
AIRE in Male Fertility: A New Hypothesis



Laboratory of Immunobiology, Institute of Molecular Genetics of the Czech Academy of Sciences, Vídeňská 1083, 142 20 Prague, Czech Republic
*
Author to whom correspondence should be addressed.
Cells 2022, 11(19), 3168; https://doi.org/10.3390/cells11193168
Submission received: 12 September 2022
/
Revised: 29 September 2022
/
Accepted: 8 October 2022
/
Published: 9 October 2022
(This article belongs to the Section Reproductive Cells and Development)
{kind=link}
{kind=link}
Abstract
:Male infertility affects approximately 14% of all European men, of which ~44% are characterized as idiopathic. There is an urgency to identify the factors that affect male fertility. One such factor, Autoimmune Regulator (AIRE), a protein found in the thymus, has been studied in the context of central tolerance functioning as a nuclear transcription modulator, responsible for the expression of tissue-restricted antigens in specialized thymic cells that prevent autoimmunity. While its expression in the testes remains enigmatic, we recently observed that sterility in mice correlates with the absence of Aire in the testes, regardless of the deficient expression in medullary thymic epithelial cells or cells of the hematopoietic system. By assessing the Aire transcript levels, we discovered that Sertoli cells are the exclusive source of Aire in the testes, where it most likely plays a non-immune role, suggesting an unknown mechanism by which testicular Aire regulates fertility. Here, we discuss these results in the context of previous reports which have suggested that infertility observed in Aire deficient mice is of an autoimmune aetiology. We present an alternative point of view for the role of Aire in testes in respect to fertility altering the perspective of how Aire’s function in the testes is currently perceived.
1. Introduction
The autoimmune regulator (Aire) is one of the most studied transcription factors and is known for its critical contribution to the establishment of immunological central tolerance, which occurs in the thymus [1]. Even though the expression of Aire in testes was recognized around the same time as in the thymus, its physiological function in this extrathymic organ remains enigmatic. Importantly, Aire deficient mice are infertile, but whether this phenotype is a consequence of the lack of Aire in the thymus, testes, or both is unclear. What is also puzzling is the exact cellular source of Aire in testes i.e., if its role is solely transcriptional and how it’s function is delivered. Several groups have presented data that suggests that testicular Aire performs a non-immune function, however their conclusions are not convincing. Therefore, we have decided to compare current published data to data recently generated in our lab. From what we have observed, we hope to bring to the forefront novel ideas regarding the role of Aire in testes and its relationship to the pool of Aire that is expressed in the thymus.
2. Materials and Methods
2.1. Breeding Experiment
Male mice used: wild type (Aire+/+), total body knock-out (Aire−/−), and males with conditionally depleted Aire in spermatogonia stem cells (VasaCreAirefl/fl) and primary spermatocytes (Smc1βiCreAirefl/fl). All males were housed in pairs with one wild–type female (C57Bl6). The breeding pairs were established at the beginning of their fertility period, i.e., six weeks of age. We established five independent breeding pairs (A–E) and monitored them for six-consecutive months. Each month the number of pups born was recorded. All mice were bred and raised at the animal facility of the Institute of Molecular Genetics in Prague. The protocols for their care were approved by the institutional review committee.
2.2. Testes Collection and Statistical Analysis
Testes were taken immediately after euthanizing the animal by cervical dislocation. All males were at the age of six weeks, i.e. at the beginning of their reproductive period. The epididymis and any other tissue (seminal vesicle, fat, etc.) was completely removed from the dissected testes and each testicle was weighted separately. The testis weight represents the average value from two testicles of a single male. We evaluated testicular size and weight from 10 males of each genotype. All graphs were created with GraphPad Prism version 5.00 for Windows (GraphPad Software, San Diego, CA, USA), and data was statistically evaluated by two-way ANOVA analysis.
2.3. Collection of Cells from Seminiferous Tubule
Testes from six-week old wild type males were processed as described elsewhere to obtain a single-cell suspension [2]. Briefly, decapsulated testes were treated with collagenase and DNase, followed by filtration. For identification of germ cell populations, we took advantage of Hoechst 33342 staining, which enables the discrimination of the cells according to ploidy [3,4]. Cells were FACS-sorted using a BD Influx high speed cell sorter (BD Biosciences, Franklin Lakes, NJ, USA). At least 10,000 cells per each population were dropped directly into lysis buffer (RLT), followed by the immediate isolation of RNA.
2.4. Isolation of RNA and qRT–PCR
Total RNA from FACS-sorted cells was immediately isolated using a commercially available kit (RNeasy MikroKit, QIAGEN, Hilden, Germany). Reverse transcription was done using a RevertAid RT Reverse Transcription Kit (Thermo Fisher Scientific, Waltham, MA, USA) and qPCR with SYBR™ Green PCR Master Mix (Applied Biosystems, Waltham, MA, USA). For the qPCR reaction, we used the LightCycler® 480 System (Roche, F. Hoffmann-La Roche AG, Basel, Switzerland) equipped with software for the analysis of gene expression. The level of gene expression was quantified relative to the house-keeping gene, Casc3. The sequences of primers used in reaction were as follows: Thy1: Fw 5’ AAGTCGGAACTCTTGGCACC 3’, Rev 5’ TCCAGGCGAAGGTTTTGGTT 3’; Scp1: Fw 5’ ACCGTTGGACAACGATTGCT 3’, Rev 5’ ATCCATTGCAAGTAAAAGCAACA 3’; Catsper 3: Fw 5’ CTTCAGTTTGGCCACGGTTG 3’, Rev 5’ CCCAGCTACGGCTACCTCTA 3’; Izumo1: Fw 5’GGGATGACCGGTGACTCTTG 3’, Rev 5’ ATGCTTCACCCAGCTCAGTC 3’; Wt1: Fw 5’ GAGAATCCGCAGGATCGCAG 3’, Rev 5’ TGAACTGGCCCGAGAAGTG 3’; and Casc3: Fw 5’ TTCGAGGTGTGCCTAACCA 3’, Rev 5’ GCTTAGCTCGACCACTCTGG 3’. All mentioned primers annealed at a temperature of 60 °C.
