
Metabolic and Transcriptional Stress Memory in Sorbus pohuashanensis Suspension Cells Induced by Yeast Extract

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Treatment, and Experimental Design
2.2. Determination of Cellulose, Oxidative Stress Parameters, and Antioxidant Enzyme Levels
2.3. Metabolite Extraction, UPLC-QTOF/MS Analysis, and Data Processing
2.4. RNA Extraction, Library Preparation, and Sequencing
2.5. Statistical Analysis
3. Results
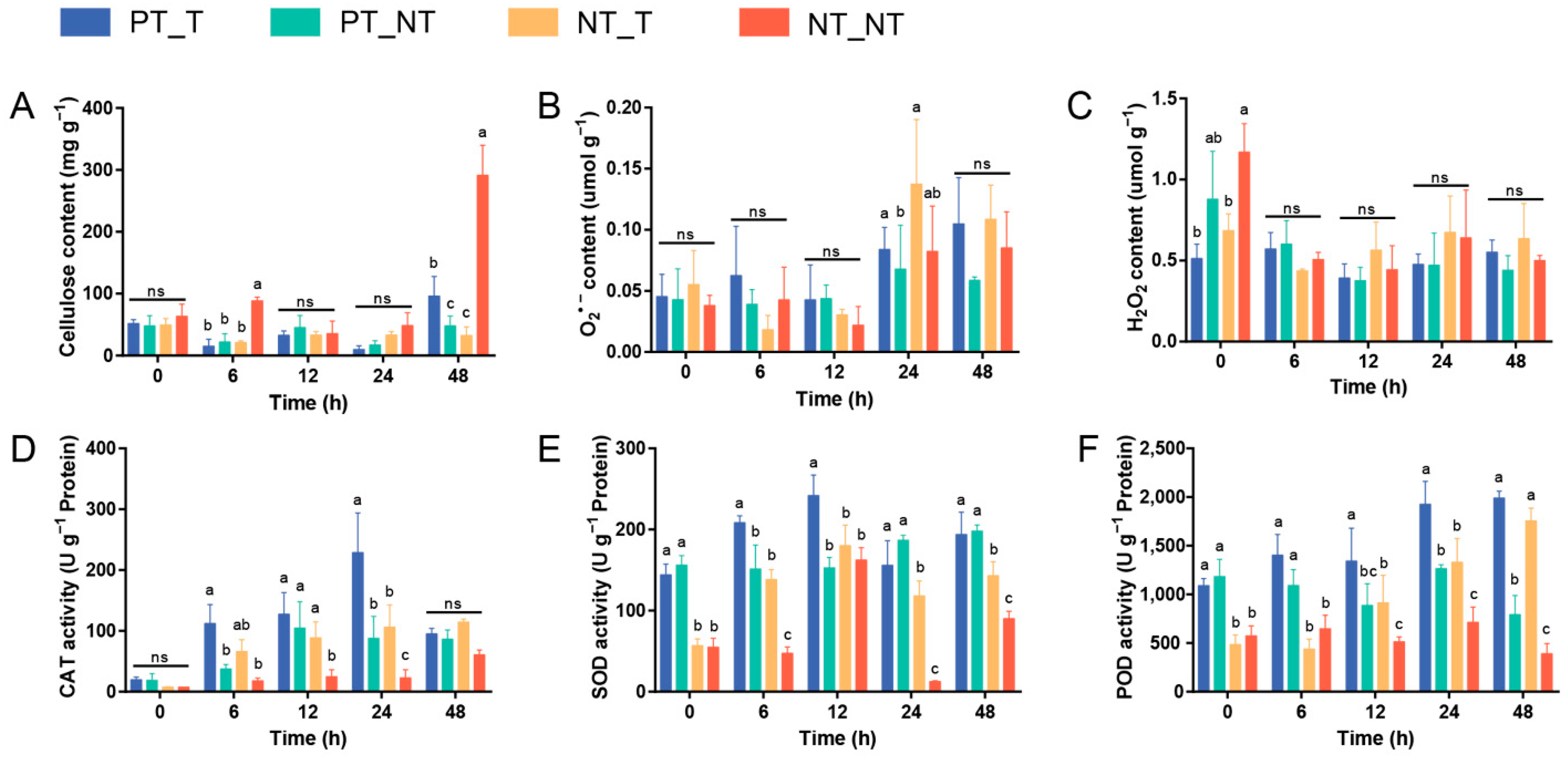
3.1. Cell Wall Integrity of SPSCs Induced by Recurrent YE Stress
3.2. Antioxidation Capacity of SPSCs Induced by Recurrent YE Stress
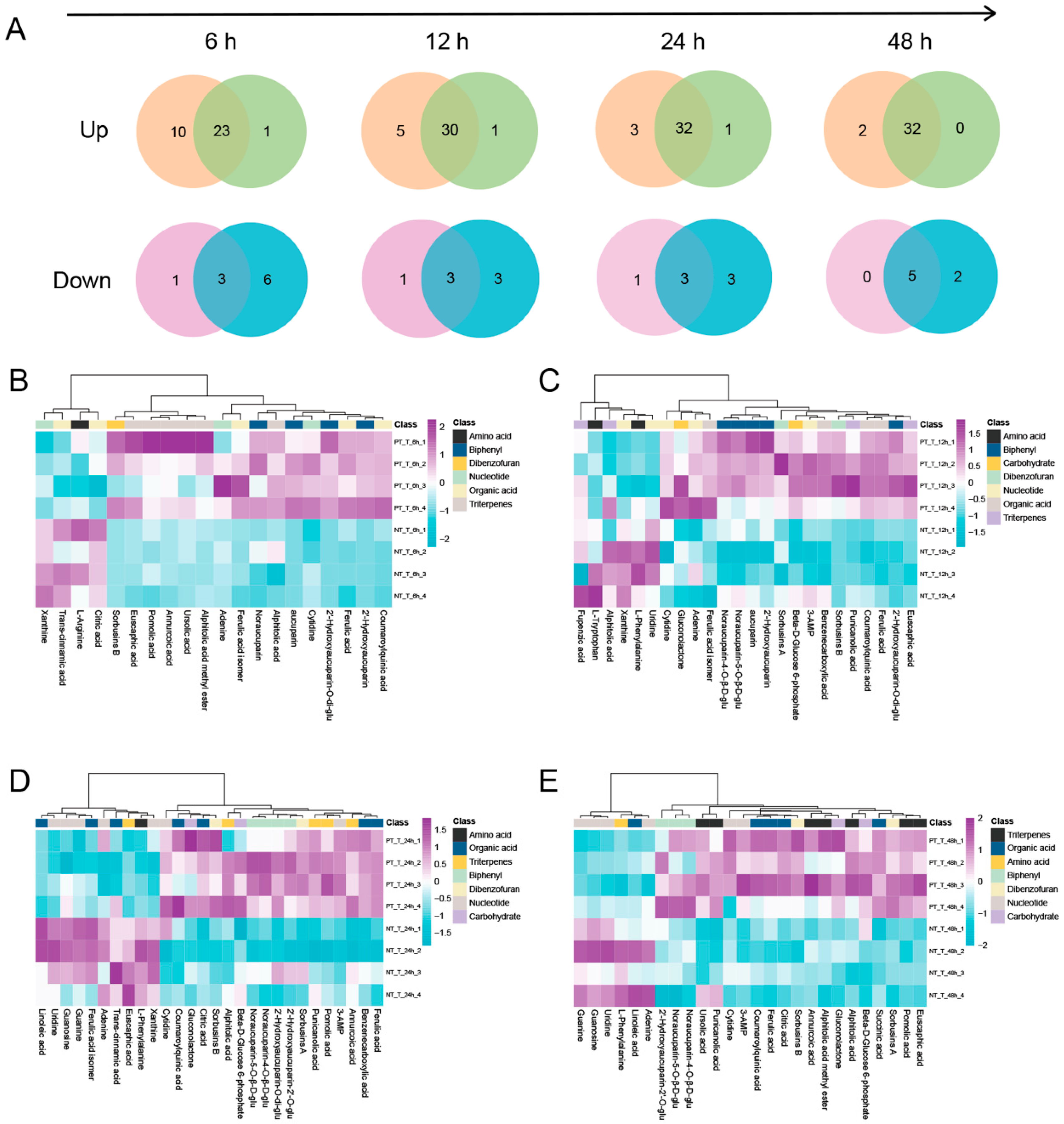
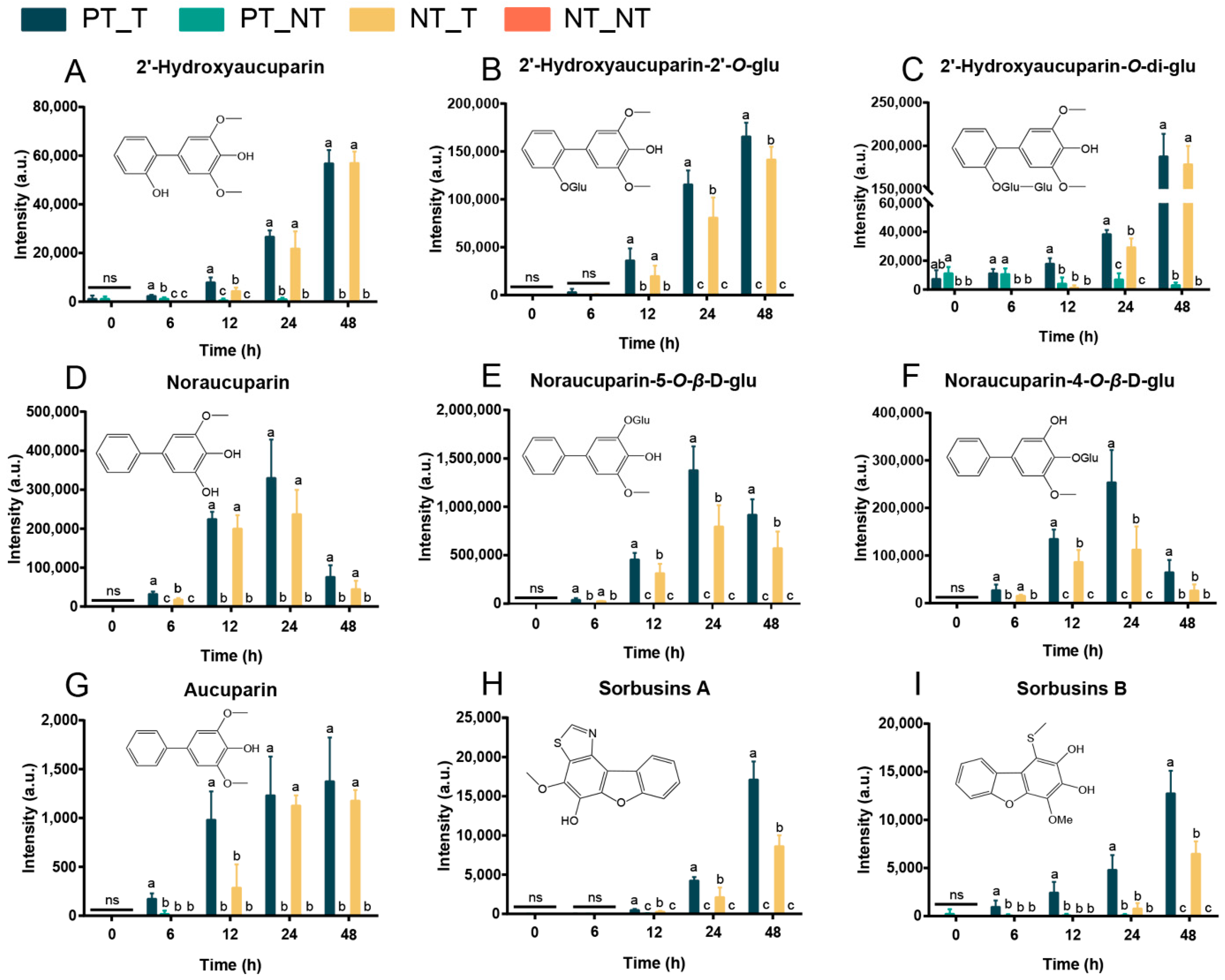
3.3. Metabolomic Memory Response in SPSCs Induced by YE
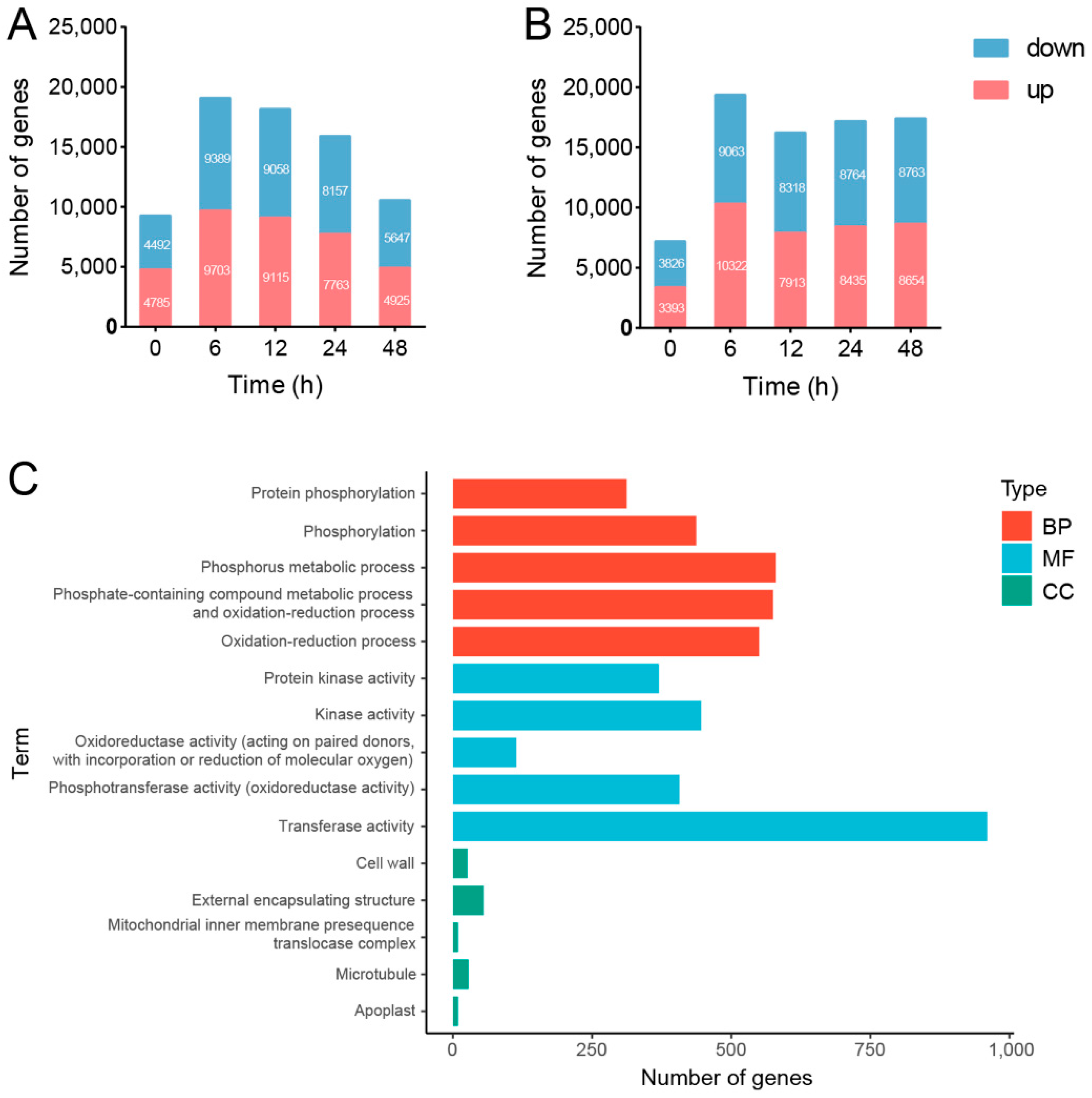
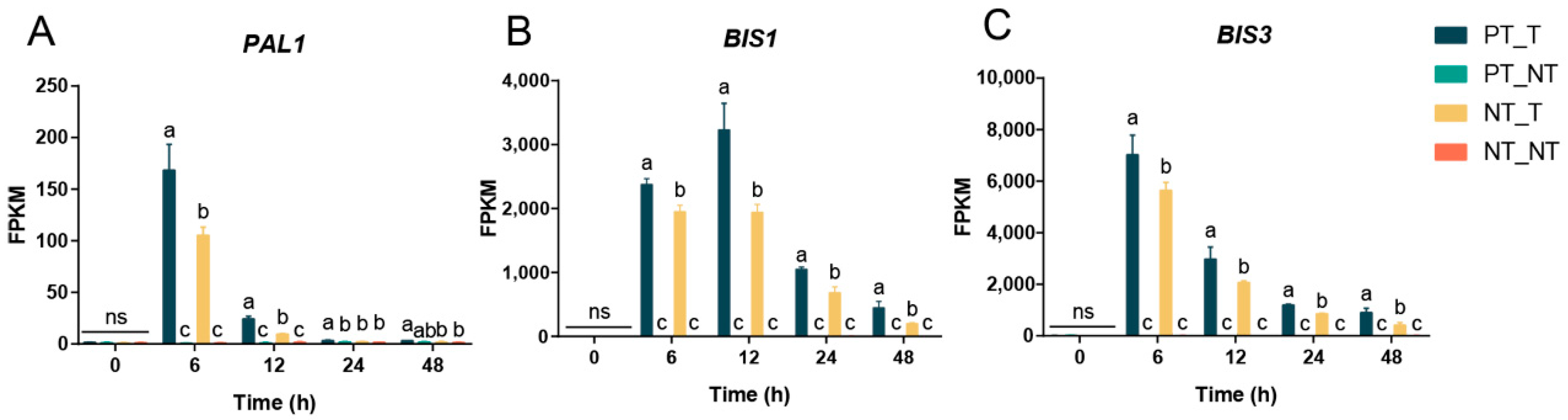
3.4. Transcriptional Memory Response in SPSCs Induced by YE
3.5. Correlation Analysis between Transcripts and Phytoalexins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lämke, J.; Brzezinka, K.; Altmann, S.; Bäurle, I. A hit-and-run heat shock factor governs sustained histone methylation and transcriptional stress memory. EMBO J. 2016, 35, 162–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, T.; Jin, Y.; Li, H.; Amombo, E.; Fu, J. Stress memory induced transcriptional and metabolic changes of perennial ryegrass (Lolium perenne) in response to salt stress. Physiol. Plant 2016, 156, 54–69. [Google Scholar] [CrossRef] [PubMed]
- Anjar, W.; Claude, B.; Gianpiero, M.; Julius, D.; Jonathan, P.; Jorg, H.; Ranjith, P.; Hadi, P.; Jorge, K.; Jorg, B.; et al. Hyperosmotic stress memory in Arabidopsis is mediated by distinct epigenetically labile sites in the genome and is restricted in the male germline by DNA glycosylase activity. eLife 2016, 5, e13546. [Google Scholar]
