


Cutaneous Injection of Resiniferatoxin Completely Alleviates and Prevents Nerve-Injury-Induced Neuropathic Pain
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. L5 Nerve-Injury-Induced Neuropathic Pain Model, Intraplantar Injection of RTX/Vehicle and Behavioral Testing
2.2.1. Surgery Rats
2.2.2. Behavioral Testing
2.3. Immunohistochemistry
2.3.1. Spinal Cord and DRG Staining
2.3.2. c-Fos Staining in the Spinal Cord (ABC Method)
2.3.3. Skin Staining
2.4. Quantitative Real-Time Polymerase Chain Reaction
2.5. Western Blot
2.6. Imaging and Data Analysis
2.7. Dorsal Root Ganglion
2.8. Antibody Characterization
2.9. Statistical Analysis
3. Results
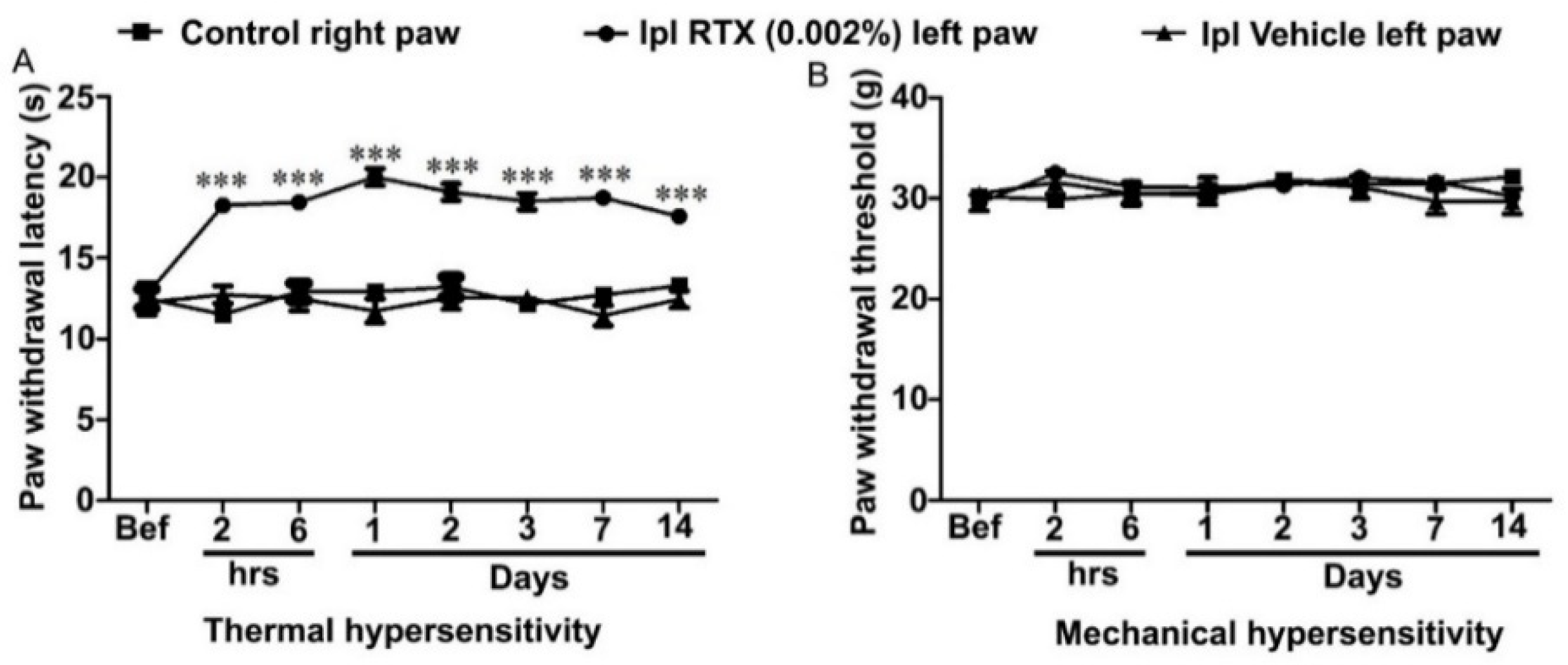
3.1. Behavioural and Analgesic Effects of Plantar Injection RTX in Normal Rats
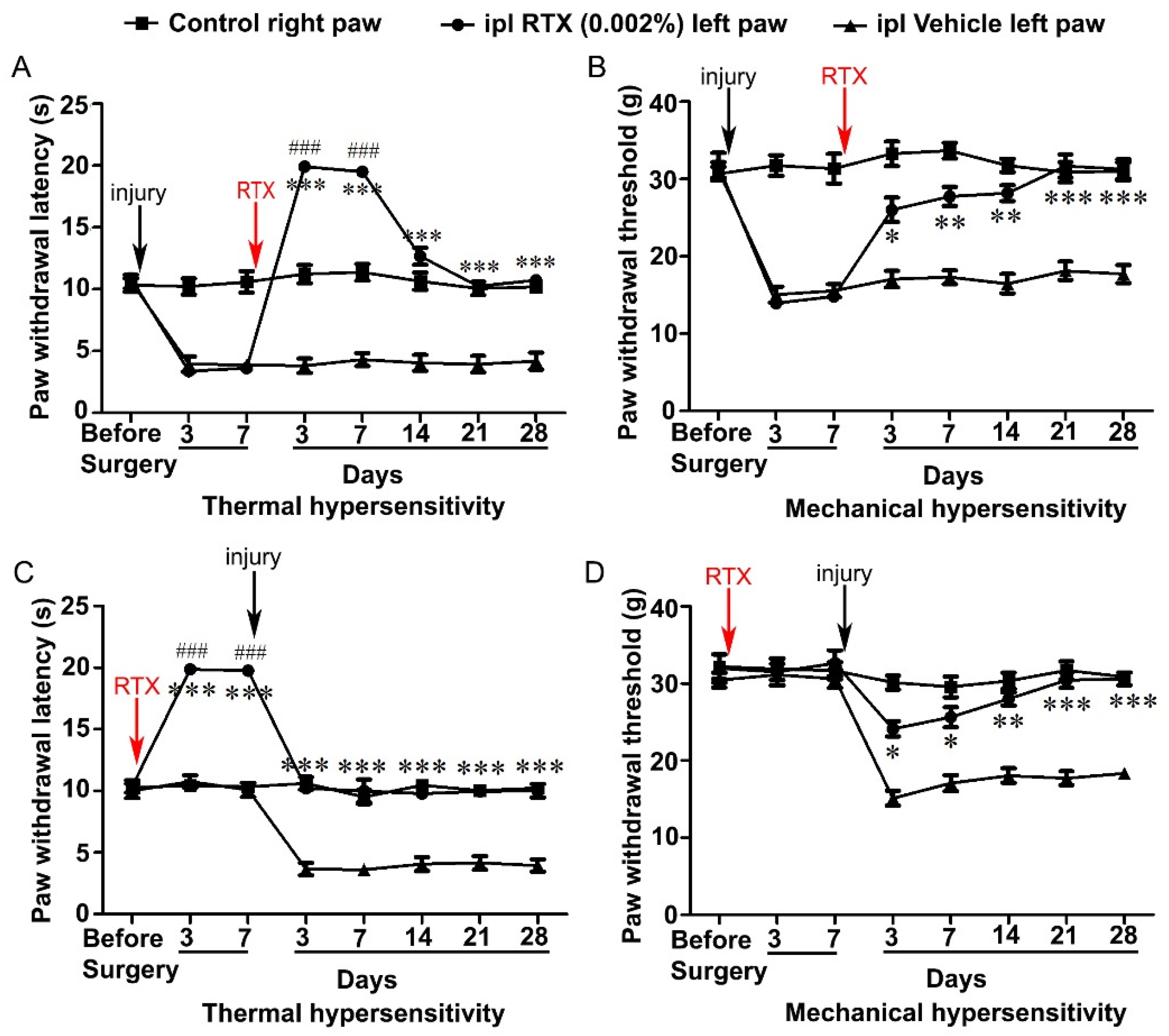
3.2. Intraplantar Injection of RTX Completely Alleviates Thermal and Mechanical Hypersensitivity Caused by L5 Nerve Injury
3.3. Intraplantar Injection of RTX Completely Prevents the Development of Thermal and Mechanical Hypersensitivity Caused by L5 Nerve Injury
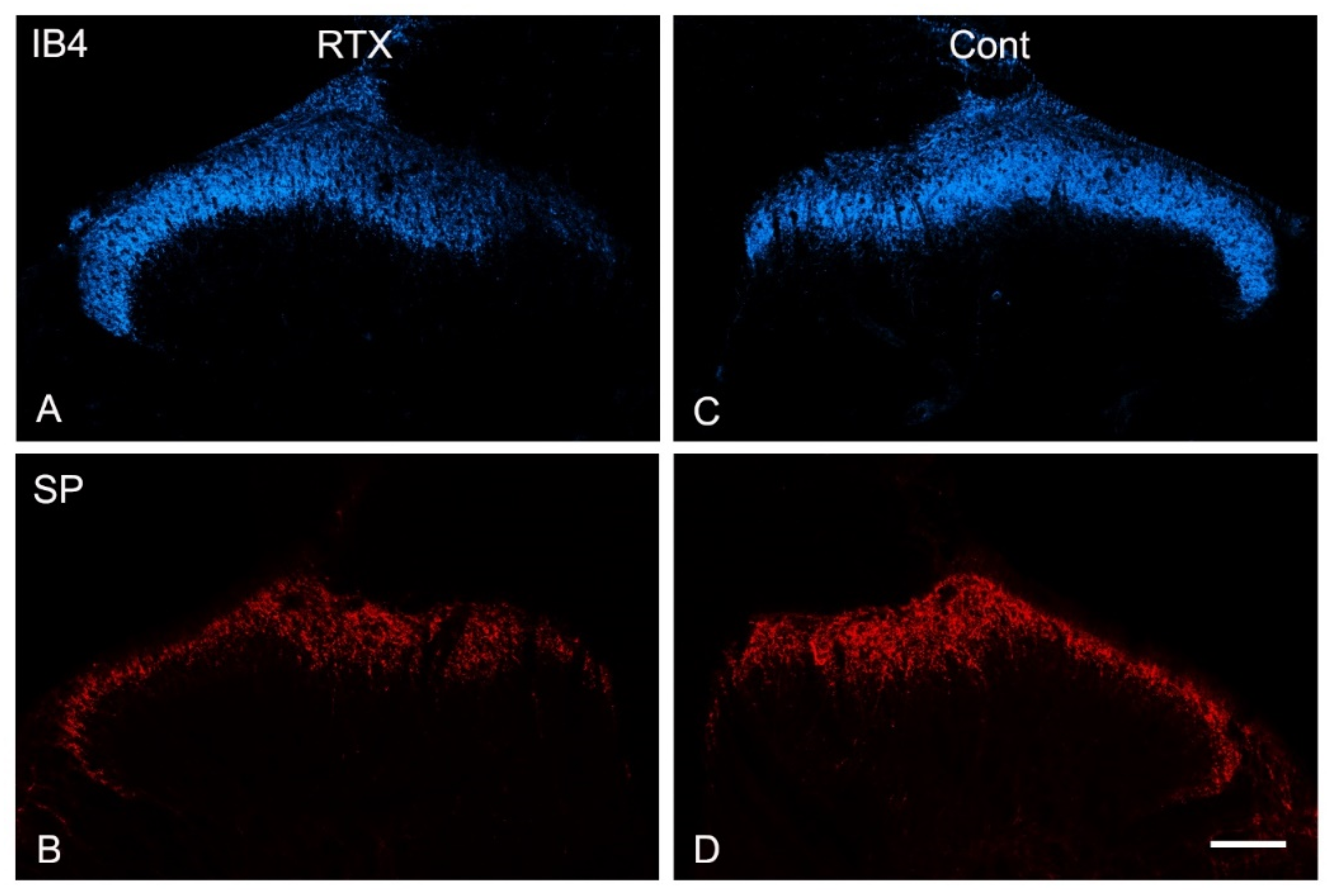
3.4. Effects of Intraplantar Injection of RTX on the Immunoreactivity of TRPV1, IB4, CGRP, SP and VIP in the Dorsal Horn of the Spinal Cord
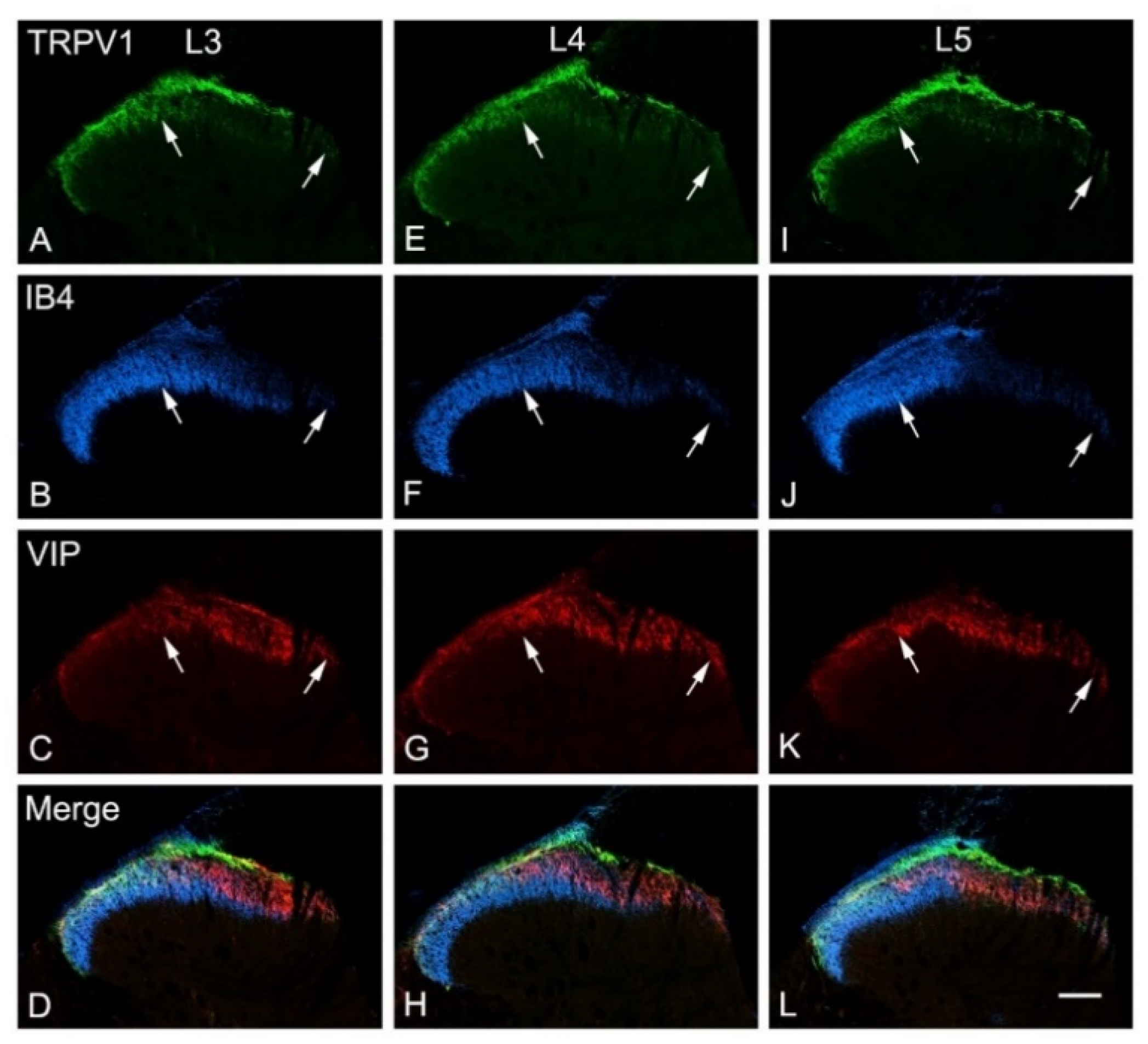
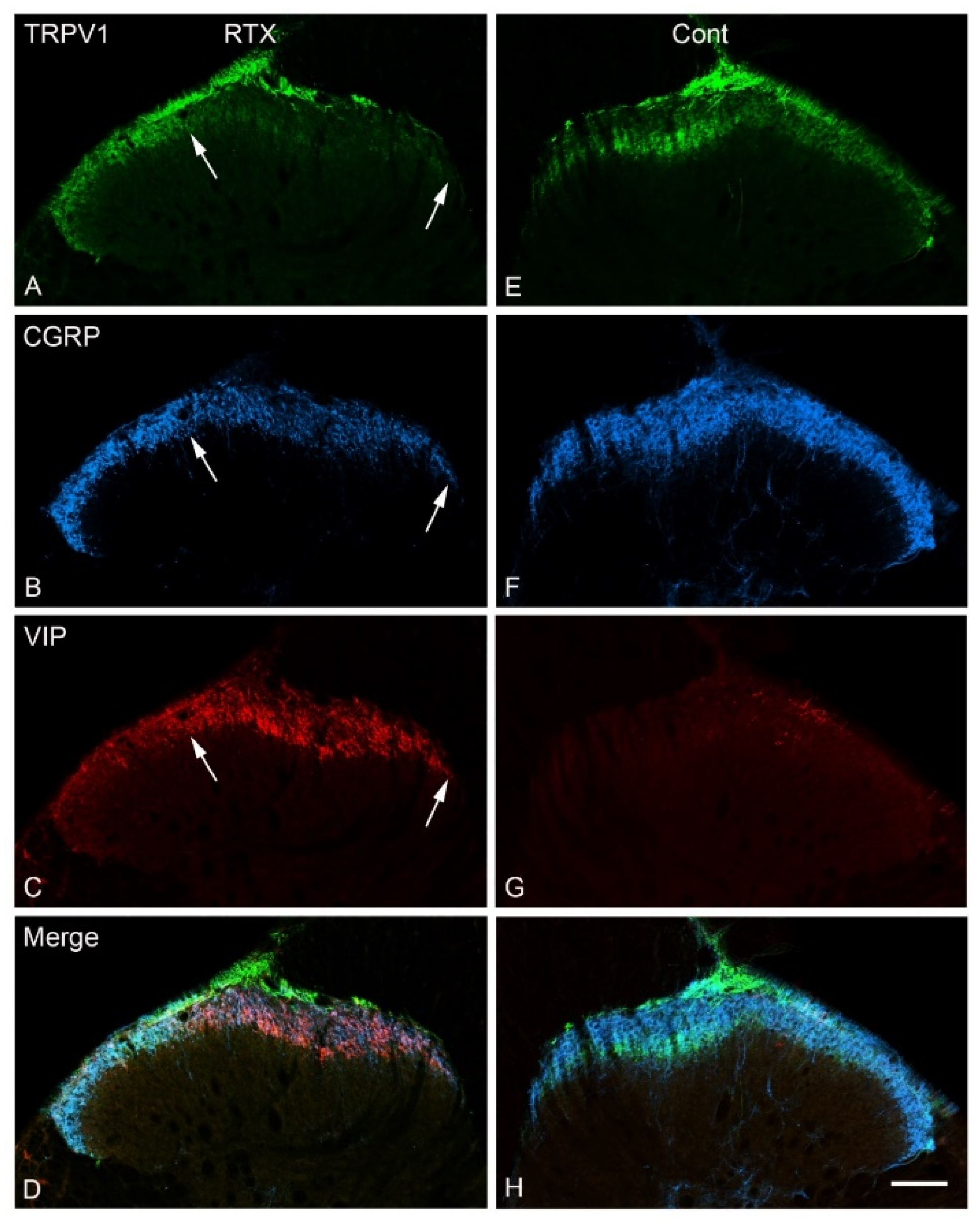
3.5. Effects of Intraplantar Injection of RTX on the Immunoreactivity of TRPV1, IB4, CGRP and VIP in the DRGs
3.6. Effects of Intraplantar Injection of RTX on the mRNA and Protein Expression Level of TRPV1 in the DRG
3.7. Effects of Intraplantar Injection of RTX on the c-Fos Activation in the Spinal Cord
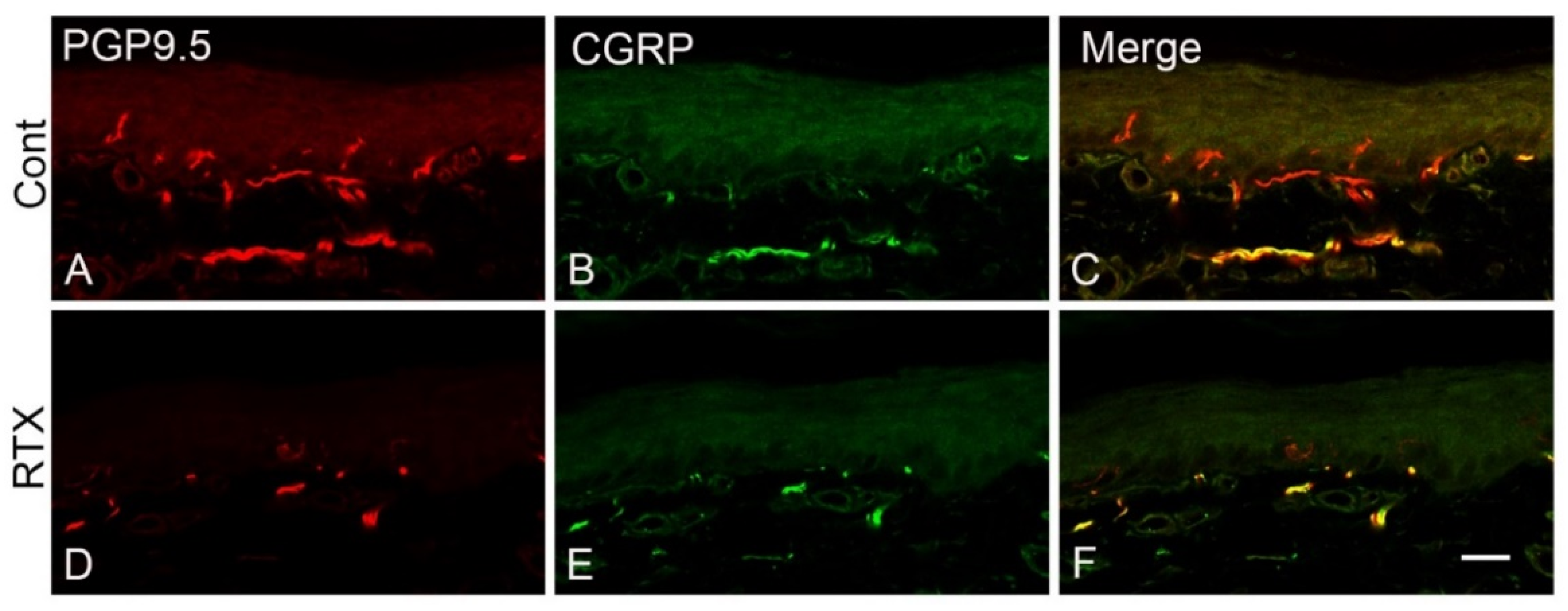
3.8. Effects of Intraplantar Injection of RTX on the Immunoreactivity of PGP 9.5 and CGRP in the Skin of the Hind Paw of Rats
4. Discussion
4.1. Neurochemical Changes Following Intraplantar Injection of RTX
4.2. Intraplantar RTX Injection Caused VIP Upregulation in the Primary Sensory Neurons
4.3. Perineural vs. Intraplantar RTX Application in Treating Nerve-Injury-Induced Neuropathic Pain
4.4. Mechanisms of Analgesic Action of RTX
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Campbell, J.N.; Meyer, R.A. Mechanisms of neuropathic pain. Neuron 2006, 52, 77–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ossipov, M.H.; Lai, J.; Porreca, F. Mechanisms of Experimental Neuropathic Pain: Integration from Animal Models. In Wall and Melzack’s Textbook of Pain, 5th ed.; Wall, P.D., McMahon, S.B., Koltzenburg, M., Eds.; Churchhill Livingstone, Elsevier Limited: London, UK, 2006; pp. 929–946. [Google Scholar]
