
Measurement of Chromosomal Arms and FISH Reveal Complex Genome Architecture and Standardized Karyotype of Model Fish, Genus Carassius



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fish Sampling and Origin
2.2. Chromosome Analysis
2.3. Measurements of Chromosomal Arm Length
2.4. Standardized Carassius Karyotype
2.5. Preparation of 5S and 28S Ribosomal Probes
2.6. Fluorescent In Situ Hybridization
3. Results
3.1. Karyotype Analysis
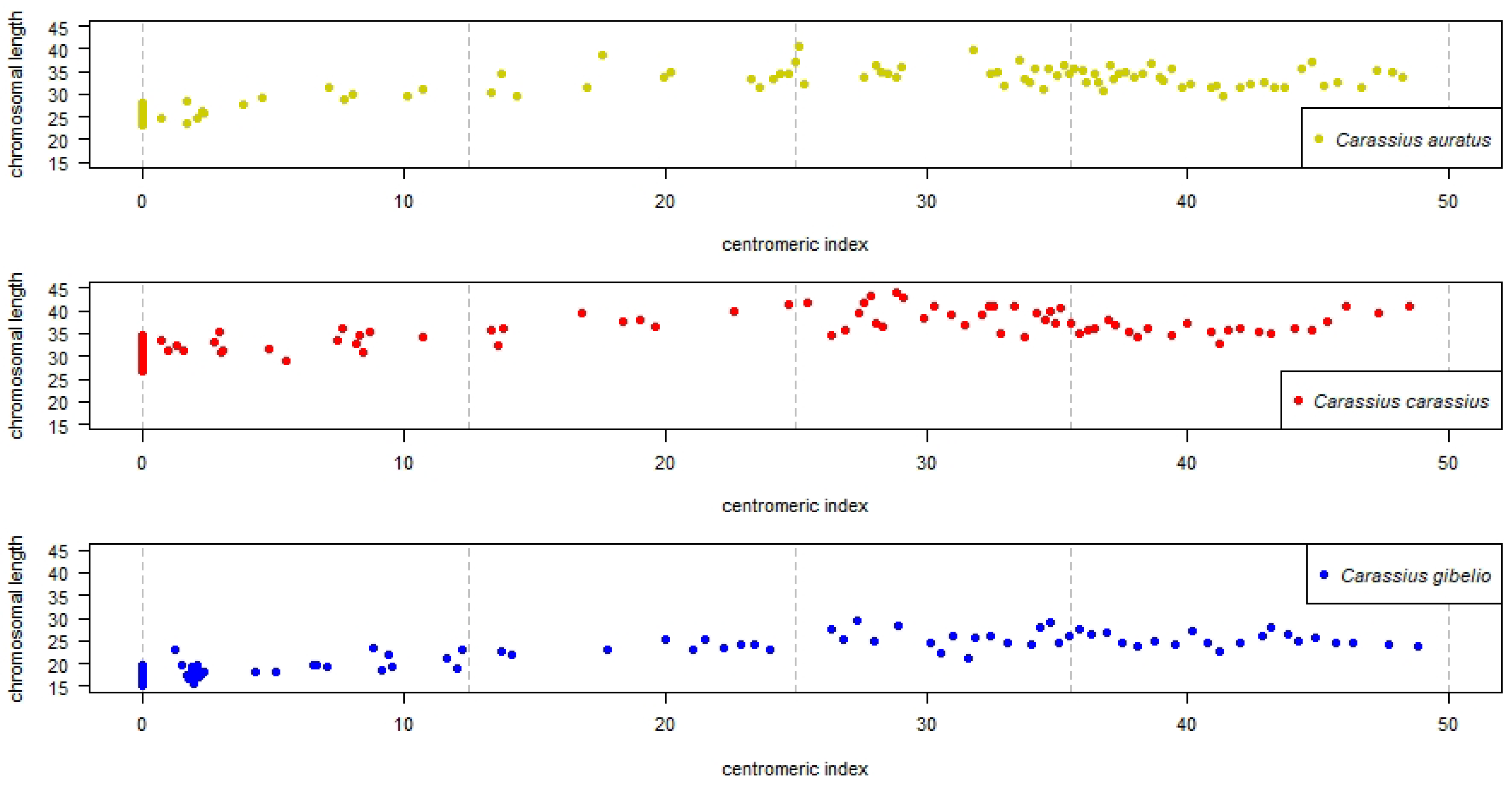
