
Cross-Talk between CB1, AT1, AT2 and Mas Receptors Responsible for Blood Pressure Control in the Paraventricular Nucleus of Hypothalamus in Conscious Spontaneously Hypertensive Rats and Their Normotensive Controls

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Placement of Cannula for PVN Microinjection
2.3. Placement of Catheters for BP Measurements and Injections
2.4. Microinjections
2.5. Experimental Protocol
2.6. PVN Tissue Microdissection
2.7. Western Blot Analysis
2.8. Data Analysis
2.9. Drugs
3. Results
3.1. General
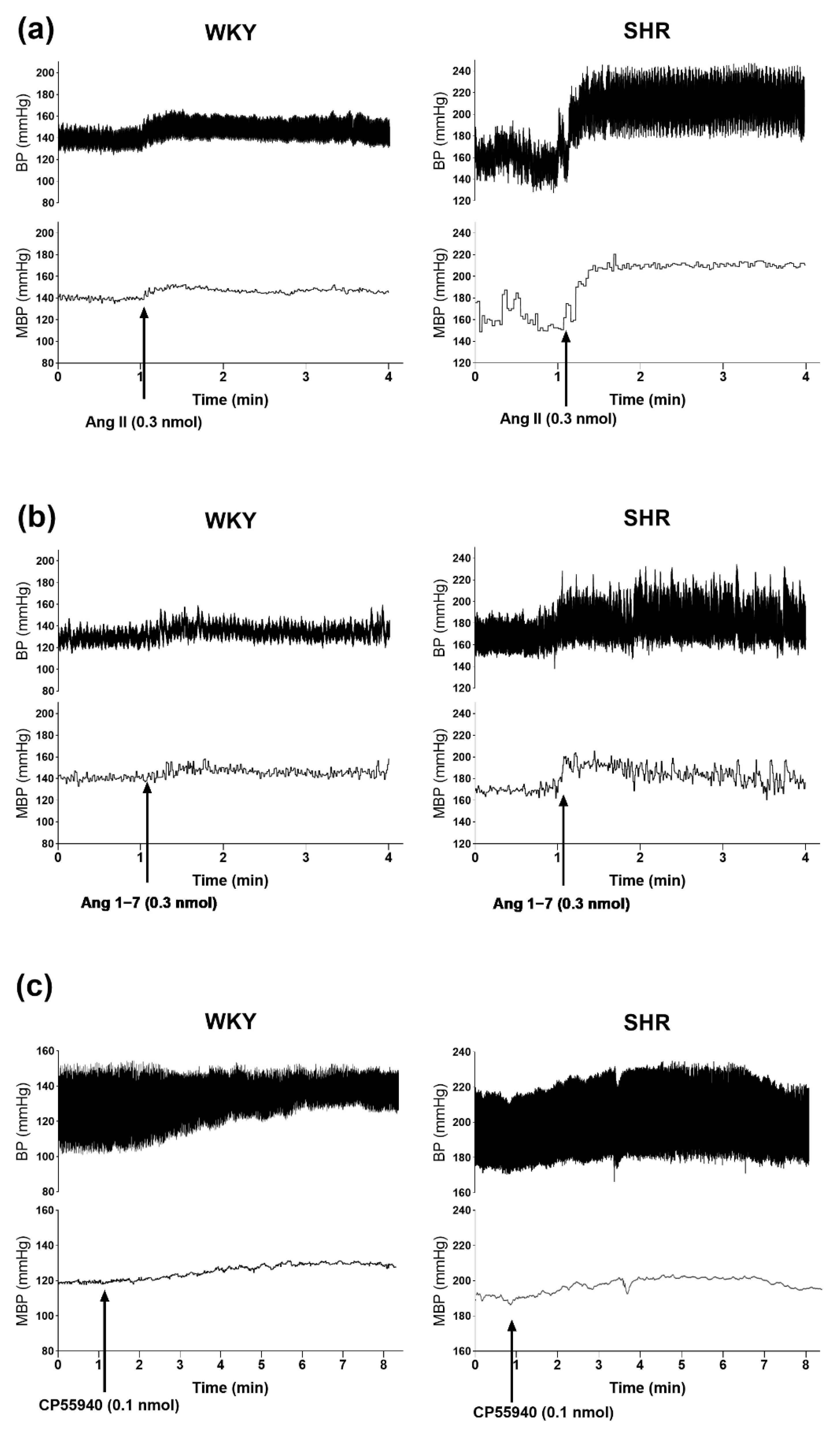
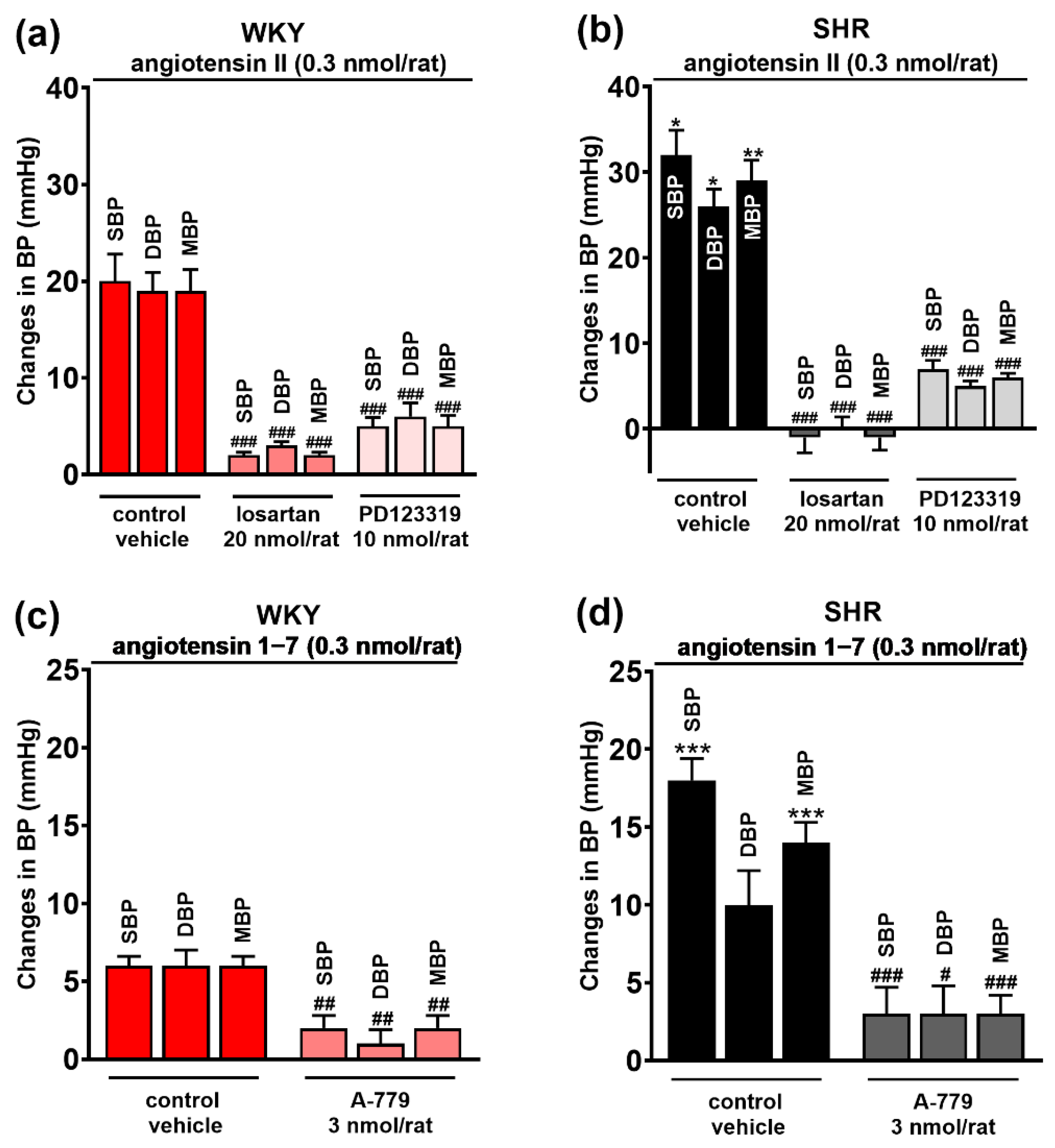
3.2. Involvement of AT1, AT2 and Mas Receptors in the Pressor Responses of Ang II and Ang 1–7 Microinjected into PVN
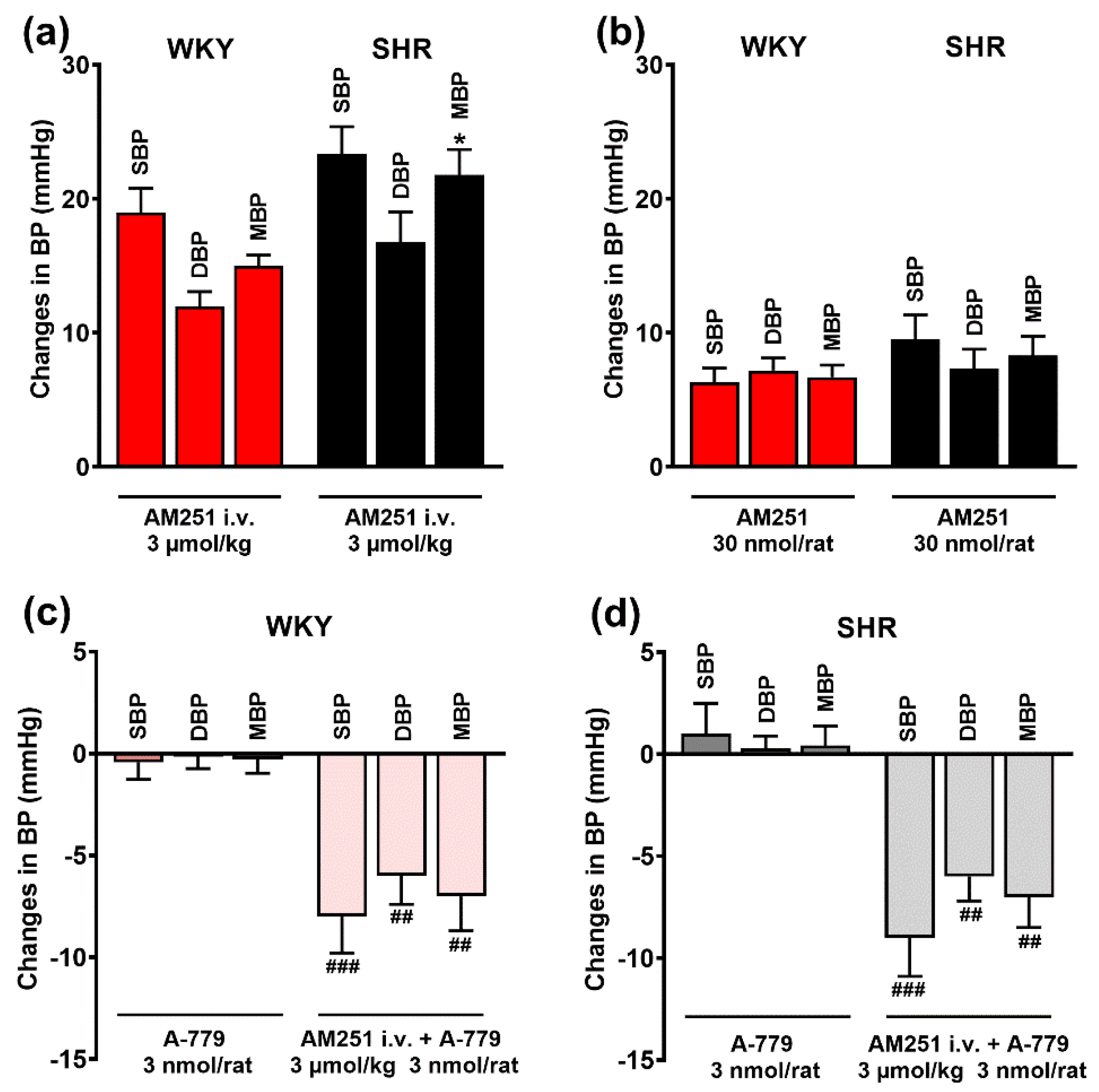
3.3. Involvement of CB1 Receptors in the Pressor Effects of Ang II and Ang 1–7 Microinjected into the PVN
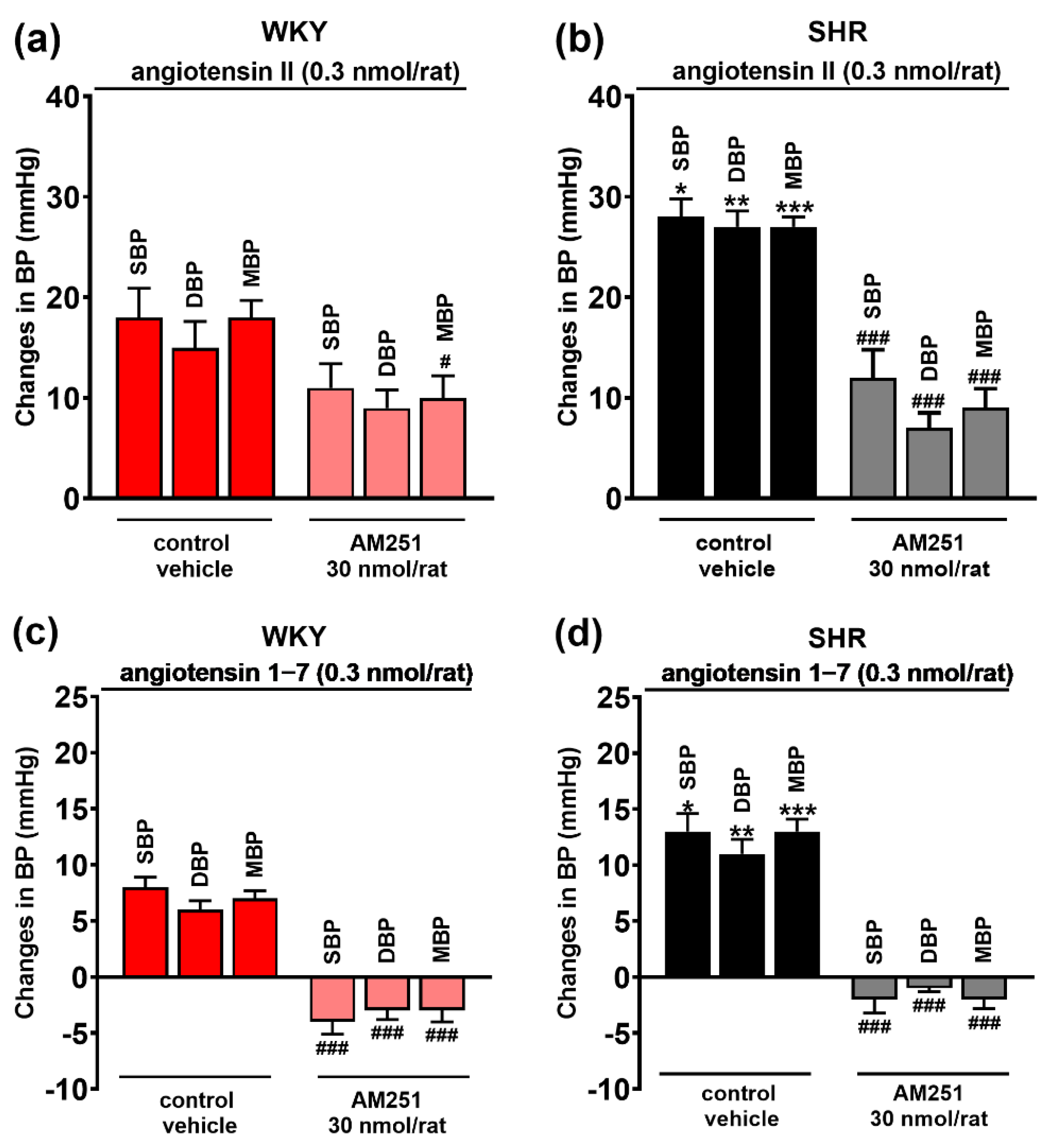
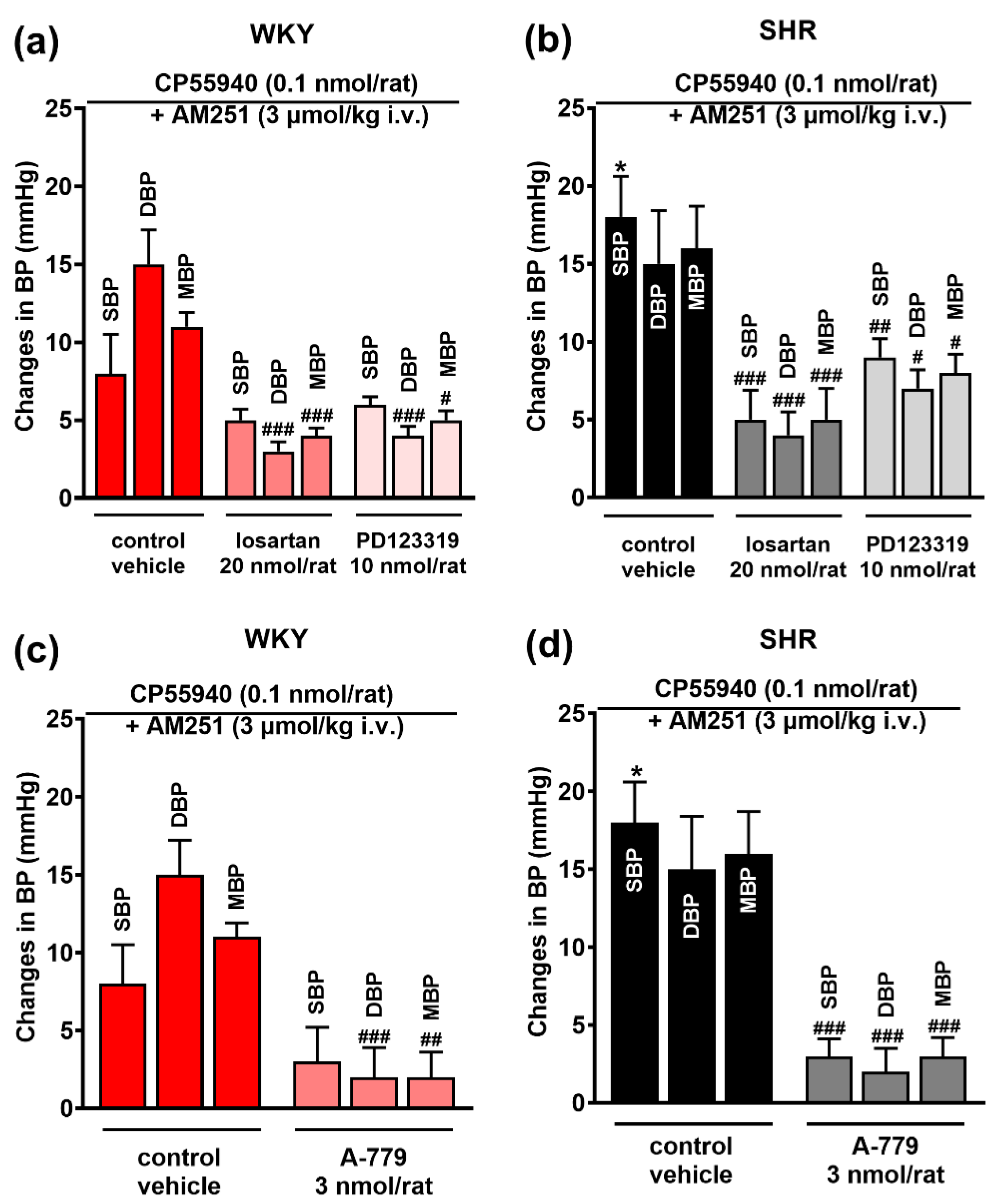
3.4. Involvement of AT1, AT2 and Mas Receptors in the Pressor Effect of the CB1 Receptor Agonist CP55940 Microinjected into the PVN
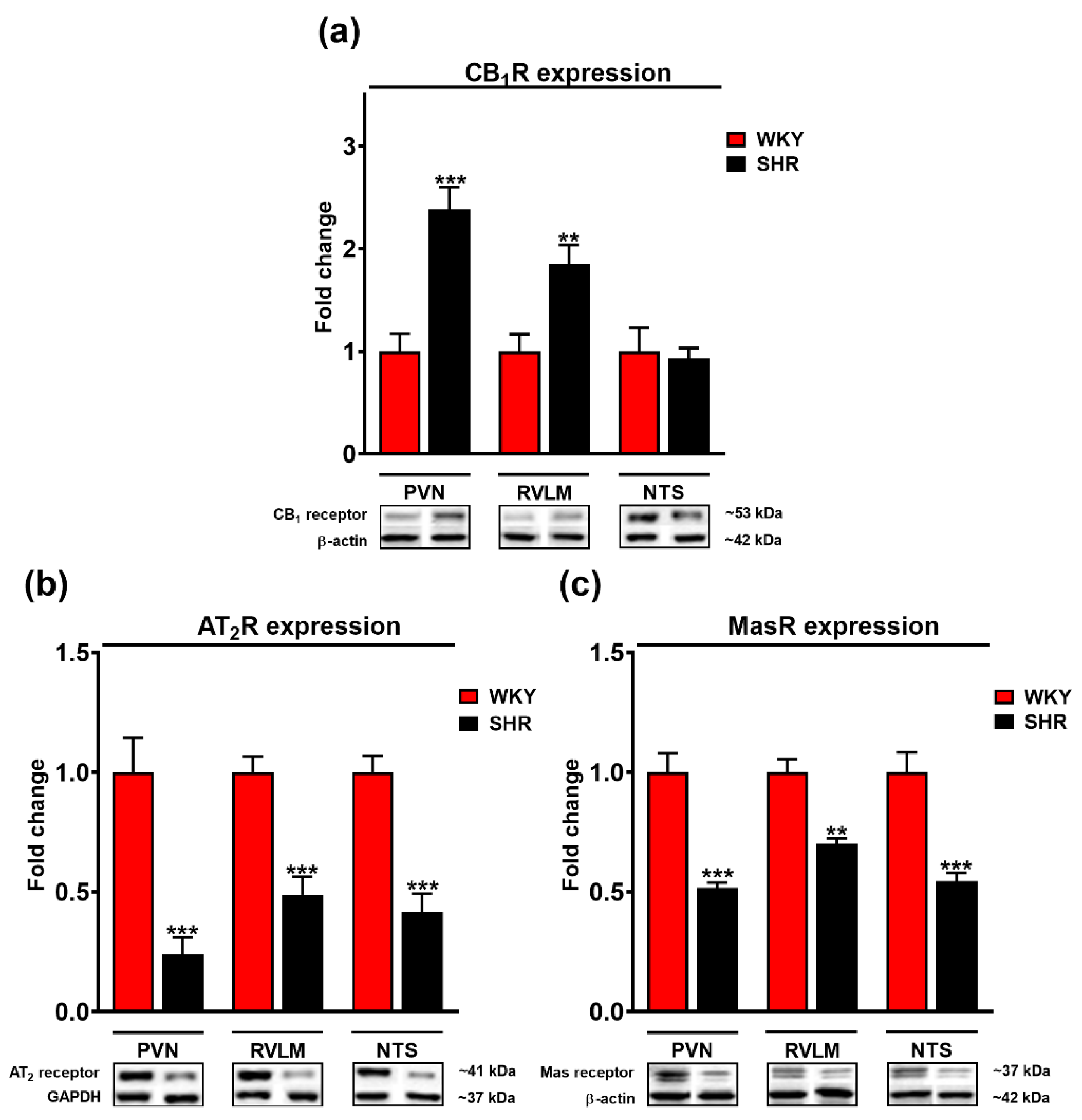
3.5. Comparison of Cannabinoid CB1, Mas, and AT2 Receptor Expression in the PVN, RVLM and NTS of WKY and SHR Rats
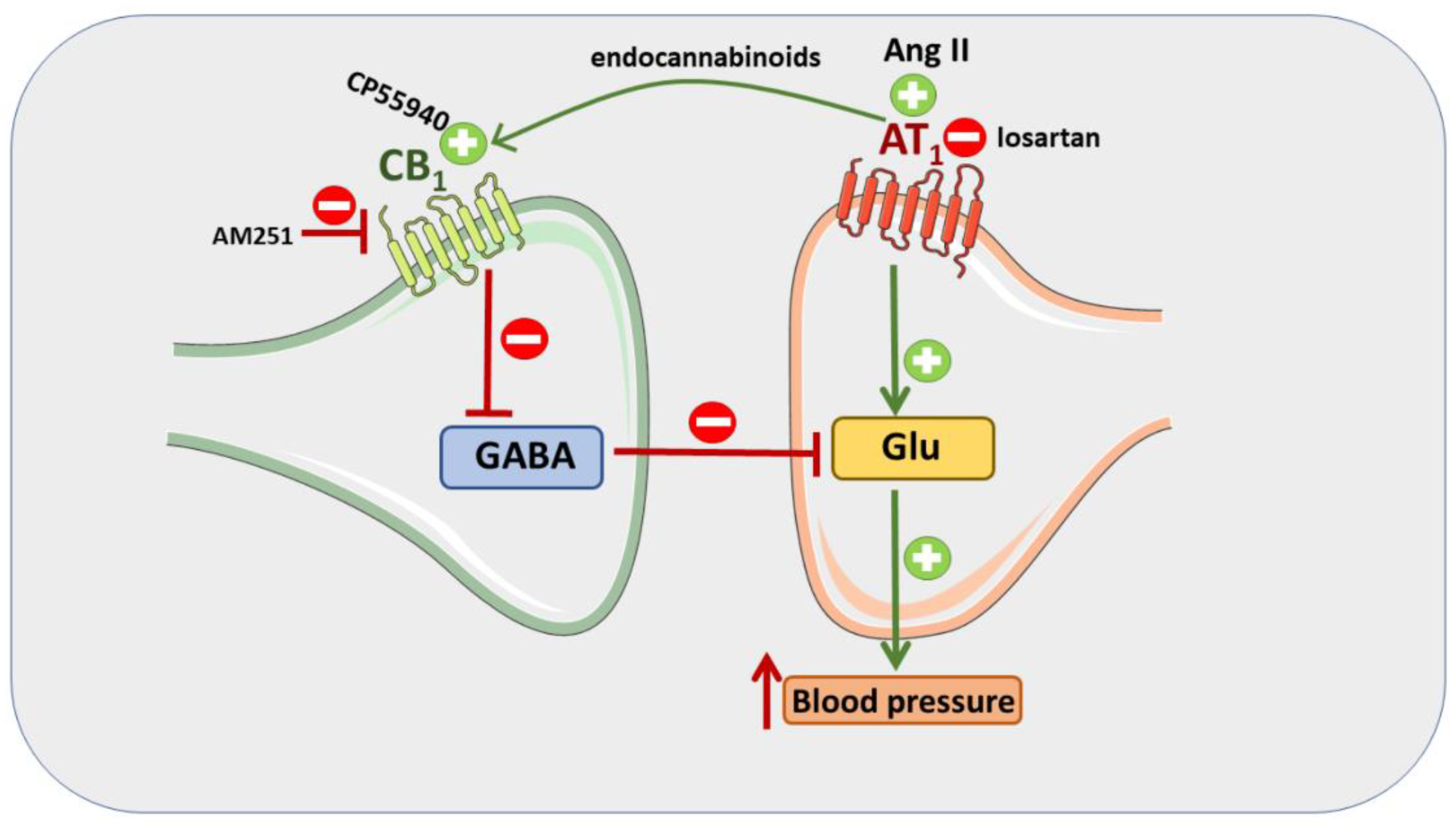
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goncharuk, V.D. The hypothalamus and its role in hypertension. Handb. Clin. Neurol. 2021, 182, 333–354. [Google Scholar] [CrossRef] [PubMed]
- Saxena, T.; Ali, A.O.; Saxena, M. Pathophysiology of essential hypertension: An update. Expert. Rev. Cardiovasc. Ther. 2018, 16, 879–887. [Google Scholar] [CrossRef] [PubMed]