- Kim, S.H.; Chung, J.M. An experimental model for peripheral neuropathy produced by segmental spinal nerve ligation in the rat. Pain 1992, 50, 355–363. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, A.B.; Dworkin, R.H. Treatment of neuropathic pain: An overview of recent guidelines. Am. J. Med. 2009, 122, S22–S32. [Google Scholar] [CrossRef] [PubMed]
- Finnerup, N.B.; Attal, N.; Haroutounian, S.; McNicol, E.; Baron, R.; Dworkin, R.H.; Gilron, I.; Haanpää, M.; Hansson, P.; Jensen, T.S. Pharmacotherapy for neuropathic pain in adults: A systematic review and meta-analysis. Lancet Neurol. 2015, 14, 162–173. [Google Scholar] [CrossRef] [Green Version]
- Baron, R.; Binder, A.; Wasner, G. Neuropathic pain: Diagnosis, pathophysiological mechanisms, and treatment. Lancet Neurol. 2010, 9, 807–819. [Google Scholar] [CrossRef]
- Cruccu, G.; Truini, A. A review of neuropathic pain: From guidelines to clinical practice. Pain Ther. 2017, 6, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Caterina, M.J.; Leffler, A.; Malmberg, A.B.; Martin, W.; Trafton, J.; Petersen-Zeitz, K.; Koltzenburg, M.; Basbaum, A.; Julius, D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 2000, 288, 306–313. [Google Scholar] [CrossRef]
- Davis, J.B.; Gray, J.; Gunthorpe, M.J.; Hatcher, J.P.; Davey, P.T.; Overend, P.; Harries, M.H.; Latcham, J.; Clapham, C.; Atkinson, K. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature 2000, 405, 183–187. [Google Scholar] [CrossRef]
- Szallasi, A.; Blumberg, P. Resiniferatoxin, a phorbol-related diterpene, acts as an ultrapotent analog of capsaicin, the irritant constituent in red pepper. Neuroscience 1989, 30, 515–520. [Google Scholar] [CrossRef] [Green Version]
- Javed, H.; Rehmathulla, S.; Tariq, S.; Emerald, B.S.; Ljubisavljevic, M.; Shehab, S. Perineural application of resiniferatoxin on uninjured L3 and L4 nerves completely alleviates thermal and mechanical hypersensitivity following L5 nerve injury in rats. J. Comp. Neurol. 2020, 528, 2195–2217. [Google Scholar] [CrossRef]
- Pecze, L.; Blum, W.; Schwaller, B. Mechanism of capsaicin receptor TRPV1-mediated toxicity in pain-sensing neurons focusing on the effects of Na+/Ca2+ fluxes and the Ca2+-binding protein calretinin. Biochim. Biophys. Acta Mol. Cell Res. 2013, 1833, 1680–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, K.; Bates, B.D.; Keller, J.M.; Lopez, M.; Scholl, L.; Navarro, J.; Madian, N.; Haspel, G.; Nemenov, M.I.; Iadarola, M.J. Ablation of rat TRPV1-expressing Adelta/C-fibers with resiniferatoxin: Analysis of withdrawal behaviors, recovery of function and molecular correlates. Mol. Pain 2010, 6, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caterina, M.J.; Julius, D. The vanilloid receptor: A molecular gateway to the pain pathway. Annu. Rev. Neurosci. 2001, 24, 487–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, A.; Vulchanova, L.; Wang, J.; Li, X.; Elde, R. Immunocytochemical localization of the vanilloid receptor 1 (VR1): Relationship to neuropeptides, the P2X3 purinoceptor and IB4 binding sites. Eur. J. Neurosci. 1999, 11, 946–958. [Google Scholar] [CrossRef] [PubMed]
