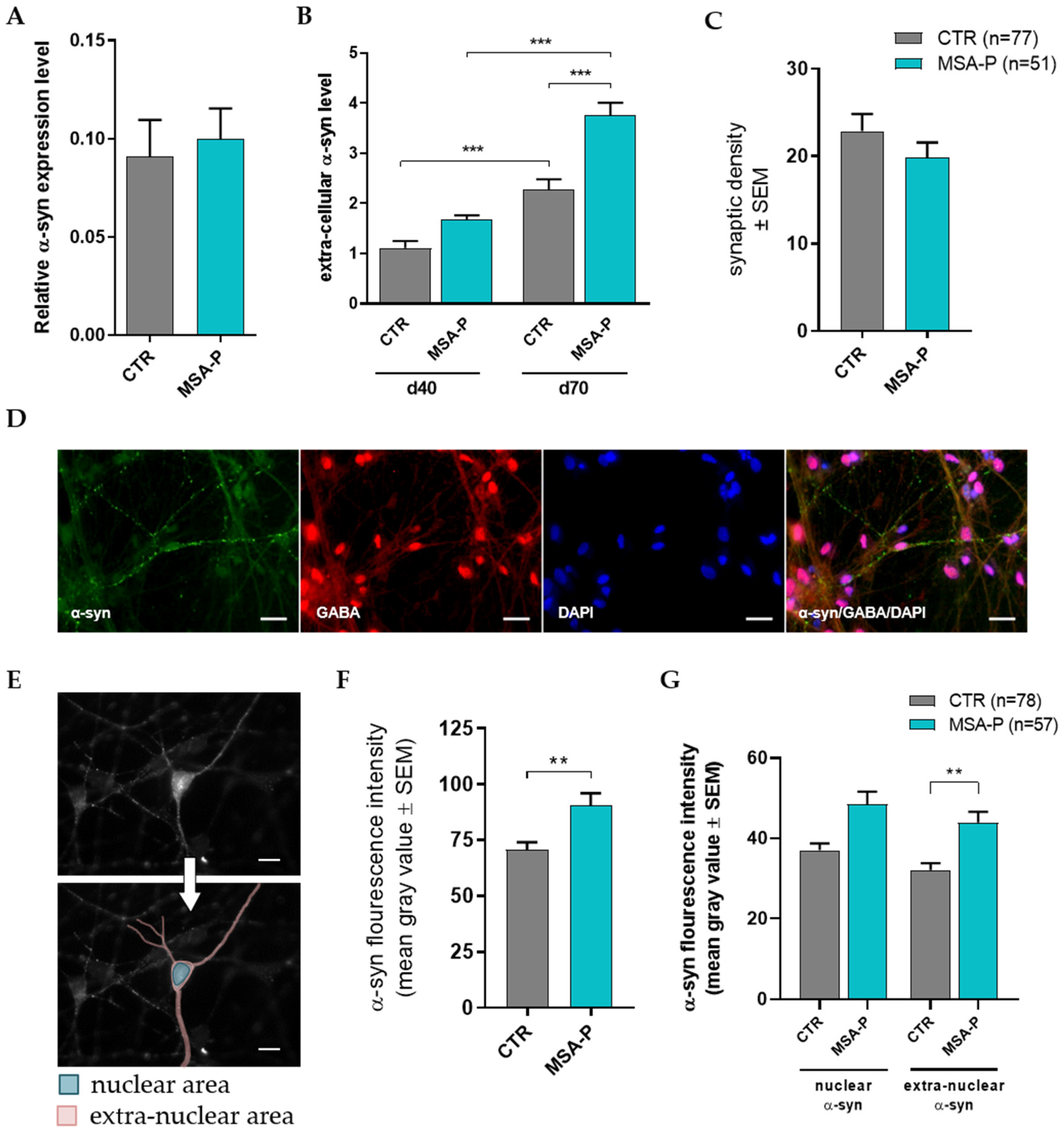
3.5.1. Elevated Release of α-Synuclein by Differentiated MSA-P MSNs
3.5.2. Unaltered GABAergic Synaptic Density
3.5.3. MSA-P MSNs Contain Significantly More α-Synuclein Than Control MSNs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gilman, S.; Wenning, G.; Low, P.A.; Brooks, D.; Mathias, C.J.; Trojanowski, J.Q.; Wood, N.; Colosimo, C.; Durr, A.; Fowler, C.J.; et al. Second consensus statement on the diagnosis of multiple system atrophy. Neurology 2008, 71, 670–676. [Google Scholar] [CrossRef]
- Wenning, G.K.; Stankovic, I.; Vignatelli, L.; Fanciulli, A.; Calandra-Buonaura, G.; Seppi, K.; Palma, J.; Meissner, W.G.; Krismer, F.; Berg, D.; et al. The Movement Disorder Society Criteria for the Diagnosis of Multiple System Atrophy. Mov. Disord. 2022, 37, 1131–1148. [Google Scholar] [CrossRef] [PubMed]
- Spillantini, M.; Growther, R.; Jakes, R.; Cairns, N.; Lantos, P.; Goedert, M. Filamentous a-synuclein inclusions link multiple system atrophy with Parkinson’s disease and dementia with Lewy bodies. Neurosci. Lett. 1998, 251, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, T.; Paviour, D.; Quinn, N.P.; Josephs, K.A.; Sangha, H.; Kilford, L.; Healy, D.G.; Wood, N.W.; Lees, A.J.; Holton, J.L.; et al. The spectrum of pathological involvement of the striatonigral and olivopontocerebellar systems in multiple system atrophy: Clinicopathological correlations. Brain 2004, 127, 2657–2671. [Google Scholar] [CrossRef] [Green Version]
- Cykowski, M.D.; Coon, E.A.; Powell, S.Z.; Jenkins, S.M.; Benarroch, E.E.; Low, P.A.; Schmeichel, A.M.; Parisi, J.E. Expanding the spectrum of neuronal pathology in multiple system atrophy. Brain 2015, 138, 2293–2309. [Google Scholar] [CrossRef]
- Salvesen, L.; Ullerup, B.H.; Sunay, F.B.; Brudek, T.; Løkkegaard, A.; Agander, T.K.; Winge, K.; Pakkenberg, B. Changes in total cell numbers of the basal ganglia in patients with multiple system atrophy—A stereological study. Neurobiol. Dis. 2015, 74, 104–113. [Google Scholar] [CrossRef]
- Goedert, M.; Masuda-Suzukake, M.; Falcon, B. Like prions: The propagation of aggregated tau and α-synuclein in neurodegeneration. Brain 2017, 140, 266–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasili, E.; Dominguez-Meijide, A.; Outeiro, T.F. Spreading of α-Synuclein and Tau: A Systematic Comparison of the Mechanisms Involved. Front. Mol. Neurosci. 2019, 12, 107. [Google Scholar] [CrossRef] [Green Version]