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E., Jr.; Collins, K.J.; Dennison Himmelfarb, C.; DePalma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W.; et al. ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the Prevention, Detection, Evaluation, and Management of High Blood Pressure in Adults: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 2018, 138, e484–e594. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.J.; Ma, H.J.; Shao, J.Y.; Pan, H.L.; Li, D.P. Impaired Hypothalamic Regulation of Sympathetic Outflow in Primary Hypertension. Neurosci. Bull. 2019, 35, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.A.S.; Oudit, G.Y.; Verano-Braga, T.; Canta, G.; Steckelings, U.M.; Bader, M. The renin-angiotensin system: Going beyond the classical paradigms. Am. J. Physiol. Heart Circ. Physiol. 2019, 316, H958–H970. [Google Scholar] [CrossRef]
- Miller, A.J.; Arnold, A.C. The renin-angiotensin system in cardiovascular autonomic control: Recent developments and clinical implications. Clin. Auton. Res. 2019, 29, 231–243. [Google Scholar] [CrossRef]
- Vargas Vargas, R.A.; Varela Millán, J.M.; Fajardo Bonilla, E. Renin-angiotensin system: Basic and clinical aspects-A general perspective. Endocrinol. Diabetes Nutr. 2022, 69, 52–62. [Google Scholar] [CrossRef]
- Williams, B.; Mancia, G.; Spiering, W.; Rosei, E.A.; Azizi, M.; Burnier, M.; Clement, D.L.; Coca, A.; de Simone, G.; Dominiczak, A.; et al. Linee guida ESC/ESH 2018 per la diagnosi e il trattamento dell’ipertensione arteriosa. Task Force per la Diagnosi e il Trattamento dell’Ipertensione Arteriosa della Società Europea di Cardiologia (ESC) e della Società Europea dell’Ipertensione Arteriosa (ESH) [2018 ESC/ESH Guidelines for the management of arterial hypertension. The Task Force for the management of arterial hypertension of the European Society of Cardiology (ESC) and the European Society of Hypertension (ESH)]. G. Ital. Cardiol. 2006, 19, 3S–73S. [Google Scholar] [CrossRef]
