
Dynamic 3D Modeling for Human Sperm Motility through the Female Cervical Canal and Uterine Cavity to Predict Sperm Chance of Reaching the Oocyte

Abstract
:1. Introduction
2. Methods and Materials
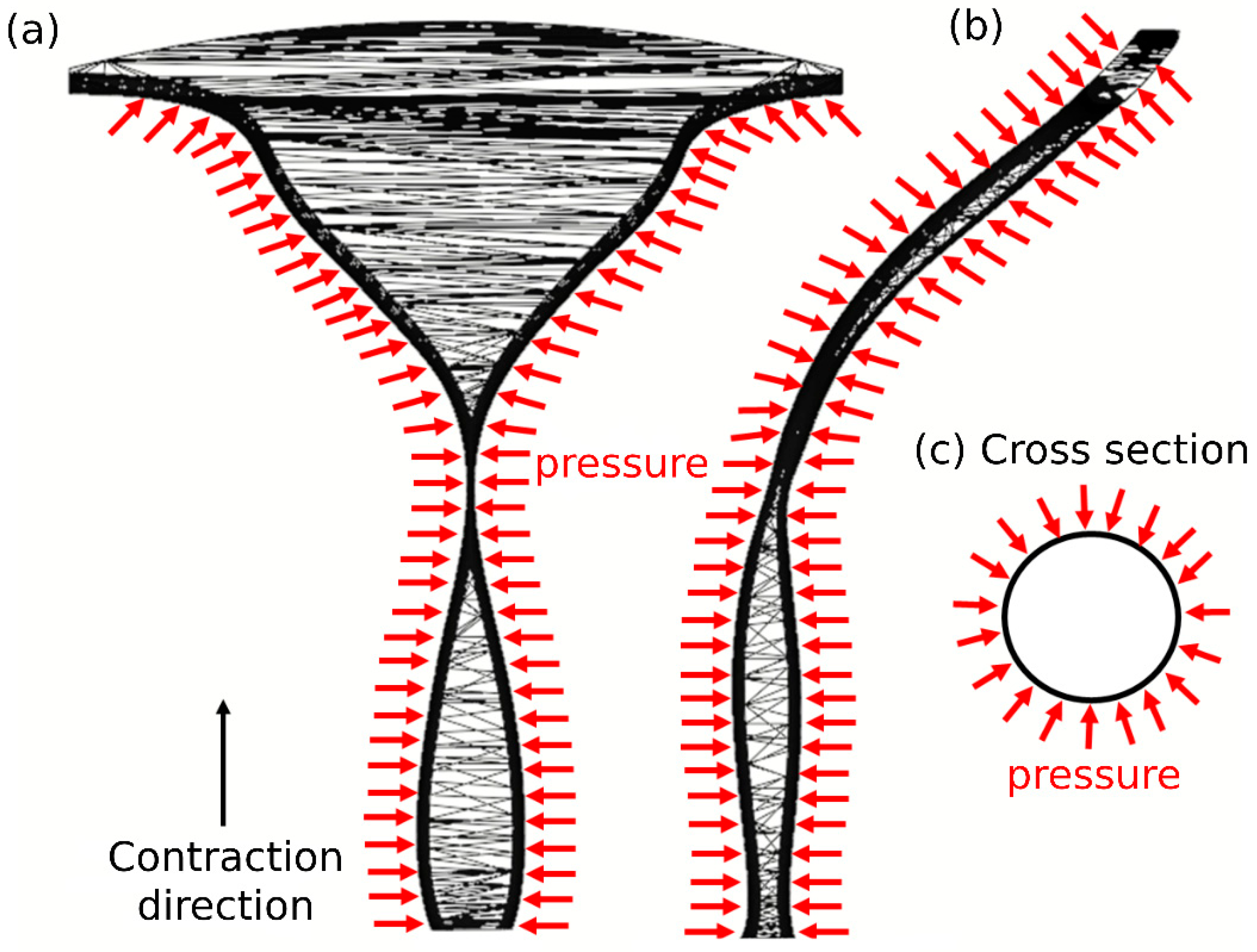
2.1. 3D Model of Human Female Cervical Canal and Uterine Cavity
2.2. Uterine Peristalsis
2.3. 3D Reconstruction of the Human Sperm Models
2.4. Beat Pattern
2.5. Computerized Fluid Dynamic
2.6. Dynamic Simulations
2.7. Outcome Measurements
- The 3D swimming trajectories of the different models were analyzed for position maps, calculated by SolidWorks 2022 as mentioned above. The localized displacements that developed in the sperm head centroid of the normal and pathological models were determined as a function of time. The outcome measures included the following:
- The 3D swimming trajectories of the sperm head centroid of the normal and pathological sperm models across the 3D human female cervical canal and uterine cavity model over 5 h. To ensure that an adequate sample of semen is available and contains the healthiest sperm with the highest chances for success, the recommendation is to perform IVF no later than one and a half hours from ejaculation. We decided to extend the simulation to five hours to make sure we covered a larger dynamic range over time.