- Woerman, A.L.; Patel, S.; Kazmi, S.A.; Oehler, A.; Lee, J.; Mordes, D.A.; Olson, S.H.; Prusiner, S.B. Kinetics of α-synuclein prions preceding neuropathological inclusions in multiple system atrophy. PLoS Pathog. 2020, 16, e1008222. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Kaji, R.; Matsumoto, S.; Nagahiro, S.; Goto, S. Compartmental loss of striatal medium spiny neurons in multiple system atrophy of parkinsonian type. Mov. Disord. 2007, 22, 2365–2370. [Google Scholar] [CrossRef]
- Stanslowsky, N.; Reinhardt, P.; Glaß, H.; Kalmbach, N.; Naujock, M.; Hensel, N.; Lübben, V.; Pal, A.; Venneri, A.; Lupo, F.; et al. Neuronal Dysfunction in iPSC-Derived Medium Spiny Neurons from Chorea-Acanthocytosis Patients Is Reversed by Src Kinase Inhibition and F-Actin Stabilization. J. Neurosci. 2016, 36, 12027–12043. [Google Scholar] [CrossRef] [Green Version]
- Capetian, P.; Stanslowsky, N.; Bernhardi, E.; Grütz, K.; Domingo, A.; Brüggemann, N.; Naujock, M.; Seibler, P.; Klein, C.; Wegner, F. Altered glutamate response and calcium dynamics in iPSC-derived striatal neurons from XDP patients. Exp. Neurol. 2018, 308, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Compagnoni, G.M.; Kleiner, G.; Samarani, M.; Aureli, M.; Faustini, G.; Bellucci, A.; Ronchi, D.; Bordoni, A.; Garbellini, M.; Salani, S.; et al. Mitochondrial Dysregulation and Impaired Autophagy in iPSC-Derived Dopaminergic Neurons of Multiple System Atrophy. Stem Cell Rep. 2018, 11, 1185–1198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klein, T.; Henkel, L.; Klug, K.; Kwok, C.K.; Klopocki, E.; Üçeyler, N. Generation of the human induced pluripotent stem cell line UKWNLi002-A from dermal fibroblasts of a woman with a heterozygous c.608 CT (p.Thr203Met) mutation in exon 3 of the nerve growth factor gene potentially associated with hereditary sensory and autonomic neuropathy type 5. Stem Cell Res. 2018, 33, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef] [Green Version]
- Staege, S.; Kutschenko, A.; Baumann, H.; Glaß, H.; Henkel, L.; Gschwendtberger, T.; Kalmbach, N.; Klietz, M.; Hermann, A.; Lohmann, K.; et al. Reduced Expression of GABAA Receptor Alpha2 Subunit Is Associated With Disinhibition of DYT-THAP1 Dystonia Patient-Derived Striatal Medium Spiny Neurons. Front. Cell Dev. Biol. 2021, 9, 650586. [Google Scholar] [CrossRef]
- Hensel, N.; Schön, A.; Konen, T.; Lübben, V.; Förthmann, B.; Baron, O.; Grothe, C.; Leifheit-Nestler, M.; Claus, P.; Haffner, D. Fibroblast growth factor 23 signaling in hippocampal cells: Impact on neuronal morphology and synaptic density. J. Neurochem. 2016, 137, 756–769. [Google Scholar] [CrossRef] [Green Version]
