
Subgenome Discrimination in Brassica and Raphanus Allopolyploids Using Microsatellites

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microsatellite Mining
2.2. Designing Microsatellite Pre-Labeled Oligoprobes
2.3. Plant Samples
2.4. Chromosome Preparation
2.5. FISH and Karyotyping
3. Results
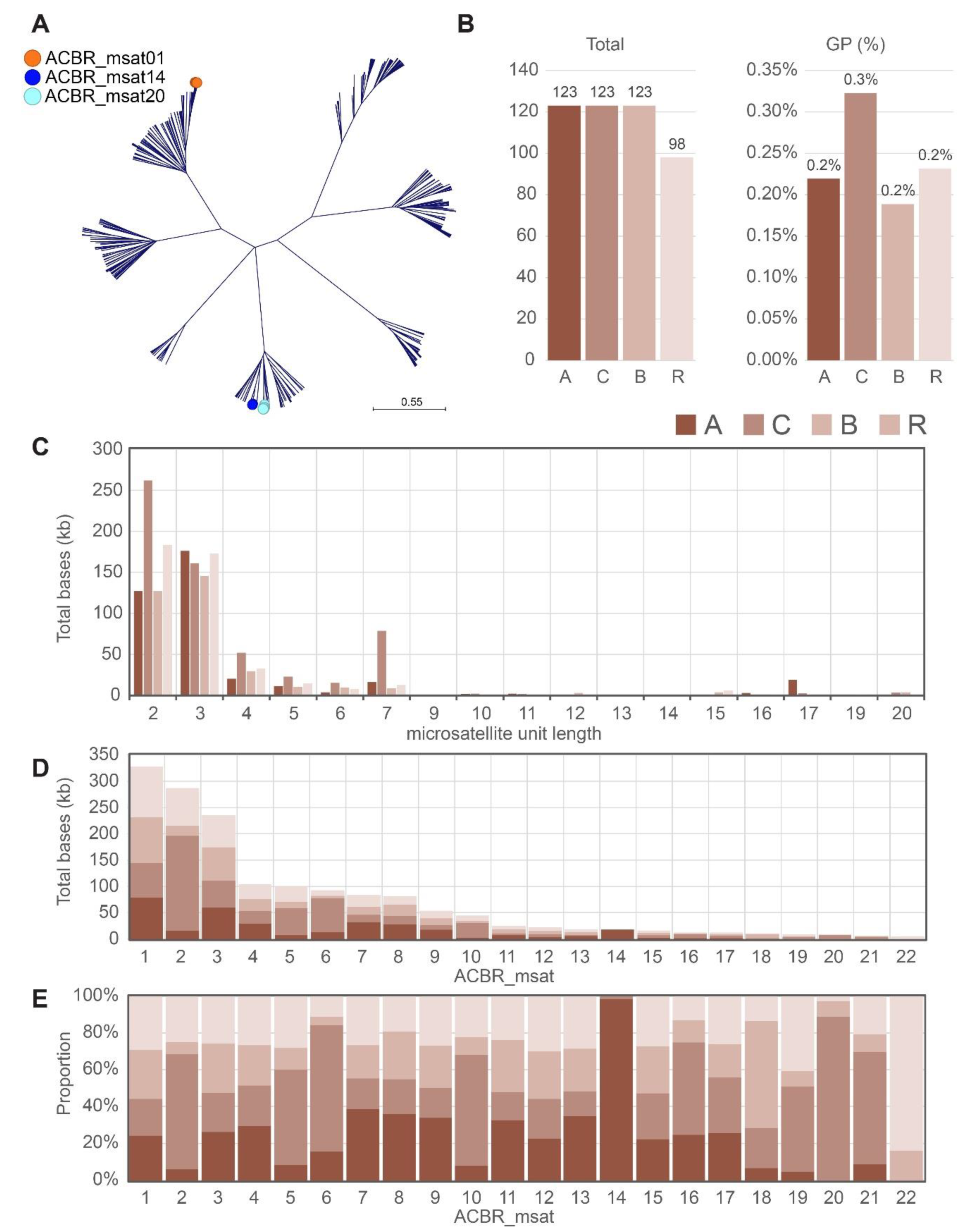
3.1. Identification of Major Microsatellites in the A, C, B, and R Genomes
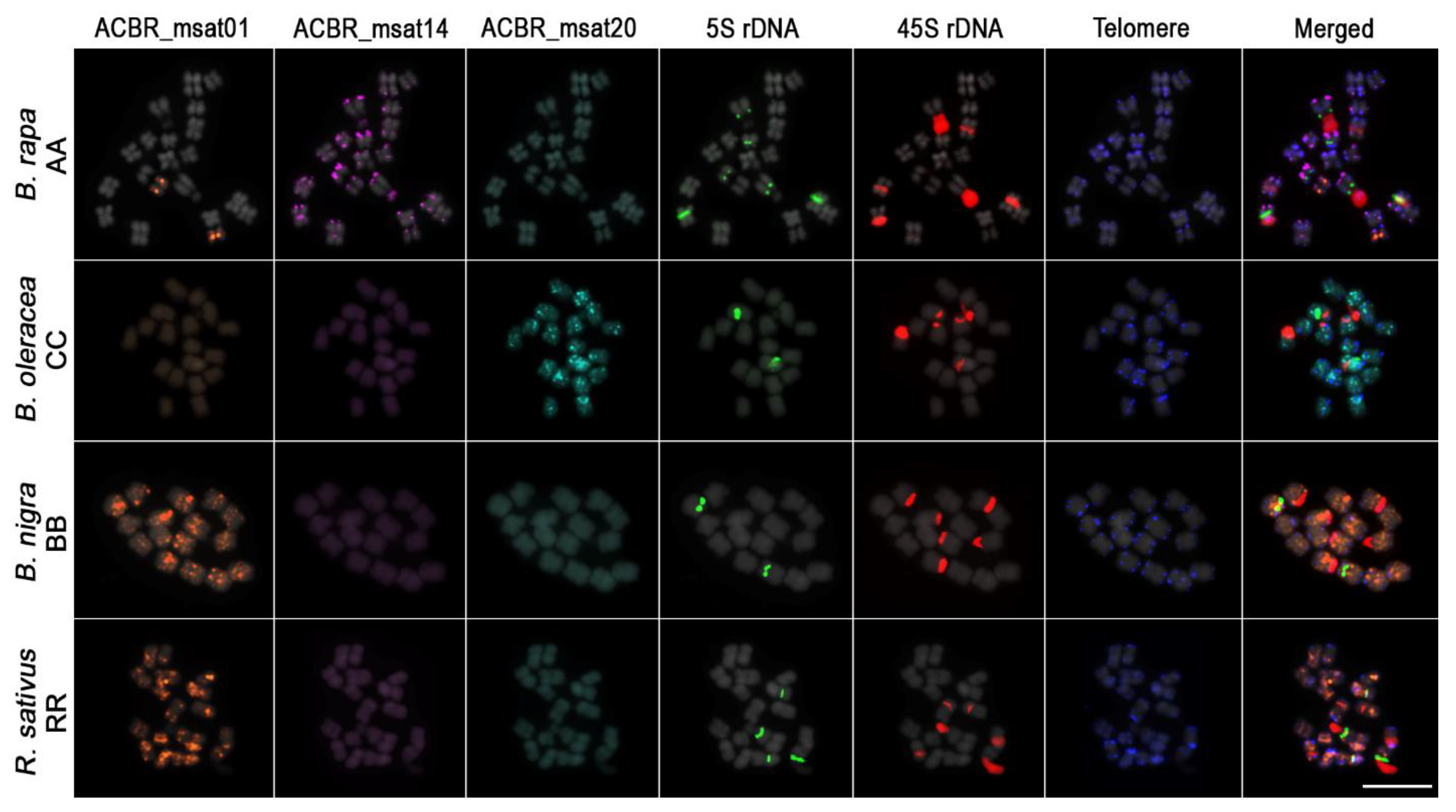
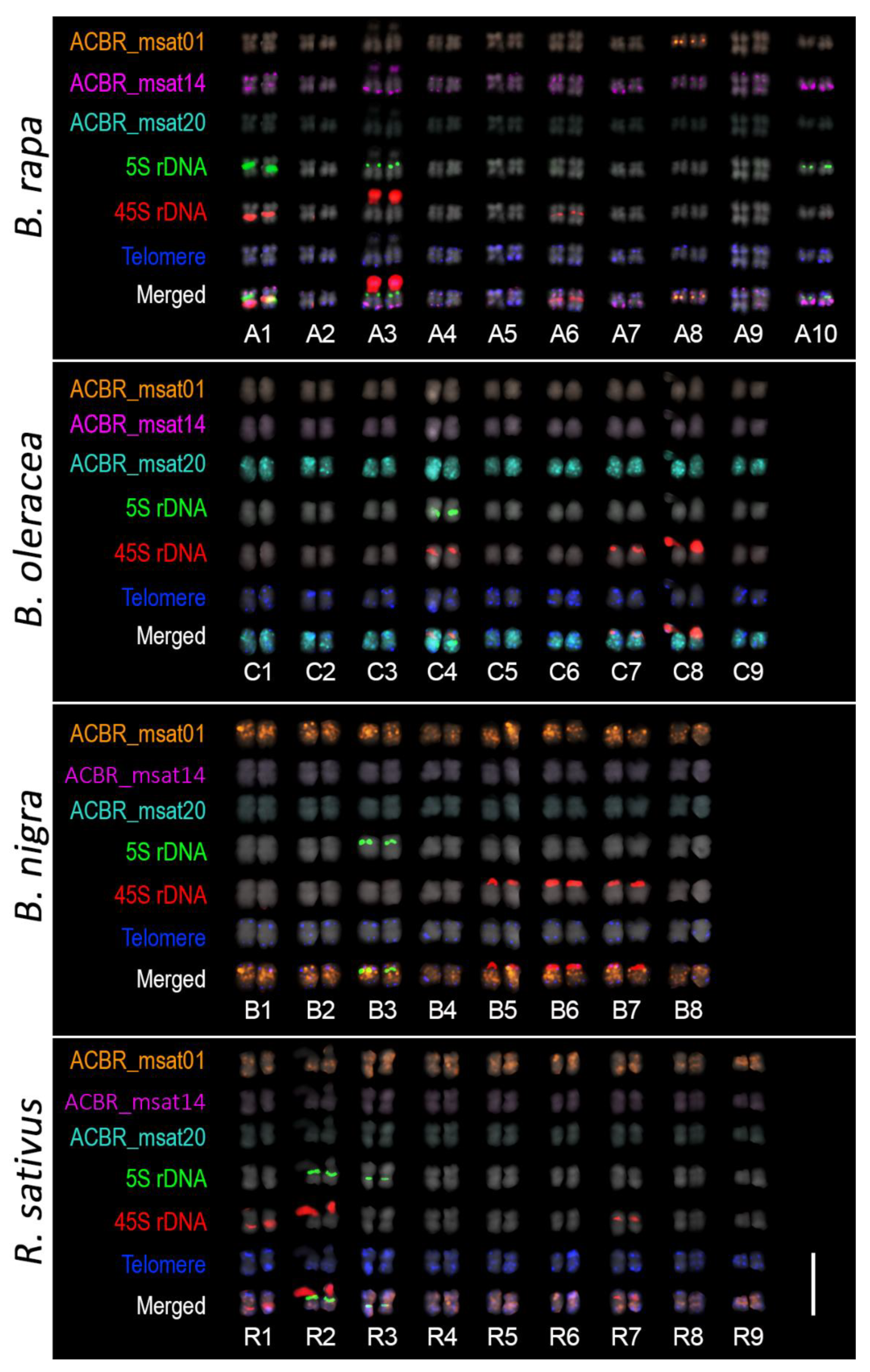
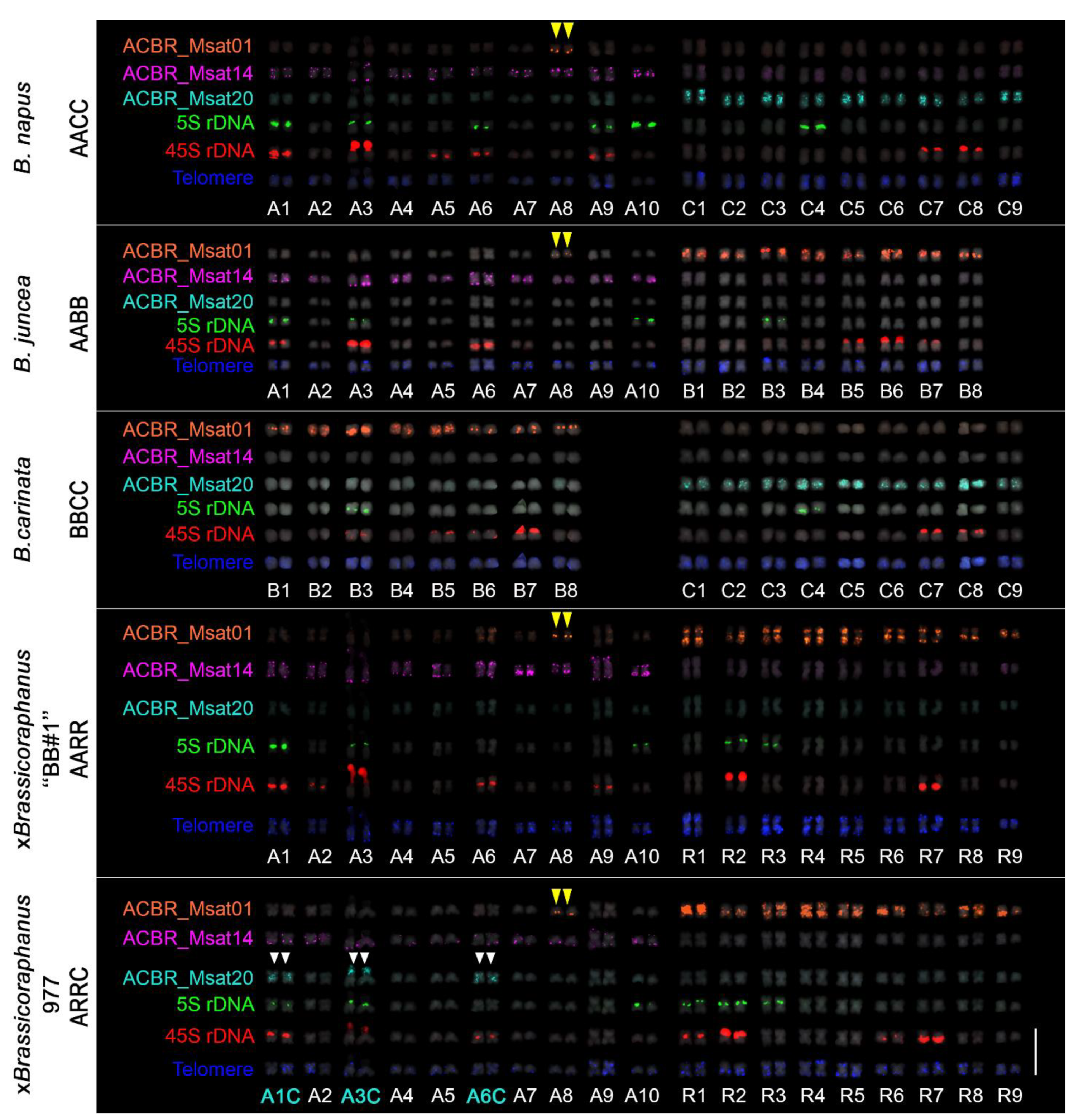
3.2. FISH Revealed Different Chromosome and Genome Distribution Patterns of the Microsatellites
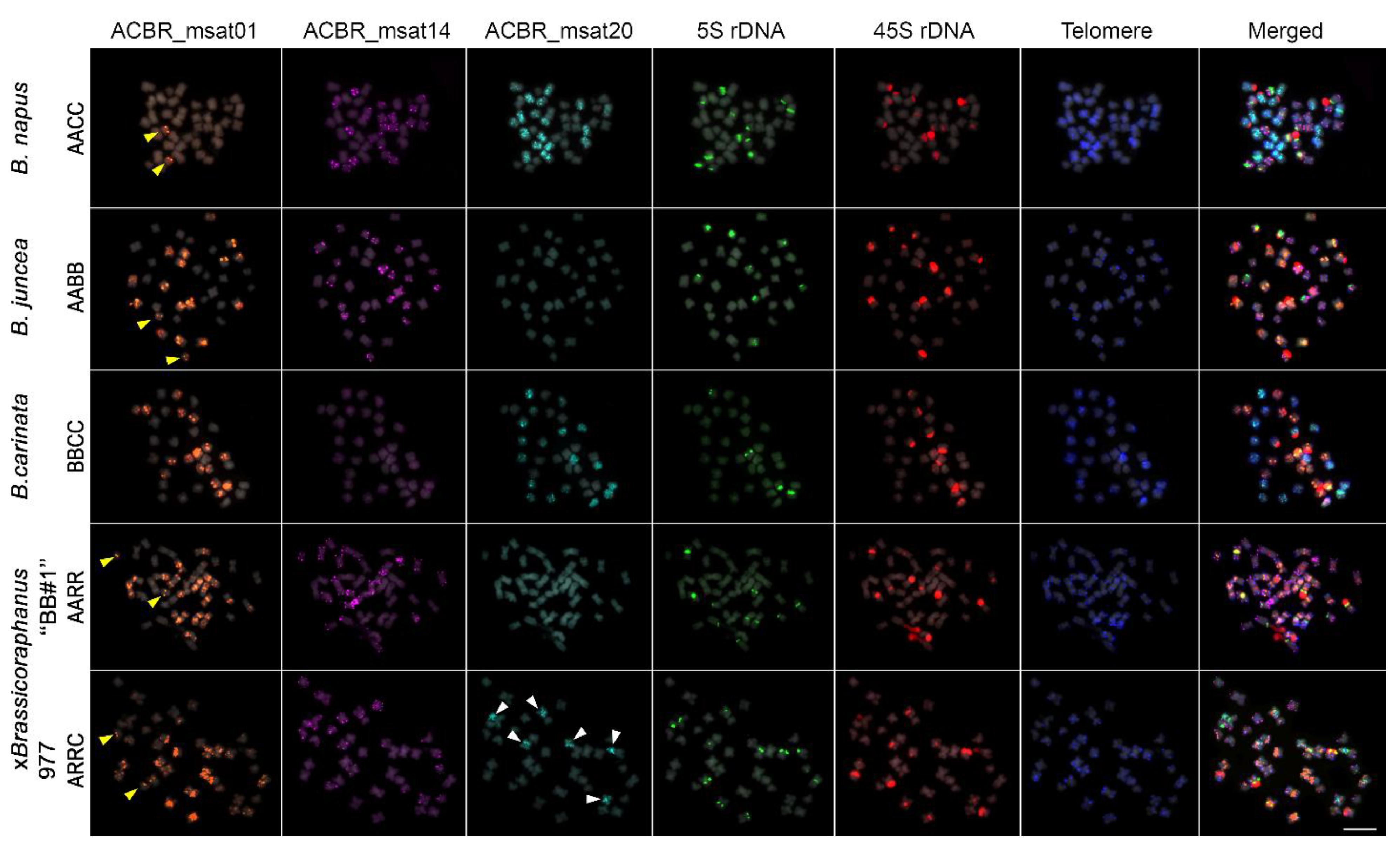
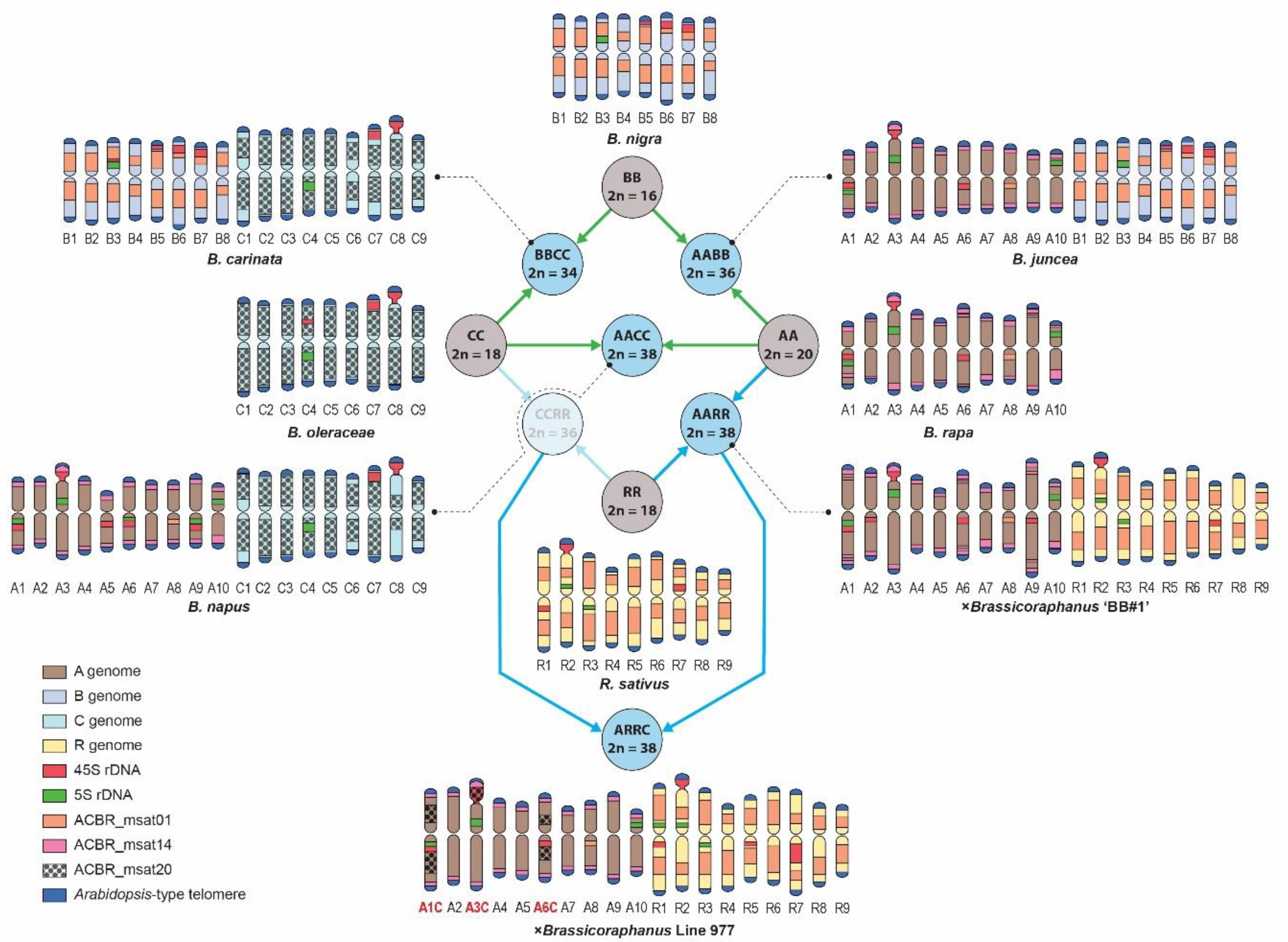
3.3. Genome-Specific Microsatellites Discriminated Subgenomes in Allotetraploids
3.4. FISH Revealed Genome Rearrangement and Chromatin Elimination in ×Brassicoraphanus 977
3.5. Ribosomal DNA and Telomeric Repeat Distribution
4. Discussion
4.1. Importance of Subgenome Discrimination in Allopolyploids
4.2. Organization of Microsatellite in the Chromosomes Determine FISH Detectability
4.3. Genome-Specific Microsatellites for the A, C, and B Brassica Genomes
4.4. Microsatellite Distribution Supports Phylogenetic Relationships of the Four Diploid Species
4.5. Association of 45S rDNA in Genome Reshuffling
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kiefer, M.; Schmickl, R.; German, D.A.; Mandáková, T.; Lysak, M.A.; Al-Shehbaz, I.A.; Franzke, A.; Mummenhoff, K.; Stamatakis, A.; Koch, M.A.J.P.; et al. BrassiBase: Introduction to a novel knowledge database on Brassicaceae evolution. J. Plant Cell Physiol. 2014, 55, e3. [Google Scholar] [CrossRef] [Green Version]
- Ebeed, H.T. Bioinformatics Studies on the Identification of New Players and Candidate Genes to Improve Brassica Response to Abiotic Stress, In The Plant Family Brassicaceae: Biology and Physiological Responses to Environmental Stresses; Hasanuzzaman, E., Ed.; Springer Nature: Singapore, 2020; pp. 483–496. [Google Scholar]
- Mandrich, L.; Caputo, E.J.N. Brassicaceae-derived anticancer agents: Towards a green approach to beat cancer. J. Nutr. 2020, 12, 868. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Balibrea, S.; Moreno, D.A.; García-Viguera, C. Glucosinolates in broccoli sprouts (Brassica oleracea var. italica) as conditioned by sulphate supply during germination. J. Food Sci. 2010, 75, C673–C677. [Google Scholar]
- Waminal, N.E.; Perumal, S.; Lee, J.; Kim, H.H.; Yang, T.-J. Repeat evolution in Brassica rapa (AA), B. oleracea (CC), and B. napus (AACC) genomes. Plant Breed. Biotechnol. 2016, 4, 107–122. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.; Zhao, M.; Ma, J.; Yu, J.; Huang, S. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef] [PubMed]