2.5. Immunofluorescence and Microscopy
Testes from six-week old wild type males were dissected, immediately immersed in Tissue-Tek® O.C.T. Compound (Sakura Finetek USA, Torrance, CA, USA) and frozen on dry ice. Immunofluorescence detection of proteins was done on cryo-sectioned tissue after fixation and permeabilization. Diluted antibodies were applied to these tissue sections and incubated for 2 h at room temperature. The primary antibodies used were as follows: SOX9 antibody (dilution 1:200, mouse monoclonal sc-166505), AIRE antibody (dilution 1:100, goat polyclonal sc-17986), and SCP3 antibody (dilution 1:200, rabbit polyclonal sc-33195). The origin of all antibodies was from Santa Cruz Biotechnology (Dallas, TX, USA). After several washes, these secondary antibodies were applied to the tissue sections: Alexa Fluor 488 (donkey anti-rabbit), Alexa Fluor 555 (donkey anti-mouse) and Alexa Fluor 647 (donkey anti-goat); all diluted 1:1000 and purchased from Thermo Fisher Scientific (Waltham, MA, USA). Images were taken using a Leica TCS SP5 confocal microscope (Wetzlar, Germany).
3. Results
3.1. Thymic Aire as an Essential Mediator of Central Immune Tolerance
The story of Aire started in 1997, when positional cloning identified mutations in the Aire gene as the single genetic defect underlying a disease called autoimmune polyendocrinopathy–candidiasis–ectodermal dystrophy (APECED, OMIM: 240300) [5,6]. APECED belongs to a category of rare human monogenic autoimmune diseases where the immune system malfunctions by attacking the body’s own tissues and organs. It is characterized by multiple autoimmune endocrinopathies, chronic mucocutaneous candidiasis, ectodermal dystrophies, as well as a plethora of symptoms including testicular failure [7]. Several Aire-deficient mouse strains were shown to recapitulate human APECED disease, including infertility [8,9]. A major finding explaining the role of Aire in the prevention of autoimmunity was that its expression was confined to medullary thymic epithelial cells (mTECs) [10]. In this respect, Aire functions as a transcription factor which promotes the “promiscuous” expression of a large repertoire of repressed genes, which are otherwise only expressed in the immune periphery as tissue-restricted antigens [11]. Aire achieves this “promiscuous” expression of tissue-restricted antigens by decoding epigenetic marks of repressive chromatin via its PHD1 and PHD2 domains and navigates transcriptional machinery to these silenced genes [12,13,14]. Recent studies estimate that mTECs express ~18,000 genes which represents ~85% of the protein encoded genome. Of these, Aire regulates ~4000 genes, which mainly encodes tissue-restricted antigens [15,16,17,18]. mTECs present the products of these genes on its surface as self-antigens that are required for the elimination of self-reactive T cells, or their conversion to Foxp3+ regulatory T cell lineage, both of which are required for the prevention of autoimmunity. Thus, the absence or malfunction of AIRE, such as that observed in APECED patients, leads to the non-expression and presentation of tissue-restricted antigens on mTECs and the non-removal of self-reactive thymocytes, resulting in the disruption of central tolerance and, consequently, the onset of autoimmunity [19].
3.2. Extrathymic Aire Expression
Because of its multiorgan manifestation and severity of autoimmunity, it was originally thought that nearly all symptoms observed in patients with APECED exhibited an autoimmune etiology [9]. However, this would only be credible if Aire is exclusively expressed in mTECs. By using various biochemical and genetic approaches, several groups convincingly showed that Aire is also expressed on the protein level in secondary lymphoid organs, mainly in the lymph nodes and spleen [20,21,22]. Thus, the question arises whether Aire in these extrathymic tissues acts in a similar manner to what has been observed in central tolerance. Indeed, initial studies using Aire-EGFP-reporter (Adig) mice [23] localized Aire expression to a unique subset of lymph node hematopoietic cells which were described as extrathymic Aire-expressing cells. The authors concluded that these cells represented a specialized, distinct, and nonconventional APC population that exhibited a CD45+ MHCII+ CD11cLOW EpCAM+ phenotype, influencing the induction of peripheral tolerance in an Aire-dependent fashion similar to that mediated by mTECs in the thymus [23,24].
Recently, we conducted experiments exploring the potential phenotypic diversity and the specific function of extrathymic Aire-expressing cells. We have shown that the expression of Aire protein in lymph nodes is confined to a rare subset of innate lymphoid cells (ILC) that display the characteristics of MHCII+ILC type 3 cells [25]. Since Aire in MHCII+ILC3 cells regulates IL-17 immune responses to Candida albicans infections [26], it is clear that Aire deficiency in these cells is the underpinning cause of chronic mucocutaneous candidiasis in patients with APECED. Hence, the immune function of Aire as a transcription regulator can be attuned in a cell-specific manner and functionally differs from its well-established role in mTECs in the enforcement of central tolerance.
3.3. Aire in Testes and Male Sterility
Interestingly, in addition to lymph nodes and spleen, the expression of the Aire protein has also been described in the testes [27,28]. Interestingly, since the testes are not classified as secondary lymphoid organs, a non-immune function of Aire could potentially be envisioned. It is important to reiterate that Aire whole body knock-out (Aire−/−) mice are infertile and patients with APECED suffer from testicular failure, which draws an interesting parallel. Thus, the obvious question is whether these reproduction-affected phenotypes are a consequence of Aire malfunction in mTECs, secondary lymphoid organs or testes. Interestingly, Kekäläinen et al. found that sterility in Aire−/− mouse males could be reversed in the absence of T and B cells, i.e., Aire−/−Rag1−/− mice were found to be fully fertile which led the authors to suggest an autoimmune aetiology of infertility in Aire−/− males [29]. This finding argues that it is either Aire expression in mTECs or in the peripheral hematopoietic system which controls the processes leading to autoimmune attack that results in infertility and precludes the involvement of testicular Aire in this process. However, several reports failed to provide support for such a conclusion.
While these studies confirmed that male Aire−/− mice suffer from infertility, no prominent lymphocyte infiltrates in the gonads were detected [8]. It was shown that autoantibodies against seminal vesicles and the epididymis appeared between weeks 21–25, although a full cadre was documented between weeks 48–49 [30]. The exception was the detection of autoantibodies against the prostate gland in 7–8 weeks old animals and peaks at 21–25 weeks of age, which was accompanied by pronounced prostatitis, which may affect the quality of sperm and hence fertility [30,31,32]. Thus, while it seems that lymphocyte infiltration, autoantibody production, and autoimmunity towards multiple reproductive targets contributes to infertility in male Aire−/− mice [33], these phenotypes are observed predominantly later in the life of these animals. However, this is in contrast to the fact that 85% of males are sterile from the beginning of their reproductive cycle, and incapable of producing any litters [9]. Moreover, no evidence has been presented demonstrating that autoantibodies in Aire−/− mice have a pathogenic role in the onset of autoimmunity [34].
