
4-Methylumebelliferone Enhances Radiosensitizing Effects of Radioresistant Oral Squamous Cell Carcinoma Cells via Hyaluronan Synthase 3 Suppression

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
2.3. Irradiation Condition
2.4. SiRNA Transfection
2.5. HA Density Quantitation
2.6. Clonogenic Survival Assay
2.7. Monolayer Wound Healing Assay
2.8. RNA Extraction and Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
2.9. Flow Cytometric Analysis
2.10. SDS-PAGE and Western Blotting
2.11. Assay of SOD Activity
2.12. Statistical Analysis
3. Results
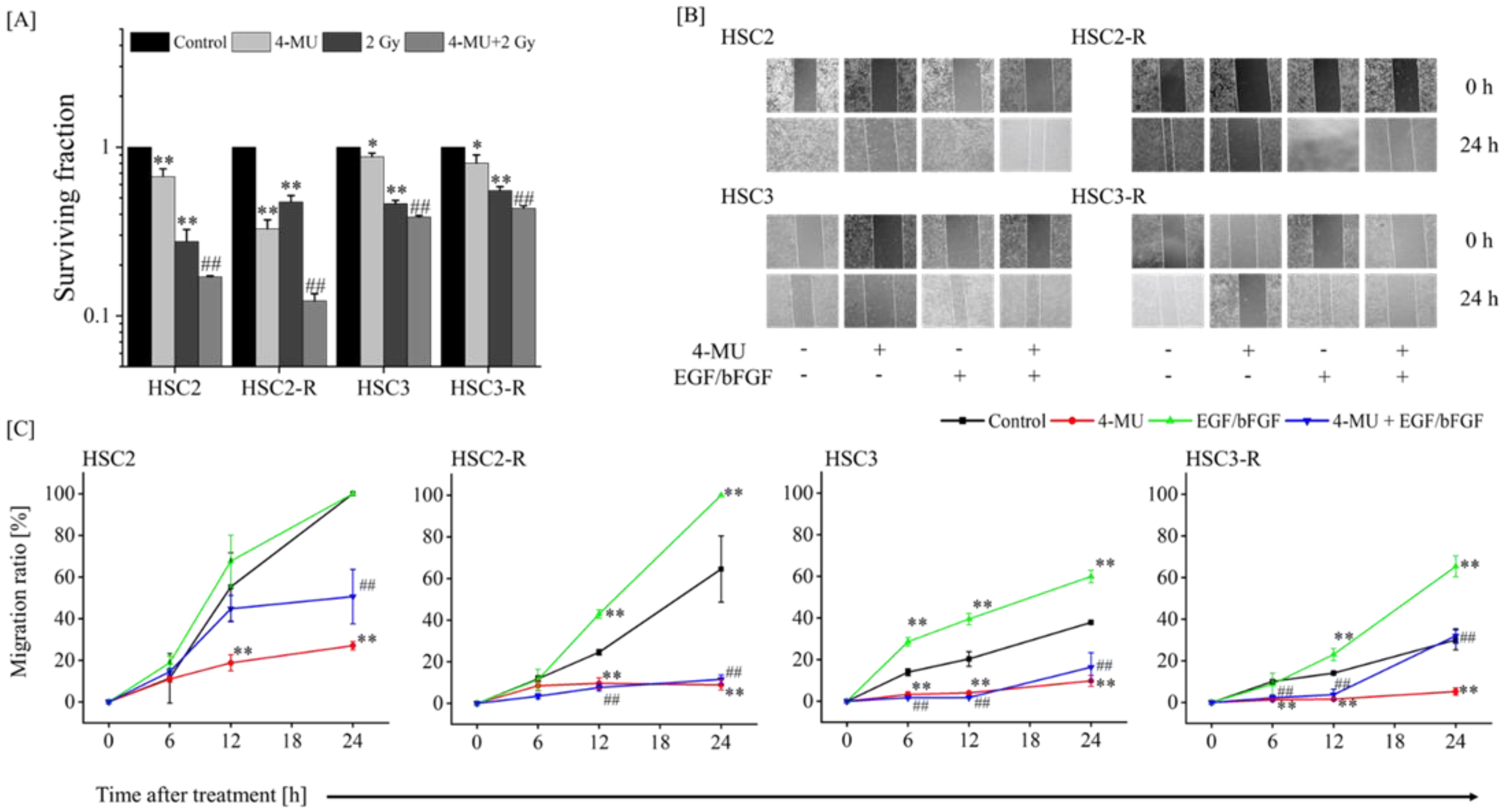
3.1. Evaluation of the Effect of 4-MU on Radioresistance
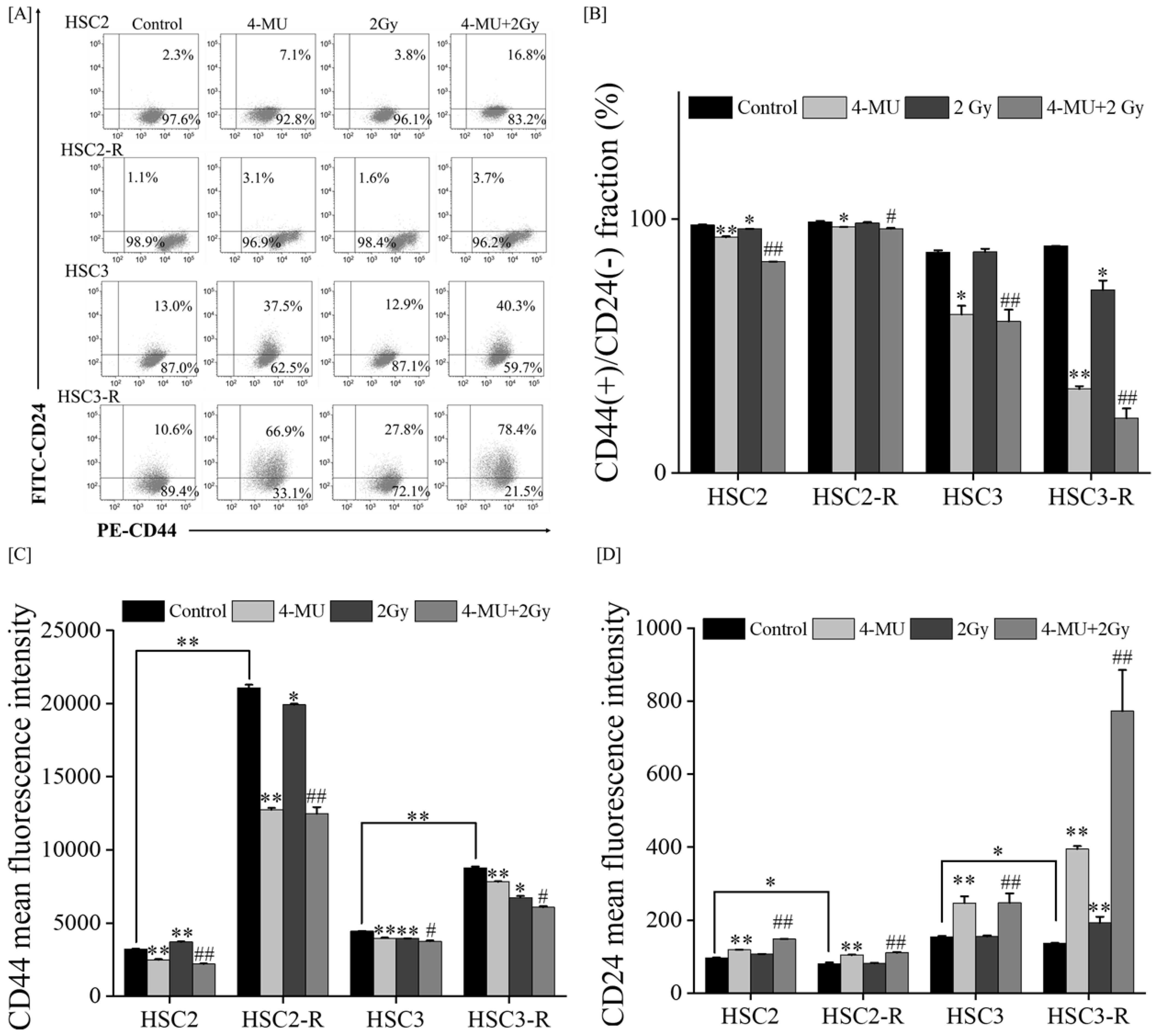
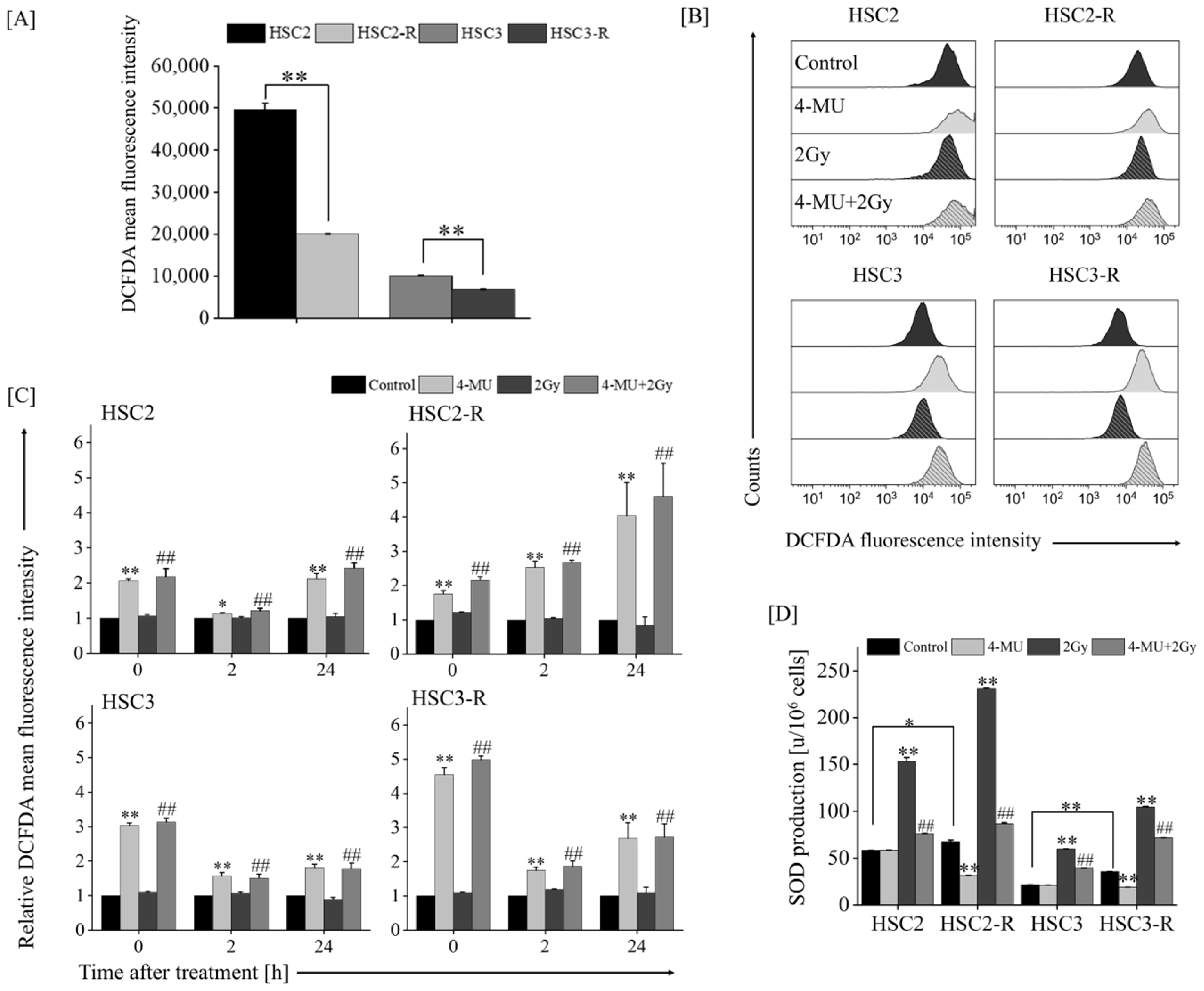
3.2. Investigation of Mechanisms of Radiosensitization with 4-MU
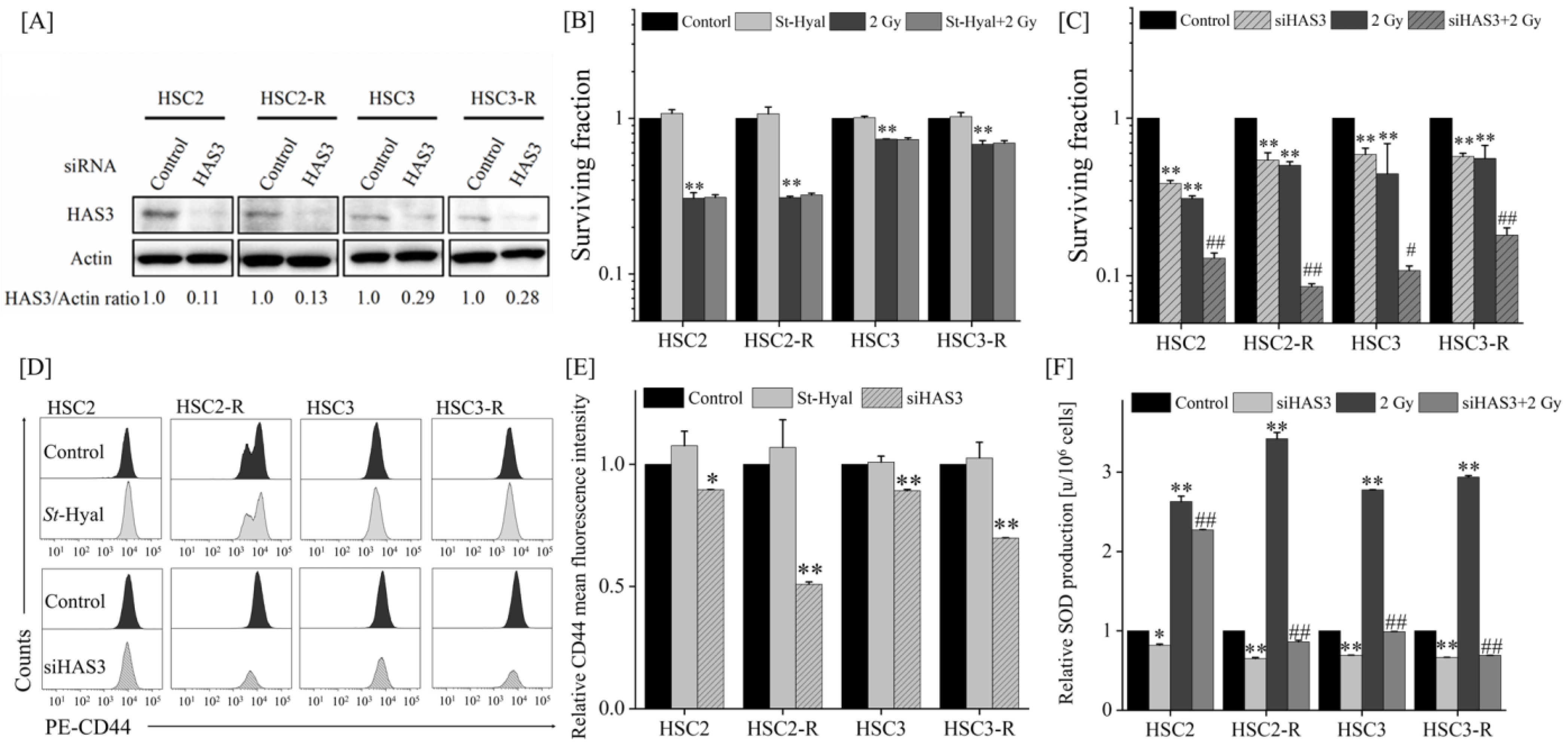
3.3. Radiosensitizing Effects after Extracellular HA Elimination or HAS Inhibition
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2017. CA Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Seigel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narita, Y.; Kato, T.; Ono, T.; Oyama, S.; Komori, S.; Arai, K.; Abe, Y.; Harada, T.; Nakamura, T.; Wada, H.; et al. Effect of anatomical change on dose distribution during radiotherapy for maxillary sinus carcinoma: Passive scattering proton therapy versus volumetric-modulated arc therapy. Br. J. Radiol. 2018, 92, 20180273. [Google Scholar] [CrossRef] [PubMed]
- Pfister, D.G.; Spencer, S.; Adelstein, D.; Adkins, D.; Anzai, Y.; Brizel, D.M.; Bruce, J.Y.; Busse, P.M.; Caudell, J.J.; Cmelak, A.J.; et al. Head and Neck Cancers, Version 2.2020, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Canc. Netw. 2020, 18, 873–898. [Google Scholar] [CrossRef] [PubMed]
- Ou, D.; Blanchard, P.; El Khoury, C.; De Felice, F.; Even, C.; Levy, A.; Nguyen, F.; Janot, F.; Gorphe, P.; Deutsch, E.; et al. Induction chemotherapy with docetaxel, cisplatin and fluorouracil followed by concurrent chemoradiotherapy or chemoradiotherapy alone in locally advanced non-endemic nasopharyngeal carcinoma. Oral Oncol. 2016, 62, 114–121. [Google Scholar] [CrossRef]
- Tangthongkum, M.; Kirtstreesakul, V.; Supanimitjaroenporn, P.; Leelasawatsuk, P. Treatment outcome of advance staged oral cavity cancer: Concurrent chemoradiotherapy compared with primary surgery. Eur. Arch. Otorhinolaryngol. 2017, 274, 2567–2572. [Google Scholar] [CrossRef] [PubMed]
