
Physiological Function during Exercise and Environmental Stress in Humans—An Integrative View of Body Systems and Homeostasis



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Brain Function during Exercise and Environmental Stress
2.1. Exercise Responses
2.2. Impact of Adding Thermal Stress to Exercise
2.3. The Control of Brain Blood Flow during Exercise, with and without Environmental Stress
3. Lung Function during Exercise and Environmental Stress
3.1. Exercise Responses
3.2. Impact of Adding Environmental Stress to Exercise
3.3. Breathing Mechanics and Control of Breathing during Exercise, with and without Environmental Stress
4. Heart Function during Exercise and Environmental Stress
4.1. Exercise Responses
4.2. Impact of Adding Environmental Stress to Exercise
4.3. Cardiac and Peripheral Mechanisms Regulating the Systemic Circulation
5. Skeletal Muscle Function during Exercise and Environmental Stress
5.1. Exercise Responses
5.2. Impact of Adding Thermal Stress to Exercise
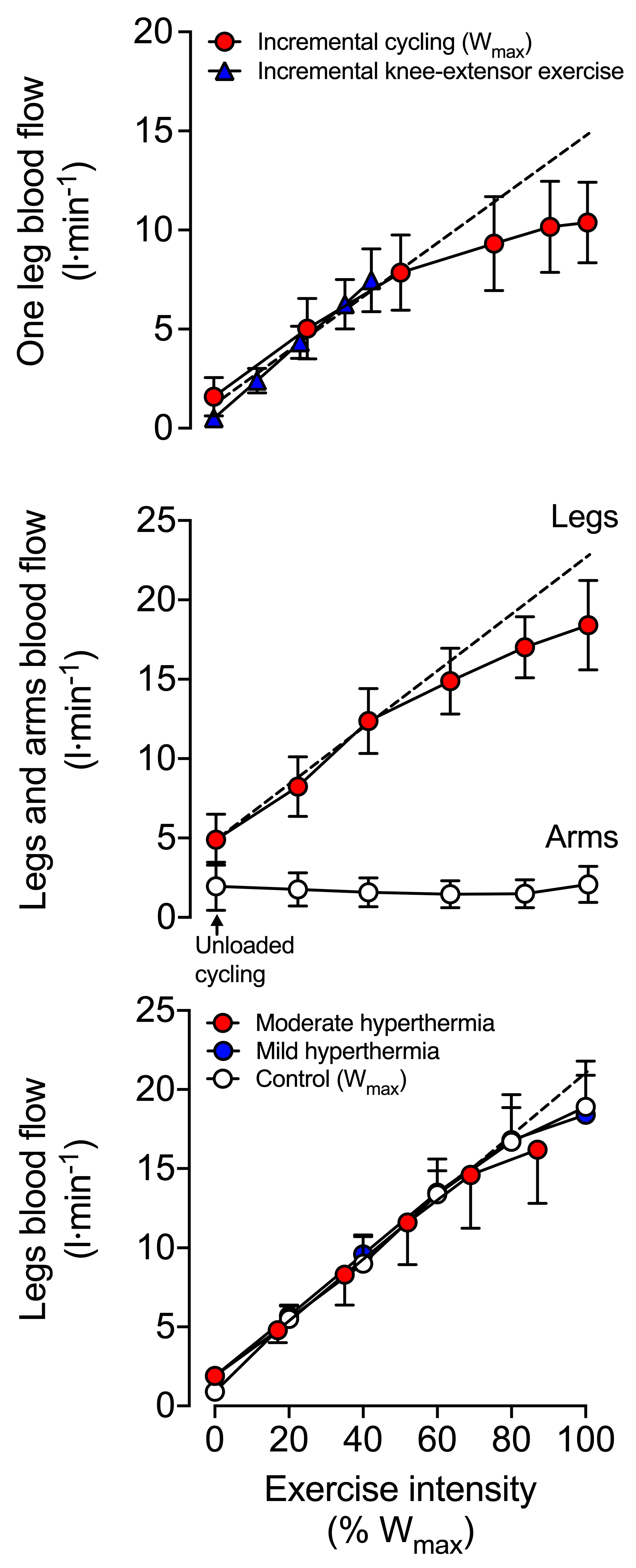
5.3. Mechanisms of Blood Flow Control—Impact of Exercise, Hyperthermia and Dehydration
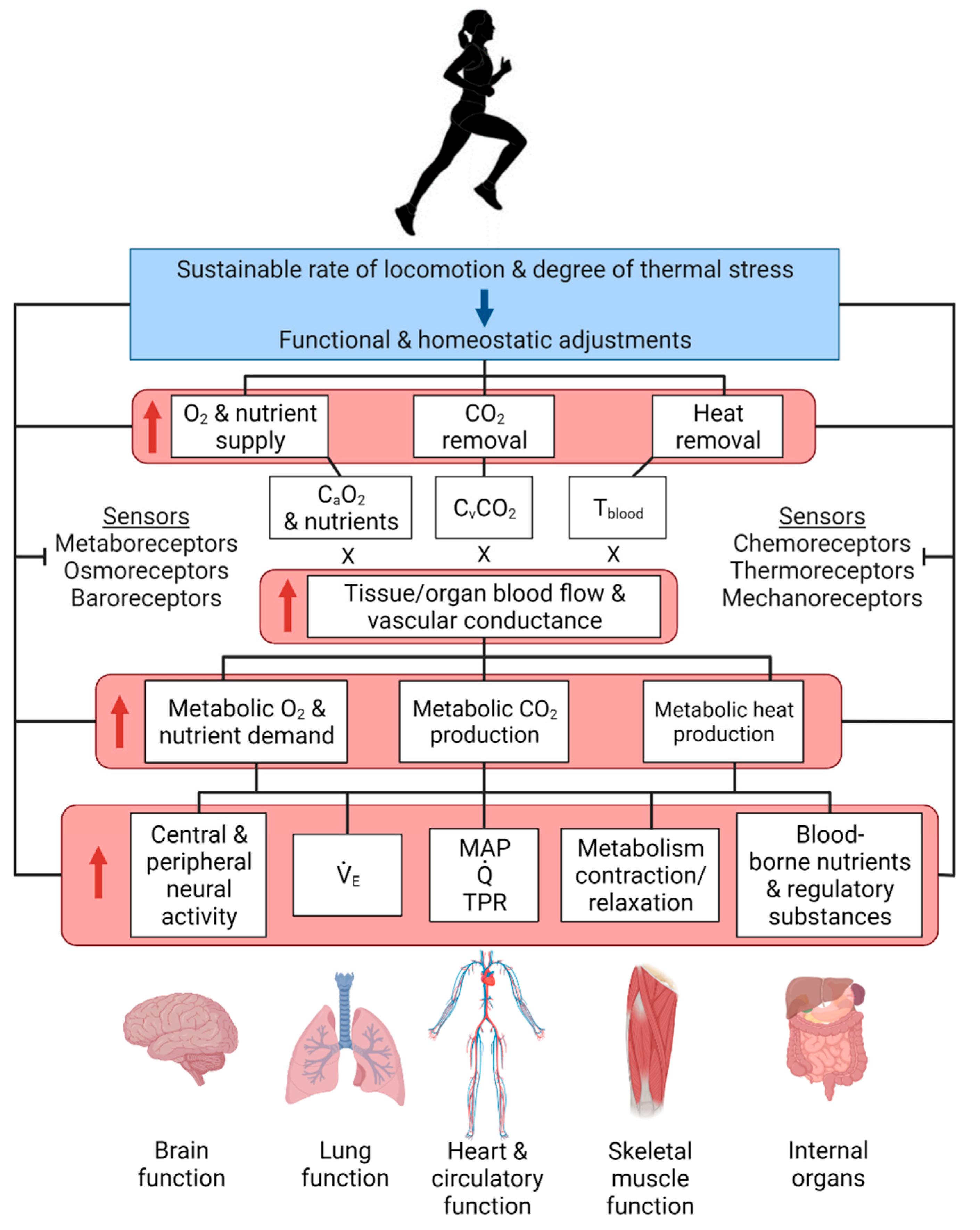
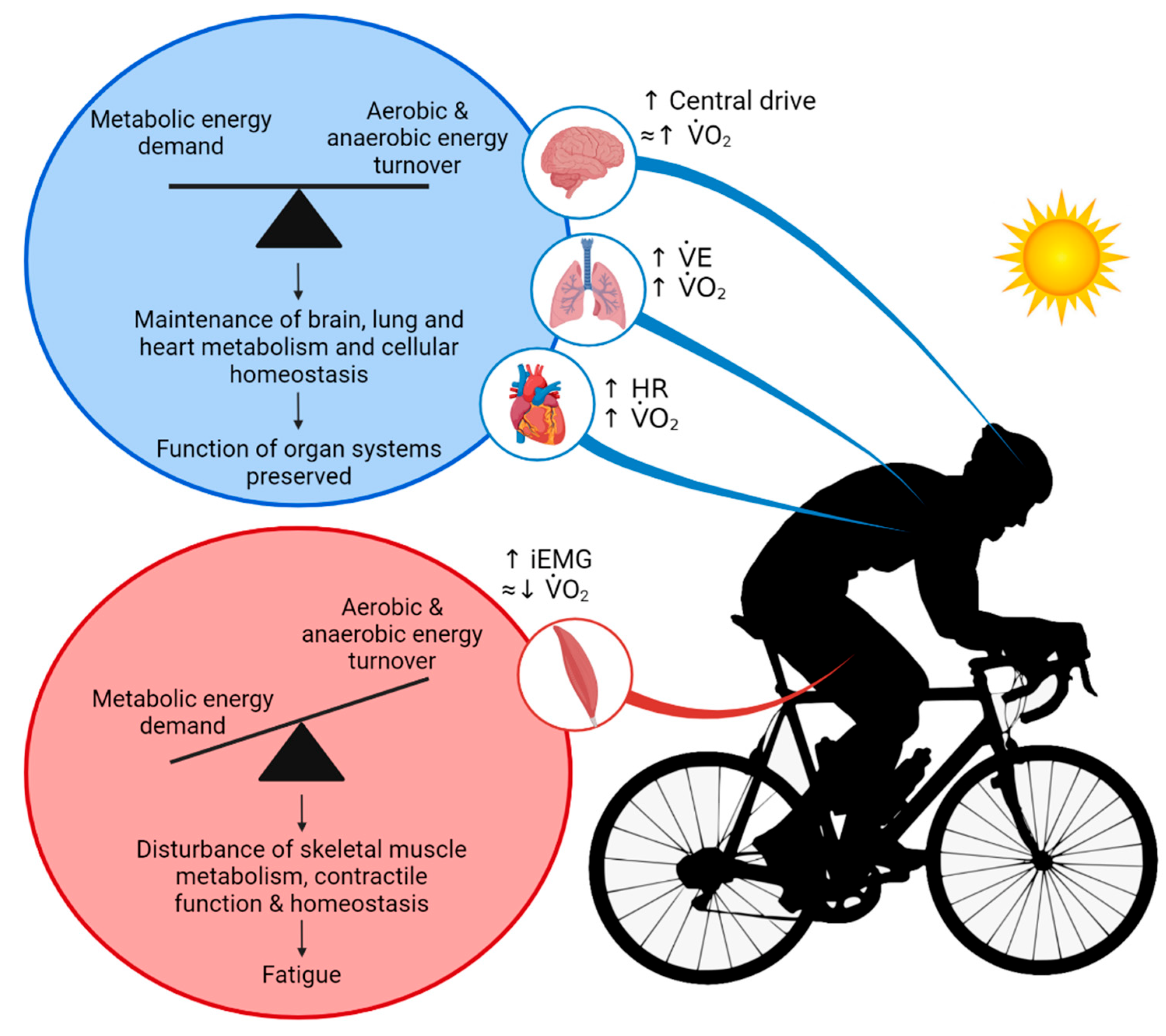
6. Integrative View of Physiological Systems and Homeostasis during Exercise and Environmental Stress
7. Organ Systems Interactions—Future Research Directions
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bernard, C. Leçons sur les Phénomènes de la vie Communs aux Animaux et aux Végétaus/Par Claude Bernard.; JB Baillière: Paris, France, 1879. [Google Scholar]
- Cannon, W.B. Organization for physiological homeostasis. Physiol. Rev. 1929, 9, 399–431. [Google Scholar] [CrossRef]
- Rowell, L.B. Human Cardiovascular Control; Oxford University Press: New York, NY, USA, 1993; ISBN 9-19-507362-2. [Google Scholar]
- Joyner, M.J.; Casey, D.P. Regulation of Increased blood flow (hyperemia) to muscles during exercise: A hierarchy of competing physiological needs. Physiol. Rev. 2015, 95, 549–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saltin, B.; Rådegran, G.; Koskolou, M.D.; Roach, R.C. Skeletal muscle blood flow in humans and its regulation during exercise. Acta Physiol. Scand. 1998, 162, 421–436. [Google Scholar] [CrossRef] [PubMed]
- González-Alonso, J.; Calbet, J.A.L.; Nielsen, B. Muscle blood flow is reduced with dehydration during prolonged exercise in humans. J. Physiol. 1998, 513, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, M.N.; Raven, P.B.; Snell, P.G.; Stray-Gundersen, J.; Levine, B.D. Maximal oxygen uptake as a parametric measure of cardiorespiratory capacity. Med. Sci. Sports Exerc. 2007, 39, 103–107. [Google Scholar] [CrossRef] [PubMed]
- Roca, J.; Agusti, A.G.; Alonso, A.; Poole, D.C.; Viegas, C.; Barbera, J.A.; Rodriguez-Roisin, R.; Ferrer, A.; Wagner, P.D. Effects of training on muscle O2 transport at VO2max. J. Appl. Physiol. 1992, 73, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- Roca, J.; Hogan, M.C.; Story, D.; Bebout, D.E.; Haab, P.; Gonzalez, R.; Ueno, O.; Wagner, P.D. Evidence for tissue diffusion limitation of VO2max in normal humans. J. Appl. Physiol. 1989, 67, 291–299. [Google Scholar] [CrossRef]
- McGuire, D.K.; Levine, B.D.; Williamson, J.W.; Snell, P.G.; Blomqvist, C.G.; Saltin, B.; Mitchell, J.H. A 30-year follow-up of the Dallas Bedrest and Training Study: I. Effect of age on the cardiovascular response to exercise. Circulation 2001, 104, 1350–1357. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, S.P.; Dawson, E.A.; Yoshiga, C.C.; Dalsgaard, M.K.; Damsgaard, R.; Secher, N.H.; González-Alonso, J. Limitations to systemic and locomotor limb muscle oxygen delivery and uptake during maximal exercise in humans. J. Physiol. 2005, 566, 273–285. [Google Scholar] [CrossRef]
- Rowell, L.B.; Murray, J.A.; Brengelmann, G.L.; Kraning, K.K. Human cardiovascular adjustments to rapid changes in skin temperature during exercise. Circ. Res. 1969, 24, 711–724. [Google Scholar] [CrossRef] [Green Version]
- Andersen, P.; Saltin, B. Maximal perfusion of skeletal muscle in man. J. Physiol. 1985, 366, 233–249. [Google Scholar] [CrossRef]
- Ekblom, B.; Goldbarg, A.N.; Gullbring, B. Response to exercise after blood loss and reinfusion. J. Appl. Physiol. 1972, 33, 175–180. [Google Scholar] [CrossRef]
- Dempsey, J.A.; La Gerche, A.; Hull, J.H. Is the healthy respiratory system built just right, overbuilt, or underbuilt to meet the demands imposed by exercise? J. Appl. Physiol. 2020, 129, 1235–1256. [Google Scholar] [CrossRef]
- Mitchell, J.H.; Sproule, B.J.; Chapman, C.B. The physiological meaning of the maximal oxygen intake test. J. Clin. Investig. 1958, 37, 538–547. [Google Scholar] [CrossRef] [Green Version]
- Saltin, B.; Astrand, P.O. Maximal oxygen uptake in athletes. J. Appl. Physiol. 1967, 23, 353–358. [Google Scholar] [CrossRef]
- Sutton, J.R.; Reeves, J.T.; Wagner, P.D.; Groves, B.M.; Cymerman, A.; Malconian, M.K.; Rock, P.B.; Young, P.M.; Walter, S.D.; Houston, C.S. Operation Everest II: Oxygen transport during exercise at extreme simulated altitude. J. Appl. Physiol. 1988, 64, 1309–1321. [Google Scholar] [CrossRef]
- González-Alonso, J. A novel role for the red blood cell in the regulation of tissue O2 supply. J. Physiol. 2012, 590, 4983–4984. [Google Scholar] [CrossRef] [Green Version]
- Åstrand, P.-O. Human physical fitness with special reference to sex and age. Physiol. Rev. 1956, 36, 307–335. [Google Scholar] [CrossRef]
- Kokkinos, P.; Myers, J.; Kokkinos, J.P.; Pittaras, A.; Narayan, P.; Manolis, A.; Karasik, P.; Greenberg, M.; Papademetriou, V.; Singh, S. Exercise capacity and mortality in black and white men. Circulation 2008, 117, 614–622. [Google Scholar] [CrossRef] [Green Version]
- Rowland, T.W. Developmental aspects of physiological function relating to aerobic exercise in children. Sports Med. 1990, 10, 255–266. [Google Scholar] [CrossRef]
- Edvardsen, E.; Hansen, B.H.; Holme, I.M.; Dyrstad, S.M.; Anderssen, S.A. Reference values for cardiorespiratory response and fitness on the treadmill in a 20- to 85-year-old population. Chest 2013, 144, 241–248. [Google Scholar] [CrossRef]
- Madsen, P.L.; Sperling, B.K.; Warming, T.; Schmidt, J.F.; Secher, N.H.; Wildschiodtz, G.; Holm, S.; Lassen, N.A. Middle cerebral artery blood velocity and cerebral blood flow and O2 uptake during dynamic exercise. J. Appl. Physiol. 1993, 74, 245–250. [Google Scholar] [CrossRef]
- Clarke, D.D.; Sokoloff, L. Regulation of Cerebral Metabolic Rate. In Basic Neurochemistry: Molecular, Cellular and Medical Aspects; Siegel, G.J., Arganoff, B.W., Albers, R.W., Fisher, S.K., Uhler, M.D., Eds.; Lippincott-Raven: Philadelphia, PA, USA, 1999. [Google Scholar]
- Delp, M.D.; Armstrong, R.B.; Godfrey, D.A.; Laughlin, M.H.; Ross, C.D.; Wilkerson, M.K. Exercise increases blood flow to locomotor, vestibular, cardiorespiratory and visual regions of the brain in miniature swine. J. Physiol. 2001, 533, 849–859. [Google Scholar] [CrossRef]
- Brümmer, V.; Schneider, S.; Strüder, H.K.; Askew, C.D. Primary motor cortex activity is elevated with incremental exercise intensity. Neuroscience 2011, 181, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Christensen, L.O.D.; Johannsen, P.; Sinkjær, T.; Petersen, N.; Pyndt, H.S.; Nielsen, J.B. Cerebral activation during bicycle movements in man. Exp. Brain Res. 2000, 135, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Lassen, N.A. Normal Average Value of Cerebral Blood Flow in Younger Adults is 50 mL/100 g/min. J. Cereb. Blood Flow Metab. 1985, 5, 347–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olesen, J. Contralateral focal increase of cerebral blood flow in man during arm work. Brain 1971, 94, 635–646. [Google Scholar] [CrossRef]
- Jørgensen, L.G.; Perko, M.; Hanel, B.; Schroeder, T.V.; Secher, N.H. Middle cerebral artery flow velocity and blood flow during exercise and muscle ischemia in humans. J. Appl. Physiol. 1992, 72, 1123–1132. [Google Scholar] [CrossRef]
- Hellstrom, G.; Fischer-Colbrie, W.; Wahlgren, N.G.; Jogestrand, T. Carotid artery blood flow and middle cerebral artery blood flow velocity during physical exercise. J. Appl. Physiol. 1996, 81, 413–418. [Google Scholar] [CrossRef]
- Fisher, J.P.; Hartwich, D.; Seifert, T.; Olesen, N.D.; Mcnulty, C.L.; Nielsen, H.B.; van Lieshout, J.J.; Secher, N.H. Cerebral perfusion, oxygenation and metabolism during exercise in young and elderly individuals. J. Physiol. 2013, 591, 1859–1870. [Google Scholar] [CrossRef]
- Sato, K.; Ogoh, S.; Hirasawa, A.; Oue, A.; Sadamoto, T. The distribution of blood flow in the carotid and vertebral arteries during dynamic exercise in humans. J. Physiol. 2011, 589, 2847–2856. [Google Scholar] [CrossRef]
- Trangmar, S.J.; Chiesa, S.T.; Kalsi, K.K.; Secher, N.H.; González-Alonso, J. Whole body hyperthermia, but not skin hyperthermia, accelerates brain and locomotor limb circulatory strain and impairs exercise capacity in humans. Physiol. Rep. 2017, 5, e13108. [Google Scholar] [CrossRef]