Considering this finding, an alternative view is rather apparent: the primary cause of infertility in male Aire−/− mice is not the result of the autoimmunity caused by the breach of central tolerance. To validate this notion, the prediction that Aire deficiency in mTECs or secondary lymphoid organs (lymph nodes and spleen) does not affect male fertility has already been functionally tested using a mouse strain with the floxed Aire allele (Airefl/fl). This strain was mated with mice harboring a TEC-specific Cre driver, Foxn1Cre, or with a strain with a hematopoietic-specific Cre driver, Vav1Cre [35]. A detailed analysis revealed a significant decrease of Aire mRNA in the thymus of Foxn1Cre+Airefl/fl and secondary lymphoid organs of Vav1Cre+Airefl/fl animals. The expression of Aire in testes from both Foxn1Cre+Airefl/fl and Vav1Cre+Airefl/fl was comparable to testes from Aire sufficient animals. Importantly, both Foxn1Cre+Airefl/fl and Vav1Cre+Airefl/fl males were fully fertile. Since Aire protein expression had been localized only to the thymus, secondary lymphoid organs, and testes, the infertility observed in male Aire−/− mice correlated with the absence of Aire expression in the testes and not with its deficiency in the thymic stroma or hematopoietic system [35]. To the best of our knowledge, this experiment provided the first genetic evidence for a functional link between fertility and Aire expression in the testes.
3.4. Which Cells in Seminiferous Tubule Express Aire?
The paradigm-changing observation described above leads to one question: what is the mechanism(s) by which Aire deficiency in the testes initiates an animal’s infertility? The first part of this question concerns the precise cellular source of Aire in the testes, which then allows one to consider the possible scenarios regarding how the function of Aire is delivered to support fertility.
Thus far, the precise cellular localization of the Aire protein production in the testes remains highly controversial. Notably, Schaller et al. claimed to find Aire protein in the nuclei of germ cells, specifically in early spermatocytes and spermatogonia but not in spermatids or Sertoli cells. For the immunohistochemical detection of Aire, the authors used the Aire specific monoclonal antibody, clone 5H12, which is routinely used by many labs for detection in thymic medullary cells [27]. While the cells which they stated contained Aire were located close to the inner peripheral circumference of the seminiferous tubules, their identification was only positional, with no cell specific colocalization marker (Smc1β for early spermatocytes or Thy1 for spermatogonia stem cells). Therefore, the only conclusion which can be made from this study is that Aire expression in the testes is confined to the seminiferous tubules.
Radhakrishnan et al. found that Aire mRNA is expressed in all evaluated stages of germ cell development, including the spermatogonia from neonatal and adult testes, primary and secondary spermatocytes, and spermatids [28]. The authors also showed that the Aire expression pattern in mouse testes is dynamic, with the highest level of Aire mRNA observed during the first week of life [28]. The authors proposed that this pattern of expression correlates with the requirement for Aire abundancy during the first wave of spermatogenesis. Interestingly, four weeks after birth, a dramatic ~10-fold decline in Aire expression was observed as the animals sexually matured. However, the presence of Aire mRNA and its decrease between the first and fourth week of life did not correlate with protein levels where no changes were seen [28]. Nevertheless, the common denominator in both studies discussed above is the suggestion that Aire is expressed in germ cells residing within seminiferous tubules.