- The number of normal and pathological sperm models passing through the cervical canal and reaching the fallopian tube site.
- The positions of normal and pathological sperm models inside the 3D cervical and uterine cavity model as a function of swimming time.
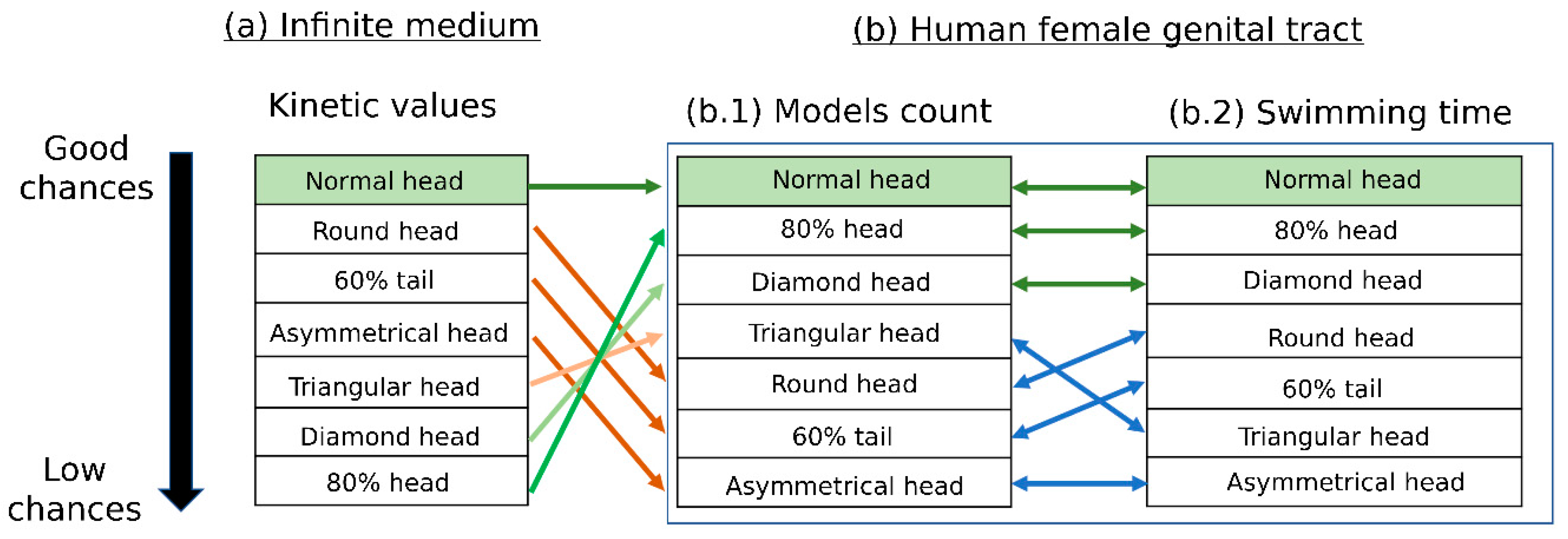
- Scoring the various sperm cell pathologies according to their estimated success rates of reaching the oocyte and comparing them with the scoring results of the infinite medium sperm simulation performed in [19].