- Bonner, J.A.; Harari, P.M.; Giralt, J.; Azarnia, N.; Shin, D.M.; Cohen, R.B.; Jones, C.U.; Sur, R.; Raben, D.; Jassem, J.; et al. Radiotherapy plus cetuximab for squamous-cell carcinoma of the head and neck. N. Engl. J. Med. 2006, 354, 567–678. [Google Scholar] [CrossRef] [Green Version]
- Shahinas, J.; Hysi, D. Methods and risk of bias in molecular marker prognosis studies in oral squamous cell carcinoma. Oral Dis. 2018, 24, 115–119. [Google Scholar] [CrossRef]
- McDermott, N.; Meunier, A.; Mooney, B.; Nortey, G.; Hernandez, C.; Hurley, S.; Lynam-Lennon, N.; Baesoom, S.H.; Bowman, K.J.; Marples, B.; et al. Fractionated radiation exposure amplifies the radioresistant nature of prostate cancer cells. Sci. Rep. 2016, 6, 34796. [Google Scholar] [CrossRef] [Green Version]
- Arechaga-Ocampo, E.; Lopez-Camarillo, C.; Villegas-Sepulveda, N.; Gonzalez-De la Rosa, C.H.; Perez-Añorve, I.X.; Roldan-Perez, R.; Flores-Perez, A.; Peña-Curiel, O.; Angeles-Zaragoza, O.; Rangel Corona, R.; et al. Tumor suppressor miR-29c regulates radioresistance in lung cancer cells. Tumor Biol. 2017, 39, 1010428317695010. [Google Scholar] [CrossRef]
- Hara, T.; Iwadate, M.; Tachibana, K.; Waguri, S.; Takenoshita, S.; Hamada, N. Metastasis of breast cancer cells to the bone, lung, and lymph nodes promotes resistance to ionizing radiation. Strahlenther. Onkol. 2017, 193, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Chang, J.Y.; Yun, S.H.; Yang, K.M.; Hong, E.H.; Kim, D.H.; Um, H.D.; Lee, K.H.; Lee, S.J.; Hwang, S.G. Chloride intracellular channel 1 identified using proteomic analysis plays an important role in the radiosensitivity of Hep-2 cells via reactive oxygen species production. Proteomics 2010, 10, 2589–2604. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Xu, K.; Chen, Q.; Wang, B.; Pan, J.; Huang, S.; Wei, Y.; Ma, H. Simvastatin inhibits the development of radioresistant esophageal cancer cells by increasing the radiosensitivity and reversing EMT process via the PTEN-PI3K/AKT pathway. Exp. Cell Res. 2018, 362, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, T.; Takagaki, K.; Shibata, S.; Tanaka, K.; Higuchi, T.; Endo, M. Hyaluronic-acid-deficient extracellular matrix induced by addition of 4-methylumbelliferone to the medium of cultured human skin fibroblasts. Biochem. Biophys. Res. Commun. 1995, 208, 470–475. [Google Scholar] [CrossRef]