- Hu, T.; Liu, S.Q.; Amombo, E.; Fu, J.M. Stress memory induced rearrangements of HSP transcription, photosystem II photochemistry and metabolism of tall fescue (Festuca arundinacea Schreb.) in response to high-temperature stress. Front. Plant Sci. 2015, 6, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulak, M. Recurrent drought stress effects on essential oil profile of Lamiaceae plants: An approach regarding stress memory. Ind. Crops Prod. 2020, 154, 112695. [Google Scholar] [CrossRef]
- Liu, H.; Able, A.J.; Able, J.A. Priming crops for the future: Rewiring stress memory. Trends Plant Sci. 2022, 27, 699–716. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [Green Version]
- Jänkänpää, H.J.; Mishra, Y.; Schröder, W.P.; Jansson, S. Metabolic profiling reveals metabolic shifts in Arabidopsis plants grown under different light conditions. Plant Cell Environ. 2012, 35, 1824–1836. [Google Scholar] [CrossRef]
- Zuther, E.; Schaarschmidt, S.; Fischer, A.; Erban, A.; Pagter, M.; Mubeen, U.; Giavalisco, P.; Kopka, J.; Sprenger, H.; Hincha, D.K. Molecular signatures associated with increased freezing tolerance due to low temperature memory in Arabidopsis. Plant Cell Environ. 2019, 42, 854–873. [Google Scholar]
- Li, P.; Yang, H.; Wang, L.; Liu, H.; Huo, H.; Zhang, C.; Liu, A.; Zhu, A.; Hu, J.; Lin, Y.; et al. Physiological and transcriptome analyses reveal short-term responses and formation of memory under drought stress in rice. Front. Genet. 2019, 10, 55. [Google Scholar] [CrossRef] [Green Version]
- de Leonardis, A.M.; Fragasso, M.; Beleggia, R.; Ficco, D.B.; de Vita, P.; Mastrangelo, A.M. Effects of heat stress on metabolite accumulation and composition, and nutritional properties of durum wheat grain. Int. J. Mol. Sci. 2015, 16, 30382–30404. [Google Scholar] [CrossRef] [Green Version]