- Michael, G.J.; Priestley, J.V. Differential expression of the mRNA for the vanilloid receptor subtype 1 in cells of the adult rat dorsal root and nodose ganglia and its downregulation by axotomy. J. Neurosci. 1999, 19, 1844–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, M.-K.; Campbell, J.N. Use of capsaicin to treat pain: Mechanistic and therapeutic considerations. Pharmaceuticals 2016, 9, 66. [Google Scholar] [CrossRef] [Green Version]
- Derry, S.; Moore, R.A. Topical capsaicin (low concentration) for chronic neuropathic pain in adults. Cochrane Database Syst. Rev. 2012, 9, CD010111. [Google Scholar] [CrossRef]
- Anand, P.; Bley, K. Topical capsaicin for pain management: Therapeutic potential and mechanisms of action of the new high-concentration capsaicin 8% patch. Br. J. Anaesth. 2011, 107, 490–502. [Google Scholar] [CrossRef] [Green Version]
- Iadarola, M.J.; Brown, D.C.; Nahama, A.; Sapio, M.R.; Mannes, A.J. Pain treatment in the companion canine model to validate rodent results and incentivize the transition to human clinical trials. Front. Pharmacol. 2021, 12, 1923. [Google Scholar] [CrossRef]
- Sapio, M.R.; Neubert, J.K.; LaPaglia, D.M.; Maric, D.; Keller, J.M.; Raithel, S.J.; Rohrs, E.L.; Anderson, E.M.; Butman, J.A.; Caudle, R.M. Pain control through selective chemo-axotomy of centrally projecting TRPV1+ sensory neurons. J. Clin. Investig. 2018, 128, 1657–1670. [Google Scholar] [CrossRef]
- Fattori, V.; Hohmann, M.S.; Rossaneis, A.C.; Pinho-Ribeiro, F.A.; Verri Jr, W.A. Capsaicin: Current understanding of its mechanisms and therapy of pain and other pre-clinical and clinical uses. Molecules 2016, 21, 844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jancsó, G. Pathobiological reactions of C-fibre primary sensory neurones to peripheral nerve injury. Exp. Physiol. 1992, 77, 405–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.-Q.; Premkumar, L.S. Ablation and regeneration of peripheral and central TRPV1 expressing nerve terminals and the consequence of nociception. Open Pain J. 2015, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Karai, L.; Brown, D.C.; Mannes, A.J.; Connelly, S.T.; Brown, J.; Gandal, M.; Wellisch, O.M.; Neubert, J.K.; Olah, Z.; Iadarola, M.J. Deletion of vanilloid receptor 1_expressing primary afferent neurons for pain control. J. Clin. Investig. 2004, 113, 1344–1352. [Google Scholar] [CrossRef]