- Mohsin, M.; Souza, L.A.C.; Aliabadi, S.; Worker, C.J.; Cooper, S.G.; Afrin, S.; Murata, Y.; Xiong, Z.; Feng Earley, Y. Increased (Pro)renin Receptor Expression in the Hypertensive Human Brain. Front. Physiol. 2020, 11, 606811. [Google Scholar] [CrossRef]
- Chen, A.D.; Zhang, S.J.; Yuan, N.; Xu, Y.; De, W.; Gao, X.Y.; Zhu, G.Q. Angiotensin AT1 receptors in paraventricular nucleus contribute to sympathetic activation and enhanced cardiac sympathetic afferent reflex in renovascular hypertensive rats. Exp. Physiol. 2011, 96, 94–103. [Google Scholar] [CrossRef]
- Sun, H.J.; Li, P.; Chen, W.W.; Xiong, X.Q.; Han, Y. Angiotensin II and angiotensin-(1-7) in paraventricular nucleus modulate cardiac sympathetic afferent reflex in renovascular hypertensive rats. PLoS ONE 2012, 7, e52557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reja, V.; Goodchild, A.K.; Phillips, J.K.; Pilowsky, P.M. Upregulation of angiotensin AT1 receptor and intracellular kinase gene expression in hypertensive rats. Clin. Exp. Pharmacol. Physiol. 2006, 33, 690–695. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Chen, S.R.; Chen, H.; Pan, H.L. Endogenous AT1 receptor-protein kinase C activity in the hypothalamus augments glutamatergic input and sympathetic outflow in hypertension. J. Physiol. 2019, 597, 4325–4340. [Google Scholar] [CrossRef] [PubMed]