- Zhao, D.; Zhang, Y.; Lu, Y.; Fan, L.; Zhang, Z.; Zheng, J.; Chai, M. Genome sequence and transcriptome of Sorbus pohuashanensis provide insights into population evolution and leaf sunburn response. J. Genet. Genom. 2022, 49, 547–558. [Google Scholar] [CrossRef]
- Xu, M.; Yu, X.; Zheng, Y.; Zhang, T.; Xia, X.; Fu, Q.; Zhang, C. Study on nutritive substances and medicinal components of Sorbus pohuashanensis. For. Res. 2020, 33, 154–160. [Google Scholar]
- Gao, Y.; Yang, J.; Yang, X.; Zhang, L.; Wang, J.; Li, Q.; Lin, D.-M.; Zhang, M.; Xia, S.; Xu, L.; et al. Novel dibenzofuran and biphenyl phytoalexins from Sorbus pohuashanensis suspension cell and their antimicrobial activities. Fitoterapia 2021, 152, 104914. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Li, J.; Yuan, J.; Wang, S.; Zhou, L.; Zhou, L.; Kang, C.; Guo, L. High-throughput screening of secondary metabolites by Sorbus pohuashanensis cells under environmental stress using UHPLC-QTOF combined with AntDAS. Physiol. Plant 2021, 173, 2216–2225. [Google Scholar] [CrossRef]
- Song, C.; Wang, X.; Yang, J.; Kuang, Y.; Wang, Y.; Yang, S.; Qin, J.; Guo, L. Antifungal biphenyl derivatives from Sorbus pohuashanensis leaves infected by Alternaria tenuissi and their effect against crop pathogens. Chem. Biodivers. 2021, 18, e2100079. [Google Scholar] [CrossRef]
- Zhou, L.; Yang, J.; Yang, G.; Kang, C.; Xiao, W.; Lv, C.; Wang, S.; Tang, J.; Guo, L. Biphenyl phytoalexin in Sorbus pohuashanensis suspension cell induced by yeast extract. Molecules 2016, 21, 1180. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Wang, S.; Liu, Y.; Li, J.; Zhou, L.; Li, T.; Zhou, L.; Zhang, W.; Guo, L.; Huang, L. A preliminary study on memory function of Sorbus aucuparia suspension cell to biotic stress. Chin. J. Chin. Mat. Med. 2021, 46, 2467–2473. [Google Scholar]
- Updegraff, D.M. Semimicro determination of cellulose in biological materials. Anal. Biochem. 1969, 32, 420–424. [Google Scholar] [CrossRef]