- Meijering, E.; Jacob, M.; Sarria, J.-C.F.; Steiner, P.; Hirling, H.; Unser, M. Design and validation of a tool for neurite tracing and analysis in fluorescence microscopy images. Cytometry A 2004, 58, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Fred, A.; Gamboa, H. (Eds.) Biomedical Engeneering Systems and Technologies. In 9th International Joint Conference, BIOSTEC 2016, Rome, Italy, 21–23 February 2016, Revised Selected Papers; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Stanslowsky, N.; Haase, A.; Martin, U.; Naujock, M.; Leffler, A.; Dengler, R.; Wegner, F. Functional differentiation of midbrain neurons from human cord blood-derived induced pluripotent stem cells. Stem Cell Res. Ther. 2014, 5, 35. [Google Scholar] [CrossRef]
- Kutschenko, A.; Staege, S.; Grütz, K.; Glaß, H.; Kalmbach, N.; Gschwendtberger, T.; Henkel, L.; Heine, J.; Grünewald, A.; Hermann, A.; et al. Functional and Molecular Properties of DYT-SGCE Myoclonus-Dystonia Patient-Derived Striatal Medium Spiny Neurons. Int. J. Mol. Sci. 2021, 22, 3565. [Google Scholar] [CrossRef] [PubMed]
- Wegner, F.; Kraft, R.; Busse, K.; Härtig, W.; Schaarschmidt, G.; Schwarz, S.C.; Schwarz, J.; Hevers, W. Functional and molecular analysis of GABAA receptors in human midbrain-derived neural progenitor cells. J. Neurochem. 2008, 107, 1056–1069. [Google Scholar] [CrossRef] [PubMed]
- Wegner, F.; Kraft, R.; Busse, K.; Härtig, W.; Ahrens, J.; Leffler, A.; Dengler, R.; Schwarz, J. Differentiated human midbrain-derived neural progenitor cells express excitatory strychnine-sensitive glycine receptors containing α2β subunits. PLoS ONE 2012, 7, e36946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Precious, S.; Kelly, C.; Reddington, A.; Vinh, N.; Stickland, R.; Pekarik, V.; Scherf, C.; Jeyasingham, R.; Glasbey, J.; Holeiter, M.; et al. FoxP1 marks medium spiny neurons from precursors to maturity and is required for their differentiation. Exp. Neurol. 2016, 282, 9–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emmanouilidou, E.; Minakaki, G.; Keramioti, M.V.; Xylaki, M.; Balafas, E.; Chrysanthou-Piterou, M.; Kloukina, I.; Vekrellis, K. GABA transmission via ATP-dependent K+ channels regulates α-synuclein secretion in mouse striatum. Brain 2016, 139, 871–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azevedo, C.; Teku, G.; Pomeshchik, Y.; Reyes, J.F.; Chumarina, M.; Russ, K.; Savchenko, E.; Hammarberg, A.; Lamas, N.J.; Collin, A.; et al. Parkinson’s disease and multiple system atrophy patient iPSC-derived oligodendrocytes exhibit alpha-synuclein–induced changes in maturation and immune reactive properties. Proc. Natl. Acad. Sci. USA 2022, 119, e2111405119. [Google Scholar] [CrossRef]