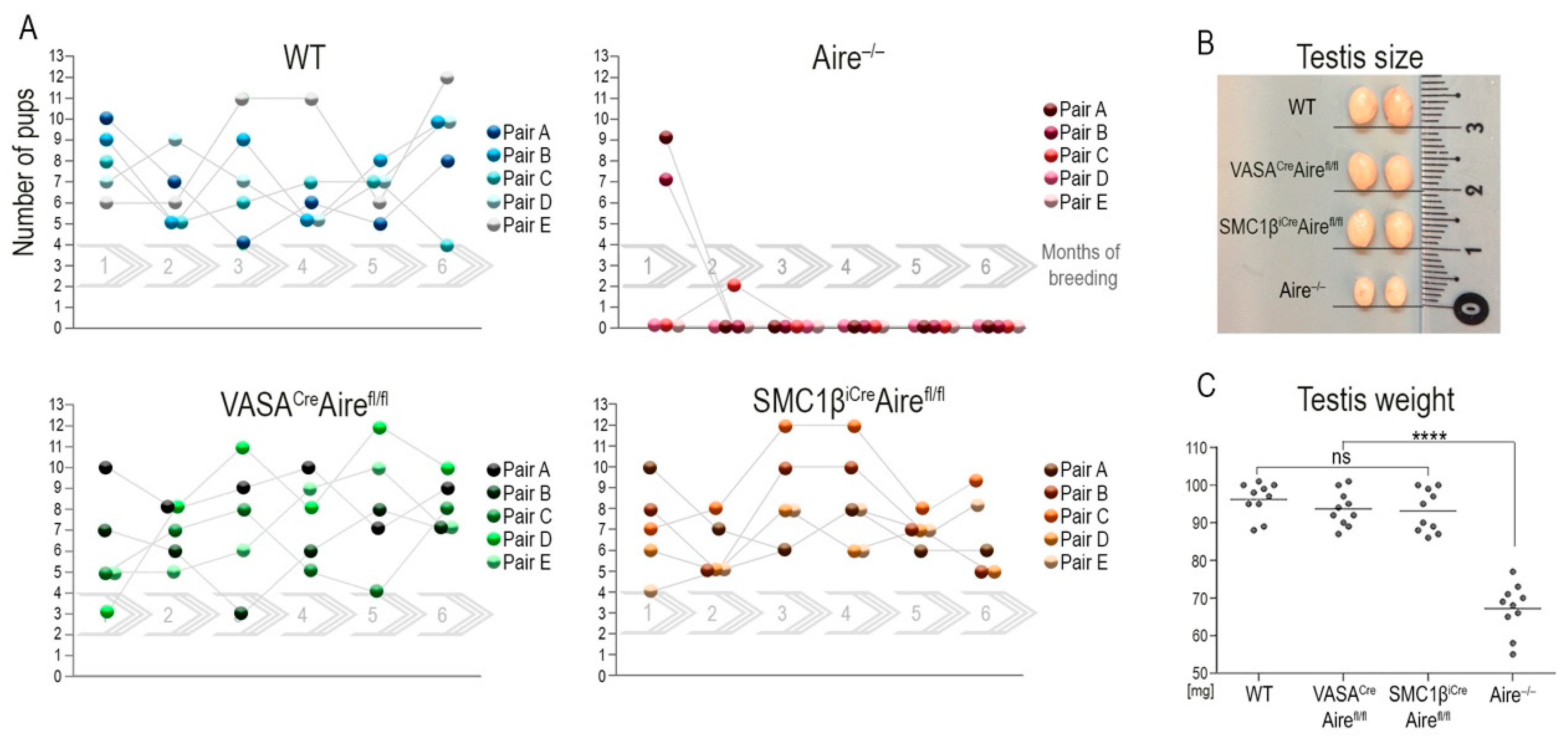
If the cellular source of testicular Aire in the testes is indeed in germ cells and this pool of Aire is required for fertility, then the Aire deficiency in germ cells should recapitulate the infertility observed in male Aire−/− mice. To test this prediction, we again utilized Airefl/fl mice which were crossed with animals that expressed either VasaCre or Smc1βiCre drivers. The mouse vasa homologue (Vasa) gene showed spermatogonia stem cell expression [36] from embryonic day 12.5 onwards after entering the gonadal anlage [37]. The Smc1β gene product is a member of a cohesion complex that forms a ring structure around meiotic chromosomes and is expressed at the onset of meiosis I of primary spermatocytes (SCI). Vasa as well as Smc1β are specifically expressed in germ cells but not in testicular somatic cells [38]. Thus, the utility of the Vasa and Smc1β promoters as Cre drivers makes them suitable for conditional depletion of desired floxed alleles with a specific effect on spermatogonia stem cells and primary spermatocytes, and their progeny-secondary spermatocytes (SCII), round spermatids and sperm (RS+S). VasaCreAirefl/fl and Smc1βiCreAirefl/fl males were cross-bred with wild type females over a six-month period. The breeding performance was closely monitored and compared to wild-type couples. There was no significant difference in fertility between the two transgenic males, whereas the total body Aire KO gave birth to only a few pups at the beginning of the reproduction cycle (Figure 1A). The size and weight of the testes correlated with the results (Figure 1B) with no significant difference observed between wild-type, VasaCreAirefl/fl and Smc1βiCreAirefl/fl males. These parameters of Aire−/− male testes were significantly reduced to 55–80% when compared to wild-type males (Figure 1C).
It is important to note that, in order to identify Aire expressing cells in testes, Radhakrishnan and colleagues FACS-sorted cell populations from the flow cytometric profile of spermatocyte populations stained with Hoechst 33342. However, the generated Hoechst profile seemed rather non-standard when compared to previously published analyses [3,4,39]. Additionally, it is also unclear as to which cell types were sorted and analyzed [28]. This caveat weakens the conclusion that Aire mRNA is present in all germ cell types. Moreover, the authors omitted the analysis of Sertoli cells, which are also present in the seminiferous tubules, as a potential source of Aire.
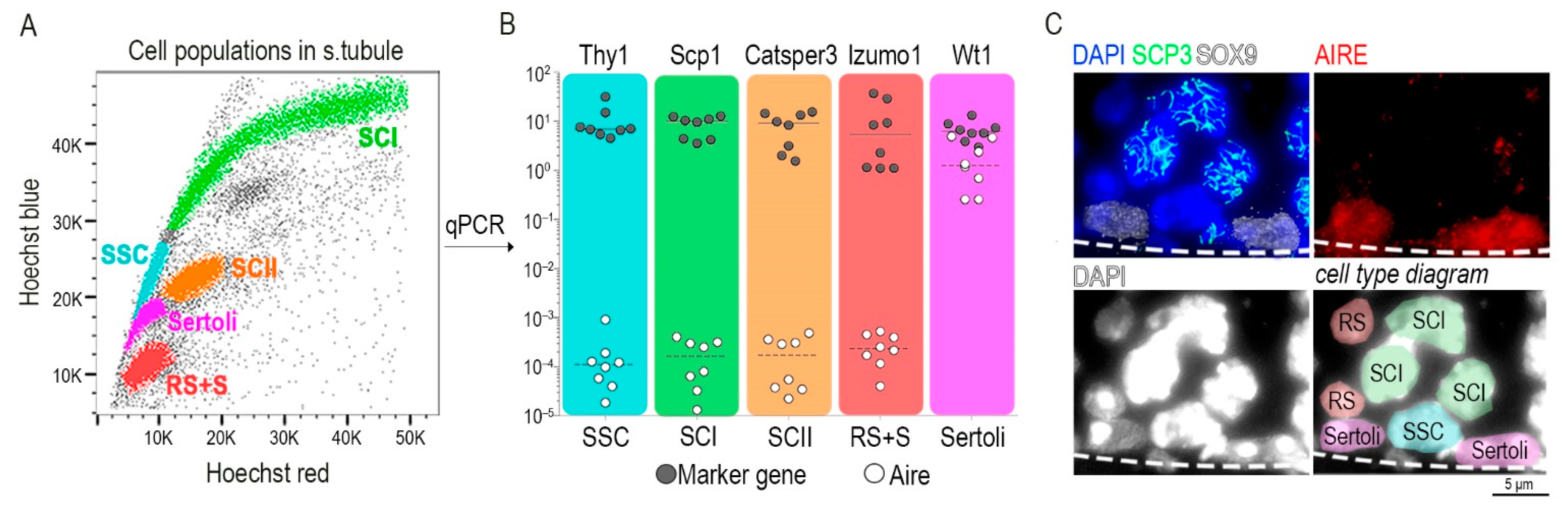
To precisely identify the cellular source of Aire, we isolated all germ cell types present in seminiferous tubules (Figure 2A), along with Sertoli cells [2]. All indicated cell types were FACS-sorted directly into the lysis buffer for isolation of mRNA. Using RT-qPCR analysis for each sorted cell type, we quantified Aire transcript levels and utilized the following cell-type specific marker genes: Thy1 (Thy1 cell surface antigen; [40]) in spermatogonia stem cells, Scp1 (Synaptonemal complex protein 1; [41]) in primary spermatocytes, Catsper 3 (Cation Channel Sperm Associated 3; [42]) in secondary spermatocytes, Izumo1 (Izumo Sperm-Egg Fusion 1; [43]) from round spermatid and sperm, and Wt1 (Wilms Tumor Protein; [44]) for Sertoli cells. Unexpectedly, we found that Aire was exclusively present in Sertoli cells. All other cell types showed a 10,000–100,000 times lower expression of Aire mRNA in comparison to Sertoli cells (Figure 2B). In addition, Aire on the protein level is also detectable only in Sertoli and no other cells in the seminiferous tubule (Figure 2C).