- Bors, W.; Lengfelder, E.; Saran, M. Oxidation of hydroxylamine to nitrite as an assay for the combined presence of superoxide anions and hydroxyl radicals. Biochem. Biophy. Res. Commun. 1977, 75, 973–979. [Google Scholar] [CrossRef]
- Satterfield, C.N.; Bonnell, A.H. Interferences in titanium sulfate method for hydrogen peroxide. Anal. Chem. 1955, 27, 1174–1175. [Google Scholar] [CrossRef]
- Johansson, L.H.; Håkan, B.L.A. A spectrophotometric method for determination of catalase activity in small tissue samples. Anal. Biochem. 1988, 174, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, C.; Fridovich, I. Superoxide dismutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef] [PubMed]
- Doerge, D.R.; Divi, R.L.; Churchwell, M.I. Identification of the colored guaiacol oxidation product produced by peroxidases. Anal. Biochem. 1997, 250, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Khasin, M.; Bernhardson, L.F.; O’Neill, P.M.; Palmer, N.A.; Scully, E.D.; Sattler, S.E.; Funnell-Harris, D.L. Pathogen and drought stress affect cell wall and phytohormone signaling to shape host responses in a sorghum COMT bmr12 mutant. BMC Plant Biol. 2021, 21, 391. [Google Scholar] [CrossRef]
- Malinovsky, F.G.; Fangel, J.U.; Willats, W.G. The role of the cell wall in plant immunity. Front. Plant Sci. 2014, 5, 178. [Google Scholar] [CrossRef] [Green Version]
- Yong, S.Y.; Wickneswari, R. Molecular characterization of a cellulose synthase gene (AaxmCesA1) isolated from an Acacia auriculiformis x Acacia mangium hybrid. Plant Mol. Biol. Rep. 2013, 31, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Zeng, D.; Cui, J.; Yin, Y.; Xiong, Y.; Yu, W.; Zhao, H.; Guan, S.; Cheng, D.; Sun, Y.; Lu, W. The memory of rice response to spaceflight stress: From the perspective of metabolomics and proteomics. Int. J. Mol. Sci. 2022, 23, 3390. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, H.; Zhao, Z.; An, L. Nitric oxide synthase like activity-dependent nitric oxide production protects against chilling-induced oxidative damage in Chorispora bungeana suspension cultured cells. Plant Physiol. Biochem. 2010, 48, 936–944. [Google Scholar] [CrossRef]