- Trangmar, S.J.; Chiesa, S.T.; Stock, C.G.; Kalsi, K.K.; Secher, N.H.; González-Alonso, J. Dehydration affects cerebral blood flow but not its metabolic rate for oxygen during maximal exercise in trained humans. J. Physiol. 2014, 592, 3143–3160. [Google Scholar] [CrossRef]
- Smith, K.J.; MacLeod, D.; Willie, C.K.; Lewis, N.C.S.; Hoiland, R.L.; Ikeda, K.; Tymko, M.M.; Donnelly, J.; Day, T.A.; MacLeod, N.; et al. Influence of high altitude on cerebral blood flow and fuel utilization during exercise and recovery. J. Physiol. 2014, 592, 5507–5527. [Google Scholar] [CrossRef]
- Nybo, L.; Nielsen, B. Middle cerebral artery blood velocity is reduced with hyperthermia during prolonged exercise in humans. J. Physiol. 2001, 534, 279–286. [Google Scholar] [CrossRef]
- Périard, J.D.; Racinais, S. Heat stress exacerbates the reduction in middle cerebral artery blood velocity during prolonged self-paced exercise. Scand. J. Med. Sci. Sports 2015, 25, 135–144. [Google Scholar] [CrossRef]
- Nybo, L.; Møller, K.; Volianitis, S.; Nielsen, B.; Secher, N.H. Effects of hyperthermia on cerebral blood flow and metabolism during prolonged exercise in humans. J. Appl. Physiol. 2002, 93, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Keiser, S.; Flück, D.; Hüppin, F.; Stravs, A.; Hilty, M.P.; Lundby, C. Heat training increases exercise capacity in hot but not in temperate conditions: A mechanistic counter-balanced cross-over study. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H750–H761. [Google Scholar] [CrossRef] [Green Version]
- González-Alonso, J.; Dalsgaard, M.K.; Osada, T.; Volianitis, S.; Dawson, E.A.; Yoshiga, C.C.; Secher, N.H. Brain and central haemodynamics and oxygenation during maximal exercise in humans. J. Physiol. 2004, 557, 331–342. [Google Scholar] [CrossRef]
- Trangmar, S.J.; Chiesa, S.T.; Llodio, I.; Garcia, B.; Kalsi, K.K.; Secher, N.H.; González-Alonso, J. Dehydration accelerates reductions in cerebral blood flow during prolonged exercise in the heat without compromising brain metabolism. Am. J. Physiol. Heart Circ. Physiol. 2015, 309, H1598–H1607. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, K.; Stöhr, E.J.; Akiyama, K.; Watanabe, S.; González-Alonso, J. Dehydration reduces stroke volume and cardiac output during exercise because of impaired cardiac filling and venous return, not left ventricular function. Physiol. Rep. 2020, 8, e14433. [Google Scholar] [CrossRef]
- Attwell, D.; Buchan, A.M.; Charpak, S.; Lauritzen, M.; MacVicar, B.A.; Newman, E.A. Glial and neuronal control of brain blood flow. Nature 2010, 468, 232–243. [Google Scholar] [CrossRef] [Green Version]
- Dienel, G.A. Fueling and Imaging Brain Activation. ASN Neuro 2012, 4, AN20120021. [Google Scholar] [CrossRef] [Green Version]
- Hoiland, R.L.; Caldwell, H.G.; Howe, C.A.; Nowak-Flück, D.; Stacey, B.S.; Bailey, D.M.; Paton, J.F.R.; Green, D.J.; Sekhon, M.S.; Macleod, D.B.; et al. Nitric oxide is fundamental to neurovascular coupling in humans. J. Physiol. 2020, 598, 4927–4939. [Google Scholar] [CrossRef]
- Iadecola, C. The neurovascular unit coming of age: A journey through neurovascular coupling in health and disease. Neuron 2017, 96, 17–42. [Google Scholar] [CrossRef] [Green Version]
- Madsen, P.L.; Hasselbalch, S.G.; Hagemann, L.P.; Olsen, K.S.; Bülow, J.; Holm, S.; Wildschiødtz, G.; Paulson, O.B.; Lassen, N.A. Persistent resetting of the cerebral oxygen/glucose uptake ratio by brain activation: Evidence obtained with the Kety—Schmidt technique. J. Cereb. Blood Flow Metab. 1995, 15, 485–491. [Google Scholar] [CrossRef] [Green Version]
- Fox, P.T.; Raichle, M.E.; Mintun, M.A.; Dence, C. Nonoxidative glucose consumption during focal physiologic neural activity. Science 1988, 241, 462–464. [Google Scholar] [CrossRef]
- Willie, C.K.; Cowan, E.C.; Ainslie, P.N.; Taylor, C.E.; Smith, K.J.; Sin, P.Y.W.; Tzeng, Y.C. Neurovascular coupling and distribution of cerebral blood flow during exercise. J. Neurosci. Methods 2011, 198, 270–273. [Google Scholar] [CrossRef]
- Rupp, T.; Perrey, S. Prefrontal cortex oxygenation and neuromuscular responses to exhaustive exercise. Eur. J. Appl. Physiol. 2007, 102, 153–163. [Google Scholar] [CrossRef]
- Subudhi, A.W.; Lorenz, M.C.; Fulco, C.S.; Roach, R.C. Cerebrovascular responses to incremental exercise during hypobaric hypoxia: Effect of oxygenation on maximal performance. Am. J. Physiol. Circ. Physiol. 2008, 294, H164–H171. [Google Scholar] [CrossRef]
- Vogiatzis, I.; Louvaris, Z.; Habazettl, H.; Athanasopoulos, D.; Andrianopoulos, V.; Cherouveim, E.; Wagner, H.; Roussos, C.; Wagner, P.D.; Zakynthinos, S. Frontal cerebral cortex blood flow, oxygen delivery and oxygenation during normoxic and hypoxic exercise in athletes. J. Physiol. 2011, 589, 4027–4039. [Google Scholar] [CrossRef] [PubMed]
- Willie, C.K.; Tzeng, Y.C.; Fisher, J.A.; Ainslie, P.N. Integrative regulation of human brain blood flow. J. Physiol. 2014, 592, 841–859. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.J.; Ainslie, P.N. Regulation of cerebral blood flow and metabolism during exercise. Exp. Physiol. 2017, 102, 1356–1371. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, L.G.; Perko, G.; Secher, N.H. Regional cerebral artery mean flow velocity and blood flow during dynamic exercise in humans. J. Appl. Physiol. 1992, 73, 1825–1830. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, H.G.; Coombs, G.B.; Howe, C.A.; Hoiland, R.L.; Patrician, A.; Lucas, S.J.E.; Ainslie, P.N. Evidence for temperature-mediated regional increases in cerebral blood flow during exercise. J. Physiol. 2020, 598, 1459–1473. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.D.; Haykowsky, M.J.; Stickland, M.K.; Altamirano-Diaz, L.A.; Willie, C.K.; Smith, K.J.; Petersen, S.R.; Ainslie, P.N. Reductions in cerebral blood flow during passive heat stress in humans: Partitioning the mechanisms. J. Physiol. 2011, 589, 4053–4064. [Google Scholar] [CrossRef] [PubMed]
- Bain, A.R.; Smith, K.J.; Lewis, N.C.; Foster, G.E.; Wildfong, K.W.; Willie, C.K.; Hartley, G.L.; Cheung, S.S.; Ainslie, P.N. Regional changes in brain blood flow during severe passive hyperthermia: Effects of PaCO2 and extracranial blood flow. J. Appl. Physiol. 2013, 115, 653–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogoh, S.; Sato, K.; Okazaki, K.; Miyamoto, T.; Hirasawa, A.; Morimoto, K.; Shibasaki, M. Blood flow distribution during heat stress: Cerebral and systemic blood flow. J. Cereb. Blood Flow Metab. 2013, 33, 1915–1920. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, P.; Nybo, L.; Volianitis, S.; Møller, K.; Secher, N.H.; Gjedde, A. Cerebral oxygenation is reduced during hyperthermic exercise in humans. Acta Physiol. 2010, 199, 63–70. [Google Scholar] [CrossRef]
- Rasmussen, P.; Dawson, E.A.; Nybo, L.; van Lieshout, J.J.; Secher, N.H.; Gjedde, A. Capillary-oxygenation-level-dependent near-infrared spectrometry in frontal lobe of humans. J. Cereb. Blood Flow Metab. 2007, 27, 1082–1093. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, S.P.; Damsgaard, R.; Dawson, E.A.; Secher, N.H.; González-Alonso, J. Restrictions in systemic and locomotor skeletal muscle perfusion, oxygen supply and VO2 during high-intensity whole-body exercise in humans. J. Physiol. 2008, 586, 2621–2635. [Google Scholar] [CrossRef]
- González-alonso, J.; Mortensen, S.P.; Jeppesen, T.D.; Ali, L.; Barker, H.; Damsgaard, R.; Secher, N.H.; Dawson, E.A.; Dufour, S.P. Haemodynamic responses to exercise, ATP infusion and thigh compression in humans: Insight into the role of muscle mechanisms on cardiovascular function. J. Physiol. 2008, 586, 2405–2417. [Google Scholar] [CrossRef]
- González-Alonso, J.; Teller, C.; Andersen, S.L.; Jensen, F.B.; Hyldig, T.; Nielsen, B. Influence of body temperature on the development of fatigue during prolonged exercise in the heat. J. Appl. Physiol. 1999, 86, 1032–1039. [Google Scholar] [CrossRef] [Green Version]
- Nybo, L.; Jensen, T.; Nielsen, B.; González-Alonso, J. Effects of marked hyperthermia with and without dehydration on VO2 kinetics during intense exercise. J. Appl. Physiol. 2001, 90, 1057–1064. [Google Scholar] [CrossRef] [Green Version]
- Nybo, L.; González-Alonso, J. Critical core temperature: A hypothesis too simplistic to explain hyperthermia-induced fatigue. Scand. J. Med. Sci. Sports 2015, 25, 4–5. [Google Scholar] [CrossRef] [Green Version]
- Nybo, L.; Secher, N.H.; Nielsen, B. Inadequate heat release from the human brain during prolonged exercise with hyperthermia. J. Physiol. 2002, 545, 697–704. [Google Scholar] [CrossRef]
- Yablonskiy, D.A.; Ackerman, J.J.H.; Raichle, M.E. Coupling between changes in human brain temperature and oxidative metabolism during prolonged visual stimulation. Proc. Natl. Acad. Sci. USA 2000, 97, 7603–7608. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, B.; Hyldig, T.; Bidstrup, F.; González-Alonso, J.; Christoffersen, G.R.J. Brain activity and fatigue during prolonged exercise in the heat. Pflügers Arch. 2001, 442, 41–48. [Google Scholar] [CrossRef]