- Neubert, J.K.; Mannes, A.J.; Karai, L.J.; Jenkins, A.C.; Zawatski, L.; Abu-Asab, M.; Iadarola, M.J. Perineural resiniferatoxin selectively inhibits inflammatory hyperalgesia. Mol. Pain 2008, 4, 1744-8069-4-3. [Google Scholar] [CrossRef] [Green Version]
- Neubert, J.K.; Karai, L.; Jun, J.H.; Kim, H.-S.; Olah, Z.; Iadarola, M.J. Peripherally induced resiniferatoxin analgesia. Pain 2003, 104, 219–228. [Google Scholar] [CrossRef]
- Salas, M.M.; Clifford, J.L.; Hayden, J.R.; Iadarola, M.J.; Averitt, D.L. Local resiniferatoxin induces long-lasting analgesia in a rat model of full thickness thermal injury. Pain Med. 2017, 18, 2453–2465. [Google Scholar] [CrossRef] [Green Version]
- Tender, G.C.; Li, Y.-Y.; Cui, J.-G. Vanilloid receptor 1-positive neurons mediate thermal hyperalgesia and tactile allodynia. Spine J. 2008, 8, 351–358. [Google Scholar] [CrossRef]
- Tender, G.C.; Walbridge, S.; Olah, Z.; Karai, L.; Iadarola, M.; Oldfield, E.H.; Lonser, R.R. Selective ablation of nociceptive neurons for elimination of hyperalgesia and neurogenic inflammation. J. Neurosurg. 2005, 102, 522–525. [Google Scholar] [CrossRef] [Green Version]
- Shehab, S.A.-D.S. Fifth lumbar spinal nerve injury causes neurochemical changes in corresponding as well as adjacent spinal segments: A possible mechanism underlying neuropathic pain. J. Chem. Neuroanat. 2014, 55, 38–50. [Google Scholar] [CrossRef]
- Llewellyn-Smith, I.J.; Minson, J.B. Complete penetration of antibodies into vibratome sections after glutaraldehyde fixation and ethanol treatment: Light and electron microscopy for neuropeptides. J. Histochem. Cytochem. 1992, 40, 1741–1749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emerald, B.S.; Chng, K.; Masuda, S.; Sloboda, D.M.; Vickers, M.H.; Kambadur, R.; Gluckman, P.D. Gene expression profiling in the Cynomolgus macaque Macaca fascicularis shows variation within the normal birth range. BMC Genom. 2011, 12, 509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Baiou, D.; Santha, P.; Avelino, A.; Charrua, A.; Bacskai, T.; Matesz, K.; Cruz, F.; Nagy, I. Neurochemical characterization of insulin receptor-expressing primary sensory neurons in wild-type and vanilloid type 1 transient receptor potential receptor knockout mice. J. Comp. Neurol. 2007, 503, 334–347. [Google Scholar] [CrossRef]
- Shehab, S.; Spike, R.; Todd, A. Evidence against cholera toxin B subunit as a reliable tracer for sprouting of primary afferents following peripheral nerve injury. Brain Res. 2003, 964, 218–227. [Google Scholar] [CrossRef]
- Reilly, D.; Ferdinando, D.; Johnston, C.; Shaw, C.; Buchanan, K.; Green, M. The epidermal nerve fibre network: Characterization of nerve fibres in human skin by confocal microscopy and assessment of racial variations. Br. J. Dermatol. 1997, 137, 163–170. [Google Scholar] [CrossRef]