- Fan, Z.D.; Zhang, L.; Shi, Z.; Gan, X.B.; Gao, X.Y.; Zhu, G.Q. Artificial microRNA interference targeting AT(1a) receptors in paraventricular nucleus attenuates hypertension in rats. Gene Ther. 2012, 19, 810–817. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Sun, H.J.; Li, P.; Gao, Q.; Zhou, Y.B.; Zhang, F.; Gao, X.Y.; Zhu, G.Q. Angiotensin-(1-7) in paraventricular nucleus modulates sympathetic activity and cardiac sympathetic afferent reflex in renovascular hypertensive rats. PLoS ONE 2012, 7, e48966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, X.J.; Miao, Y.W.; Li, H.B.; Su, Q.; Liu, K.L.; Fu, L.Y.; Hou, Y.K.; Shi, X.L.; Li, Y.; Mu, J.J.; et al. Blockade of Endogenous Angiotensin-(1-7) in Hypothalamic Paraventricular Nucleus Attenuates High Salt-Induced Sympathoexcitation and Hypertension. Neurosci. Bull. 2019, 35, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Malinowska, B.; Baranowska-Kuczko, M.; Schlicker, E. Triphasic blood pressure responses to cannabinoids: Do we understand the mechanism. Br. J. Pharmacol. 2012, 16 5, 2073–2088. [Google Scholar] [CrossRef] [Green Version]
- Malinowska, B.; Toczek, M.; Pędzińska-Betiuk, A.; Schlicker, E. Cannabinoids in arterial, pulmonary and portal hypertension—Mechanisms of action and potential therapeutic significance. Br. J. Pharmacol. 2019, 176, 1395–1411. [Google Scholar] [CrossRef]
- Grzeda, E.; Schlicker, E.; Luczaj, W.; Harasim, E.; Baranowska-Kuczko, M.; Malinowska, B. Bi-directional CB1 receptor-mediated cardiovascular effects of cannabinoids in anaesthetized rats: Role of the paraventricular nucleus. J. Physiol. Pharmacol. 2015, 66, 343–353. [Google Scholar]
- Grzęda, E.; Schlicker, E.; Toczek, M.; Zalewska, I.; Baranowska-Kuczko, M.; Malinowska, B. CB1 receptor activation in the rat paraventricular nucleus induces bi-directional cardiovascular effects via modification of glutamatergic and GABAergic neurotransmission. Naunyn Schmiedebergs Arch. Pharmacol. 