- Rasmussen, P.; Stie, H.; Nybo, L.; Nielsen, B. Heat induced fatigue and changes of the EEG is not related to reduced perfusion of the brain during prolonged exercise in humans. J. Therm. Biol. 2004, 29, 731–737. [Google Scholar] [CrossRef]
- Périard, J.D.; De Pauw, K.; Zanow, F.; Racinais, S. Cerebrocortical activity during self-paced exercise in temperate, hot and hypoxic conditions. Acta Physiol. 2018, 222, e12916. [Google Scholar] [CrossRef]
- Périard, J.D.; Christian, R.J.; Knez, W.L.; Racinais, S. Voluntary muscle and motor cortical activation during progressive exercise and passively induced hyperthermia. Exp. Physiol. 2014, 99, 136–148. [Google Scholar] [CrossRef] [PubMed]
- De Pauw, K.; Roelands, B.; Marušič, U.; Tellez, H.F.; Knaepen, K.; Meeusen, R. Brain mapping after prolonged cycling and during recovery in the heat. J. Appl. Physiol. 2013, 115, 1324–1331. [Google Scholar] [CrossRef] [Green Version]
- Racinais, S.; Gaoua, N.; Grantham, J. Hyperthermia impairs short-term memory and peripheral motor drive transmission. J. Physiol. 2008, 586, 4751–4762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodall, S.; González-Alonso, J.; Ali, L.; Ross, E.Z.; Romer, L.M. Supraspinal fatigue after normoxic and hypoxic exercise in humans. J. Physiol. 2012, 590, 2767–2782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodall, S.; Charlton, K.; Hignett, C.; Prichard, J.; Barwood, M.; Howatson, G.; Thomas, K. Augmented supraspinal fatigue following constant-load cycling in the heat. Scand. J. Med. Sci. Sports 2015, 25, 164–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Racinais, S.; Buchheit, M.; Girard, O. Breakpoints in ventilation, cerebral and muscle oxygenation, and muscle activity during an incremental cycling exercise. Front. Physiol. 2014, 5, 142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, A.; St Clair Gibson, A.; Mbambo, Z.; Lambert, M.; Noakes, T. The effects of heat stress on neuromuscular activity during endurance exercise. Pflügers Arch. Eur. J. Physiol. 2002, 444, 738–743. [Google Scholar] [CrossRef] [Green Version]
- Forster, H.V.; Pan, L.G.; Funahashi, A. Temporal pattern of arterial CO2 partial pressure during exercise in humans. J. Appl. Physiol. 1986, 60, 653–660. [Google Scholar] [CrossRef]
- Wasserman, K.; Van Kessel, A.L.; Burton, G.G. Interaction of physiological mechanisms during exercise. J. Appl. Physiol. 1967, 22, 71–85. [Google Scholar] [CrossRef]
- Dempsey, J.A.; Hanson, P.G.; Henderson, K.S. Exercise-induced arterial hypoxaemia in healthy human subjects at sea level. J. Physiol. 1984, 355, 161–175. [Google Scholar] [CrossRef]
- Nielsen, H.B.; Madsen, P.; Svendsen, L.B.; Roach, R.C.; Secher, N.H. The influence of PaO2, pH and SaO2 on maximal oxygen uptake. Acta Physiol. Scand. 1998, 164, 89–97. [Google Scholar] [CrossRef]
- Dominelli, P.B.; Foster, G.E.; Dominelli, G.S.; Henderson, W.R.; Koehle, M.S.; McKenzie, D.C.; Sheel, A.W. Exercise-induced arterial hypoxaemia and the mechanics of breathing in healthy young women. J. Physiol. 2013, 591, 3017–3034. [Google Scholar] [CrossRef]
- Durand, F.; Mucci, P.; Préfaut, C. Evidence for an inadequate hyperventilation inducing arterial hypoxemia at submaximal exercise in all highly trained endurance athletes. Med. Sci. Sports Exerc. 2000, 32, 926–932. [Google Scholar] [CrossRef]
- Hopkins, S.R.; McKenzie, D.C.; Schoene, R.B.; Glenny, R.W.; Robertson, H.T. Pulmonary gas exchange during exercise in athletes. I. Ventilation-perfusion mismatch and diffusion limitation. J. Appl. Physiol. 1994, 77, 912–917. [Google Scholar] [CrossRef]
- Wagner, P.D.; Gale, G.E.; Moon, R.E.; Torre-Bueno, J.R.; Stolp, B.W.; Saltzman, H.A. Pulmonary gas exchange in humans exercising at sea level and simulated altitude. J. Appl. Physiol. 1986, 61, 260–270. [Google Scholar] [CrossRef]
- Marshall, H.; Gibson, O.R.; Romer, L.M.; Illidi, C.; Hull, J.H.; Kippelen, P. Systemic but not local rehydration restores dehydration-induced changes in pulmonary function in healthy adults. J. Appl. Physiol. 2021, 130, 517–527. [Google Scholar] [CrossRef]
- Govindaraj, M. The effect of dehydration on the ventilatory capacity in normal subjects. Am. Rev. Respir. Dis. 1972, 105, 842–844. [Google Scholar] [CrossRef]
- Tipton, M.J.; Kadinopoulos, P.; de Sa, D.R.; Barwood, M.J. Changes in lung function during exercise are independently mediated by increases in deep body temperature. BMJ Open Sport Exerc. Med. 2017, 3, e000210. [Google Scholar] [CrossRef] [Green Version]
- Haldane, J.S. The influence of high air temperatures No. I. J. Hyg. 1905, 5, 494–513. [Google Scholar] [CrossRef]
- Cabanac, M.; White, M.D. Core temperature thresholds for hyperpnea during passive hyperthermia in humans. Eur. J. Appl. Physiol. Occup. Physiol. 1995, 71, 71–76. [Google Scholar] [CrossRef]
- Fujii, N.; Honda, Y.; Hayashi, K.; Soya, H.; Kondo, N.; Nishiyasu, T. Comparison of hyperthermic hyperpnea elicited during rest and submaximal, moderate-intensity exercise. J. Appl. Physiol. 2008, 104, 998–1005. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, B.; Honda, Y.; Fujii, N.; Kondo, N.; Nishiyasu, T. Comparison of hyperthermic hyperventilation during passive heating and prolonged light and moderate exercise in the heat. J. Appl. Physiol. 2012, 113, 1388–1397. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, T.D.; Ainslie, P.N.; Thomas, K.N.; Wilson, L.C.; Akerman, A.P.; Donnelly, J.; Campbell, H.A.; Cotter, J.D. Influence of the mode of heating on cerebral blood flow, non-invasive intracranial pressure and thermal tolerance in humans. J. Physiol. 2021, 599, 1977–1996. [Google Scholar] [CrossRef] [PubMed]
- Cotes, J.E. The role of body temperature in controlling ventilation during exercise in one normal subject breathing oxygen. J. Physiol. 1955, 129, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Chu, A.L.; Jay, O.; White, M.D. The effects of hyperthermia and hypoxia on ventilation during low-intensity steady-state exercise. Am. J. Physiol. Integr. Comp. Physiol. 2007, 292, R195–R203. [Google Scholar] [CrossRef]
- White, M.D.; Cabanac, M. Exercise hyperpnea and hyperthermia in humans. J. Appl. Physiol. 1996, 81, 1249–1254. [Google Scholar] [CrossRef]
- White, M.D.; Cabanac, M. Core temperature thresholds for ventilation during exercise. In Modeling and Control of Ventilation; Semple, S.J.G., Adams, L., Brian, B.J., Eds.; Plenum Press: New York, NY, USA, 1995; pp. 173–177. [Google Scholar]
- Gaudio, R.; Abramson, N. Heat-induced hyperventilation. J. Appl. Physiol. 1968, 25, 742–746. [Google Scholar] [CrossRef]
- Hayashi, K.; Honda, Y.; Ogawa, T.; Kondo, N.; Nishiyasu, T. Relationship between ventilatory response and body temperature during prolonged submaximal exercise. J. Appl. Physiol. 2006, 100, 414–420. [Google Scholar] [CrossRef]
- Tsuji, B.; Hayashi, K.; Kondo, N.; Nishiyasu, T. Characteristics of hyperthermia-induced hyperventilation in humans. Temperature 2016, 3, 146–160. [Google Scholar] [CrossRef] [Green Version]
- White, M.D. Components and mechanisms of thermal hyperpnea. J. Appl. Physiol. 2006, 101, 655–663. [Google Scholar] [CrossRef]
- Hanson, R.d.G. Respiratory heat loss at increased core temperature. J. Appl. Physiol. 1974, 37, 103–107. [Google Scholar] [CrossRef]
- Rasch, W.; Samson, P.; Cote, J.; Cabanac, M. Heat loss from the human head during exercise. J. Appl. Physiol. 1991, 71, 590–595. [Google Scholar] [CrossRef]
- White, M.D.; Greiner, J.G.; McDonald, P.L.L. Point: Humans do demonstrate selective brain cooling during hyperthermia. J. Appl. Physiol. 2011, 110, 569–571. [Google Scholar] [CrossRef]
- Nybo, L.; Secher, N.H. Counterpoint: Humans do not demonstrate selective brain cooling during hyperthermia. J. Appl. Physiol. 2011, 110, 571–573. [Google Scholar] [CrossRef]
- Kety, S.S.; Polis, B.D. The blood flow and oxygen consumption of the human brain in diabetic acidosis and coma. J. Clin. Investig. 1948, 27, 500–510. [Google Scholar] [CrossRef]
- Brothers, R.M.; Wingo, J.E.; Hubing, K.A.; Crandall, C.G. The effects of reduced end-tidal carbon dioxide tension on cerebral blood flow during heat stress. J. Physiol. 2009, 587, 3921–3927. [Google Scholar] [CrossRef]
- Fujii, N.; Honda, Y.; Hayashi, K.; Kondo, N.; Koga, S.; Nishiyasu, T. Effects of chemoreflexes on hyperthermic hyperventilation and cerebral blood velocity in resting heated humans. Exp. Physiol. 2008, 93, 994–1001. [Google Scholar] [CrossRef]
- Johnson, B.D.; Babcock, M.A.; Suman, O.E.; Dempsey, J.A. Exercise-induced diaphragmatic fatigue in healthy humans. J. Physiol. 1993, 460, 385–405. [Google Scholar] [CrossRef]
- Taylor, B.J.; How, S.C.; Romer, L.M. Exercise-induced abdominal muscle fatigue in healthy humans. J. Appl. Physiol. 2006, 100, 1554–1562. [Google Scholar] [CrossRef]
- Dempsey, J.A.; Romer, L.; Rodman, J.; Miller, J.; Smith, C. Consequences of exercise-induced respiratory muscle work. Respir. Physiol. Neurobiol. 2006, 151, 242–250. [Google Scholar] [CrossRef]