Our finding is further supported by the recent work of Grive et al. who identified the genes specifically expressed by meiocytes in the first and second round of meiotic division as well as the population of spermatogonia stem cells [45]. They not only identified conserved markers of germ cells but also their novel genes. Nevertheless, Aire was not detected in any of the described spermatogenic populations (GEO: GSE121904, blasted Aire exon 1 in dataset) [45,46,47]. In contrast, we identified an in silico Aire gene expressed in highly purified Sertoli cells from a transgenic SOX9-GFP male dataset. The Aire transcript was present beginning at day 10, and higher levels were observed from day 18 onwards during male development (GEO: GSE59698) [48].
4. Discussion
The main objective of this article was to compare our recently generated data to previously published data sets, some of which present inconsistent or unclear outcomes that describe the role of Aire in male fertility. Our work is an alternative to recent formulated thoughts of the potential role of Aire in testes, which are based on previously published data that Aire in testes is localized to germ cells [50]. Recent genetic data [35], in combination with our presented results, support the view that it is the Aire in Sertoli cells which might be important for the maintenance of fertility. Even if Aire is present in spermatogonia stem cells or primary spermatocytes as suggested by other studies [27,28], its depletion in these cells would not impact male fertility. This alternative point of view regarding the function of testicular Aire provides a new perspective on the role of Aire in Sertoli cells in respect to the control of male fertility.
What could be the possible role and mechanism of Aire in testes? Some insight may be provided by Foxn1Cre+Airefl/fl mice which appear to be fully fertile for at least the first twelve months of life [2,35]. This provides evidence that even in the absence of Aire in the thymic stroma resulting in the continuous process of multiorgan autoimmunity, the testes are not functionally affected. Thus, since infertility which arises as a result of autoimmunity can only occur in the absence of Aire in the testes and later in life, this may provide a clue for the role of testicular Aire. Notably, the testes are viewed as an immune privileged organ in which Sertoli cells are critical for the generation and maintenance of a barrier within seminiferous tubules [51]. Aire in Sertoli cells could be directly involved in this process either as a transcription factor which regulates the expression of genes required for barrier function or some other function which has yet to be determined. Consistent with this proposal, since the barrier in Foxn1Cre+Airefl/fl mice remains intact and provides protection from autoimmune attack, it maintains fertility throughout life. However, this barrier in Aire−/− mice may be improperly formed and non-functional allowing the penetration of autoantibodies and autoreactive cells to be critical reproduction targets. Since in Aire−/− mice an autoimmune attack is the result of Aire deficiency in mTECs, the accompanying lack of testicular Aire is not a direct causative factor of infertility but rather a consequence of its specific role in Sertoli cells. While this scenario would explain the onset of infertility with autoimmune aetiology later in life, it still does not provide an explanation which states why the vast majority of Aire−/− mice cannot reproduce at all. Clearly, other mechanisms dictate how Aire in Sertoli cells modulate fertility, which warrants further investigation.
Another aspect of this issue is the fact that dominant mutations in the Aire gene affect approximately 1 in 550 people [12]. However, the effect of dominant mutations in Aire on patient fertility has not been adequately addressed.
5. Conclusions
Together, the presence of Aire in testes challenges the traditional view of Aire as a unique immune-related transcription factor and should be an invitation to members of this field of study to revisit its physiological role in a much broader context examining multiple regulatory mechanisms that operate in a cell/tissue-dependent fashion. In this context, it was our intention to provide a conceptually new hypothesis that could begin to resolve the issue of Aire-mediated fertility from a completely different perspective. As our studies continue to progress, future experimentation and analysis of a generated conditional Aire knock-out in Sertoli cells will be an import step in strengthening our current data identifying Sertoli cells as the source of Aire in seminiferous tubules.
Author Contributions
Conceptualization, J.P., J.M. and D.F.; methodology, J.P.; formal analysis, J.P., J.M. and D.F.; resources, D.F.; data curation, J.P. and D.F.; writing—original draft preparation, J.P. and D.F.; writing—review and editing, J.P., J.M. and D.F.; visualization, J.P.; supervision, D.F.; project administration, D.F.; funding acquisition, D.F. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Grant 20-30350S from The Grant Agency of the Czech Republic (GACR).
Institutional Review Board Statement
All animal experiments were approved by the Ethical committee of the Institute of Molecular Genetics of the CAS, and by the ethical committee of the Czech Academy of Sciences (protocol 27/2020). Animals care and handling were in accordance with the Czech Act No. 246/1992 Coll. and European directive 2010/63/EU.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We would like to thank to rer. nat. Rolf Jessberger (Institute of Physiological Chemistry, TU Dresden, Germany) for providing the transgenic mouse strains VASACre and SCM1βiCre.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial interests which could be construed as a potential conflict of interest.