- Shehab, S.; Javed, H.; Khan, G. Somatotopic localization of c-Fos expression in the spinal cord in response to noxious heat sensation. Front. Neuroanat. 2022, 16, 1035257. [Google Scholar] [CrossRef]
- Molander, C.; Grant, G. Laminar distribution and somatotopic organization of primary afferent fibers from hindlimb nerves in the dorsal horn. A study by transganglionic transport of horseradish peroxidase in the rat. Neuroscience 1986, 19, 297–312. [Google Scholar] [CrossRef]
- Shehab, S.A.; Hughes, D.I. Simultaneous identification of unmyelinated and myelinated primary somatic afferents by co-injection of isolectin B4 and Cholera toxin subunit B into the sciatic nerve of the rat. J. Neurosci. Methods 2011, 198, 213–221. [Google Scholar] [CrossRef]
- Shehab, S.A.S. Acute and chronic sectioning of fifth lumbar spinal nerve has equivalent effects on the primary afferents of sciatic nerve in rat spinal cord. J. Comp. Neurol. 2009, 517, 481–492. [Google Scholar] [CrossRef]
- Shehab, S.A.; Al-Marashda, K.; Al-Zahmi, A.; Abdul-Kareem, A.; Al-Sultan, M.A. Unmyelinated primary afferents from adjacent spinal nerves intermingle in the spinal dorsal horn: A possible mechanism contributing to neuropathic pain. Brain Res. 2008, 1208, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Grant, G. Projection patterns of primary sensory neurons studied by transganglionic methods: Somatotopy and target-related organization. Brain Res. Bull 1993, 30, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Bernardini, N.; Neuhuber, W.; Reeh, P.; Sauer, S. Morphological evidence for functional capsaicin receptor expression and calcitonin gene-related peptide exocytosis in isolated peripheral nerve axons of the mouse. Neuroscience 2004, 126, 585–590. [Google Scholar] [CrossRef] [PubMed]
- Simone, D.A.; Nolano, M.; Johnson, T.; Wendelschafer-Crabb, G.; Kennedy, W.R. Intradermal injection of capsaicin in humans produces degeneration and subsequent reinnervation of epidermal nerve fibers: Correlation with sensory function. J. Neurosci. 1998, 18, 8947–8959. [Google Scholar] [CrossRef] [Green Version]
- Nolano, M.; Simone, D.A.; Wendelschafer-Crabb, G.; Johnson, T.; Hazen, E.; Kennedy, W.R. Topical capsaicin in humans: Parallel loss of epidermal nerve fibers and pain sensation. Pain 1999, 81, 135–145. [Google Scholar] [CrossRef]
- Kissin, I.; Freitas, C.F.; Bradley, E.L., Jr. Perineural resiniferatoxin prevents the development of hyperalgesia produced by loose ligation of the sciatic nerve in rats. Anesth. Analg. 2007, 104, 1210–1216. [Google Scholar] [CrossRef]
- Kissin, I.; Freitas, C.F.; Mulhern, H.L.; DeGirolami, U. Sciatic nerve block with resiniferatoxin: An electron microscopic study of unmyelinated fibers in the rat. Anesth. Analg. 2007, 105, 825–831. [Google Scholar] [CrossRef]