- Ichinose, M.; Saito, M.; Fujii, N.; Ogawa, T.; Hayashi, K.; Kondo, N.; Nishiyasu, T. Modulation of the control of muscle sympathetic nerve activity during incremental leg cycling. J. Physiol. 2008, 586, 2753–2766. [Google Scholar] [CrossRef]
- Rosenmeier, J.B.; Hansen, J.; González-Alonso, J. Circulating ATP-induced vasodilatation overrides sympathetic vasoconstrictor activity in human skeletal muscle. J. Physiol. 2004, 558, 351–365. [Google Scholar] [CrossRef] [Green Version]
- Calbet, J.A.L.; González-Alonso, J.; Helge, J.W.; Søndergaard, H.; Munch-Andersen, T.; Boushel, R.; Saltin, B. Cardiac output and leg and arm blood flow during incremental exercise to exhaustion on the cycle ergometer. J. Appl. Physiol. 2007, 103, 969–978. [Google Scholar] [CrossRef]
- Harms, C.A.; Babcock, M.A.; McClaran, S.R.; Pegelow, D.F.; Nickele, G.A.; Nelson, W.B.; Dempsey, J.A. Respiratory muscle work compromises leg blood flow during maximal exercise. J. Appl. Physiol. 1997, 82, 1573–1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harms, C.A.; Wetter, T.J.; St. Croix, C.M.; Pegelow, D.F.; Dempsey, J.A. Effects of respiratory muscle work on exercise performance. J. Appl. Physiol. 2000, 89, 131–138. [Google Scholar] [CrossRef]
- Dominelli, P.B.; Archiza, B.; Ramsook, A.H.; Mitchell, R.A.; Peters, C.M.; Molgat-Seon, Y.; Henderson, W.R.; Koehle, M.S.; Boushel, R.; Sheel, A.W. Effects of respiratory muscle work on respiratory and locomotor blood flow during exercise. Exp. Physiol. 2017, 102, 1535–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harms, C.A.; Wetter, T.J.; McClaran, S.R.; Pegelow, D.F.; Nickele, G.A.; Nelson, W.B.; Hanson, P.; Dempsey, J.A. Effects of respiratory muscle work on cardiac output and its distribution during maximal exercise. J. Appl. Physiol. 1998, 85, 609–618. [Google Scholar] [CrossRef]
- Aaron, E.A.; Johnson, B.D.; Seow, C.K.; Dempsey, J.A. Oxygen cost of exercise hyperpnea: Measurement. J. Appl. Physiol. 1992, 72, 1810–1817. [Google Scholar] [CrossRef] [PubMed]
- Dominelli, P.B.; Render, J.N.; Molgat-Seon, Y.; Foster, G.E.; Romer, L.M.; Sheel, A.W. Oxygen cost of exercise hyperpnoea is greater in women compared with men. J. Physiol. 2015, 593, 1965–1979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogiatzis, I.; Athanasopoulos, D.; Habazettl, H.; Kuebler, W.M.; Wagner, H.; Roussos, C.; Wagner, P.D.; Zakynthinos, S. Intercostal muscle blood flow limitation in athletes during maximal exercise. J. Physiol. 2009, 587, 3665–3677. [Google Scholar] [CrossRef] [PubMed]
- Munch, G.D.W.; Svendsen, J.H.; Damsgaard, R.; Secher, N.H.; González-Alonso, J.; Mortensen, S.P. Maximal heart rate does not limit cardiovascular capacity in healthy humans: Insight from right atrial pacing during maximal exercise. J. Physiol. 2014, 592, 377–390. [Google Scholar] [CrossRef]
- Parkes, M.J. Reappraisal of systemic venous chemoreceptors: Might they explain the matching of breathing to metabolic rate in humans? Exp. Physiol. 2017, 102, 1567–1583. [Google Scholar] [CrossRef]
- Parkes, M.J. Evaluating the importance of the carotid chemoreceptors in controlling breathing during exercise in man. Biomed. Res. Int. 2013, 2013, 893506. [Google Scholar] [CrossRef]
- Forster, H.V.; Haouzi, P.; Dempsey, J.A. Control of breathing during exercise. Compr. Physiol. 2012, 2, 743–777. [Google Scholar] [CrossRef]
- Paterson, D.J. Defining the neurocircuitry of exercise hyperpnoea. J. Physiol. 2014, 592, 433–444. [Google Scholar] [CrossRef]
- Amann, M.; Blain, G.M.; Proctor, L.T.; Sebranek, J.J.; Pegelow, D.F.; Dempsey, J.A. Group III and IV muscle afferents contribute to ventilatory and cardiovascular response to rhythmic exercise in humans. J. Appl. Physiol. 2010, 109, 966–976. [Google Scholar] [CrossRef] [Green Version]
- Bruce, R.M.; Jolley, C.; White, M.J. Control of exercise hyperpnoea: Contributions from thin-fibre skeletal muscle afferents. Exp. Physiol. 2019, 104, 1605–1621. [Google Scholar] [CrossRef]
- Fujii, N.; Kashihara, M.; Kenny, G.P.; Honda, Y.; Fujimoto, T.; Cao, Y.; Nishiyasu, T. Carotid chemoreceptors have a limited role in mediating the hyperthermia-induced hyperventilation in exercising humans. J. Appl. Physiol. 2019, 126, 305–313. [Google Scholar] [CrossRef]
- Tsuji, B.; Honda, Y.; Ikebe, Y.; Fujii, N.; Kondo, N.; Nishiyasu, T. Voluntary suppression of hyperthermia-induced hyperventilation mitigates the reduction in cerebral blood flow velocity during exercise in the heat. Am. J. Physiol. Integr. Comp. Physiol. 2015, 308, R669–R679. [Google Scholar] [CrossRef]
- Tryba, A.K.; Ramirez, J.-M. Response of the respiratory network of mice to hyperthermia. J. Neurophysiol. 2003, 89, 2975–2983. [Google Scholar] [CrossRef]
- Asmussen, E.; Johansen, S.H.; Jørgensen, M.; Nielsen, M. On the nervous factors controlling respiration and circulation during exercise experiments with curarization. Acta Physiol. Scand. 1965, 63, 343–350. [Google Scholar] [CrossRef]
- Kumazawa, T.; Mizumura, K. Thin-fibre receptors responding to mechanical, chemical, and thermal stimulation in the skeletal muscle of the dog. J. Physiol. 1977, 273, 179–194. [Google Scholar] [CrossRef] [PubMed]
- Hertel, H.-C.; Howaldt, B.; Mense, S. Responses of group IV and group III muscle afferents to thermal stimuli. Brain Res. 1976, 113, 201–205. [Google Scholar] [CrossRef]
- Rowell, L.B. Human Circulation: Regulation During Physical Stress; Oxford University Press: Oxford, UK, 1986. [Google Scholar]
- Åstrand, P.-O.; Cuddy, T.E.; Saltin, B.; Stenberg, J. Cardiac output during submaximal and maximal work. J. Appl. Physiol. 1964, 19, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Grimby, G.; Nilsson, N.J.; Saltin, B. Cardiac output during submaximal and maximal exercise in active middle-aged athletes. J. Appl. Physiol. 1966, 21, 1150–1156. [Google Scholar] [CrossRef] [PubMed]
- Richardson, R.S.; Noyszewski, E.A.; Kendrick, K.F.; Leigh, J.S.; Wagner, P.D. Myoglobin O2 desaturation during exercise. Evidence of limited O2 transports. J. Clin. Investig. 1995, 96, 1916–1926. [Google Scholar] [CrossRef] [Green Version]
- Hill, A.V.; Long, C.N.H.; Lupton, H. Muscular exercise, lactic acid and the supply and utilisation of oxygen.—Parts VII–VIII. Proc. R. Soc. London. Ser. B Contain. Pap. Biol. Character 1924, 97, 155–176. [Google Scholar] [CrossRef]
- Åstrand, P.-O.; Saltin, B. Maximal oxygen uptake and heart rate in various types of muscular activity. J. Appl. Physiol. 1961, 16, 977–981. [Google Scholar] [CrossRef]
- Beck, K.C.; Randolph, L.N.; Bailey, K.R.; Wood, C.M.; Snyder, E.M.; Johnson, B.D. Relationship between cardiac output and oxygen consumption during upright cycle exercise in healthy humans. J. Appl. Physiol. 2006, 101, 1474–1480. [Google Scholar] [CrossRef]
- Stöhr, E.J.; González-Alonso, J.; Shave, R. Left ventricular mechanical limitations to stroke volume in healthy humans during incremental exercise. Am. J. Physiol. Circ. Physiol. 2011, 301, H478–H487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmgren, A. Circulatory changes during muscular work in man; with special reference to arterial and central venous pressures in the systemic circulation. Scand. J. Clin. Lab. Investig. 1956, 8 (Suppl. 24), 1–97. [Google Scholar]
- Knight, D.R.; Poole, D.C.; Schaffartzik, W.; Guy, H.J.; Prediletto, R.; Hogan, M.C.; Wagner, P.D. Relationship between body and leg VO2 during maximal cycle ergometry. J. Appl. Physiol. 1992, 73, 1114–1121. [Google Scholar] [CrossRef] [PubMed]
- Pokan, R.; Von Duvillard, S.P.; Hofmann, P.; Smekal, G.; Fruhwald, F.M.; Gasser, R.; Tscha, H.; Baron, R.; Schmid, P.; Bachl, N. Change in left atrial and ventricular dimensions during and immediately after exercise. Med. Sci. Sports Exerc. 2000, 32, 1713–1718. [Google Scholar] [CrossRef] [PubMed]
- Pokan, R.; Hofmann, P.; Von Duvillard, S.P.; BeaufortT, F.; Schumacher, M.; Frauhwald, F.M.; Zweiker, R.; Eber, B.; Gasser, R.; Brandt, D.; et al. Left ventricular function in response to the transition from aerobic to anaerobic metabolism. Med. Sci. Sports Exerc. 1997, 29, 1040–1047. [Google Scholar] [CrossRef]
- González-Alonso, J.; Calbet, J.A.L. Reductions in systemic and skeletal muscle blood flow and oxygen delivery limit maximal aerobic capacity in humans. Circulation 2003, 107, 824–830. [Google Scholar] [CrossRef] [Green Version]
- Rowell, L.B.; Marx, H.J.; Bruce, R.A.; Conn, R.D.; Kusumi, F. Reductions in cardiac output, central blood volume, and stroke volume with thermal stress in normal men during exercise. J. Clin. Investig. 1966, 45, 1801–1816. [Google Scholar] [CrossRef] [Green Version]
- Brengelmann, G.L.; Johnson, J.M.; Hermansen, L.; Rowell, L.B. Altered control of skin blood flow during exercise at high internal temperatures. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1977, 43, 790–794. [Google Scholar] [CrossRef]