References
- Kyewski, B.; Klein, L. A central role for central tolerance. Annu. Rev. Immunol. 2006, 24, 571–606. [Google Scholar] [CrossRef] [PubMed]
- Petrusova, J.; Manning, J.; Kubovčiak, J.; Kolář, M.; Filipp, D. Two complementary approaches for efficient isolation of Sertoli cells for transcriptomic analysis. Front. Cell Dev. Biol. 2022, 10, 972017. [Google Scholar] [CrossRef] [PubMed]
- Bastos, H.; Lassalle, B.; Chicheportiche, A.; Riou, L.; Testart, J.; Allemand, I.; Fouchet, P. Flow cytometric characterization of viable meiotic and postmeiotic cells by Hoechst 33342 in mouse spermatogenesis. Cytom. Part A 2005, 65, 40–49. [Google Scholar] [CrossRef]
- Gaysinskaya, V.; Soh, I.Y.; van der Heijden, G.W.; Bortvin, A. Optimized flow cytometry isolation of murine spermatocytes. Cytom. Part A 2014, 85, 556–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finnish-German, A.C. An autoimmune disease, APECED, caused by mutations in a novel gene featuring two PHD-type zinc-finger domains. Nat. Genet. 1997, 17, 399–403. [Google Scholar] [CrossRef]
- Nagamine, K.; Peterson, P.; Scott, H.S.; Kudoh, J.; Minoshima, S.; Heino, M.; Krohn, K.J.; Lalioti, M.D.; Mullis, P.E.; Antonarakis, S.E.; et al. Positional cloning of the APECED gene. Nat. Genet. 1997, 17, 393–398. [Google Scholar] [CrossRef]
- Bjorses, P.; Aaltonen, J.; Horelli-Kuitunen, N.; Yaspo, M.L.; Peltonen, L. Gene defect behind APECED: A new clue to autoimmunity. Hum. Mol. Genet. 1998, 7, 1547–1553. [Google Scholar] [CrossRef] [Green Version]
- Anderson, M.S.; Venanzi, E.S.; Klein, L.; Chen, Z.; Berzins, S.P.; Turley, S.J.; von Boehmer, H.; Bronson, R.; Dierich, A.; Benoist, C.; et al. Projection of an immunological self shadow within the thymus by the aire protein. Science 2002, 298, 1395–1401. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, C.; Winqvist, O.; Puhakka, L.; Halonen, M.; Moro, A.; Kampe, O.; Eskelin, P.; Pelto-Huikko, M.; Peltonen, L. Aire deficient mice develop multiple features of APECED phenotype and show altered immune response. Hum. Mol. Genet. 2002, 11, 397–409. [Google Scholar] [CrossRef]
- Mathis, D.; Benoist, C. Aire. Annu. Rev. Immunol. 2009, 27, 287–312. [Google Scholar] [CrossRef]
- Klein, L.; Kyewski, B.; Allen, P.M.; Hogquist, K.A. Positive and negative selection of the T cell repertoire: What thymocytes see (and don’t see). Nat. Rev. Immunol. 2014, 14, 377–391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramson, J.; Husebye, E.S. Autoimmune regulator and self-tolerance-molecular and clinical aspects. Immunol. Rev. 2016, 271, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Bansal, K.; Yoshida, H.; Benoist, C.; Mathis, D. The transcriptional regulator Aire binds to and activates super-enhancers. Nat. Immunol. 2017, 18, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Abramson, J.; Giraud, M.; Benoist, C.; Mathis, D. Aire’s partners in the molecular control of immunological tolerance. Cell 2010, 140, 123–135. [Google Scholar] [CrossRef] [Green Version]
- Danan-Gotthold, M.; Guyon, C.; Giraud, M.; Levanon, E.Y.; Abramson, J. Extensive RNA editing and splicing increase immune self-representation diversity in medullary thymic epithelial cells. Genome Biol. 2016, 17, 219. [Google Scholar] [CrossRef] [Green Version]
- Sansom, S.N.; Shikama-Dorn, N.; Zhanybekova, S.; Nusspaumer, G.; Macaulay, I.C.; Deadman, M.E.; Heger, A.; Ponting, C.P.; Hollander, G.A. Population and single-cell genomics reveal the Aire dependency, relief from Polycomb silencing, and distribution of self-antigen expression in thymic epithelia. Genome Res. 2014, 24, 1918–1931. [Google Scholar] [CrossRef] [Green Version]
- Brennecke, P.; Reyes, A.; Pinto, S.; Rattay, K.; Nguyen, M.; Kuchler, R.; Huber, W.; Kyewski, B.; Steinmetz, L.M. Single-cell transcriptome analysis reveals coordinated ectopic gene-expression patterns in medullary thymic epithelial cells. Nat. Immunol. 2015, 16, 933–941. [Google Scholar] [CrossRef] [Green Version]
- Meredith, M.; Zemmour, D.; Mathis, D.; Benoist, C. Aire controls gene expression in the thymic epithelium with ordered stochasticity. Nat. Immunol. 2015, 16, 942–949. [Google Scholar] [CrossRef] [Green Version]
- Husebye, E.S.; Anderson, M.S.; Kampe, O. Autoimmune polyendocrine syndromes. New Engl. J. Med. 2018, 378, 1132–1141. [Google Scholar] [CrossRef]
- Heino, M.; Peterson, P.; Kudoh, J.; Nagamine, K.; Lagerstedt, A.; Ovod, V.; Ranki, A.; Rantala, I.; Nieminen, M.; Tuukkanen, J.; et al. Autoimmune regulator is expressed in the cells regulating immune tolerance in thymus medulla. Biochem. Biophys. Res. Commun 1999, 257, 821–825. [Google Scholar] [CrossRef]
- Halonen, M.; Pelto-Huikko, M.; Eskelin, P.; Peltonen, L.; Ulmanen, I.; Kolmer, M. Subcellular location and expression pattern of autoimmune regulator (Aire), the mouse orthologue for human gene defective in autoimmune polyendocrinopathy candidiasis ectodermal dystrophy (APECED). J. Histochem. Cytochem. 2001, 49, 197–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adamson, K.A.; Pearce, S.H.; Lamb, J.R.; Seckl, J.R.; Howie, S.E. A comparative study of mRNA and protein expression of the autoimmune regulator gene (Aire) in embryonic and adult murine tissues. J. Pathol. 2004, 202, 180–187. [Google Scholar] [CrossRef]
- Gardner, J.M.; Devoss, J.J.; Friedman, R.S.; Wong, D.J.; Tan, Y.X.; Zhou, X.; Johannes, K.P.; Su, M.A.; Chang, H.Y.; Krummel, M.F.; et al. Deletional tolerance mediated by extrathymic Aire-expressing cells. Science 2008, 321, 843–847. [Google Scholar] [CrossRef] [Green Version]
- Gardner, J.M.; Metzger, T.C.; McMahon, E.J.; Au-Yeung, B.B.; Krawisz, A.K.; Lu, W.; Price, J.D.; Johannes, K.P.; Satpathy, A.T.; Murphy, K.M.; et al. Extrathymic Aire-expressing cells are a distinct bone marrow-derived population that induce functional inactivation of CD4(+) T cells. Immunity 2013, 39, 560–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamano, T.; Dobes, J.; Voboril, M.; Steinert, M.; Brabec, T.; Zietara, N.; Dobesova, M.; Ohnmacht, C.; Laan, M.; Peterson, P.; et al. Aire-expressing ILC3-like cells in the lymph node display potent APC features. J. Exp. Med. 2019, 216, 1027–1037. [Google Scholar] [CrossRef] [Green Version]
- Dobes, J.; Ben-Nun, O.; Binyamin, A.; Stoler-Barak, L.; Oftedal, B.E.; Goldfarb, Y.; Kadouri, N.; Gruper, Y.; Givony, T.; Zalayat, I.; et al. Extrathymic expression of Aire controls the induction of effective TH17 cell-mediated immune response to Candida albicans. Nat. Immunol. 2022, 23, 1098–1108. [Google Scholar] [CrossRef] [PubMed]