- Ragé, M.; Van Acker, N.; Facer, P.; Shenoy, R.; Knaapen, M.W.; Timmers, M.; Streffer, J.; Anand, P.; Meert, T.; Plaghki, L. The time course of CO2 laser-evoked responses and of skin nerve fibre markers after topical capsaicin in human volunteers. Clin. Neurophysiol. 2010, 121, 1256–1266. [Google Scholar] [CrossRef]
- Oszlács, O.; Jancsó, G.; Kis, G.; Dux, M.; Sántha, P. Perineural capsaicin induces the uptake and transganglionic transport of choleratoxin b subunit by nociceptive c-fiber primary afferent neurons. Neuroscience 2015, 311, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Tran, P.V.; Johns, M.E.; McAdams, B.; Abrahante, J.E.; Simone, D.A.; Banik, R.K. Global transcriptome analysis of rat dorsal root ganglia to identify molecular pathways involved in incisional pain. Mol. Pain 2020, 16, 1744806920956480. [Google Scholar] [CrossRef]
- Ishida, H.; Zhang, Y.; Gomez, R.; Shannonhouse, J.; Son, H.; Banik, R.; Kim, Y.S. In Vivo Calcium Imaging Visualizes Incision-Induced Primary Afferent Sensitization and Its Amelioration by Capsaicin Pretreatment. J. Neurosci. 2021, 41, 8494–8507. [Google Scholar] [CrossRef] [PubMed]
- Shehab, S.; Atkinson, M. Vasoactive intestinal polypeptide (VIP) increases in the spinal cord after peripheral axotomy of the sciatic nerve originate from primary afferent neurons. Brain Res. 1986, 372, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Kim, K.H.; Nam, T.S.; Lee, W.T.; Park, K.A.; Kim, D.-W.; Leem, J.W. The role of uninjured C-afferents and injured afferents in the generation of mechanical hypersensitivity after partial peripheral nerve injury in the rat. Exp. Neurol. 2007, 204, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.J.; Xie, Y.-K. A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain 1988, 33, 87–107. [Google Scholar] [CrossRef]
- Kefauver, J.M.; Ward, A.B.; Patapoutian, A. Discoveries in structure and physiology of mechanically activated ion channels. Nature 2020, 587, 567–576. [Google Scholar] [CrossRef]
- Szallasi, A.; Blumberg, P.M. Vanilloid (Capsaicin) receptors and mechanisms. Pharmacol. Rev. 1999, 51, 159–212. [Google Scholar]
- Price, T.J.; Flores, C.M. Critical evaluation of the colocalization between calcitonin gene-related peptide, substance P, transient receptor potential vanilloid subfamily type 1 immunoreactivities, and isolectin B4 binding in primary afferent neurons of the rat and mouse. J. Pain 2007, 8, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Cavanaugh, D.J.; Chesler, A.T.; Bráz, J.M.; Shah, N.M.; Julius, D.; Basbaum, A.I. Restriction of transient receptor potential vanilloid-1 to the peptidergic subset of primary afferent neurons follows its developmental downregulation in nonpeptidergic neurons. J. Neurosci. 2011, 31, 10119–10127. [Google Scholar] [CrossRef]