- Kellogg, D.L.; Johnson, J.M.; Kenney, W.L.; Pergola, P.E.; Kosiba, W.A. Mechanisms of control of skin blood flow during prolonged exercise in humans. Am. J. Physiol. Circ. Physiol. 1993, 265, H562–H568. [Google Scholar] [CrossRef]
- Kenney, W.L.; Zappe, D.H.; Tankersley, C.G.; Derr, J.A. Effect of systemic yohimbine on the control of skin blood flow during local heating and dynamic exercise. Am. J. Physiol. Circ. Physiol. 1994, 266, H371–H376. [Google Scholar] [CrossRef]
- Savard, G.K.; Nielsen, B.; Laszczynska, J.; Larsen, B.E.; Saltin, B. Muscle blood flow is not reduced in humans during moderate exercise and heat stress. J. Appl. Physiol. 1988, 64, 649–657. [Google Scholar] [CrossRef]
- Nielsen, B.; Savard, G.; Richter, E.A.; Hargreaves, M.; Saltin, B. Muscle blood flow and muscle metabolism during exercise and heat stress. J. Appl. Physiol. 1990, 69, 1040–1046. [Google Scholar] [CrossRef]
- González-Alonso, J.; Mora-Rodríguez, R.; Coyle, E.F. Stroke volume during exercise: Interaction of environment and hydration. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, 321–330. [Google Scholar] [CrossRef] [Green Version]
- Pearson, J.; Low, D.A.; Stöhr, E.; Kalsi, K.; Ali, L.; Barker, H.; González-Alonso, J. Hemodynamic responses to heat stress in the resting and exercising human leg: Insight into the effect of temperature on skeletal muscle blood flow. Am. J. Physiol. Integr. Comp. Physiol. 2011, 300, R663–R673. [Google Scholar] [CrossRef] [Green Version]
- Pitts, G.C.; Johnson, R.E.; Consolazio, F.C. Work in the heat as affected by intake of water, salt and glucose. Am. J. Physiol. Content 1944, 142, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Ladell, W.S.S. The effects of water and salt intake upon the performance of men working in hot and humid environments. J. Physiol. 1955, 127, 11–46. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, M.T.; González-Alonso, J.; Montain, S.J.; Coyle, E.F. Fluid replacement and glucose infusion during exercise prevent cardiovascular drift. J. Appl. Physiol. 1991, 71, 871–877. [Google Scholar] [CrossRef] [Green Version]
- González-Alonso, J.; Mora-Rodriguez, R.; Below, P.R.; Coyle, E.F. Dehydration reduces cardiac output and increases systemic and cutaneous vascular resistance during exercise. J. Appl. Physiol. 1995, 79, 1487–1496. [Google Scholar] [CrossRef]
- González-Alonso, J. Separate and combined influences of dehydration and hyperthermia on cardiovascular responses to exercise. Int. J. Sports Med. 1998, 19, 6–9. [Google Scholar] [CrossRef]
- Sawka, M.N.; Knowlton, R.G.; Critz, J.B. Thermal and circulatory responses to repeated bouts of prolonged running. Med. Sci. Sports Exerc. 1979, 11, 177–180. [Google Scholar]
- Sawka, M.N.; Young, A.J.; Francesconi, P.; Muza, S.R.; Pandolf, K.B. Thermoregulatory and blood responses during exercise at graded hypohydration levels. J. Appl. Physiol. 1985, 59, 1394–1401. [Google Scholar] [CrossRef]
- Arngrimsson, S.; Petitt, D.; Borrani, F.; Skinner, K.; Cureton, K. Hyperthermia and maximal oxygen uptake in men and women. Eur. J. Appl. Physiol. 2004, 92. [Google Scholar] [CrossRef] [PubMed]
- Stöhr, E.J.; González-Alonso, J.; Pearson, J.; Low, D.A.; Ali, L.; Barke, H.; Shave, R. Dehydration reduces left ventricular filling at rest and during exercise independent of twist mechanics. J. Appl. Physiol. 2011, 111, 891–897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, J.; Kalsi, K.K.; Stöhr, E.J.; Low, D.A.; Barker, H.; Ali, L.; González-Alonso, J. Haemodynamic responses to dehydration in the resting and exercising human leg. Eur. J. Appl. Physiol. 2013, 113, 1499–1509. [Google Scholar] [CrossRef] [PubMed]
- Kenefick, R.W.; Mahood, N.V.; Hazzard, M.P.; Quinn, T.J.; Castellani, J.W. Hypohydration effects on thermoregulation during moderate exercise in the cold. Eur. J. Appl. Physiol. 2004, 92, 565–570. [Google Scholar] [CrossRef] [Green Version]
- Montain, S.J.; Coyle, E.F. Influence of graded dehydration on hyperthermia and cardiovascular drift during exercise. J. Appl. Physiol. 1992, 73, 1340–1350. [Google Scholar] [CrossRef]
- Saltin, B. Aerobic and anaerobic work capacity after dehydration. J. Appl. Physiol. 1964, 19, 1114–1118. [Google Scholar] [CrossRef]
- Buskirk, E.R.; Iampietro, P.F.; Bass, D.E. Work performance after dehydration: Effects of physical conditioning and heat acclimatization. J. Appl. Physiol. 1958, 12, 189–194. [Google Scholar] [CrossRef]
- Patterson, S.W.; Starling, E.H. On the mechanical factors which determine the output of the ventricles. J. Physiol. 1914, 48, 357–379. [Google Scholar] [CrossRef]
- Rowell, L.B.; Blackmon, J.R.; Bruce, R.A. Indocyanine green clearance and estimated hepatic blood Flow during mild to maximal exercise in upright man. J. Clin. Investig. 1964, 43, 1677–1690. [Google Scholar] [CrossRef] [Green Version]
- Rowell, L.B.; Blackmon, J.R.; Martin, R.H.; Mazzarella, J.A.; Bruce, R.A. Hepatic clearance of indocyanine green in man under thermal and exercise stresses. J. Appl. Physiol. 1965, 20, 384–394. [Google Scholar] [CrossRef]
- White, H.L.; Rolf, D. Effects of exercise and some other influences on the renal circulation in man. Am. J. Physiol. Content 1948, 152, 505–516. [Google Scholar] [CrossRef] [Green Version]
- Chapman, C.B.; Henschel, A.; Minckler, J.; Forsgren, A.; Keys, A. The effect of exercise on renal plasma flow in normal male subjects. J. Clin. Investig. 1948, 27, 639–644. [Google Scholar] [CrossRef]
- Grimby, G. Renal clearances during prolonged supine exercise at different loads. J. Appl. Physiol. 1965, 20, 1294–1298. [Google Scholar] [CrossRef]
- Levy, M.N. The cardiac and vascular factors that determine systemic blood flow. Circ. Res. 1979, 44, 739–747. [Google Scholar] [CrossRef] [Green Version]
- Guyton, A.C.; Lindsey, A.W.; Kaufmann, B.N. Effect of mean circulatory filling pressure and other peripheral circulatory factors on cardiac output. Am. J. Physiol. Content 1955, 180, 463–468. [Google Scholar] [CrossRef] [Green Version]
- Guyton, A.C.; Lindsey, A.W.; Abernathy, B.; Richardson, T. Venous return at various right atrial pressures and the normal venous return curve. Am. J. Physiol. Content 1957, 189, 609–615. [Google Scholar] [CrossRef] [Green Version]
- Rowell, L.B.; O’Leary, D.S.; Kellogg, D.L. Section 12: Exercise: Regulation and integration of multiple systems. In Handbook of Physiology; Rowell, L.B., Shepherd, J.T., Eds.; Oxford University Press: New York, NY, USA, 1996; pp. 770–840. [Google Scholar]
- Sheriff, D.D.; Zhou, X.P.; Scher, A.M.; Rowell, L.B. Dependence of cardiac filling pressure on cardiac output during rest and dynamic exercise in dogs. Am. J. Physiol. Circ. Physiol. 1993, 265, H316–H322. [Google Scholar] [CrossRef]
- Notarius, C.F.; Magder, S. Central venous pressure during exercise: Role of muscle pump. Can. J. Physiol. Pharmacol. 1996, 74, 647–651. [Google Scholar] [CrossRef]
- Sonnenblick, E.H.; Braunwald, E.; Williams, J.F.; Glick, G. Effects of exercise on myocardial force-velocity relations in intact unanesthetized man: Relative roles of changes in heart rate, sympathetic activity, and ventricular dimensions. J. Clin. Investig. 1965, 44, 2051–2062. [Google Scholar] [CrossRef]
- Heiss, H.W.; Barmeyer, J.; Wink, K.; Hell, G.; Cerny, F.J.; Keul, J.; Reindell, H. Studies on the regulation of myocardial blood flow in man. Basic Res. Cardiol. 1976, 71, 658–675. [Google Scholar] [CrossRef]
- Jørgensen, C.R.; Gobel, F.L.; Taylor, H.L.; Wang, Y. Myocardial blood flow and oxygen consumption during exercise. Ann. N. Y. Acad. Sci. 1977, 301, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Ekström-Jodal, B.; Häggendal, E.; Malmberg, R.; Svedmyr, N. The effect of adrenergic β-receptor blockade on coronary circulation in man during work. Acta Med. Scand. 1972, 191, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, C.R.; Wang, K.; Wang, Y.; Gobel, F.L.; Nelson, R.R.; TAaylor, H.; Gams, F.R.; Vilandre, J.E. Effect of propranolol on myocardial oxygen consumption and its hemodynamic correlates during upright exercise. Circulation 1973, 48, 1173–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmberg, S.; Serzysko, W.; Varnauskas, E. Coronary circulation during heavy exercise in control subjects and patients with coronary heart disease. Acta Med. Scand. 1971, 190, 465–480. [Google Scholar] [CrossRef]
- Nelson, R.R.; Gobel, F.L.; Jøorgensen, C.R.; Wang, K.; Wang, Y.; Taylor, H.L. Hemodynamic predictors of myocardial oxygen consumption during static and dynamic exercise. Circulation 1974, 50, 1179–1189. [Google Scholar] [CrossRef] [Green Version]
- Poliner, L.R.; Dehmer, G.J.; Lewis, S.E.; Parkey, R.W.; Blomqvist, C.G.; Willerson, J.T. Left ventricular performance in normal subjects: A comparison of the responses to exercise in the upright and supine positions. Circulation 1980, 62, 528–534. [Google Scholar] [CrossRef] [Green Version]
- Wyss, C.A.; Koepfli, P.; Mikolajczyk, K.; Burger, C.; von Schulthess, G.K.; Kaufmann, P.A. Bicycle exercise stress in PET for assessment of coronary flow reserve: Repeatability and comparison with adenosine stress. J. Nucl. Med. 2003, 44, 146–154. [Google Scholar]