- Nagaharu, U. Genome analysis in Brassica with special reference to the experimental formation of B. napus and peculiar mode of fertilization. J. Jap. Bot. 1935, 7, 389–452. [Google Scholar]
- Khalid, M.; Ayayda, R.; Gheith, N.; Salah, Z.; Abu-Lafi, S.; Jaber, A.; Al-Rimawi, F.; Al-Mazaideh, G. Assessment of antimicrobial and anticancer activity of radish sprouts extracts. Jordan J. Biol. Sci. 2020, 13, 567–574. [Google Scholar]
- Chang, M.J.; Kim, B.; Lee, H.S.; Kim, W.K.; Cho, K.J.T.F.J. Anti-cancer effect of Korean radish (Raphanus sativus L.) leaf extract through induction of apoptosis and down-regulate Erb B2 signaling and Akt pathway in human breast MDA-MB-231 cell lines. FASEB J. 2010, 24, 720.4. [Google Scholar] [CrossRef]
- Lee, S.-S.; Choi, W.-J.; Woo, J.-G. Development of a new vegetable crop in xBrassicoraphanus by hybridization of Brassica campestris and Raphanus sativus. KHSH 2002, 43, 693–698. [Google Scholar]
- Prakash, S.; Bhat, S.; Quiros, C.; Kirti, P.; Chopra, V. Brassica and its close allies: Cytogenetics and evolution. Plant Breed. Rev. 2009, 31, 21–187. [Google Scholar]
- Bang, S.; Tsutsui, K.; Shim, S.; Kaneko, Y. Production and characterization of the novel CMS line of radish (Raphanus sativus) carrying Brassica maurorum cytoplasm. Plant Breed. 2011, 130, 410–412. [Google Scholar] [CrossRef]
- Wang, X.; Zeng, L.; Xu, L.; Chen, W.; Liu, F.; Yang, H.; Chi, P.; Ren, L.; Yan, R.; Lu, G. Clubroot resistance introgression in interspecific hybrids between Raphanus sativus and Brassica napus. OCS 2019, 4, 139–151. [Google Scholar]
- Gong, T.; Ray, Z.T.; Butcher, K.E.; Black, Z.E.; Zhao, X.; Brecht, J.K. A novel graft between Pac Choi (Brassica rapa var. chinensis) and Daikon Radish (Raphanus sativus var. longipinnatus). Agronomy 2020, 10, 1464. [Google Scholar]
- Zhan, Z.; Nwafor, C.C.; Hou, Z.; Gong, J.; Zhu, B.; Jiang, Y.; Zhou, Y.; Wu, J.; Piao, Z.; Tong, Y.J. Cytological and morphological analysis of hybrids between Brassicoraphanus, and Brassica napus for introgression of clubroot resistant trait into Brassica napus L. PLoS ONE 2017, 12, e0177470. [Google Scholar] [CrossRef]
- Lee, S.; Kim, H.; Oh, D.; Woo, J.K. Breeding a fertile intergeneric allotetraploid plant between heading Chinese cabbage and Korean radish. Korean J. Hortic. Sci. Technol. 1999, 17, 653. [Google Scholar]