3. Results
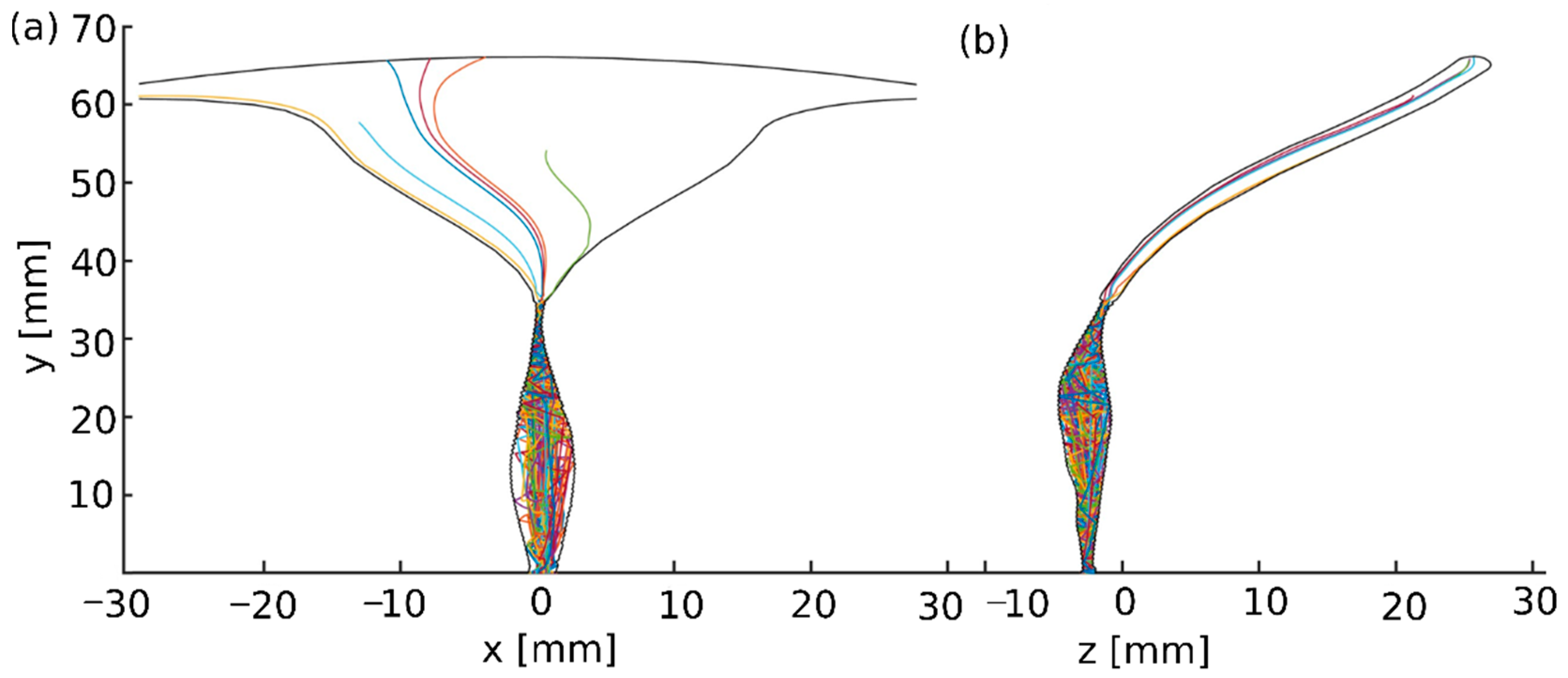
3.1. 3D Swimming Trajectories of the Sperm Models through the 3D Tract Model
3.2. Scoring of Sperm Pathologies Based on Their Positions in the 3D Tract Model
3.3. Scoring the 3D Swimming Patterns of Normal Sperm Models as a Function of Time
3.4. Scoring of Sperm Pathologies Based on Their Swimming Time in the 3D Tract Model
3.5. Scoring of Sperm Pathologies—Summary
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reynaud, K.; Schuss, Z.; Rouach, N.; Holcman, D.; Reynaud, K.; Schuss, Z.; Rouach, N.; Holcman, D.; Reynaud, K.; Schuss, Z.; et al. Why so Many Sperm Cells? Commun. Integr. Biol. 2015, 8, e1017156. [Google Scholar] [CrossRef] [PubMed]
- Schwalm, H.; Dubrauszky, V. The Structure of the Musculature of the Human Uterus—Muscles and Connective Tissue. Am. J. Obstet. Gynecol. 1966, 94, 391–404. [Google Scholar] [CrossRef]
- Rorie, D.K.; Newton, M. Histologic and Chemical Studies of the Smooth Muscle in the Human Cervix and Uterus. Am. J. Obstet. Gynecol. 1967, 99, 466–469. [Google Scholar] [CrossRef] [PubMed]
- Ludmir, J.; Sehdev, H.M. Anatomy and Physiology of the Uterine Cervix. Clin. Obstet. Gynecol. 2000, 43, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Aspden, R.M. Collagen Organisation in the Cervix and Its Relation to Mechanical Function. Coll. Relat. Res. 1988, 8, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Baah-Dwomoh, A.; McGuire, J.; Tan, T.; De Vita, R. Mechanical Properties of Female Reproductive Organs and Supporting Connective Tissues: A Review of the Current State of Knowledge. Appl. Mech. Rev. 2016, 68, 060801. [Google Scholar] [CrossRef] [Green Version]
- Katz, D.F.; Slade, D.A.; Nakajima, S.T. Analysis of Pre-Ovulatory Changes in Cervical Mucus Hydration and Sperm Penetrability. Adv. Contracept. 1997, 13, 143–151. [Google Scholar] [CrossRef]
- Mullins, K.J.; Saacke, R.G. Study of the Functional Anatomy of Bovine Cervical Mucosa with Special Reference to Mucus Secretion and Sperm Transport. Reprod. Biol. 1989, 117, 106–117. [Google Scholar] [CrossRef]
- Tung, C.; Hu, L.; Fiore, A.G.; Ardon, F.; Hickman, D.G.; Gilbert, R.O. Microgrooves and Fluid Flows Provide Preferential Passageways for Sperm over Pathogen Tritrichomonas Foetus. Proc. Natl. Acad. Sci. USA 2015, 112, 5431–5436. [Google Scholar] [CrossRef] [Green Version]
- Weiss, S.; Jaermann, T.; Schmid, P.; Staempfli, P.; Boesiger, P.; Niederer, P.; Caduff, R.; Bajka, M. Three-Dimensional Fiber Architecture of the Nonpregnant Human Uterus Determined Ex Vivo Using Magnetic Resonance Diffusion Tensor Imaging. Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. 2006, 90, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Hawk, H.W. Transport and Fate of Spermatozoa After Insemination of Cattle. J. Dairy Sci. 1986, 70, 1487–1503. [Google Scholar] [CrossRef] [PubMed]