- Tanudjojo, B.; Shaikh, S.S.; Fenyi, A.; Bousset, L.; Agarwal, D.; Marsh, J.; Zois, C.; Heman-Ackah, S.; Fischer, R.; Sims, D.; et al. Phenotypic manifestation of α-synuclein strains derived from Parkinson’s disease and multiple system atrophy in human dopaminergic neurons. Nat. Commun. 2021, 12, 3817. [Google Scholar] [CrossRef]
- Schapira, A.H. Calcium dysregulation in Parkinson’s disease. Brain 2013, 136, 2015–2016. [Google Scholar] [CrossRef] [Green Version]
- Schöndorf, D.C.; Aureli, M.; McAllister, F.E.; Hindley, C.J.; Mayer, F.; Schmid, B.; Sardi, S.P.; Valsecchi, M.; Hoffmann, S.; Schwarz, L.K.; et al. iPSC-derived neurons from GBA1-associated Parkinson’s disease patients show autophagic defects and impaired calcium homeostasis. Nat. Commun. 2014, 5, 4028. [Google Scholar] [CrossRef] [Green Version]
- Smith-Dijak, A.I.; Sepers, M.D.; Raymond, L.A. Alterations in synaptic function and plasticity in Huntington disease. J. Neurochem. 2019, 150, 346–365. [Google Scholar] [CrossRef]
- Marambaud, P.; Dreses-Werringloer, U.; Vingtdeux, V. Calcium signaling in neurodegeneration. Mol. Neurodegener. 2009, 4, 20. [Google Scholar] [CrossRef] [PubMed]
- Bezprozvanny, I.; Mattson, M.P. Neuronal calcium mishandling and the pathogenesis of Alzheimer’s disease. Trends Neurosci. 2008, 31, 454–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selkoe, D.J. Alzheimer’s disease: Genes, proteins, and therapy. Physiol. Rev. 2001, 81, 741–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zampese, E.; Surmeier, D.J. Calcium, Bioenergetics, and Parkinson’s Disease. Cells 2020, 9, 2045. [Google Scholar] [CrossRef]
- Ferreiro, E.; Resende, R.; Costa, R.; Oliveira, C.R.; Pereira, C.M. An endoplasmic-reticulum-specific apoptotic pathway is involved in prion and amyloid-beta peptides neurotoxicity. Neurobiol. Dis. 2006, 23, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, J.; Alvarez-Illera, P.; García-Casas, P.; Fonteriz, R.I.; Montero, M. The Role of Ca2+ Signaling in Aging and Neurodegeneration: Insights from Caenorhabditis elegans Models. Cells 2020, 9, 204. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Joshi, P.R.; Cepeda, C.; Masliah, E.; Levine, M.S. Alpha-synuclein overexpression in mice alters synaptic communication in the corticostriatal pathway. J. Neurosci. Res. 2009, 88, 1764–1776. [Google Scholar] [CrossRef] [Green Version]