- Lee, S.-S.; Lee, S.-A.; Yang, J.; Kim, J. Developing stable progenies of ×Brassicoraphanus, an intergeneric allopolyploid between Brassica rapa and Raphanus sativus, through induced mutation using microspore culture. Theor. Appl. Genet. 2011, 122, 885–891. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-S.; Son, C.Y.; Kim, J.; Park, J.E.; Yu, S.H.; Yi, G.; Huh, J.H. Properties of self-sterile but cross-fertile allopolyploids synthesized between Brassica rapa and Raphanus sativus. Korean J. Hortic. Sci. Technol. 2020, 61, 163–171. [Google Scholar] [CrossRef]
- Chen, H.; Wu, J. Characterization of fertile amphidiploid between Raphanus sativus and Brassica alboglabra and the crossability with Brassica species. Genet. Resour. Crop Evol. 2008, 55, 143–150. [Google Scholar] [CrossRef]
- Schwarzacher, T.; Leitch, A.; Bennett, M.D.; Heslop-Harrison, J. In situ localization of parental genomes in a wide hybrid. Ann. Bot. 1989, 64, 315–324. [Google Scholar] [CrossRef]
- Ramzan, F.; Younis, A.; Lim, K.-B. Application of genomic in situ hybridization in horticultural science. Int. J. Genomics 2017, 2017, 7561909. [Google Scholar] [CrossRef] [Green Version]
- Howell, E.C.; Kearsey, M.J.; Jones, G.H.; King, G.J.; Armstrong, S.J. A and C genome distinction and chromosome identification in Brassica napus by sequential fluorescence in situ hybridization and genomic in situ hybridization. Genetics 2008, 180, 1849–1857. [Google Scholar] [CrossRef] [Green Version]
- Snowdon, R.; Köhler, W.; Friedt, W.; Köhler, A. Genomic in situ hybridization in Brassica amphidiploids and interspecific hybrids. Theor. Appl. Genet. 1997, 95, 1320–1324. [Google Scholar] [CrossRef]
- Ohmido, N.; Ueda, K.; Fujii, K. Chromosome instability of allopolyploid resynthesized Brassica napus. Chromosome Sci. 2015, 18, 79–84. [Google Scholar]
- Dos Santos, K.G.B.; Becker, H.; Ecke, W.; Bellin, U. Molecular characterisation and chromosomal localisation of a telomere-like repetitive DNA sequence highly enriched in the C genome of Brassica. Cytogenet. Genome Res. 2007, 119, 147–153. [Google Scholar] [CrossRef]
- Cuadrado, A.; Cardoso, M.; Jouve, N. Physical organisation of simple sequence repeats (SSRs) in Triticeae: Structural, functional and evolutionary implications. Cytogenet. Genome Res. 2008, 120, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Gilmore, S.; Keys, W.; Morgante, M.; Rafalski, A. Cross-species amplification of soybean (Glycine max) simple sequence repeats (SSRs) within the genus and other legume genera: Implications for the transferability of SSRs in plants. Mol. Biol. Evol. 1998, 15, 1275–1287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlin, E.F.; Boles, S.B.; Seppelt, R.D.; Terracciano, S.; Shaw, A.J. The Peat Moss Sphagnum cuspidatum in Australia: Microsatellites Provide a Global Perspective. Syst. Bot. 2011, 36, 22–32. [Google Scholar] [CrossRef]