- Derry, S.; Rice, A.S.; Cole, P.; Tan, T.; Moore, R.A. Topical capsaicin (high concentration) for chronic neuropathic pain in adults. Cochrane Database Syst. Rev. 2017, 1, CD007393. [Google Scholar]
- Brown, J.D.; Saeed, M.; Do, L.; Braz, J.; Basbaum, A.I.; Iadarola, M.J.; Wilson, D.M.; Dillon, W.P. CT-guided injection of a TRPV1 agonist around dorsal root ganglia decreases pain transmission in swine. Sci. Transl. Med. 2015, 7, ra145–ra305. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.L.; Zhang, F.X.; Dong, F.; Bao, L.; Zhang, X. Experimental evidence for alleviating nociceptive hypersensitivity by single application of capsaicin. Mol. Pain 2015, 11, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arora, V.; Li, T.; Kumari, S.; Wang, S.; Asgar, J.; Chung, M.K. Capsaicin-induced depolymerization of axonal microtubules mediates analgesia for trigeminal neuropathic pain. Pain 2022, 163, 1479–1488. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Bian, C.; Yang, J.; Arora, V.; Gao, Y.; Wei, F.; Chung, M.K. Ablation of TRPV1+ Afferent Terminals by Capsaicin Mediates Long-Lasting Analgesia for Trigeminal Neuropathic Pain. eNeuro 2020, 7, eneuro.0118-20.2020. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Experimental Procedures | |||
|---|---|---|---|---|
| Neuropathic Pain | Immunohistochemistry/Western Blotting/qRT-PCR | c-Fos Immunohistochemistry | ||
| Treatment | Prevention | |||
| Plantar RTX injection (0.002%, 100 µL) | Left L5 nerve injury to induce neuropathic pain, and after 7 days, RTX was injected in the plantar skin of the left hind paw. | RTX was injected in the plantar skin of the left hind paw, and after 7 days, the left L5 nerve was injured to induce neuropathic pain | RTX was injected in the plantar skin of the left hind paw, and rats were sacrificed after 14 days. | RTX was injected in the plantar skin of the left hind paw, and after 14 days, a second injection of the same dose and volume of RTX was injected. The rats were sacrificed 90 min after the second injection. |
| Plantar vehicle injection (10% Tween 80, 10% EtOH, 80% normal saline, 100 µL) | Left L5 nerve injury to induce neuropathic pain, and after 7 days, vehicle was injected in the plantar skin of the left hind paw. | The vehicle was injected in the skin of the left hind paw, and after 7 days, left L5 nerve was injured to induce neuropathic pain | The vehicle was injected in the plantar skin of the left hind paw, and rats were sacrificed after 14 days | Vehicle was injected in plantar skin of the left hind paw, and after 14 days, RTX was injected in the same left hind paw. The rats were sacrificed 90 min after the second injection. |