- Sukyoon, K.; Youngpyo, L.; Soon, L.; Haengsoon, L.; Sangsoo, K. Transgenic plants with enhanced tolerance to environmental stress by metabolic engineering of antioxidative mechanism in chloroplasts. J. Plant Biotechnol. 2005, 32, 151–159. [Google Scholar]
- Sako, K.; Nguyen, H.M.; Seki, M. Advances in chemical priming to enhance abiotic stress tolerance in plants. Plant Cell Physiol. 2021, 61, 1995–2003. [Google Scholar] [CrossRef]
- Chinnusamy, V.; Zhu, J.K. Epigenetic regulation of stress responses in plants. Curr. Opin. Plant Biol. 2009, 12, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Alves de Freitas Guedes, F.; Menezes-Silva, P.E.; DaMatta, F.M.; Alves-Ferreira, M. Using transcriptomics to assess plant stress memory. Theor. Exp. Plant Phys. 2019, 31, 47–58. [Google Scholar] [CrossRef]
- Duan, L.; Liu, H.; Li, X.; Xiao, J.; Wang, S. Multiple phytohormones and phytoalexins are involved in disease resistance to Magnaporthe oryzae invaded from roots in rice. Physiol. Plant 2014, 152, 486–500. [Google Scholar] [CrossRef]
- Liu, B.; Beuerle, T.; Klundt, T.; Beerhues, L. Biphenyl synthase from yeast-extract-treated cell cultures of Sorbus aucuparia. Planta 2004, 218, 492–496. [Google Scholar] [CrossRef]
- Beerhues, L.; Liu, B. Biosynthesis of biphenyls and benzophenones-Evolution of benzoic acid-specific type III polyketide synthases in plants. Phytochemistry 2009, 70, 1719–1727. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Mohamed, H.I.; Aldaej, M.I.; Al-Khayri, J.M.; Rezk, A.A.; Al-Mssallem, M.Q.; Sattar, M.N.; Ramadan, K.M.A. Production and antioxidant activity of secondary metabolites in Hassawi rice (Oryza sativa L.) cell suspension under salicylic acid, yeast extract, and pectin elicitation. Vitro Cell. Dev.-Plant 2022, 58, 615–629. [Google Scholar] [CrossRef]