- Kuroda, Y.; Kasai, K.; Nanashima, N.; Nozaka, H.; Nakano, M.; Chiba, M.; Yoneda, M.; Nakamura, T. 4-Methylumbelliferone inhibits the phosphorylation of hyaluronan synthase 2 induced by 12-O-tetradecanoyl-phorbol-13-acetate. Biomed. Res. 2013, 34, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Kultti, A.; Pasonen-Seppanen, S.; Jauhiainen, M.; Rilla, K.J.; Karna, R.; Pyoria, E.; Tammi, R.H.; Tammi, M.I. 4-methylumbelliferone inhibits hyaluronan synthesis by depletion of cellular UDP-glucuronic acid and downregulation of hyaluronan synthase 2 and 3. Exp. Cell Res. 2009, 315, 1914–1923. [Google Scholar] [CrossRef]
- Yates, T.J.; Lopez, L.E.; Lokeshwar, S.D.; Ortiz, N.; Kallifatidis, G.; Jordan, A.; Hoye, K.; Altman, N.; Lokeshwar, V.B. Dietary supplement 4-methylumbelliferone: An effective chemopreventive and therapeutic agent for prostate cancer. J. Natl. Cancer Inst. 2015, 107, djv085. [Google Scholar] [CrossRef] [Green Version]
- Piccioni, F.; Fiore, E.; Bayo, J.; Atorrasagasti, C.; Peixoto, E.; Rizzo, M.; Malvicini, M.; Tirado-González, I.; García, M.G.; Alaniz, L.; et al. 4-methylumbelliferone inhibits hepatocellular carcinoma growth by decreasing IL-6 production and angiogenesis. Glycobiology 2015, 25, 825–835. [Google Scholar] [CrossRef] [Green Version]
- Saga, R.; Hasegawa, K.; Murata, K.; Chiba, M.; Nakamura, T.; Okumura, K.; Tsuruga, E.; Hosokawa, Y. Regulation of radiosensitivity by 4-methylumbelliferone via the suppression of interleukin-1 in fibrosarcoma cells. Oncol. Lett. 2019, 14, 3555–3561. [Google Scholar] [CrossRef]
- Hasegawa, K.; Saga, R.; Takahashi, R.; Fukui, R.; Chiba, M.; Okumura, K.; Tsuruga, E.; Hosokawa, Y. 4-methylubelliferone inhibits clonogenic potency by suppressing high molecular weight-hyaluronan in fibrosarcoma cells. Oncol. Lett. 2020, 19, 2801–2808. [Google Scholar] [CrossRef]
- Saga, R.; Matsuya, Y.; Takahashi, R.; Hasegawa, K.; Date, H.; Hosokawa, Y. 4-Methylumbelliferone administration enhances radiosensitivity of human fibrosarcoma by intercellular communication. Sci. Rep. 2021, 11, 8258. [Google Scholar] [CrossRef]