- Schaller, C.E.; Wang, C.L.; Beck-Engeser, G.; Goss, L.; Scott, H.S.; Anderson, M.S.; Wabl, M. Expression of Aire and the early wave of apoptosis in spermatogenesis. J. Immunol. 2008, 180, 1338–1343. [Google Scholar] [CrossRef] [Green Version]
- Radhakrishnan, K.; Bhagya, K.P.; Kumar, A.T.; Devi, A.N.; Sengottaiyan, J.; Kumar, P.G. Autoimmune regulator (AIRE) is expressed in spermatogenic cells, and it altered the expression of several nucleic-acid-binding and cytoskeletal proteins in Germ cell 1 spermatogonial (GC1-spg) cells. Mol. Cell Proteomics 2016, 15, 2686–2698. [Google Scholar] [CrossRef] [Green Version]
- Kekalainen, E.; Pontynen, N.; Meri, S.; Arstila, T.P.; Jarva, H. Autoimmunity, not a developmental defect, is the cause for subfertility of Autoimmune regulator (Aire) deficient mice. Scand. J. Immunol. 2015, 81, 298–304. [Google Scholar] [CrossRef]
- Warren, B.D.; Ahn, S.H.; Brittain, K.S.; Nanjappa, M.K.; Wang, H.; Wang, J.; Blanco, G.; Sanchez, G.; Fan, Y.; Petroff, B.K.; et al. Multiple lesions contribute to infertility in males lacking autoimmune regulator. Am. J. Pathol. 2021, 191, 1592–1609. [Google Scholar] [CrossRef]
- Hou, Y.; DeVoss, J.; Dao, V.; Kwek, S.; Simko, J.P.; McNeel, D.G.; Anderson, M.S.; Fong, L. An aberrant prostate antigen-specific immune response causes prostatitis in mice and is associated with chronic prostatitis in humans. J. Clin. Invest. 2009, 119, 2031–2041. [Google Scholar] [CrossRef] [PubMed]
- Motrich, R.D.; Maccioni, M.; Molina, R.; Tissera, A.; Olmedo, J.; Riera, C.M.; Rivero, V.E. Reduced semen quality in chronic prostatitis patients that have cellular autoimmune response to prostate antigens. Hum. Reprod. 2005, 20, 2567–2572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, X.; Zhang, Y.; Wang, X.; Zhang, R.; Yang, W. The role of AIRE deficiency in infertility and its potential pathogenesis. Front. Immunol. 2021, 12, 641164. [Google Scholar] [CrossRef] [PubMed]
- Gavanescu, I.; Benoist, C.; Mathis, D. B cells are required for Aire-deficient mice to develop multi-organ autoinflammation: A therapeutic approach for APECED patients. Proc. Natl. Acad. Sci. USA 2008, 105, 13009–13014. [Google Scholar] [CrossRef] [Green Version]
- Dobes, J.; Edenhofer, F.; Voboril, M.; Brabec, T.; Dobesova, M.; Cepkova, A.; Klein, L.; Rajewsky, K.; Filipp, D. A novel conditional Aire allele enables cell-specific ablation of the immune tolerance regulator Aire. Eur. J. Immunol. 2018, 48, 546–548. [Google Scholar] [CrossRef] [Green Version]
- Fujiwara, Y.; Komiya, T.; Kawabata, H.; Sato, M.; Fujimoto, H.; Furusawa, M.; Noce, T. Isolation of a DEAD-family protein gene that encodes a murine homolog of Drosophila vasa and its specific expression in germ cell lineage. Proc. Natl. Acad. Sci. USA 1994, 91, 12258–12262. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.S.; Toyooka, Y.; Akasu, R.; Katoh-Fukui, Y.; Nakahara, Y.; Suzuki, R.; Yokoyama, M.; Noce, T. The mouse homolog of Drosophila Vasa is required for the development of male germ cells. Genes Dev. 2000, 14, 841–853. [Google Scholar] [CrossRef]
- Revenkova, E.; Eijpe, M.; Heyting, C.; Gross, B.; Jessberger, R. Novel meiosis-specific isoform of mammalian SMC1. Mol. Cell Biol. 2001, 21, 6984–6998. [Google Scholar] [CrossRef] [Green Version]
- Petrusova, J.; Havalda, R.; Flachs, P.; Venit, T.; Darasova, A.; Hulkova, L.; Sztacho, M.; Hozak, P. Focal adhesion protein Vinculin is required for proper meiotic progression during mouse spermatogenesis. Cells 2022, 11, 2013. [Google Scholar] [CrossRef]
- Baazm, M.; Abolhassani, F.; Abbasi, M.; Habibi Roudkenar, M.; Amidi, F.; Beyer, C. An improved protocol for isolation and culturing of mouse spermatogonial stem cells. Cell Reprogram 2013, 15, 329–336. [Google Scholar] [CrossRef]
- Yuan, L.; Liu, J.G.; Zhao, J.; Brundell, E.; Daneholt, B.; Hoog, C. The murine SCP3 gene is required for synaptonemal complex assembly, chromosome synapsis, and male fertility. Mol. Cell 2000, 5, 73–83. [Google Scholar] [CrossRef]
- Mata-Rocha, M.; Hernandez-Sanchez, J.; Guarneros, G.; de la Chesnaye, E.; Sanchez-Tusie, A.A.; Trevino, C.L.; Felix, R.; Oviedo, N. The transcription factors Sox5 and Sox9 regulate Catsper1 gene expression. FEBS Lett. 2014, 588, 3352–3360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, S.A.; Aitken, J.; Baker, M.A. Phosphorylation of Izumo1 and Its Role in Male Infertility. Asian J. Androl. 2015, 17, 708–710. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zhang, L.; Cui, X.; Lin, X.; Li, Y.; Wang, Y.; Wang, Y.; Qin, Y.; Chen, D.; Han, C.; et al. Wt1 directs the lineage specification of sertoli and granulosa cells by repressing Sf1 expression. Development 2017, 144, 44–53. [Google Scholar] [CrossRef] [Green Version]
- Grive, K.J.; Hu, Y.; Shu, E.; Grimson, A.; Elemento, O.; Grenier, J.K.; Cohen, P.E. Dynamic transcriptome profiles within spermatogonial and spermatocyte populations during postnatal testis maturation revealed by single-cell sequencing. PLoS Genet. 2019, 15, e1007810. [Google Scholar] [CrossRef] [Green Version]
- Hermann, B.P.; Cheng, K.; Singh, A.; Roa-De La Cruz, L.; Mutoji, K.N.; Chen, I.C.; Gildersleeve, H.; Lehle, J.D.; Mayo, M.; Westernstroer, B.; et al. The mammalian spermatogenesis single-cell transcriptome, from spermatogonial stem cells to spermatids. Cell Rep. 2018, 25, 1650–1667 e1658. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef]
- Zimmermann, C.; Stevant, I.; Borel, C.; Conne, B.; Pitetti, J.L.; Calvel, P.; Kaessmann, H.; Jegou, B.; Chalmel, F.; Nef, S. Research resource: The dynamic transcriptional profile of sertoli cells during the progression of spermatogenesis. Mol. Endocrinol. 2015, 29, 627–642. [Google Scholar] [CrossRef] [Green Version]
- Jégou, B.; Skinner, M.K. Male Reproduction. In Encyclopedia of Reproduction, 2nd ed.; Skinner, M.K., Ed.; Academic Press: Oxford, 2018; ISBN 9780128118993. [Google Scholar]
- Forsdyke, D.R. When few survive to tell the tale: Thymus and gonad as auditioning organs: Historical overview. Theory Biosci. 2020, 139, 95–104. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Mruk, D.D.; Xia, W.; Bonanomi, M.; Silvestrini, B.; Cheng, C.Y. Effective delivery of male contraceptives behind the blood-testis barrier (BTB)-lesson from adjudin. Curr. Med. Chem. 2016, 23, 701–713. [Google Scholar] [CrossRef]
Figure 1.