- van Gestel, I.; Ijland, M.M.; Hoogland, H.J.; Evers, J.L.H. Endometrial Wave-like Activity in the Non-Pregnant Uterus. Hum. Reprod. Update 2003, 9, 131–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartoov, B.; Eltes, F.; Langsam, J.; Snyder, M.; Fisher, J. Ultrastructural Studies in Morphological Assessment of Human Spermatozoa. Int. J. Androl. 1982, 5, 81–96. [Google Scholar] [CrossRef]
- Freundl, G.; Grimm, H.J.; Hofmann, N. Selective Filtration of Abnormal Spermatozoa by the Cervical Mucus. Hum. Reprod. 1988, 3, 277–280. [Google Scholar] [CrossRef] [PubMed]
- Fredricsson, B.; Björk, G. Morphology of Postcoital Spermatozoa in the Cervical Secretion and Its Clinical Significance. Fertil. Steril. 1977, 28, 841–845. [Google Scholar] [CrossRef]
- Jeulin, C.; Soumah, A.; Jouannet, P. Morphological Factors Influencing the Penetration of Human Sperm into Cervical Mucus in Vitro. Int. J. Androl. 1985, 8, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, D.; Leslie, E.E.; Kelly, R.W.; Templeton, A.A. Morphological Selection of Human Spermatozoa in Vivo and in Vitro. J. Reprod. Fertil. 1982, 64, 391–399. [Google Scholar] [CrossRef] [Green Version]
- Geboren, J.K. The in Vitro Spermatozoal Penetration Test in Fertility Investigations. Ph.D. Thesis, University of Groningen, Groningen, The Netherlands, 1968. [Google Scholar]
- Nassir, M.; Levi, M.; Dardikman-Yoffe, G.; Mirsky, S.K.; Shaked, N.T. Prediction of Sperm Progression in Three Dimensions Using Rapid Optical Imaging and Dynamic Mechanical Modeling. Cells 2022, 11, 1319. [Google Scholar] [CrossRef]
- Dardikman-Yoffe, G.; Mirsky, S.K.; Barnea, I.; Shaked, N.T. High-Resolution 4-D Acquisition of Freely Swimming Human Sperm Cells without Staining. Sci. Adv. 2020, 6, eaay7619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. WHO Laboratory Manual for the Examination and Processing of Human Semen, 6th ed.; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Merz, E.; Miric-Tesanic, D.; Bahlmann, F.; Weber, G.; Wellek, S. Sonographic Size of Uterus and Ovaries in Pre- and Postmenopausal Women. Ultrasound Obstet. Gynecol. 1996, 7, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Kelsey, T.W.; Ginbey, E.; Chowdhury, M.M.; Bath, L.E.; Anderson, R.A.; Wallace, W.H.B. A Validated Normative Model for Human Uterine Volume from Birth to Age 40 Years. PLoS ONE 2016, 11, e0157375. [Google Scholar] [CrossRef] [Green Version]
- Reusch, L.M.; Feltovich, H.; Carlson, L.C.; Hall, G.; Campagnola, P.J.; Eliceiri, K.W.; Hall, T.J. Nonlinear Optical Microscopy and Ultrasound Imaging of Human Cervical Structure. J. Biomed. Opt. 2013, 18, 031110. [Google Scholar] [CrossRef] [PubMed]