- Guo, G.; Yan-Sanders, Y.; Lyn-Cook, B.D.; Wang, T.; Tamae, D.; Ogi, J.; Khaletskiy, A.; Li, Z.; Weydert, C.; Longmate, J.A.; et al. Manganese Superoxide Dismutase-Mediated Gene Expression in Radiation-Induced Adaptive Responses. Mol. Cell Biol. 2003, 23, 2362–2378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murley, J.S.; Kataoka, Y.; Cao, D.; Li, J.J.; Oberley, L.W.; Grdina, D.J. Delayed radioprotection by NFkappaB-mediated induction of Sod2 (MnSOD) in SA-NH tumor cells after exposure to clinically used thiol-containing drugs. Radiat. Res. 2004, 162, 536–546. [Google Scholar] [CrossRef]
- Piao, L.; Canguo, Z.; Wenjie, L.; Xiaoli, C.; Wenli, S.; Li, L. Lipopolysaccharides-stimulated macrophage products enhance Withaferin: A-induced apoptosis via activation of caspases and inhibition of NF-κB pathway in human cancer cells. Mol. Immunol. 2017, 81, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Aravindan, N.; Xu, J.; Natarajan, M. Inter- and intra-cellular mechanism of NF-kB-dependent survival advantage and clonal expansion of radio-resistant cancer cells. Cell Signal. 2017, 31, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, Y.; Li, L.; Baba, T.; Nakagawa, H.; Shimura, T.; Yamamoto, Y.; Ohkubo, Y.; Fukumoto, M. Clinically relevant radioresistant cells efficiently repair DNA double-strand breaks induced by X-rays. Cancer Sci. 2009, 100, 747–752. [Google Scholar] [CrossRef]
- Nagano, O.; Okazaki, S.; Saya, H. Redox regulation in stem-like cancer cells by CD44 variant isoforms. Oncogene 2013, 32, 5191–5198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alshareeda, A.T.; Negm, O.H.; Albarakati, N.; Green, A.R.; Nolan, C.; Sultana, R.; Madhusudan, S.; Benhasouna, A.; Tighe, P.; Ellis, I.O.; et al. Clinicopathological significance of KU70/KU80, a key DNA damage repair protein in breast cancer. Breast Cancer Res. Treat. 2013, 139, 301–310. [Google Scholar] [CrossRef]
- Groselj, B.; Kerr, M.; Kiltie, A.E. Radiosensitisation of bladder cancer cells by panobinostat is modulated by Ku80 expression. Radiother. Oncol. 2013, 108, 429–433. [Google Scholar] [CrossRef] [Green Version]
- Kuwahara, Y.; Mori, M.; Oikawa, T.; Shimura, T.; Ohtake, Y.; Mori, S.; Ohkubo, Y.; Fukumoto, M. The modified high-density survival assay is the useful tool to predict the effectiveness of fractionated radiation exposure. J. Radiat. Res. 2010, 51, 297–302. [Google Scholar] [CrossRef]