- Cepeda, C.; Hurst, R.S.; Calvert, C.R.; Hernández-Echeagaray, E.; Nguyen, O.K.; Jocoy, E.; Christian, L.J.; Ariano, M.A.; Levine, M.S. Transient and progressive electrophysiological alterations in the corticostriatal pathway in a mouse model of Huntington’s disease. J. Neurosci. 2003, 23, 961–969. [Google Scholar] [CrossRef] [Green Version]
- Ben-Ari, Y. Excitatory actions of gaba during development: The nature of the nurture. Nat. Rev. Neurosci. 2002, 3, 728–739. [Google Scholar] [CrossRef]
- Obrietan, K.; Pol, A.N.V.D. GABA neurotransmission in the hypothalamus: Developmental reversal from Ca2+ elevating to depressing. J. Neurosci. 1995, 15, 5065–5077. [Google Scholar] [CrossRef]
- Pol, A.V.D.; Obrietan, K.; Cao, V.; Trombley, P. Embryonic hypothalamic expression of functional glutamate receptors. Neuroscience 1995, 67, 419–439. [Google Scholar] [CrossRef] [PubMed]
- Pol, A.N.V.D.; Gao, X.-B.; Patrylo, P.R.; Ghosh, P.K.; Obrietan, K. Glutamate Inhibits GABA Excitatory Activity in Developing Neurons. J. Neurosci. 1998, 18, 10749–10761. [Google Scholar] [CrossRef] [Green Version]
- Saint-Amant, L.; Drapeau, P. Motoneuron Activity Patterns Related to the Earliest Behavior of the Zebrafish Embryo. J. Neurosci. 2000, 20, 3964–3972. [Google Scholar] [CrossRef] [Green Version]
- Takayama, C.; Inoue, Y. GABAergic signaling in the developing cerebellum. Anat. Sci. Int. 2004, 79, 124–136. [Google Scholar] [CrossRef] [Green Version]
- Zafeiriou, M.-P.; Bao, G.; Hudson, J.; Halder, R.; Blenkle, A.; Schreiber, M.-K.; Fischer, A.; Schild, D.; Zimmermann, W.-H. Developmental GABA polarity switch and neuronal plasticity in Bioengineered Neuronal Organoids. Nat. Commun. 2020, 11, 3791. [Google Scholar] [CrossRef] [PubMed]
- Asi, Y.T.; Simpson, J.E.; Heath, P.R.; Wharton, S.B.; Lees, A.J.; Revesz, T.; Houlden, H.; Holton, J.L. Alpha-synuclein mRNA expression in oligodendrocytes in MSA. Glia 2014, 62, 964–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reyes, J.F.; Rey, N.L.; Bousset, L.; Melki, R.; Brundin, P.; Angot, E. Alpha-synuclein transfers from neurons to oligodendrocytes. Glia 2013, 62, 387–398. [Google Scholar] [CrossRef]
- Kisos, H.; Pukaß, K.; Ben-Hur, T.; Richter-Landsberg, C.; Sharon, R. Increased neuronal α-synuclein pathology associates with its accumulation in oligodendrocytes in mice modeling α-synucleinopathies. PLoS ONE 2012, 7, e46817. [Google Scholar] [CrossRef] [Green Version]
- Bonanno, G. Selective block of rat and human neocortex GABAB receptors regulating somatostatin release by a GABAB antagonist endowed with cognition enhancing activity. Neuropharmacology 1999, 38, 1789–1795. [Google Scholar] [CrossRef]