2017, 390, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Gyombolai, P.; Pap, D.; Turu, G.; Catt, K.J.; Bagdy, G.; Hunyady, L. Regulation of endocannabinoid release by G proteins: A paracrine mechanism of G protein-coupled receptor action. Mol. Cell. Endocrinol. 2012, 353, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosnihan, K.B.; Pulgar, V.M.; Gallagher, P.E.; Neves, L.A.; Yamaleyeva, L.M. Local uterine Ang-(1-7) infusion augments the expression of cannabinoid receptors and differentially alters endocannabinoid metabolizing enzymes in the decidualized uterus of pseudopregnant rats. Reprod. Biol. Endocrinol. 2015, 13, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerman, L.O.; Kurtz, T.W.; Touyz, R.M.; Ellison, D.H.; Chade, A.R.; Crowley, S.D.; Mattson, D.L.; Mullins, J.J.; Osborn, J.; Eirin, A.; et al. Animal Models of Hypertension: A Scientific Statement from the American Heart Association. Hypertension 2019, 73, e87–e120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Elsevier Academic Press: London, UK, 2007; ISBN 978-0-12-374121-9. [Google Scholar]
- Chen, Q.H.; Toney, G.M. Responses to GABA-A receptor blockade in the hypothalamic PVN are attenuated by local AT1 receptor antagonism. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 285, R1231–R1239. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G.; Howlett, A.C.; Abood, M.E.; Alexander, S.P.; Di Marzo, V.; Elphick, M.R.; Greasley, P.J.; Hansen, H.S.; Kunos, G.; Mackie, K.; et al. International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid receptors and their ligands: Beyond CB₁ and CB₂. Pharmacol. Rev. 2010, 62, 588–631. [Google Scholar] [CrossRef] [Green Version]