- Grubbström, J.; Berglund, B.; Kaijser, L. Myocardial blood flow and lactate metabolism at rest and during exercise with reduced arterial oxygen content. Acta Physiol. Scand. 1991, 142, 467–474. [Google Scholar] [CrossRef]
- Wyss, C.A.; Koepfli, P.; Fretz, G.; Seebauer, M.; Schirlo, C.; Kaufmann, P.A. Influence of altitude exposure on coronary flow reserve. Circulation 2003, 108, 1202–1207. [Google Scholar] [CrossRef] [Green Version]
- Fehling, P.C.; Haller, J.M.; Lefferts, W.K.; Hultquist, E.M.; Wharton, M.; Rowland, T.W.; Smith, D.L. Effect of exercise, heat stress and dehydration on myocardial performance. Occup. Med. (Chic. Ill.) 2015, 65, 317–323. [Google Scholar] [CrossRef] [Green Version]
- Travers, G.; González-Alonso, J.; Riding, N.; Nichols, D.; Shaw, A.; Périard, J.D. Exercise heat acclimation has minimal effects on left ventricular volumes, function and systemic hemodynamics in euhydrated and dehydrated trained humans. Am. J. Physiol. Circ. Physiol. 2020, 319, H965–H979. [Google Scholar] [CrossRef]
- Chantler, P.D.; Lakatta, E.G. Arterial-ventricular coupling with aging and disease. Front. Physiol. 2012, 105, 1342–1351. [Google Scholar] [CrossRef] [Green Version]
- Chantler, P.D.; Lakatta, E.G.; Najjar, S.S. Arterial-ventricular coupling: Mechanistic insights into cardiovascular performance at rest and during exercise. J. Appl. Physiol. 2008, 105, 1342–1351. [Google Scholar] [CrossRef] [Green Version]
- Travers, G.; González-Alonso, J.; Riding, N.; Nichols, D.; Shaw, A.; Périard, J.D. Exercise heat acclimation with dehydration does not affect vascular and cardiac volumes or systemic hemodynamics during endurance exercise. Front. Physiol. 2021, 12, 740121. [Google Scholar] [CrossRef]
- Rowell, L.B.; Brengelmann, G.L.; Detry, J.M.; Wyss, C. Venomotor responses to rapid changes in skin temperature in exercising man. J. Appl. Physiol. 1971, 30, 64–71. [Google Scholar] [CrossRef]
- Tatterson, A.J.; Hahn, A.G.; Martini, D.T.; Febbraio, M.A. Effects of heat stress on physiological responses and exercise performance in elite cyclists. J. Sci. Med. Sport 2000, 3, 186–193. [Google Scholar] [CrossRef]
- Montain, S.J.; Coyle, E.F. Fluid ingestion during exercise increases skin blood flow independent of increases in blood volume. J. Appl. Physiol. 1992, 73, 903–910. [Google Scholar] [CrossRef]
- Trinity, J.D.; Pahnke, M.D.; Lee, J.F.; Coyle, E.F. Interaction of hyperthermia and heart rate on stroke volume during prolonged exercise. J. Appl. Physiol. 2010, 109, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.-H.; Allen, J.R.; Hahn, D.; Leary, B.K.; Coyle, E.F. Cardiovascular responses to exercise when increasing skin temperature with narrowing of the core-to-skin temperature gradient. J. Appl. Physiol. 2018, 125, 697–705. [Google Scholar] [CrossRef]
- Chou, T.H.; Akins, J.D.; Crawford, C.K.; Allen, J.R.; Coyle, E.F. Low stroke volume during exercise with hot skin is due to elevated heart rate. Med. Sci. Sports Exerc. 2019, 51, 2025–2032. [Google Scholar] [CrossRef]
- Lee, J.F.; Christmas, K.M.; Machin, D.R.; MClean, B.D.; Coyle, E.F. Warm skin alters cardiovascular responses to cycling after preheating and precooling. Med. Sci. Sports Exerc. 2015, 47, 1168–1176. [Google Scholar] [CrossRef]
- Jose, A.D.; Stitt, F.; Collison, D. The effects of exercise and changes in body temperature on the intrinsic heart rate in man. Am. Heart J. 1970, 79, 488–498. [Google Scholar] [CrossRef]
- Fritzsche, R.G.; Switzer, T.W.; Hodgkinson, B.J.; Coyle, E.F. Stroke volume decline during prolonged exercise is influenced by the increase in heart rate. J. Appl. Physiol. 1999, 86, 799–805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Alonso, J.; Mora-Rodríguez, R.; Below, P.R.; Coyle, E.F. Dehydration markedly impairs cardiovascular function in hyperthermic endurance athletes during exercise. J. Appl. Physiol. 1997, 82, 1229–1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romijn, J.A.; Coyle, E.F.; Sidossis, L.S.; Gastaldelli, A.; Horowitz, J.F.; Endert, E.; Wolfe, R.R. Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am. J. Physiol. Metab. 1993, 265, E380–E391. [Google Scholar] [CrossRef] [Green Version]
- Medbø, J.I.; Tabata, I. Anaerobic energy release in working muscle during 30 s to 3 min of exhausting bicycling. J. Appl. Physiol. 1993, 75, 1654–1660. [Google Scholar] [CrossRef]
- Sahlin, K.; Tonkonogi, M.; Söderlund, K. Energy supply and muscle fatigue in humans. Acta Physiol. Scand. 1998, 162, 261–266. [Google Scholar] [CrossRef]
- Hargreaves, M.; Spriet, L.L. Skeletal muscle energy metabolism during exercise. Nat. Metab. 2020, 2, 817–828. [Google Scholar] [CrossRef]
- Bangsbo, J.; Krustrup, P.; González-Alonso, J.; Saltin, B. ATP production and efficiency of human skeletal muscle during intense exercise: Effect of previous exercise. Am. J. Physiol. Metab. 2001, 280, E956–E964. [Google Scholar] [CrossRef]
- Loon, L.J.C.; Greenhaff, P.L.; Constantin-Teodosiu, D.; Saris, W.H.M.; Wagenmakers, A.J.M. The effects of increasing exercise intensity on muscle fuel utilisation in humans. J. Physiol. 2001, 536, 295–304. [Google Scholar] [CrossRef]
- Richardson, R.S.; Poole, D.C.; Knight, D.R.; Kurdak, S.S.; Hogan, M.C.; Grassi, B.; Johnson, E.C.; Kendrick, K.F.; Erickson, B.K.; Wagner, P.D. High muscle blood flow in man: Is maximal O2 extraction compromised? J. Appl. Physiol. 1993, 75, 1911–1916. [Google Scholar] [CrossRef]
- Rådegran, G. Ultrasound Doppler estimates of femoral artery blood flow during dynamic knee extensor exercise in humans. J. Appl. Physiol. 1997, 83, 1383–1388. [Google Scholar] [CrossRef]
- Saltin, B. Exercise hyperaemia: Magnitude and aspects on regulation in humans. J. Physiol. 2007, 583, 819–823. [Google Scholar] [CrossRef]
- Armstrong, R.B.; Laughlin, M.H. Rat muscle blood flows during high-speed locomotion. J. Appl. Physiol. 1985, 59, 1322–1328. [Google Scholar] [CrossRef]
- Musch, T.I.; Haidet, G.C.; Ordway, G.A.; Longhurst, J.C.; Mitchell, J.H. Training effects on regional blood flow response to maximal exercise in foxhounds. J. Appl. Physiol. 1987, 62, 1724–1732. [Google Scholar] [CrossRef]
- Laughlin, M.H.; Davis, M.J.; Secher, N.H.; Lieshout, J.J.; Arce-Esquivel, A.A.; Simmons, G.H.; Bender, S.B.; Padilla, J.; Bache, R.J.; Merkus, D.; et al. Peripheral Circulation. Compr. Physiol. 2012, 2, 321–447. [Google Scholar] [CrossRef]
- Rowell, L.B. Ideas about control of skeletal and cardiac muscle blood flow (1876–2003): Cycles of revision and new vision. J. Appl. Physiol. 2004, 97, 384–392. [Google Scholar] [CrossRef]
- Poole, D.C.; Pittman, R.N.; Musch, T.I.; Østergaard, L. August Krogh’s theory of muscle microvascular control and oxygen delivery: A paradigm shift based on new data. J. Physiol. 2020, 598, 4473–4507. [Google Scholar] [CrossRef]
- González-Alonso, J.; Calbet, J.A.L.; Boushel, R.; Helge, J.W.; Søndergaard, H.; Munch-Andersen, T.; Hall, G.; Mortensen, S.P.; Secher, N.H. Blood temperature and perfusion to exercising and non-exercising human limbs. Exp. Physiol. 2015, 100, 1118–1131. [Google Scholar] [CrossRef]
- Johnson, J.M.; Minson, C.T.; Kellogg, D.L. Cutaneous Vasodilator and Vasoconstrictor Mechanisms in Temperature Regulation. Comp Physiol. 2014, 4, 33–89. [Google Scholar] [CrossRef]
- Crandall, C.G.; Wilson, T.E. Human Cardiovascular Responses to Passive Heat Stress. Comp Physiol. 2014, 5, 17–43. [Google Scholar] [CrossRef] [Green Version]
- Richardson, R.S.; Noyszewski, E.A.; Leigh, J.S.; Wagner, P.D. Lactate efflux from exercising human skeletal muscle: Role of intracellular PO2. J. Appl. Physiol. 1998, 85, 627–634. [Google Scholar] [CrossRef]
- González-Alonso, J.; Quistorff, B.; Krustrup, P.; Bangsbo, J.; Saltin, B. Heat production in human skeletal muscle at the onset of intense dynamic exercise. J. Physiol. 2000, 524, 603–615. [Google Scholar] [CrossRef]
- Poole, D.C.; Edward, F. Adolph Distinguished Lecture. Contemporary model of muscle microcirculation: Gateway to function and dysfunction. J. Appl. Physiol. 2019, 127, 1012–1033. [Google Scholar] [CrossRef]
- Volianitis, S.; Krustrup, P.; Dawson, E.; Secher, N.H. Arm blood flow and oxygenation on the transition from arm to combined arm and leg exercise in humans. J. Physiol. 2003, 547, 641–648. [Google Scholar] [CrossRef]
- Volianitis, S.; Yoshiga, C.C.; Nissen, P.; Secher, N.H. Effect of fitness on arm vascular and metabolic responses to upper body exercise. Am. J. Physiol. Circ. Physiol. 2004, 286, H1736–H1741. [Google Scholar] [CrossRef]
- Richter, E.A.; Kiens, B.; Hargreaves, M.; Kjaer, M. Effect of arm-cranking on leg blood flow and noradrenaline spillover during leg exercise in man. Acta Physiol. Scand. 1992, 144, 9–14. [Google Scholar] [CrossRef]
- Boushel, R.; Gnaiger, E.; Calbet, J.A.L.; González-Alonso, J.; Wright-Paradis, C.; Sondergaard, H.; Ara, I.; Helge, J.W.; Saltin, B. Muscle mitochondrial capacity exceeds maximal oxygen delivery in humans. Mitochondrion 2011, 11, 303–307. [Google Scholar] [CrossRef]