- Saga, R.; Monzen, S.; Chiba, M.; Yoshino, H.; Nakamura, T.; Hosokawa, Y. Anti-tumor and anti-invasion effects of a combination of 4-methylumbelliferone and ionizing radiation in human fibrosarcoma cells. Oncol. Lett. 2017, 13, 410–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.Y.; Lee, C.H.; Dedaj, R.; Zhao, H.; Mrabat, H.; Sheidlin, A.; Syrkina, O.; Huang, P.M.; Garg, H.G.; Hales, C.A.; et al. High-molecular-weight hyaluronan—A possible new treatment for sepsis-induced lung injury: A preclinical study in mechanically ventilated rats. Crit. Care 2008, 12, R102. [Google Scholar] [CrossRef] [Green Version]
- Todoroki, K.; Ogasawara, S.; Akiba, J.; Nakayama, M.; Naito, Y.; Seki, N.; Kusukawa, J.; Yano, H. CD44v3+/CD24− cells possess cancer stem cell-like properties in human oral squamous cell carcinoma. Int. J. Oncol. 2016, 48, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghuwalewala, S.; Ghatak, D.; Das, P.; Dey, S.; Sarkar, S.; Alam, N.; Panda, C.K.; Roychoudhury, S. CD44(high)CD24(low) molecular signature determines the Cancer Stem Cell and EMT phenotype in Oral Squamous Cell Carcinoma. Stem Cell Res. 2016, 16, 405–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bensimon, J.; Biard, D.; Paget, V.; Goislard, M.; Morel-Altmeyer, S.; Konge, J.; Chevillard, S.; Lebeau, J. Forced extinction of CD24 stem-like breast cancer marker alone promotes radiation resistance through the control of oxidative stress. Mol. Carcinog. 2016, 55, 245–254. [Google Scholar] [CrossRef]
- Schieber, M.S.; Chandel, N.S. ROS links glucose metabolism to breast cancer stem cell and EMT phenotype. Cancer Cell 2013, 23, 265–267. [Google Scholar] [CrossRef] [Green Version]
- Ishimoto, T.; Nagano, O.; Yae, T.; Tamada, M.; Motohara, M.; Oshima, H.; Ikeda, T.; Asaba, R.; Yagi, H.; Masuko, T.; et al. CD44 variant regulates redox status in cancer cells by stabilizing the xCT subunit of system xc(-) and thereby promotes tumor growth. Cancer Cell. 2011, 19, 387–400. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Ishikawa, T.; Nanashima, N.; Miura, T.; Nozaka, H.; Nakaoka, R.; Sato, T. 4-Methylumbelliferone induces the expression of membrane type 1-matrix metalloproteinase in cultured human skin fibroblasts. Biochem. Biophys. Res. Commun. 2002, 298, 646–650. [Google Scholar] [CrossRef]
- Yamada, K. The effect of digestion with Streptomyces hyaluronidase upon certain histochemical reactions of hyaluronic acid-containing tissues. J. Histochem. Cytochem. 1973, 21, 794–803. [Google Scholar] [CrossRef] [Green Version]