- Dupont, A.G.; Légat, L. GABA is a mediator of brain AT1 and AT2 receptor-mediated blood pressure responses. Hypertens. Res. 2020, 43, 995–1005. [Google Scholar] [CrossRef]
- Camargo, L.A.; Saad, W.A.; Simões, S.; Santos, T.A.; Saad, W.A. Interaction between paraventricular nucleus and septal area in the control of physiological responses induced by angiotensin II. Braz. J. Med. Biol. Res. 2002, 35, 1017–1023. [Google Scholar] [CrossRef] [Green Version]
- Khanmoradi, M.; Nasimi, A. Functions of AT1 and AT2 angiotensin receptors in the paraventricular nucleus of the rat, correlating single-unit and cardiovascular responses. Brain Res. Bull. 2017, 132, 170–179. [Google Scholar] [CrossRef]
- Elsaafien, K.; de Kloet, A.D.; Krause, E.G.; Sumners, C. Brain Angiotensin Type-1 and Type-2 Receptors in Physiological and Hypertensive Conditions: Focus on Neuroinflammation. Curr. Hypertens. Rep. 2020, 22, s11906–s020. [Google Scholar] [CrossRef]
- Dampney, R.A.; Michelini, L.C.; Li, D.P.; Pan, H.L. Regulation of sympathetic vasomotor activity by the hypothalamic paraventricular nucleus in normotensive and hypertensive states. Am. J. Physiol. Heart Circ. Physiol. 2018, 315, H1200–H1214. [Google Scholar] [CrossRef] [Green Version]
- Senst, L.; Bains, J. Neuromodulators, stress and plasticity: A role for endocannabinoid signalling. J. Exp. Biol. 2014, 217, 102–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, G.C.; Tse, J.; Hsu, Y.H.; Ho, C.Y.; Tseng, C.J.; Cheng, P.W. μ-Opioid Receptor-Mediated AT1R-TLR4 Crosstalk Promotes Microglial Activation to Modulate Blood Pressure Control in the Central Nervous System. Antioxidants 2021, 10, 1784. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Li, G.Q.; Zhang, H.P.; Zhang, Y.; Li, Q. Overactivation of cannabinoid receptor type 1 in rostral ventrolateral medulla promotes cardiovascular responses in spontaneously hypertensive rats. J. Hypertens. 2017, 35, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhang, H.; Le, K.D.; Chao, J.; Gao, L. Activation of central angiotensin type 2 receptors suppresses norepinephrine excretion and blood pressure in conscious rats. Am. J. Hypertens. 2011, 24, 724–730. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Liu, X.; Patel, K.P. Angiotensin-converting enzyme 2 overexpression improves central nitric oxide-mediated sympathetic outflow in chronic heart failure. Am. J. Physiol. Heart. Circ. Physiol. 2011, 301, H2402–H2412. [Google Scholar] [CrossRef] [Green Version]
- Nasimi, A.; Haddad, F.; Mirzaei-Damabi, N.; Rostami, B.; Hatam, M. Another controller system for arterial pressure. AngII-vasopressin neural network of the parvocellular paraventricular nucleus may regulate arterial pressure during hypotension. Brain Res. 2021, 1769, 147618. [Google Scholar] [CrossRef]
- Kwolek, G.; Zakrzeska, A.; Schlicker, E.; Göthert, M.; Godlewski, G.; Malinowska, B. Central and peripheral components of the pressor effect of anandamide in urethane-anaesthetized rats. Br. J. Pharmacol. 2005, 145, 567–575. [Google Scholar] [CrossRef] [Green Version]
- Turu, G.; Simon, A.; Gyombolai, P.; Szidonya, L.; Bagdy, G.; Lenkei, Z.; Hunyady, L. The role of diacylglycerol lipase in constitutive and angiotensin AT1 receptor-stimulated cannabinoid CB1 receptor activity. J. Biol. Chem. 2007, 282, 7753–7757. [Google Scholar] [CrossRef] [Green Version]