3.2. Interspecies Variability Based on Chromosomal Length
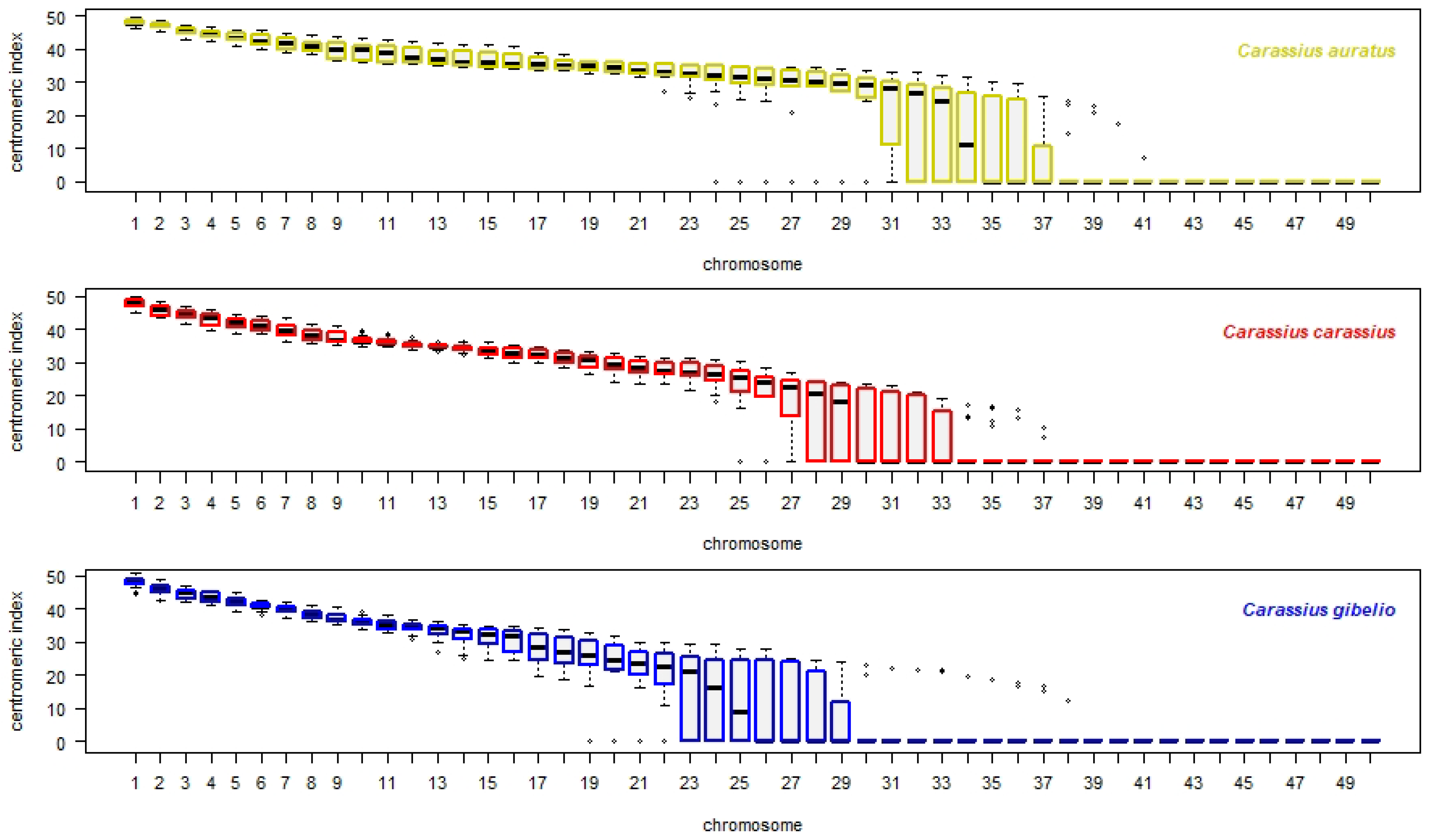
3.3. Interspecies Variability Based on Centromeric Index Linked to Individual Chromosomes of the Whole Chromosomal Complement, and Standardized Carassius Karyotype
- Each orthologous chromosome of C. carassius and C. gibelio shared i values within – range and therefore we consider that karyotypes of C. carassius and C. gibelio to be more similar based on i value.