- Bigland-Ritchie, B.; Cafarelli, E.; Vøllestad, N.K. Fatigue of submaximal static contractions. Acta Physiol. Scand. Suppl. 1986, 556, 137–148. [Google Scholar]
- Garland, S.J.; Enoka, R.M.; Serrano, L.P.; Robinson, G.A. Behavior of motor units in human biceps brachii during a submaximal fatiguing contraction. J. Appl. Physiol. 1994, 76, 2411–2419. [Google Scholar] [CrossRef]
- Taylor, J.L.; Gandevia, S.C. A comparison of central aspects of fatigue in submaximal and maximal voluntary contractions. J. Appl. Physiol. 2008, 104, 542–550. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.D.; Bronks, R.; Smith, P.; Humphries, B. Myoelectric evidence of peripheral muscle fatigue during exercise in severe hypoxia: Some references to m. vastus lateralis myosin heavy chain composition. Eur. J. Appl. Physiol. 1997, 75, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Ahlborg, G.; Felig, P.; Hagenfeldt, L.; Hendler, R.; Wahren, J. Substrate turnover during prolonged exercise in man. J. Clin. Investig. 1974, 53, 1080–1090. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, B.; Strange, S.; Christensen, N.J.; Warberg, J.; Saltin, B. Acute and adaptive responses in humans to exercise in a warm, humid environment. Pflugers Arch. Eur. J. Physiol. 1997, 434, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, B.Y.B.; Hales, J.R.S.; Strange, S.; Christensen, N.J.; Warberg, J.; Saltin, B. Human circulatory and thermoregulatory adaptations with heat acclimation and exercise in a hot, dry environment. J. Physiol. 1993, 460, 467–485. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.C.; Schaffartzik, W.; Knight, D.R.; Derion, T.; Kennedy, B.; Guy, H.J.; Prediletto, R.; Wagner, P.D. Contribution of excising legs to the slow component of oxygen uptake kinetics in humans. J. Appl. Physiol. 1991, 71, 1245–1260. [Google Scholar] [CrossRef]
- Pawelczyk, J.A.; Hanel, B.; Pawelczyk, R.A.; Warberg, J.; Secher, N.H. Leg vasoconstriction during dynamic exercise with reduced cardiac output. J. Appl. Physiol. 1992, 73, 1838–1846. [Google Scholar] [CrossRef]
- Hargreaves, M.; Dillo, P.; Angus, D.; Febbraio, M. Effect of fluid ingestion on muscle metabolism during prolonged exercise. J. Appl. Physiol. 1996, 80, 363–366. [Google Scholar] [CrossRef]
- Chiesa, S.T.; Trangmar, S.J.; Kalsi, K.K.; Rakobowchuk, M.; Banker, D.S.; Lotlikar, M.D.; Ali, L.; González-Alonso, J. Local temperature-sensitive mechanisms are important mediators of limb tissue hyperemia in the heat-stressed human at rest and during small muscle mass exercise. Am. J. Physiol. Circ. Physiol. 2015, 309, H369–H380. [Google Scholar] [CrossRef]
- Koch Esteves, N.; Gibson, O.R.; Khir, A.W.; González-Alonso, J. Regional thermal hyperemia in the human leg: Evidence of the importance of thermosensitive mechanisms in the control of the peripheral circulation. Physiol. Rep. 2021, 9, e14953. [Google Scholar] [CrossRef]
- Chiesa, S.T.; Trangmar, S.J.; González-Alonso, J. Temperature and blood flow distribution in the human leg during passive heat stress. J. Appl. Physiol. 2016, 120, 1047–1058. [Google Scholar] [CrossRef]
- Keller, D.M.; Sander, M.; Stallknecht, B.; Crandall, C.G. α-Adrenergic vasoconstrictor responsiveness is preserved in the heated human leg. J. Physiol. 2010, 588, 3799–3808. [Google Scholar] [CrossRef]
- Romero, S.A.; Gagnon, D.; Adams, A.N.; Cramer, M.N.; Kouda, K.; Crandall, C.G. Acute limb heating improves macro- and microvascular dilator function in the leg of aged humans. Am. J. Physiol. Circ. Physiol. 2017, 312, H89–H97. [Google Scholar] [CrossRef]
- Ferguson, R.A.; Krustrup, P.; Kjær, M.; Mohr, M.; Ball, D.; Bangsbo, J. Effect of temperature on skeletal muscle energy turnover during dynamic knee-extensor exercise in humans. J. Appl. Physiol. 2006, 101, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Dufour, S.P.; Patel, R.P.; Brandon, A.; Teng, X.; Pearson, J.; Barker, H.; Ali, L.; Yuen, A.H.Y.; Smolenski, R.T.; González-Alonso, J. Erythrocyte-dependent regulation of human skeletal muscle blood flow: Role of varied oxyhemoglobin and exercise on nitrite, S -nitrosohemoglobin, and ATP. Am. J. Physiol. Circ. Physiol. 2010, 299, H1936–H1946. [Google Scholar] [CrossRef] [Green Version]
- Shoemaker, J. Time course of brachial artery diameter responses to rhythmic handgrip exercise in humans. Cardiovasc. Res. 1997, 35, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, S.P.; Thaning, P.; Nyberg, M.; Saltin, B.; Hellsten, Y. Local release of ATP into the arterial inflow and venous drainage of human skeletal muscle: Insight from ATP determination with the intravascular microdialysis technique. J. Physiol. 2011, 589, 1847–1857. [Google Scholar] [CrossRef]
- Hearon, C.M.; Richards, J.C.; Racine, M.L.; Luckasen, G.J.; Larson, D.G.; Joyner, M.J.; Dinenno, F.A. Sympatholytic effect of intravascular ATP is independent of nitric oxide, prostaglandins, Na+/K+ -ATPase and K IR channels in humans. J. Physiol. 2017, 595, 5175–5190. [Google Scholar] [CrossRef]
- Ellsworth, M.L.; Sprague, R.S. Regulation of blood flow distribution in skeletal muscle: Role of erythrocyte-released ATP. J. Physiol. 2012, 590, 4985–4991. [Google Scholar] [CrossRef]
- Segal, S.S. Regulation of blood flow in the microcirculation. Microcirculation 2005, 12, 33–45. [Google Scholar] [CrossRef]
- El-Sayed, M.S.; Ali, N.; El-Sayed Ali, Z. Haemorheology in exercise and training. Sports Med. 2005, 35, 649–670. [Google Scholar] [CrossRef]
- Nader, E.; Skinner, S.; Romana, M.; Fort, R.; Lemonne, N.; Guillot, N.; Gauthier, A.; Antoine-Jonville, S.; Renoux, C.; Hardy-Dessources, M.-D.; et al. Blood Rheology: Key parameters, impact on blood flow, role in sickle cell disease and effects of exercise. Front. Physiol. 2019, 10, 1329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortensen, S.P.; Saltin, B. Regulation of the skeletal muscle blood flow in humans. Exp. Physiol. 2014, 99, 1552–1558. [Google Scholar] [CrossRef] [Green Version]
- Marshall, J.M. The roles of adenosine and related substances in exercise hyperaemia. J. Physiol. 2007, 583, 835–845. [Google Scholar] [CrossRef]
- Ellsworth, M.L.; Forrester, T.; Ellis, C.G.; Dietrich, H.H. The erythrocyte as a regulator of vascular tone. Am. J. Physiol. Circ. Physiol. 1995, 269, H2155–H2161. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised estimates for the number of human and bacteria cells in the body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [Green Version]
- Sprague, R.S.; Ellsworth, M.L.; Stephenson, A.H.; Kleinhenz, M.E.; Lonigro, A.J. Deformation-induced ATP release from red blood cells requires CFTR activity. Am. J. Physiol. Circ. Physiol. 1998, 275, H1726–H1732. [Google Scholar] [CrossRef]
- Stamler, J.S.; Jia, L.; Eu, J.P.; McMahon, T.J.; Demchenko, I.T.; Bonaventura, J.; Gernert, K.; Piantadosi, C.A. Blood flow regulation by S -Nitrosohemoglobin in the physiological oxygen gradient. Science 1997, 276, 2034–2037. [Google Scholar] [CrossRef] [Green Version]
- Gladwin, M.T.; Shelhamer, J.H.; Schechter, A.N.; Pease-Fye, M.E.; Waclawiw, M.A.; Panza, J.A.; Ognibene, F.P.; Cannon, R.O. Role of circulating nitrite and S-nitrosohemoglobin in the regulation of regional blood flow in humans. Proc. Natl. Acad. Sci. USA 2000, 97, 11482–11487. [Google Scholar] [CrossRef] [Green Version]
- González-Alonso, J.; Richardson, R.S.; Saltin, B. Exercising skeletal muscle blood flow in humans responds to reduction in arterial oxyhaemoglobin, but not to altered free oxygen. J. Physiol. 2001, 530, 331–341. [Google Scholar] [CrossRef]
- Gonzaález-Alonso, J.; Olsen, D.B.; Saltin, B. Erythrocyte and the regulation of human skeletal muscle blood flow and oxygen delivery. Circ. Res. 2002, 91, 1046–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanada, A.; Sander, M.; González-Alonso, J. Human skeletal muscle sympathetic nerve activity, heart rate and limb haemodynamics with reduced blood oxygenation and exercise. J. Physiol. 2003, 551, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Jagger, J.E.; Bateman, R.M.; Ellsworth, M.L.; Ellis, C.G. Role of erythrocyte in regulating local O2 delivery mediated by hemoglobin oxygenation. Am. J. Physiol. Circ. Physiol. 2001, 280, H2833–H2839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roach, R.C.; Koskolou, M.D.; Calbet, J.A.L.; Saltin, B. Arterial O2 content and tension in regulation of cardiac output and leg blood flow during exercise in humans. Am. J. Physiol. Circ. Physiol. 1999, 276, H438–H445. [Google Scholar] [CrossRef]
- Koskolou, M.D.; Roach, R.C.; Calbet, J.A.L.; Rådegran, G.; Saltin, B. Cardiovascular responses to dynamic exercise with acute anemia in humans. Am. J. Physiol. Circ. Physiol. 1997, 273, H1787–H1793. [Google Scholar] [CrossRef] [Green Version]