- Karpińska, O.; Baranowska-Kuczko, M.; Kloza, M.; Kozłowska, H. Endocannabinoids modulate Gq/11 protein-coupled receptor agonist-induced vasoconstriction via a negative feedback mechanism. J. Pharm. Pharmacol. 2018, 70, 214–222. [Google Scholar] [CrossRef] [Green Version]
- Miklós, Z.; Wafa, D.; Nádasy, G.L.; Tóth, Z.E.; Besztercei, B.; Dörnyei, G.; Laska, Z.; Benyó, Z.; Ivanics, T.; Hunyady, L.; et al. Angiotensin II-Induced Cardiac Effects Are Modulated by Endocannabinoid-Mediated CB1 Receptor Activation. Cells 2021, 10, 724. [Google Scholar] [CrossRef]
- Malinowska, B.; Godlewski, G.; Bucher, B.; Schlicker, E. Cannabinoid CB1 receptor-mediated inhibition of the neurogenic vasopressor response in the pithed rat. Naunyn Schmiedebergs Arch. Pharmacol. 1997, 356, 197–202. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mińczuk, K.; Schlicker, E.; Malinowska, B. Cross-Talk between CB1, AT1, AT2 and Mas Receptors Responsible for Blood Pressure Control in the Paraventricular Nucleus of Hypothalamus in Conscious Spontaneously Hypertensive Rats and Their Normotensive Controls. Cells 2022, 11, 1542. https://doi.org/10.3390/cells11091542
Mińczuk K, Schlicker E, Malinowska B. Cross-Talk between CB1, AT1, AT2 and Mas Receptors Responsible for Blood Pressure Control in the Paraventricular Nucleus of Hypothalamus in Conscious Spontaneously Hypertensive Rats and Their Normotensive Controls. Cells. 2022; 11(9):1542. https://doi.org/10.3390/cells11091542
Chicago/Turabian StyleMińczuk, Krzysztof, Eberhard Schlicker, and Barbara Malinowska. 2022. "Cross-Talk between CB1, AT1, AT2 and Mas Receptors Responsible for Blood Pressure Control in the Paraventricular Nucleus of Hypothalamus in Conscious Spontaneously Hypertensive Rats and Their Normotensive Controls" Cells 11, no. 9: 1542. https://doi.org/10.3390/cells11091542