- Lathrop, G.P. Growth of the Uterus. Arch. Dis. Child. 2017, 2, 314–330. [Google Scholar] [CrossRef]
- Da Costa, A.G.; Filho, F.M.; Ferreira, A.C.; Spara, P.; Mauad, F.M. Uterine Volume in Adolescents. Ultrasound Med. Biol. 2004, 30, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Coad, J.E.; Fugett, J.H.; Boykin, C.; Bennett, H.E.; Castrodale, D.; Castillo-Saenz, L.; Garza-Leal, J. Uterine Cavity Dimensions and Myometrial Thicknesses: Important Safety Considerations for Intrauterine Procedures. J. Minim. Invasive Gynecol. 2017, 24, S144. [Google Scholar] [CrossRef]
- Aguilar, H.N.; Mitchell, B.F. Physiological Pathways and Molecular Mechanisms Regulating Uterine Contractility. Hum. Reprod. Update 2010, 16, 725–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Ziegler, D.; Brioschi, P.A.; Fanchin, R.; Bulletti, C.; Ayoubi, J.M. Contractility of the Non-Pregnant Uterus. Infertil. Reprod. Med. Clin. N. Am. 2003, 14, 309–327. [Google Scholar] [CrossRef]
- Hendricks, C.H. A New Technique for the Study of Motility in the Non-pregnant Human Uterus. BJOG Int. J. Obstet. Gynaecol. 1964, 71, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Kuijsters, N.P.M.; Methorst, W.G.; Kortenhorst, M.S.Q.; Rabotti, C.; Mischi, M.; Schoot, B.C. Uterine Peristalsis and Fertility: Current Knowledge and Future Perspectives: A Review and Meta-Analysis. Reprod. Biomed. Online 2017, 35, 50–71. [Google Scholar] [CrossRef] [Green Version]
- Singh, G.; Chanda, A. Mechanical Properties of Whole-Body Soft Human Tissues: A Review. Biomed. Mater. 2021, 16, 62004. [Google Scholar] [CrossRef]
- Satir, P.; Matsuoka, T. Splitting the Ciliary Axoneme: Implications for a “switch-Point” Model of Dynein Arm Activity in Ciliary Motion. Cell Motil. Cytoskeleton 1989, 14, 345–358. [Google Scholar] [CrossRef]
- Satir, P.; Heuser, T.; Sale, W.S. A Structural Basis for How Motile Cilia Beat. Bioscience 2014, 64, 1073–1083. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Nicastro, D. Asymmetric Distribution and Spatial Switching of Dynein Activity Generates Ciliary Motility. Science (80-). 2018, 360, eaar1968. [Google Scholar] [CrossRef] [Green Version]
- Ishibashi, K.; Sakakibara, H.; Oiwa, K. Force-Generating Mechanism of Axonemal Dynein in Solo and Ensemble. Int. J. Mol. Sci. 2020, 21, 2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez, S.S.; Pacey, A.A. Sperm Transport in the Female Reproductive Tract. Hum. Reprod. Update 2006, 12, 23–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suarez, S.S.; Ho, H. Hyperactivated Motility in Sperm. Reprod. Domest. Anim. 2003, 124, 119–124. [Google Scholar] [CrossRef]
- Suarez, S.S.; Ho, H.C. Hyperactivation of Mammalian Sperm. Cell. Mol. Biol. 2003, 49, 351–356. [Google Scholar]