- Lokeshwar, V.B.; Lopez, L.E.; Munoz, D.; Chi, A.; Shirodkar, S.P.; Lokeshwar, S.D.; Escudero, D.O.; Dhir, N.; Altman, N. Antitumor activity of hyaluronic acid synthesis inhibitor 4-methylumbelliferone in prostate cancer cells. Cancer Res. 2010, 70, 2613–2623. [Google Scholar] [CrossRef]
- Arai, E.; Nishida, Y.; Wasa, J.; Urakawa, H.; Zhuo, L.; Kimata, K.; Kozawa, E.; Futamura, N.; Ishiguro, N. Inhibition of hyaluronan retention by 4-methylumbelliferone suppresses osteosarcoma cells in vitro and lung metastasis in vivo. Br. J. Cancer 2011, 105, 1839–1849. [Google Scholar] [CrossRef] [Green Version]
- Urakawa, H.; Nishida, Y.; Wasa, J.; Arai, E.; Zhuo, L.; Kimata, K.; Kozawa, E.; Futamura, N.; Ishiguro, N. Inhibition of hyaluronan synthesis in breast cancer cells by 4-methylumbelliferone suppresses tumorigenicity in vitro and metastatic lesions of bone in vivo. Int. J. Cancer 2012, 130, 454–466. [Google Scholar] [CrossRef]
- Thanee, M.; Loilome, W.; Techasen, A.; Sugihara, E.; Okazaki, S.; Abe, S.; Ueda, S.; Masuko, T.; Namwat, N.; Khuntikeo, N.; et al. CD44 variant-dependent redox status regulation in liver fluke-associated cholangiocarcinoma: A target for cholangiocarcinoma treatment. Cancer Sci. 2016, 107, 991–1000. [Google Scholar] [CrossRef] [PubMed]
- Bannai, S.; Ishii, T. Transport of cystine and cysteine and cell growth in cultured human diploid fibroblasts, effect of glutamate and homocysteate. J. Cell Physiol. 1982, 112, 265–272. [Google Scholar] [CrossRef]
- Hauser-Kawaguchi, A.; Luyt, L.G.; Turley, E. Design of peptide mimetics to block pro-inflammatory functions of HA fragments. Matrix Biol. 2019, 78–79, 346–356. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Kiningham, K.K.; Devalaraja, M.N.; Yeh, C.C.; Majima, H.; Kasarskis, E.J.; St Clair, D.K. An intronic NF-kappaB element is essential for induction of the human manganese superoxide dismutase gene by tumor necrosis factor-alpha and interleukin-1beta. DNA Cell Biol. 1999, 18, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Thorne, R.F.; Legg, J.W.; Isacke, C.M. The role of the CD44 transmembrane and cytoplasmic domains in co-ordinating adhesive and signalling events. J. Cell Sci. 2004, 117, 373–380. [Google Scholar] [CrossRef] [Green Version]