- Santos, J.; Serra, L.; Solé, E.; Pascual, M. FISH mapping of microsatellite loci from Drosophila subobscura and its comparison to related species. Chromosome Res. 2010, 18, 213–226. [Google Scholar] [CrossRef]
- Zhang, L.; Cai, X.; Wu, J.; Liu, M.; Grob, S.; Cheng, F.; Liang, J.; Cai, C.; Liu, Z.; Liu, B. Improved Brassica rapa reference genome by single-molecule sequencing and chromosome conformation capture technologies. Hortic. Res. 2018, 5, 50. [Google Scholar] [CrossRef] [Green Version]
- Paritosh, K.; Pradhan, A.K.; Pental, D. A highly contiguous genome assembly of Brassica nigra (BB) and revised nomenclature for the pseudochromosomes. BMC Genomics 2020, 21, 887. [Google Scholar] [CrossRef] [PubMed]
- Parkin, I.A.; Koh, C.; Tang, H.; Robinson, S.J.; Kagale, S.; Clarke, W.E.; Town, C.D.; Nixon, J.; Krishnakumar, V.; Bidwell, S.L. Transcriptome and methylome profiling reveals relics of genome dominance in the mesopolyploid Brassica oleracea. Genome Biol. 2014, 15, R77. [Google Scholar] [CrossRef] [Green Version]
- Shirasawa, K.; Hirakawa, H.; Fukino, N.; Kitashiba, H.; Isobe, S. Genome sequence and analysis of a Japanese radish (Raphanus sativus) cultivar named ‘Sakurajima Daikon’possessing giant root. DNA Res. 2020, 27, dsaa010. [Google Scholar] [CrossRef]
- Novák, P.; Neumann, P.; Macas, J. Global analysis of repetitive DNA from unassembled sequence reads using RepeatExplorer2. Nat. Protoc. 2020, 15, 3745–3776. [Google Scholar] [CrossRef]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Sobreira, T.J.P.; Durham, A.M.; Gruber, A. TRAP: Automated classification, quantification and annotation of tandemly repeated sequences. Bioinformatics 2006, 22, 361–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.-K.; Seol, Y.-J.; Perumal, S.; Lee, J.; Waminal, N.E.; Jayakodi, M.; Lee, S.-C.; Jin, S.; Choi, B.-S.; Yu, Y. Re-exploration of U’s Triangle Brassica species based on chloroplast genomes and 45S nrDNA sequences. Sci. Rep. 2018, 8, 7353. [Google Scholar] [CrossRef]
- Waminal, N.E.; Pellerin, R.J.; Kim, N.-S.; Jayakodi, M.; Park, J.Y.; Yang, T.-J.; Kim, H.H. Rapid and efficient FISH using pre-labeled oligomer probes. Sci. Rep. 2018, 8, 8224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waminal, N.E.; Kim, H.H. Dual-color FISH karyotype and rDNA distribution analyses on four Cucurbitaceae species. Hortic. Environ. Biotechnol. 2012, 53, 49–56. [Google Scholar] [CrossRef]