- Chromosomes 12–30 of C. auratus were represented by different i values within – range those of C. carassius and C. gibelio. Therefore, we consider the karyotypes of C. auratus to be most variable for this parameter.
- Chromosomes 8, 11 and 31 of C. auratus were represented by different i values within – range those of C. gibelio but C. carassius shared i values in these chromosomes with both C. auratus and C. gibelio.
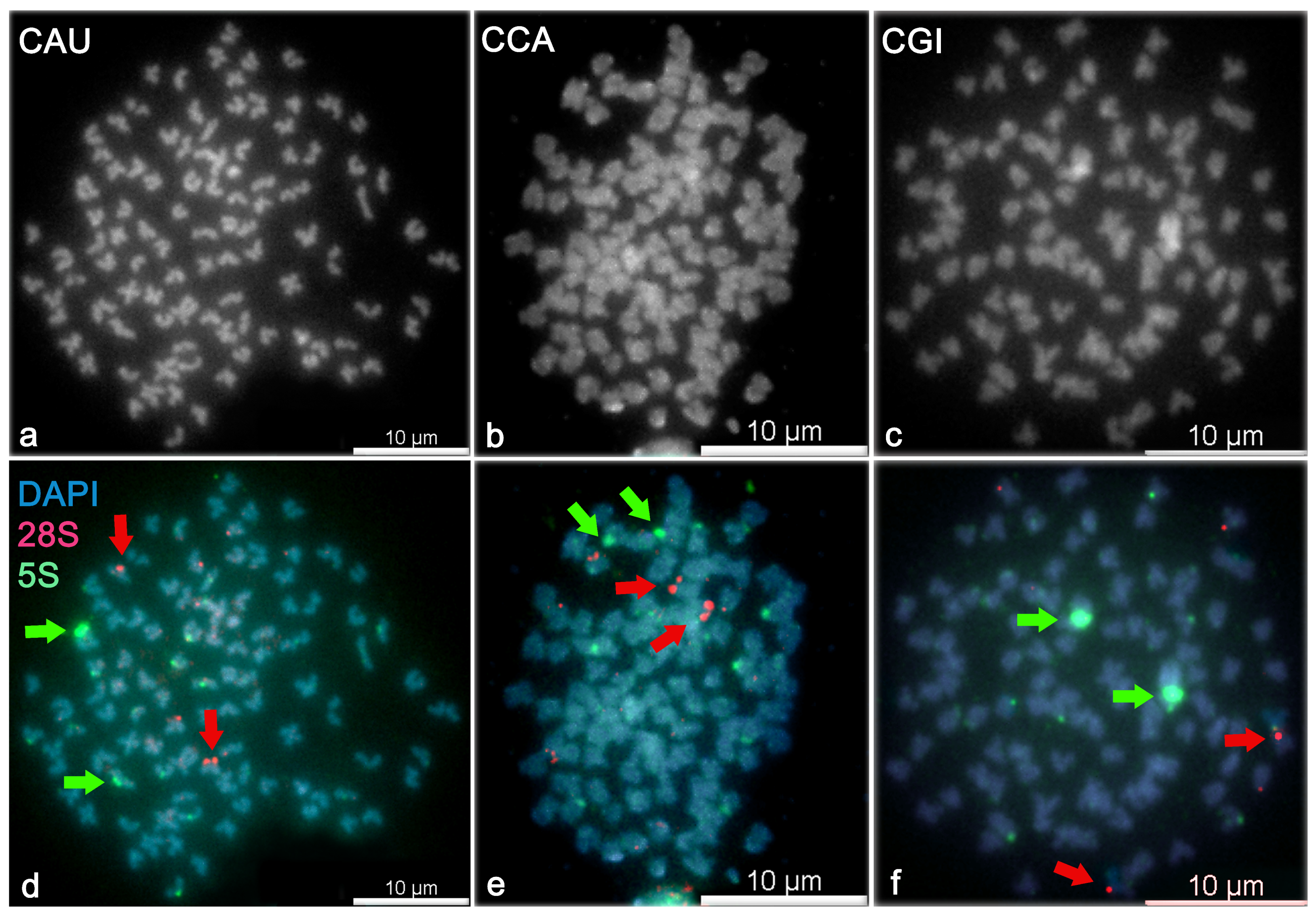
3.4. Fluorescent In Situ Hybridization with 5S and 28S Ribosomal Probes
- Two more intensive (strong) 5S rDNA signals at the p arm of chromosomes (chr28, i value ) and eight weak signals in C. auratus.
- Two strong 5S rDNA signals at the p arm of chromosomes (chr19, i value ) and six weak signals in C. carassius.
- Two strong 5S rDNA signals at the p arm of chromosomes (chr17, i value ) and eight weak signals in C. gibelio.
- two strong 28S rDNA signals at the p arm of two chromosomes (chr16, i value ) and two weak signals in C. auratus.
- two strong 28S rDNA signals at the p arm of two chromosomes (chr13, i value ) and four weak signals in C. carassius.
- two strong 28S rDNA signals at the pericentromeric region of two T chromosomes (in the range of chr26–50, i value ) and two weak 28S rDNA signals in C. gibelio.