- Rani, D.; Meelaph, T.; De-Eknamkul, W.; Vimolmangkang, S. Yeast extract elicited isoflavonoid accumulation and biosynthetic gene expression in Pueraria candollei var. mirifica cell cultures. Plant Cell Tiss. Org. 2020, 141, 661–667. [Google Scholar] [CrossRef]
- Huttner, C.; Beuerle, T.; Scharnhop, H.; Ernst, L.; Beerhues, L. Differential effect of elicitors on biphenyl and dibenzofuran formation in Sorbus aucuparia cell cultures. J. Agric. Food Chem. 2010, 58, 11977–11984. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Kwok, K.W.; Wu, J. Enhancement of tanshinone production in Salvia miltiorrhiza Bunge (red or Chinese sage) hairy-root culture by hyperosmotic stress and yeast elicitor. Biotechnol. Appl. Biochem. 2007, 46, 191–196. [Google Scholar] [PubMed]
- do Amaral, M.N.; Auler, P.A.; Rossatto, T.; Barros, P.M.; Oliveira, M.M.; Braga, E.J.B. Long-term somatic memory of salinity unveiled from physiological, biochemical and epigenetic responses in two contrasting rice genotypes. Physiol. Plant 2020, 170, 248–268. [Google Scholar] [CrossRef] [PubMed]
- Szechynska-Hebda, M.; Kruk, J.; Górecka, M.; Karpinska, B.; Karpinski, S. Evidence for Light Wavelength-Specific Photoelectrophysiological Signaling and Memory of Excess Light Episodes in Arabidopsis. Plant Cell 2010, 22, 2201–2218. [Google Scholar] [CrossRef] [Green Version]
- Górecka, M.; Lewandowska, M.; Dąbrowska-Bronk, J.; Białasek, M.; Barczak-Brzyżek, A.; Kulasek, M.; Mielecki, J.; Kozłowska-Makulska, A.; Gawroński, P.; Karpiński, S. Photosystem II 22kDa protein level—A prerequisite for excess light-inducible memory, cross-tolerance to UV-C and regulation of electrical signalling. Plant Cell Environ. 2020, 43, 649–661. [Google Scholar] [CrossRef]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2014, 5, 771. [Google Scholar] [CrossRef] [Green Version]