- Vrána, J.; Šimková, H.; Kubaláková, M.; Číhalíková, J.; Doležel, J. Flow cytometric chromosome sorting in plants: The next generation. Methods 2012, 57, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Lim, K.B.; Yang, T.J.; Hwang, Y.J.; Kim, J.S.; Park, J.Y.; Kwon, S.J.; Kim, J.; Choi, B.S.; Lim, M.H.; Jin, M.; et al. Characterization of the centromere and peri-centromere retrotransposons in Brassica rapa and their distribution in related Brassica species. Plant J. 2007, 49, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Kulak, S.; Hasterok, R.; Maluszynska, J. Karyotyping of Brassica amphidiploids using 5S and 25S rDNA as chromosome markers. Hereditas 2002, 136, 144–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belandres, H.R.; Waminal, N.E.; Hwang, Y.-J.; Park, B.-S.; Lee, S.-S.; Huh, J.H.; Kim, H.H. FISH karyotype and GISH meiotic pairing analyses of a stable intergeneric hybrid xBrassicoraphanus line BB# 5. Hortic. Sci. Technol. 2015, 33, 83–92. [Google Scholar]
- Mehrotra, S.; Goyal, V. Repetitive sequences in plant nuclear DNA: Types, distribution, evolution and function. GPB 2014, 12, 164–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, M.L.C.; Santini, L.; Diniz, A.L.; de Freitas Munhoz, C. Microsatellite markers: What they mean and why they are so useful. Genet. Mol. Biol. 2016, 39, 312–328. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ruano, F.J.; López-León, M.D.; Cabrero, J.; Camacho, J.P.M. High-throughput analysis of the satellitome illuminates satellite DNA evolution. Sci. Rep. 2016, 6, 28333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, Y.-J.; Yu, H.-J.; Mun, J.-H.; Bok, K.; Park, B.-S.; Lim, K.-B. Centromere repeat DNA originated from Brassica rapa is detected in the centromere region of Raphanus sativus chromosomes. Hortic. Sci. Technol. 2012, 30, 751–756. [Google Scholar] [CrossRef]
- Perumal, S.; Waminal, N.E.; Lee, J.; Lee, J.; Choi, B.-S.; Kim, H.H.; Grandbastien, M.-A.; Yang, T.-J. Elucidating the major hidden genomic components of the A, C, and AC genomes and their influence on Brassica evolution. Sci. Rep. 2017, 7, 17986. [Google Scholar] [CrossRef]
- Xiong, Z.; Pires, J.C. Karyotype and identification of all homoeologous chromosomes of allopolyploid Brassica napus and its diploid progenitors. Genetics 2011, 187, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Chester, M.; Gallagher, J.P.; Symonds, V.V.; da Silva, A.V.C.; Mavrodiev, E.V.; Leitch, A.R.; Soltis, P.S.; Soltis, D.E. Extensive chromosomal variation in a recently formed natural allopolyploid species, Tragopogon miscellus (Asteraceae). PNAS 2012, 109, 1176–1181. [Google Scholar] [CrossRef] [Green Version]
- Belandres, H.R.; Zhou, H.C.; Waminal, N.E.; Lee, S.-S.; Huh, J.H.; Kim, H.H. Cytogenetic analyses revealed different genome rearrangement footprints in four ×Brassicoraphanus lines with different fertility rates. Plant Breed. Biotechnol. 2019, 7, 95–105. [Google Scholar] [CrossRef]