- Cattaneo, S.; Ripamonti, M.; Bedogni, F.; Sessa, A.; Taverna, S. Somatostatin-Expressing Interneurons Co-Release GABA and Glutamate onto Different Postsynaptic Targets in the Striatum. bioRxiv 2019. Preprint. Available online: https://www.biorxiv.org/content/10.1101/566984v2 (accessed on 2 January 2023). [CrossRef]
- Song, Y.-H.; Yoon, J.; Lee, S.-H. The role of neuropeptide somatostatin in the brain and its application in treating neurological disorders. Exp. Mol. Med. 2021, 53, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Martiros, N.; Burgess, A.A.; Graybiel, A.M. Inversely Active Striatal Projection Neurons and Interneurons Selectively Delimit Useful Behavioral Sequences. Curr. Biol. 2018, 28, 560–573.e5. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID Code | Diagnosis | Sex | Age at Biopsy | Previously Published |
|---|---|---|---|---|
| CTR1 | CTRL | M | 59 | newly generated |
| CTR2 | CTRL | M | 59 | StemBANCC consortium SFC084-03-02-01A |
| CTR3 | CTRL | F | 62 | newly generated |
| P1 | Probable MSA-P | F | 78 | [13] |
| P2 | Probable MSA-P | M | 52 | newly generated |
| P3 | Probable MSA-P | F | 56 | newly generated |
| Control | MSA-P | |||||||
|---|---|---|---|---|---|---|---|---|
| Functional Properties | CTR1 (n = 23) | CTR2 (n = 13) | CTR3 (n = 19) | CTR (n = 55) | P1 (n = 14) | P2 (n = 13) | P3 (n = 14) | MSA-P (n = 41) |
| INa max. amplitudes (pA/pF) | −70.64 ± 7.1 | −85.0 ± 8.9 | −97.3 ± 9.9 | −89.4 ± 9.1 | −80.39 ± 8.8 | −137.4 ± 14.7 | −105.1 ± 11.2 | −106.6 ± 11.3 |
| IK max. amplitudes (pA/pF) | 139.9 ± 14.6 | 126.9 ± 13.3 | 152.4 ± 15.8 | 147.4 ± 15.3 | 125.6 ± 13,5 | 163.2 ± 17.2 | 174.5 ± 18.4 | 154.2 ± 16.4 |
| Resting membrane potential (mV) | −29.6 ± 2.3 | −23.4 ± 2.4 | −36.8 ± 3.6 | −30.7 ± 1.8 | −25.8 ± 2.4 | −27.4 ± 1.8 | −26.9 ± 3.1 | −26.7 ± 1.4 |
| Membrane capacitance (pF) | 20.8 ± 2.8 | 21 ± 3.5 | 18.5 ± 2.3 | 20 ± 1.6 | 14.5 ± 2.3 | 19.1 ± 3.6 | 20 ± 3.3 | 17.8 ± 1.8 |
| Input resistance (MOhm) | 336.6 ± 44.4 | 407.1 ± 46.3 | 486.4 ± 91.3 | 406.2 ± 38.9 | 440.3 ± 80.1 | 363.8 ± 49.7 | 311.4 ± 50.6 | 373.6 ± 36.4 |
| Cells with evoked APs (%) | 81.3 ± 12 | 68.8 ± 18.8 | 82.4 ± 4 | 78.9 ± 6.2 | 87.5 ± 7.2 | 93.3 ± 6.7 | 82.2 ± 9.7 | 87.7 ± 4.3 |
| Cells with single evoked APs (%) | 70.8 ± 12.5 | 46.4 ± 3.6 | 56.9 ± 21.6 | 60.8 ± 8.8 | 91.7 ± 8.3 | 62.2 ± 23.2 | 47.2 ± 12.1 | 69.5 ± 9.5 |
| Cells with repetitively evoked APs (%) | 19.4 ± 6.7 | 53.6 ± 3.6 | 21.5 ± 6.2 | 39.2 ± 8.8 | 8.33 ± 8.33 | 37.8 ± 11.8 | 39.6 ± 12 | 30.5 ± 9.8 |