Depletion of Aire in spermatogonia stem cells and spermatocytes does not affect male fertility. (A) Male mice with conditionally depleted Aire in spermatogonia stem cells (VasaCreAirefl/fl) and primary spermatocytes (Smc1βiCreAirefl/fl) exhibit fertility that is comparable to wild-type males in terms of the number of pups per litter. In contrast, whole-body Aire knock-out (Aire−/−) mice produced only a few pups at the beginning of their reproduction period. We established five independent breeding pairs (A–E) and monitored them for six-consecutive months (1–6, gray continuous arrows). (B) The size of the testes in VasaCreAirefl/fl as well as Smc1βiCreAirefl/fl males was fully comparable to WT controls. In contrast, six–week–old Aire−/− males had significantly smaller testes. (C) The decrease in testes size was confirmed by assessing their weight. p < 0.0001 = **** and ns = not significant.
Figure 1.
Depletion of Aire in spermatogonia stem cells and spermatocytes does not affect male fertility. (A) Male mice with conditionally depleted Aire in spermatogonia stem cells (VasaCreAirefl/fl) and primary spermatocytes (Smc1βiCreAirefl/fl) exhibit fertility that is comparable to wild-type males in terms of the number of pups per litter. In contrast, whole-body Aire knock-out (Aire−/−) mice produced only a few pups at the beginning of their reproduction period. We established five independent breeding pairs (A–E) and monitored them for six-consecutive months (1–6, gray continuous arrows). (B) The size of the testes in VasaCreAirefl/fl as well as Smc1βiCreAirefl/fl males was fully comparable to WT controls. In contrast, six–week–old Aire−/− males had significantly smaller testes. (C) The decrease in testes size was confirmed by assessing their weight. p < 0.0001 = **** and ns = not significant.

Figure 2.
Sertoli cells express Aire-coding mRNA. (A) The cell population from within the seminiferous tubule was stained with Hoechst 33342 and visualized in Hoechst red and blue channels. A color code was applied to the following cell types: turquoise spermatogonia stem cell (SSC), green primary spermatocytes (SCI), orange secondary spermatocytes (SCII), red round spermatids and sperm (RS+S), magenta Sertoli cells (Sertoli). (B) A single cell suspension from (A) was FACS-sorted and the indicated cell-types were prepared for mRNA isolation and qRT–PCR. Only Sertoli cells expressed physiologically relevant levels of Aire mRNA. The y axis represents the relative transcript levels of Aire and marker genes normalized to the Casc3 gene. (C) Protein immunodetection on the testicular tissue-sections showed the localization of AIRE in the nuclei of Sertoli cells (upper right panel) identified by their positivity to SOX9 (white color, upper left panel). SCP3 is a marker of primary spermatocytes (SCI). Spermatogonia stem cells (SSC), typically positioned between two Sertoli cells at the base of seminiferous tubule and round spermatid (RS), are identified as DAPI+SOX9−SCP3−, exhibiting a typical nuclear morphology [49]. The dashed line demarcates the edge of the seminiferous tubule. Cell types within the diagram (lower right panel) correspond to the color scheme used in qPCR.
Figure 2.
Sertoli cells express Aire-coding mRNA. (A) The cell population from within the seminiferous tubule was stained with Hoechst 33342 and visualized in Hoechst red and blue channels. A color code was applied to the following cell types: turquoise spermatogonia stem cell (SSC), green primary spermatocytes (SCI), orange secondary spermatocytes (SCII), red round spermatids and sperm (RS+S), magenta Sertoli cells (Sertoli). (B) A single cell suspension from (A) was FACS-sorted and the indicated cell-types were prepared for mRNA isolation and qRT–PCR. Only Sertoli cells expressed physiologically relevant levels of Aire mRNA. The y axis represents the relative transcript levels of Aire and marker genes normalized to the Casc3 gene. (C) Protein immunodetection on the testicular tissue-sections showed the localization of AIRE in the nuclei of Sertoli cells (upper right panel) identified by their positivity to SOX9 (white color, upper left panel). SCP3 is a marker of primary spermatocytes (SCI). Spermatogonia stem cells (SSC), typically positioned between two Sertoli cells at the base of seminiferous tubule and round spermatid (RS), are identified as DAPI+SOX9−SCP3−, exhibiting a typical nuclear morphology [49]. The dashed line demarcates the edge of the seminiferous tubule. Cell types within the diagram (lower right panel) correspond to the color scheme used in qPCR.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Petrusová, J.; Manning, J.; Filipp, D. AIRE in Male Fertility: A New Hypothesis. Cells 2022, 11, 3168. https://doi.org/10.3390/cells11193168
AMA Style
Petrusová J, Manning J, Filipp D. AIRE in Male Fertility: A New Hypothesis. Cells. 2022; 11(19):3168. https://doi.org/10.3390/cells11193168
Chicago/Turabian StylePetrusová, Jana, Jasper Manning, and Dominik Filipp. 2022. "AIRE in Male Fertility: A New Hypothesis" Cells 11, no. 19: 3168. https://doi.org/10.3390/cells11193168
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.