- González-Alonso, J.; Mortensen, S.P.; Dawson, E.A.; Secher, N.H.; Damsgaard, R. Erythrocytes and the regulation of human skeletal muscle blood flow and oxygen delivery: Role of erythrocyte count and oxygenation state of haemoglobin. J. Physiol. 2006, 572, 295–305. [Google Scholar] [CrossRef]
- Hellsten, Y.; Nyberg, M.; Mortensen, S.P. Contribution of intravascular versus interstitial purines and nitric oxide in the regulation of exercise hyperaemia in humans. J. Physiol. 2012, 590, 5015–5023. [Google Scholar] [CrossRef] [Green Version]
- Richardson, R.S.; Noyszewski, E.A.; Saltin, B.; González-Alonso, J. Effect of mild carboxy-hemoglobin on exercising skeletal muscle: Intravascular and intracellular evidence. Am. J. Physiol. Integr. Comp. Physiol. 2002, 283, R1131–R1139. [Google Scholar] [CrossRef]
- Clifford, P.S.; Hellsten, Y. Vasodilatory mechanisms in contracting skeletal muscle. J. Appl. Physiol. 2004, 97, 393–403. [Google Scholar] [CrossRef]
- Kalsi, K.K.; Chiesa, S.T.; Trangmar, S.J.; Ali, L.; Lotlikar, M.D.; González-Alonso, J. Mechanisms for the control of local tissue blood flow during thermal interventions: Influence of temperature-dependent ATP release from human blood and endothelial cells. Exp. Physiol. 2017, 102, 228–244. [Google Scholar] [CrossRef]
- Heinonen, I.; Brothers, R.M.; Kemppainen, J.; Knuuti, J.; Kalliokoski, K.K.; Crandall, C.G. Local heating, but not indirect whole body heating, increases human skeletal muscle blood flow. J. Appl. Physiol. 2011, 111, 818–824. [Google Scholar] [CrossRef]
- Nemkov, T.; Skinner, S.C.; Nader, E.; Stefanoni, D.; Robert, M.; Cendali, F.; Stauffer, E.; Cibiel, A.; Boisson, C.; Connes, P.; et al. Acute cycling exercise induces changes in red blood cell deformability and membrane lipid remodeling. Int. J. Mol. Sci. 2021, 22, 896. [Google Scholar] [CrossRef]
- van Beaumont, W.; Underkofler, S.; van Beaumont, S. Erythrocyte volume, plasma volume, and acid-base changes in exercise and heat dehydration. J. Appl. Physiol. 1981, 50, 1255–1262. [Google Scholar] [CrossRef]
- Mitchell, J.H. Cardiovascular control during exercise: Central and reflex neural mechanisms. Am. J. Cardiol. 1985, 55, 34D–41D. [Google Scholar] [CrossRef]
- Mitchell, J.H. Neural circulatory control during exercise: Early insights. Exp. Physiol. 2013, 98, 867–878. [Google Scholar] [CrossRef]
- Fisher, J.P.; Young, C.N.; Fadel, P.J. Autonomic adjustments to exercise in humans. Compr. Physiol. 2015, 5, 475–512. [Google Scholar] [CrossRef]
- Calbet, J.A.L.; Jensen-Urstad, M.; Van Hall, G.; Holmberg, H.-C.; Rosdahl, H.; Saltin, B. Maximal muscular vascular conductances during whole body upright exercise in humans. J. Physiol. 2004, 558, 319–331. [Google Scholar] [CrossRef]
- Furst, B. The Heart and Circulation, 2nd ed.; Springer International Publishing: Cham, Switzerland, 2020; Volume 558, ISBN 978-3-030-25061-4. [Google Scholar]
- Joyce, W.; Wang, T. What determines systemic blood flow in vertebrates? J. Exp. Biol. 2020, 223, jeb215335. [Google Scholar] [CrossRef]
- Guyton, A.C. Cardiac output, venous return, and their regulation- an integrative analysis of the circulation. In Textbook of Medical Physiology; Guyton, A.C., Ed.; W.B. Saunders Company: Philadelphia, PA, USA, 1969; pp. 337–352. [Google Scholar]
- Bada, A.A.; Svendsen, J.H.; Secher, N.H.; Saltin, B.; Mortensen, S.P. Peripheral vasodilatation determines cardiac output in exercising humans: Insight from atrial pacing. J. Physiol. 2012, 590, 2051–2060. [Google Scholar] [CrossRef]
- Ross, J.; Linhart, J.W.; Brauwald, E. Effects of changing heart rate in man by electrical stimulation of the right atrium. studies at rest, during exercise, and with isoproterenol. Circulation 1965, 32, 549–558. [Google Scholar] [CrossRef] [Green Version]
- Stein, E.; Damato, A.N.; Kosowsky, B.D.; Lau, S.H.; Lister, J.W. The relation of heart rate to cardiovascular dynamics. Circulation 1966, 33, 925–932. [Google Scholar] [CrossRef] [Green Version]
- Ilkka, H.; Bengt, S.; Jukka, K.; Sipilä, H.T.; Vesa, O.; Pirjo, N.; Juhani, K.; Kari, K.; Ylva, H. Skeletal muscle blood flow and oxygen uptake at rest and during exercise in humans: A pet study with nitric oxide and cyclooxygenase inhibition. Am. J. Physiol. Circ. Physiol. 2011, 300, H1510–H1517. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.D.; Berne, R.M. Intrinsic regulation of skeletal muscle blood flow. Circ. Res. 1964, 14, 126–138. [Google Scholar] [CrossRef] [Green Version]
- Duling, B.R.; Berne, R.M. Propagated vasodilation in the microcirculation of the hamster cheek pouch. Circ. Res. 1970, 26, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Rosenmeier, J.B.; Yegutkin, G.G.; González-Alonso, J. Activation of ATP/UTP-selective receptors increases blood flow and blunts sympathetic vasoconstriction in human skeletal muscle. J. Physiol. 2008, 586, 4993–5002. [Google Scholar] [CrossRef]
- Calbet, J.A.L.; Lundby, C.; Sander, M.; Robach, P.; Saltin, B.; Boushel, R. Effects of ATP-induced leg vasodilation on VO2 peak and leg O2 extraction during maximal exercise in humans. Am. J. Physiol. Integr. Comp. Physiol. 2006, 291, R447–R453. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, S.P.; González-Alonso, J.; Damsgaard, R.; Saltin, B.; Hellsten, Y. Inhibition of nitric oxide and prostaglandins, but not endothelial-derived hyperpolarizing factors, reduces blood flow and aerobic energy turnover in the exercising human leg. J. Physiol. 2007, 581, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Calbet, J.A.L.; Losa-Reyna, J.; Torres-Peralta, R.; Rasmussen, P.; Ponce-González, J.G.; Sheel, A.W.; de la Calle-Herrero, J.; Guadalupe-Grau, A.; Morales-Alamo, D.; Fuentes, T.; et al. Limitations to oxygen transport and utilization during sprint exercise in humans: Evidence for a functional reserve in muscle O2 diffusing capacity. J. Physiol. 2015, 593, 4649–4664. [Google Scholar] [CrossRef] [Green Version]
- Steiner, R. Introducing Anthroposopical Medicine: Lecture of Mmarch 22 1920, Donarch, Switzerland; SteinerBooks: Hudson, NY, USA, 1999. [Google Scholar]
- Havlicek, H. Arbeitet das Herz wie eine Druckpumpe oder wie ein Stoßheber. Arch. Kreislaufforsch. 1937, 1, 188–224. [Google Scholar] [CrossRef]
- Furst, B. The heart: Pressure-propulsion pump or organ of impedance? J. Cardiothorac. Vasc. Anesth. 2015, 29, 1688–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manteuffel-Szoege, L. Energy sources of blood circulation and the mechanical action of the heart. Thorax 1960, 15, 47–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manteuffel-Szoege, L.; Michalowski, J.; Grundman, J.; Pacocha, W. On the possibility of blood circulation continuing after stopping the heart. J. Cardiovasc. Surg. (Torino) 1966, 7, 201–208. [Google Scholar]
- Caldwell, H.G.; Carr, J.M.J.R.; Minhas, J.S.; Swenson, E.R.; Ainslie, P.N. Acid–base balance and cerebrovascular regulation. J. Physiol. 2021, 599, 5337–5359. [Google Scholar] [CrossRef]
- Gourine, A.V.; Kasymov, V.; Marina, N.; Tang, F.; Figueiredo, M.F.; Lane, S.; Teschemacher, A.G.; Spyer, K.M.; Deisseroth, K.; Kasparov, S. Astrocytes control breathing through pH-ependent release of ATP. Science 2010, 329, 571–575. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.C.; Ellenberger, H.H.; Ballanyi, K.; Richter, D.W.; Feldman, J.L. Pre-Bötzinger complex: A brainstem region that may generate respiratory rhythm in mammals. Science 1991, 254, 726–729. [Google Scholar] [CrossRef]
- Sheikhbahaei, S.; Turovsky, E.A.; Hosford, P.S.; Hadjihambi, A.; Theparambil, S.M.; Liu, B.; Marina, N.; Teschemacher, A.G.; Kasparov, S.; Smith, J.C.; et al. Astrocytes modulate brainstem respiratory rhythm-generating circuits and determine exercise capacity. Nat. Commun. 2018, 9, 370. [Google Scholar] [CrossRef]
- Noakes, T.D. Maximal oxygen uptake: “classical” versus “contemporary” viewpoints: A rebuttal. Med. Sci. Sports Exerc. 1998, 30, 1381–1398. [Google Scholar] [CrossRef]
- Noakes, T.D.; Marino, F.E. Arterial oxygenation, central motor output and exercise performance in humans. J. Physiol. 2007, 585, 919–921. [Google Scholar] [CrossRef]
- Noakes, T.D. Challenging beliefs: Ex Africa semper aliquid novi. Med. Sci. Sports Exerc. 1997, 29, 571–590. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Travers, G.; Kippelen, P.; Trangmar, S.J.; González-Alonso, J. Physiological Function during Exercise and Environmental Stress in Humans—An Integrative View of Body Systems and Homeostasis. Cells 2022, 11, 383. https://doi.org/10.3390/cells11030383
Travers G, Kippelen P, Trangmar SJ, González-Alonso J. Physiological Function during Exercise and Environmental Stress in Humans—An Integrative View of Body Systems and Homeostasis. Cells. 2022; 11(3):383. https://doi.org/10.3390/cells11030383
Chicago/Turabian StyleTravers, Gavin, Pascale Kippelen, Steven J. Trangmar, and José González-Alonso. 2022. "Physiological Function during Exercise and Environmental Stress in Humans—An Integrative View of Body Systems and Homeostasis" Cells 11, no. 3: 383. https://doi.org/10.3390/cells11030383