| Amplitude (mV) of evoked APs | 77.3 ± 3.6 | 85.5 ± 3.8 | 77.8 ± 3.4 | 79.2 ± 2.2 | 78.1 ± 3.2 | 84.7 ± 3.1 | 82.7 ± 2.7 | 81.8 ± 1.8 |
| Duration (ms) of evoked APs | 3.92 ± 0.45 | 2.83 ± 0.34 | 5.82 ± 1.21 | 4.39 ± 0.51 | 4.31 ± 0.42 | 3.38 ± 0.3 | 2.8 ± 0.23 | 3.52 ± 0.22 |
| AHP amplitude (mV) | 11.8 ± 1.4 | 9.6 ± 1.9 | 12.8 ± 2 | 11.7 ± 1 | 9.1 ± 0.66 | 12.9 ± 1.4 | 14.1 ± 1.6 | 12.1 ± 0.84 |
| Time to peak AHP (ms) | 17.5 ± 3.1 | 19.4 ± 4.3 | 18.3 ± 2.2 | 18.2 ± 1.7 | 19.1 ± 3.6 | 16 ± 2.9 | 17.5 ± 3.1 | 17.5 ± 1.8 |
| Cells with spontaneous APs (%) | 48.5 ± 14.7 | 29.2 ± 18.2 | 51.9 ± 1.9 | 43.7 ± 7.8 | 18.8 ± 18.8 | 31.1 ± 17.4 | 42.2 ± 8.9 | 29.5 ± 9.1 |
| Frequency of spontaneous APs (Hz) | 0.77 ± 0.15 | 0.6 ± 0.17 | 0.84 ± 0.12 | 0.76 ± 0.09 | 0.33 ± 0.11 | 0.76 ± 0.15 | 0.35 ± 0.05 | 0.48 ± 0.06 * |
| Amplitude of spontaneous APs (mV) | 45.2 ± 3.5 | 47.8 ± 4 | 53.8 ± 3.9 | 48.8 ± 2.3 | 28.57 ± 6.5 | 42.8 ± 3 | 46.5 ± 2.5 | 43.3 ± 2 |
| Cells with miniature PSCs (%) | 88.8 ± 6.6 | 87.5 ± 7.2 | 40.7 ± 12.1 | 74 ± 8.5 | 56.3 ± 21.4 | 73.3 ± 17.6 | 76.7 ± 5.1 | 67.5 ± 9.6 |
| Miniature PSC frequencies (Hz) | 0.59 ± 0.08 | 0.45 ± 0.05 | 0.88 ± 0.3 | 0.6 ± 0.07 | 0.17 ± 0.02 | 0.43 ± 0.08 | 0.31 ± 0.03 | 0.29 ± 0.03 *** |
| Miniature PSC amplitudes (pA) | 42.4 ± 3.3 | 29.4 ± 1.6 | 38.9 ± 2.8 | 37.4 ± 1.8 | 22.7 ± 2.7 | 28.4 ± 2.5 | 37.9 ± 2.9 | 29.5 ± 1.7 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henkel, L.M.; Kankowski, S.; Moellenkamp, T.M.; Smandzich, N.J.; Schwarz, S.; Di Fonzo, A.; Göhring, G.; Höglinger, G.; Wegner, F. iPSC-Derived Striatal Medium Spiny Neurons from Patients with Multiple System Atrophy Show Hypoexcitability and Elevated α-Synuclein Release. Cells 2023, 12, 223. https://doi.org/10.3390/cells12020223
Henkel LM, Kankowski S, Moellenkamp TM, Smandzich NJ, Schwarz S, Di Fonzo A, Göhring G, Höglinger G, Wegner F. iPSC-Derived Striatal Medium Spiny Neurons from Patients with Multiple System Atrophy Show Hypoexcitability and Elevated α-Synuclein Release. Cells. 2023; 12(2):223. https://doi.org/10.3390/cells12020223
Chicago/Turabian StyleHenkel, Lisa M., Svenja Kankowski, Thiemo M. Moellenkamp, Nadine J. Smandzich, Sigrid Schwarz, Alessio Di Fonzo, Gudrun Göhring, Günter Höglinger, and Florian Wegner. 2023. "iPSC-Derived Striatal Medium Spiny Neurons from Patients with Multiple System Atrophy Show Hypoexcitability and Elevated α-Synuclein Release" Cells 12, no. 2: 223. https://doi.org/10.3390/cells12020223