- Han, Y.; Zhang, T.; Thammapichai, P.; Weng, Y.; Jiang, J. Chromosome-specific painting in Cucumis species using bulked oligonucleotides. Genetics 2015, 200, 771–779. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J. Fluorescence in situ hybridization in plants: Recent developments and future applications. Chromosome Res. 2019, 27, 153–165. [Google Scholar] [CrossRef]
- Cuadrado, A.; Schwarzacher, T.; Jouve, N. Identification of different chromatin classes in wheat using in situ hybridization with simple sequence repeat oligonucleotides. Theor. Appl. Genet. 2000, 101, 711–717. [Google Scholar] [CrossRef]
- Zattera, M.L.; Gazolla, C.B.; de Araújo Soares, A.; Gazoni, T.; Pollet, N.; Recco-Pimentel, S.M.; Bruschi, D.P. Evolutionary dynamics of the repetitive DNA in the karyotypes of Pipa carvalhoi and Xenopus tropicalis (Anura, Pipidae). Front. Genet. 2020, 11, 637. [Google Scholar] [CrossRef] [PubMed]
- Utsunomia, R.; Melo, S.; Scacchetti, P.C.; Oliveira, C.; de Andrade Machado, M.; Pieczarka, J.C.; Nagamachi, C.Y.; Foresti, F. Particular chromosomal distribution of microsatellites in five species of the genus Gymnotus (Teleostei, Gymnotiformes). Zebrafish 2018, 15, 398–403. [Google Scholar] [CrossRef]
- Megyeri, M.; Mikó, P.; Farkas, A.; Molnár-Láng, M.; Molnár, I. Cytomolecular discrimination of the A chromosomes of Triticum monococcum and the A chromosomes of Triticum aestivum using microsatellite DNA repeats. J. Appl. Genet. 2017, 58, 67–70. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, H.; Wu, J.; Matsumoto, T. Characterization of chromosomal ends on the basis of chromosome-specific telomere variants and subtelomeric repeats in rice (Oryza sativa L.). In Subtelomeres; Springer: Berlin/Heidelberg, Germany, 2014; pp. 187–194. [Google Scholar]
- Mizuno, H.; Matsumoto, T.; Wu, J. Composition and structure of rice centromeres and telomeres. In Rice Genomics, Genetics and Breeding; Springer: Berlin/Heidelberg, Germany, 2018; pp. 37–52. [Google Scholar]
- Waminal, N.E.; Pellerin, R.J.; Kang, S.-H.; Kim, H.H. Chromosomal mapping of tandem repeats revealed massive chromosomal rearrangements and insights into Senna tora dysploidy. Front. Plant Sci. 2021, 12, 154. [Google Scholar] [CrossRef]
- Vershinin, A.V.; Evtushenko, E.V. What is the Specificity of Plant Subtelomeres? In Subtelomeres; Louis, E.J., Becker, M.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 195–209. [Google Scholar]
- Schelfhout, C.; Snowdon, R.; Cowling, W.; Wroth, J.M.; Genetics, A. A PCR based B-genome-specific marker in Brassica species. Theor. Appl. Genet. 2004, 109, 917–921. [Google Scholar] [CrossRef] [PubMed]
- Schelfhout, C.; Snowdon, R.; Cowling, W.; Wroth, J.M. Tracing B-genome chromatin in Brassica napus × B. juncea interspecific progeny. Genome 2006, 49, 1490–1497. [Google Scholar] [CrossRef]