- Broxterman, S.E.; Schols, H.A. Interactions between pectin and cellulose in primary plant cell walls. Carbohydr. Polym. 2018, 192, 263–272. [Google Scholar] [CrossRef]
- Clara, S.-R.; Stefan, B.; Kian, H.; Friederike, S.; Belén, I.A.; Vera, V.; Cornelia, K.; Arun, S.; Markus, R.; Ernst, A.; et al. Chitinase-like1/pom-pom1 and its homolog CTL2 are glucan-interacting proteins important for cellulose biosynthesis in Arabidopsis. Plant Cell 2012, 24, 589–607. [Google Scholar]
- Wu, H.C.; Bulgakov, V.P.; Jinn, T.L. Pectin methylesterases: Cell wall remodeling proteins are required for plant response to heat stress. Front. Plant Sci. 2018, 9, 1612. [Google Scholar] [CrossRef] [Green Version]
- Rui, Y.; Dinneny, J.R. A wall with integrity: Surveillance and maintenance of the plant cell wall under stress. New Phytol. 2020, 225, 1428–1439. [Google Scholar] [CrossRef] [Green Version]
- Saxena, I.M. The plant cell wall: Barrier and facilitator of environmental perception. In Sensory Biology of Plants; Sopory, S., Ed.; Springer: Singapore, 2019; pp. 453–476. [Google Scholar]
- Wojtaszek, P. Oxidative burst: An early plant response to pathogen infection. Biochem. J. 1997, 322, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Qiu, X.; Lei, C.; Huang, L.; Li, X.; Hao, H.; Du, Z.; Wang, H.; Ye, H.; Beerhues, L.; Liu, B. Endogenous hydrogen peroxide is a key factor in the yeast extract-induced activation of biphenyl biosynthesis in cell cultures of Sorbus aucuparia. Planta 2012, 235, 217–223. [Google Scholar] [CrossRef]
- Vibhakar, C.; Sheena, A.; Rohan, V.P.; Jigna, G.T. Physiological function of phenolic compounds in plant defense system. In Phenolic Compounds; Farid, A.B., Ed.; IntechOpen: Rijeka, Croatia, 2021; Chapter 10. [Google Scholar]