- Abdraboh, M.E.; Gaur, R.L.; Hollenbach, A.D.; Sandquist, D.; Raj, M.H.; Ouhtit, A. Survivin is a novel target of CD44-promoted breast tumor invasion. Am. J. Pathol. 2011, 179, 555–563. [Google Scholar] [CrossRef]
- Brinck, J.; Heldin, P. Expression of recombinant hyaluronan synthase (HAS) isoforms in CHO cells reduces cell migration and cell surface CD44. Exp. Cell Res. 1999, 252, 342–351. [Google Scholar] [CrossRef]
- Evanko, S.P.; Wight, T.N. Intracellular localization of hyaluronan in proliferating cells. J. Histochem. Cytochem. 1999, 47, 1331–1342. [Google Scholar] [CrossRef]
- Evanko, S.P.; Parks, W.T.; Weight, T.N. Intracellular hyaluronan in arterial smooth muscle cells, association with microtubules, RHAMM, and the mitotic spindle. J. Histochem. Cytochem. 2004, 52, 1525–1535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, J.; Hascall, V.C.; Wang, A. Cyclin, D3 mediates synthesis of a hyaluronan matrix that is adhesive for monocytes in mesangial cells stimulated to divide in hyperglycemic medium. J. Biol. Chem. 2009, 284, 16621–16632. [Google Scholar] [CrossRef] [Green Version]
- Bohaumilitzky, L.; Huber, A.K.; Stork, E.M.; Wengert, S.; Woelfl, F.; Boehm, H. A Trickster in Disguise: Hyaluronan’s Ambivalent Roles in the Matrix. Front. Oncol. 2017, 7, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misra, S.; Hascall, V.C.; Markwald, R.R.; Ghatak, S. Interactions between Hyaluronan and Its Receptors (CD44, RHAMM) Regulate the Activities of Inflammation and Cancer. Front. Immunol. 2015, 6, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.L.; Wang, M.J.; Chen, J.Y. Acetylation and activation of STAT3 mediated by nuclear translocation of CD44. J. Cell Biol. 2009, 185, 949–957. [Google Scholar] [CrossRef] [Green Version]
- Skandalis, S.S.; Karalis, T.; Heldin, P. Intracellular hyaluronan: Importance for cellular functions. Semin. Cancer Biol. 2020, 60, 20–30. [Google Scholar] [CrossRef]
- Assmann, V.; Marshall, F.J.; Fieber, C.; Hofmann, M.; Hart, I.R. The human hyaluronan receptor RHAMM is expressed as an intracellular protein in breast cancer cells. J. Cell Sci. 1998, 111, 1685–1694. [Google Scholar] [CrossRef]
- Pilarski, L.M.; Masellis-Smith, A.; Belch, A.R.; Yang, B.; Savani, R.C.; Turley, E.A. RHAMM, a receptor for hyaluronan-mediated motility, on normal human lymphocytes, thymocytes and malignant B cells: A mediator in B cell malignancy? Leuk. Lymphoma. 1994, 14, 363–374. [Google Scholar] [CrossRef]