- Sobachkin, A.; Dumnov, G. Numerical Basis of CAD-Embedded CFD. NAFEMS World Congr. 2013, 2013, 1–20. [Google Scholar]
- Chenoweth, P.J. Genetic Sperm Defects. Theriogenology 2005, 64, 457–468. [Google Scholar] [CrossRef]
- Sartori, P.; Geyer, V.F.; Scholich, A.; Jülicher, F.; Howard, J. Dynamic Curvature Regulation Accounts for the Symmetric and Asymmetric Beats of Chlamydomonas Flagella. eLife 2016, 5, e13258. [Google Scholar] [CrossRef]
- Su, T.W.; Xue, L.; Ozcan, A. High-Throughput Lensfree 3D Tracking of Human Sperms Reveals Rare Statistics of Helical Trajectories. Proc. Natl. Acad. Sci. USA 2012, 109, 16018–16022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafforgue, O.; Bouguerra, N.; Poncet, S.; Seyssiecq, I.; Favier, J.; Elkoun, S. Thermo-Physical Properties of Synthetic Mucus for the Study of Airway Clearance. J. Biomed. Mater. Res.-Part A 2017, 105, 3025–3033. [Google Scholar] [CrossRef] [PubMed]
- Lai, S.K.; O’Hanlon, D.E.; Harrold, S.; Man, S.T.; Wang, Y.Y.; Cone, R.; Hanes, J. Rapid Transport of Large Polymeric Nanoparticles in Fresh Undiluted Human Mucus. Proc. Natl. Acad. Sci. USA 2007, 104, 1482–1487. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pass the Cervical Canal | Reach the Fallopian Tube Sites | |
|---|---|---|
| Normal models | 27% | 12% |
| Diamond head models | 17% | 3% |
| 80% head models | 15% | 4% |
| Triangular head models | 11% | 4% |
| Asymmetrical head models | 6% | 0% |
| 60% tail models | 6% | 1% |
| Round head models | 4% | 2% |
| Pass the Cervical Canal [h] | Reaching the Fallopian Tube Sites [h] | |
|---|---|---|
| Normal models | 0.44 | 1.28 |
| Diamond head models | 0.65 | 1.86 |
| 80% head models | 0.8 | 1.95 |
| 60% tail models | 0.9 | 2.5 |
| Triangular head models | 0.92 | 2.72 |
| Round head models | 1.9 | 3.35 |
| Asymmetrical head models | 2.17 | Did not reach the fallopian tube site |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nassir, M.; Levi, M.; Shaked, N.T. Dynamic 3D Modeling for Human Sperm Motility through the Female Cervical Canal and Uterine Cavity to Predict Sperm Chance of Reaching the Oocyte. Cells 2023, 12, 203. https://doi.org/10.3390/cells12010203
Nassir M, Levi M, Shaked NT. Dynamic 3D Modeling for Human Sperm Motility through the Female Cervical Canal and Uterine Cavity to Predict Sperm Chance of Reaching the Oocyte. Cells. 2023; 12(1):203. https://doi.org/10.3390/cells12010203
Chicago/Turabian StyleNassir, Mayssam, Mattan Levi, and Natan T. Shaked. 2023. "Dynamic 3D Modeling for Human Sperm Motility through the Female Cervical Canal and Uterine Cavity to Predict Sperm Chance of Reaching the Oocyte" Cells 12, no. 1: 203. https://doi.org/10.3390/cells12010203