- He, Q.; Cai, Z.; Hu, T.; Liu, H.; Bao, C.; Mao, W.; Jin, W. Repetitive sequence analysis and karyotyping reveals centromere-associated DNA sequences in radish (Raphanus sativus L.). BMC Plant Biol. 2015, 15, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flynn, J.M.; Long, M.; Wing, R.A.; Clark, A.G. Evolutionary dynamics of abundant 7-bp satellites in the genome of Drosophila virilis. Mol. Biol. 2020, 37, 1362–1375. [Google Scholar] [CrossRef]
- Havlová, K.; Dvořáčková, M.; Peiro, R.; Abia, D.; Mozgová, I.; Vansáčová, L.; Gutierrez, C.; Fajkus, J. Variation of 45S rDNA intergenic spacers in Arabidopsis thaliana. Plant Mol. Biol. 2016, 92, 457–471. [Google Scholar] [CrossRef]
- Garcia, S.; Kovařík, A.J.H. Dancing together and separate again: Gymnosperms exhibit frequent changes of fundamental 5S and 35S rRNA gene (rDNA) organisation. Heredity 2013, 111, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ta, T.D.; Waminal, N.E.; Nguyen, T.H.; Pellerin, R.J.; Kim, H.H. Comparative FISH analysis of Senna tora tandem repeats revealed insights into the chromosome dynamics in Senna. Genes Genom. 2021, 43, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.; Fonsêca, A.; dos Santos, K.G.B.; Mosiolek, M.; Pedrosa-Harand, A.J.G. Contrasting evolution of a satellite DNA and its ancestral IGS rDNA in Phaseolus (Fabaceae). Genome 2012, 55, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Falquet, J.; Creusot, F.; Dron, M. Molecular analysis of Phaseolus vulgaris rDNA unit and characterization of a satellite DNA homologous to IGS subrepeats. Plant Physiol. 1997, 35, 611–622. [Google Scholar]
- Dos Santos, J.M.; Diniz, D.; Rodrigues, T.A.S.; Cioffi, M.D.B.; Waldschmidt, A.M. Heterochromatin distribution and chromosomal mapping of microsatellite repeats in the genome of Frieseomelitta stingless bees (Hymenoptera: Apidae: Meliponini). Fla. Entomol. 2018, 101, 33–39. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Cultivar | Genome | Genome Size (Mbp) | Accession Number | Reads | Coverage (×) |
|---|---|---|---|---|---|---|
| B. rapa | Chiifu-401-42 | AA | 353 | GCA_000309985.3 | 199,662 | 0.06 |
| B. oleracea | TO1000 | CC | 522 | GCA_000695525.1 | 200,880 | 0.04 |
| B. nigra | inbred line YZ12151 | BB | 468 | GCA_001682895.1 | 201,798 | 0.04 |
| R. sativus | XYB36-2 | RR | 414 | GCA_002197605.1 | 201,210 | 0.05 |
| Species | 2n | Genome Type | Source | Accession Number |
|---|---|---|---|---|
| B. rapa | 20 | AA | NAASa | IT 032730 |
| B. oleracea | 18 | CC | NPGSb | PI 24501510GI |
| B. nigra | 16 | BB | NPGS | PI 649154 |
| R. sativus | 18 | RR | danong.co.kr (accessed on 31 April 2019) | N/A |