| Right control | The right hind paw of RTX-injected animals served as control. | Right DRG and spinal cord served as control. | ||
| Antibody | Immunogen | Host | Catalogue/Source | Dilution |
|---|---|---|---|---|
| TRPV1 | YTGSLKPEDAEVFKDSMVPGEK | Guinea Pig | GP14100/Neuromics, MN, USA | 1:5000 |
| IB4 | Purified Griffonia simplicifolia lectin I whole molecule | Goat | AS2104/Vector Laboratories, Peterborough, UK | 1:1000 |
| CGRP | Synthetic peptide corresponding to a portion of rat α-calcitonin gene-related peptide (CGRP) | Rabbit | T4032/Peninsula Laboratories, San Carlos, CA, USA | 1:10,000 |
| CGRP | Synthetic peptide corresponding to Rat CGRP conjugated to Keyhole Limpet Haemocyanin (KLH). | Sheep | ab22560/Abcam, Waltham, MA, USA | 1:2000 |
| VIP | Pure Porcine VIP | Rabbit | Gift from Prof J. Allen | 1:2000 |
| SP | Synthetic peptide | Rabbit | T-4107 (IHC7451)/Peninsula Laboratories, San Carlos, CA, USA | 1:5000 |
| PGP 9.5 | Purified whole human PGP9.5 | Rabbit | RA95101, Ultraclone, Isle of Wight, UK | 1:1000 |
| c-Fos | KLH-conjugated linear peptide corresponding to 14 amino acids from the N-terminal region of human c-Fos | Rabbit | ABE457/Merck Millipore, MA, USA | 1:5000 |
| DRGs (n = 3) | TRPV1 (%) | CGRP (%) | IB4 (%) |
|---|---|---|---|
| Left L4 (RTX) | 12.12 ± 0.81 *** | 25.23 ± 1.24 *** | 19.70 ± 1.16 *** |
| Left L4 (vehicle) | 36.45 ± 0.70 | 43.14 ± 0.81 | 40.58 ± 1.70 |
| Right L4 (control) | 31.34 ± 1.58 | 39.69 ± 1.88 | 42.58 ± 1.00 |
| DRGs | 40 H (n = 3) | 14 Days (n = 3) | ||
|---|---|---|---|---|
| TRPV1 | VIP | TRPV1 | VIP | |
| Left L4 (RTX) | 23.58% ± 1.61 ** | 14.38% ± 0.65 *** | 8.19% ± 0.77 *** | 12.86% ± 1.22 *** |
| Left L4 (Vehicle) | 34.62% ± 0.41 | 0.53% ± 0.06 | 31.74% ± 0.43 | 0.86% ± 0.04 |
| Right L4 (Control) | 31.19% ± 1.28 | 3.05% ± 0.72 | 28.78% ± 0.62 | 4.37% ± 0.16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Javed, H.; Johnson, A.M.; Challagandla, A.K.; Emerald, B.S.; Shehab, S. Cutaneous Injection of Resiniferatoxin Completely Alleviates and Prevents Nerve-Injury-Induced Neuropathic Pain. Cells 2022, 11, 4049. https://doi.org/10.3390/cells11244049
Javed H, Johnson AM, Challagandla AK, Emerald BS, Shehab S. Cutaneous Injection of Resiniferatoxin Completely Alleviates and Prevents Nerve-Injury-Induced Neuropathic Pain. Cells. 2022; 11(24):4049. https://doi.org/10.3390/cells11244049
Chicago/Turabian StyleJaved, Hayate, Aishwarya Mary Johnson, Anil Kumar Challagandla, Bright Starling Emerald, and Safa Shehab. 2022. "Cutaneous Injection of Resiniferatoxin Completely Alleviates and Prevents Nerve-Injury-Induced Neuropathic Pain" Cells 11, no. 24: 4049. https://doi.org/10.3390/cells11244049