- Yang, B.; Yang, B.L.; Savani, R.C.; Turley, E.A. Identification of a common hyaluronan binding motif in the hyaluronan binding proteins RHAMM, CD44 and link protein. EMBO J. 1994, 13, 286–296. [Google Scholar] [CrossRef]
- Nguyen, N.; Kumar, A.; Chacko, S.; Ouellette, R.J.; Ghosh, A. Human hyaluronic acid synthase-1 promotes malignant transformation via epithelial-to-mesenchymal transition, micronucleation and centrosome abnormalities. Cell Commun. Signal. 2017, 15, 48. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.Z.; Fang, W.Y.; Huang, C.C.; Tsai, S.T.; Wang, Y.C.; Yang, C.L.; Wu, L.W. Hyaluronan synthase 3 mediated oncogenic action through forming inter-regulation loop with tumor necrosis factor alpha in oral cancer. Oncotarget 2017, 8, 15563–15583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, R.; Thayalan, D.K.; Padmanaban, R.; Ramadas, R.; Annasamy, R.K.; Anandan, N. Association of serum and salivary tumor necrosis factor-alpha with histological grading in oral cancer and its role in differentiating premalignant and malignant oral disease. Asian Pac. J. Cancer Prev. 2014, 15, 7141–7148. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasegawa, K.; Saga, R.; Ohuchi, K.; Kuwahara, Y.; Tomita, K.; Okumura, K.; Sato, T.; Fukumoto, M.; Tsuruga, E.; Hosokawa, Y. 4-Methylumebelliferone Enhances Radiosensitizing Effects of Radioresistant Oral Squamous Cell Carcinoma Cells via Hyaluronan Synthase 3 Suppression. Cells 2022, 11, 3780. https://doi.org/10.3390/cells11233780
Hasegawa K, Saga R, Ohuchi K, Kuwahara Y, Tomita K, Okumura K, Sato T, Fukumoto M, Tsuruga E, Hosokawa Y. 4-Methylumebelliferone Enhances Radiosensitizing Effects of Radioresistant Oral Squamous Cell Carcinoma Cells via Hyaluronan Synthase 3 Suppression. Cells. 2022; 11(23):3780. https://doi.org/10.3390/cells11233780
Chicago/Turabian StyleHasegawa, Kazuki, Ryo Saga, Kentaro Ohuchi, Yoshikazu Kuwahara, Kazuo Tomita, Kazuhiko Okumura, Tomoaki Sato, Manabu Fukumoto, Eichi Tsuruga, and Yoichiro Hosokawa. 2022. "4-Methylumebelliferone Enhances Radiosensitizing Effects of Radioresistant Oral Squamous Cell Carcinoma Cells via Hyaluronan Synthase 3 Suppression" Cells 11, no. 23: 3780. https://doi.org/10.3390/cells11233780