| B. napus | 38 | AACC | NPGS | TI 031006 |
| B. juncea | 36 | AABB | NPGS | PI 633077 |
| B. carinata | 34 | BBCC | NPGS | Ames 2779 |
| ×Brassicoraphanus ‘BB#1′ | 38 | AARR | BioBreeding Institute (Ansung, Korea) | PRJNA353741 |
| ×Brassicoraphanus 977 | 38 | ARRC | BioBreeding Institute | N/A |
| Name | Sequence | Length | Total Bases | ||||
|---|---|---|---|---|---|---|---|
| A | C | B | R | Grand Total | |||
| ACBR_msat01 | AG | 2 | 80,004 | 65,645 | 87,523 | 95,545 | 328,717 |
| ACBR_msat02 | TA | 2 | 17,429 | 180,550 | 18,852 | 71,783 | 288,614 |
| ACBR_msat03 | AAG | 3 | 62,101 | 50,474 | 63,611 | 61,099 | 237,285 |
| ACBR_msat04 | TGA | 3 | 31,243 | 23,210 | 23,246 | 28,005 | 105,704 |
| ACBR_msat05 | TAA | 3 | 8541 | 51,851 | 11,968 | 28,179 | 100,539 |
| ACBR_msat06 | TTTAGGG | 7 | 14,629 | 64,754 | 4438 | 10,471 | 94,292 |
| ACBR_msat07 | TTG | 3 | 33,032 | 14,323 | 15,292 | 22,713 | 85,360 |
| ACBR_msat08 | TG | 2 | 29,815 | 15,745 | 21,198 | 16,033 | 82,791 |
| ACBR_msat09 | GAG | 3 | 18,862 | 8880 | 12,870 | 14,823 | 55,435 |
| ACBR_msat10 | AAAT | 4 | 3720 | 27,736 | 4390 | 10,228 | 46,074 |
| ACBR_msat11 | ACC | 3 | 8494 | 4084 | 7375 | 6254 | 26,207 |
| ACBR_msat12 | CTTT | 4 | 5361 | 5163 | 6096 | 7137 | 23,757 |
| ACBR_msat13 | TGC | 3 | 6967 | 2710 | 4611 | 5726 | 20,014 |
| ACBR_msat14 | TTTAGGGTTAGGTAGGG | 17 | 18,903 | 297 | 0 | 0 | 19,200 |
| ACBR_msat15 | AAAC | 4 | 3933 | 4403 | 4519 | 4813 | 17,668 |
| ACBR_msat16 | TTCGG | 5 | 3505 | 7088 | 1709 | 1843 | 14,145 |
| ACBR_msat17 | ACT | 3 | 3484 | 4086 | 2441 | 3552 | 13,563 |
| ACBR_msat18 | ATAG | 4 | 850 | 2701 | 7258 | 1684 | 12,493 |
| ACBR_msat19 | ATTTT | 5 | 509 | 4749 | 866 | 4191 | 10,315 |
| ACBR_msat20 | TTTCGGG | 7 | 0 | 8845 | 835 | 270 | 9950 |
| ACBR_msat21 | AATT | 4 | 722 | 4925 | 792 | 1661 | 8100 |
| ACBR_msat22 | TGAACAGTGTTTCGA | 15 | 0 | 0 | 973 | 5126 | 6099 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campomayor, N.B.; Waminal, N.E.; Kang, B.Y.; Nguyen, T.H.; Lee, S.-S.; Huh, J.H.; Kim, H.H. Subgenome Discrimination in Brassica and Raphanus Allopolyploids Using Microsatellites. Cells 2021, 10, 2358. https://doi.org/10.3390/cells10092358
Campomayor NB, Waminal NE, Kang BY, Nguyen TH, Lee S-S, Huh JH, Kim HH. Subgenome Discrimination in Brassica and Raphanus Allopolyploids Using Microsatellites. Cells. 2021; 10(9):2358. https://doi.org/10.3390/cells10092358
Chicago/Turabian StyleCampomayor, Nicole Bon, Nomar Espinosa Waminal, Byung Yong Kang, Thi Hong Nguyen, Soo-Seong Lee, Jin Hoe Huh, and Hyun Hee Kim. 2021. "Subgenome Discrimination in Brassica and Raphanus Allopolyploids Using Microsatellites" Cells 10, no. 9: 2358. https://doi.org/10.3390/cells10092358