- Vriet, C.; Hennig, L.; Laloi, C. Stress-induced chromatin changes in plants: Of memories, metabolites and crop improvement. Cell. Mol. Life Sci. 2015, 72, 1261–1273. [Google Scholar] [CrossRef]
- Yang, D.; Huang, Z.; Jin, W.; Xia, P.; Jia, Q.; Yang, Z.; Hou, Z.; Zhang, H.; Ji, W.; Han, R. DNA methylation: A new regulator of phenolic acids biosynthesis in Salvia miltiorrhiza. Ind. Crops Prod. 2018, 124, 402–411. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KEGG Pathway | ID | p-Value (NT_NT vs. NT_T) | p-Value (NT_T vs. PT_T) |
|---|---|---|---|
| Phenylpropanoid biosynthesis a,b | ko00940 | 2.72 × 10−9 | 0.045694574 |
| Terpenoid backbone biosynthesis a | ko00900 | 1.57 × 10−5 | ns |
| Plant-pathogen interaction a,b | ko04626 | 0.000256399 | 2.37 × 10−9 |
| Flavonoid biosynthesis a | ko00941 | 0.000422929 | ns |
| Plant hormone signal transduction a | ko04075 | 0.000495331 | ns |
| Circadian rhythm–plant b | ko04712 | ns | 0.011897832 |
| Oxidative phosphorylation b | ko00190 | ns | 0.025599648 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Luo, Z.-Q.; Yuan, J.; Wang, S.; Liu, J.; Su, P.; Zhou, J.-H.; Li, X.; Yang, J.; Guo, L.-P. Metabolic and Transcriptional Stress Memory in Sorbus pohuashanensis Suspension Cells Induced by Yeast Extract. Cells 2022, 11, 3757. https://doi.org/10.3390/cells11233757
Li Y, Luo Z-Q, Yuan J, Wang S, Liu J, Su P, Zhou J-H, Li X, Yang J, Guo L-P. Metabolic and Transcriptional Stress Memory in Sorbus pohuashanensis Suspension Cells Induced by Yeast Extract. Cells. 2022; 11(23):3757. https://doi.org/10.3390/cells11233757
Chicago/Turabian StyleLi, Yuan, Zhi-Qiang Luo, Jie Yuan, Sheng Wang, Juan Liu, Ping Su, Jun-Hui Zhou, Xiang Li, Jian Yang, and Lan-Ping Guo. 2022. "Metabolic and Transcriptional Stress Memory in Sorbus pohuashanensis Suspension Cells Induced by Yeast Extract" Cells 11, no. 23: 3757. https://doi.org/10.3390/cells11233757