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Delaunay, L. Comparative karyological study of species Muscari Mill. and Bellevalia Lapeyr. Bull. Tiflis Bot. Gard. 1922, 2, 1–32. (In Russian) [Google Scholar]
- Levan, A.; Fredga, K.; Sandberg, A.A. Nomenclature for centromeric position on chromosomes. Hereditas 1964, 52, 201–220. [Google Scholar] [CrossRef]
- White, M.J.D. Animal Cytology and Evolution, 3rd ed.; Cambridge University Press: Cambridge, UK, 1973; p. 468. [Google Scholar]
- White, M.J.D. The Chromosomes, 6th ed.; Chapman Hall: London, UK, 1973; p. 188. [Google Scholar]
- Kretschmer, R.; Gunski, R.J.; Garnero, A.D.V.; de Freitas, T.R.O.; Toma, G.A.; Cioffi, M.D.B.; de Oliveira, E.H.C.; O’Connor, R.E.; Griffin, D.K. Chromosomal Analysis in Crotophaga ani (Aves, Cuculiformes) Reveals Extensive Genomic Reorganization and an Unusual Z-Autosome Robertsonian Translocation. Cells 2020, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, R.E.; Kiazim, L.G.; Rathje, C.C.; Jennings, R.L.; Griffin, D.K. Rapid Multi-Hybridisation FISH Screening for Balanced Porcine Reciprocal Translocations Suggests a Much Higher Abnormality Rate Than Previously Appreciated. Cells 2021, 10, 250. [Google Scholar] [CrossRef] [PubMed]
- Cauret, C.M.; Gansauge, M.T.; Tupper, A.S.; Furman, B.L.; Knytl, M.; Song, X.Y.; Greenbaum, E.; Meyer, M.; Evans, B.J.; Wilson, M. Developmental Systems Drift and the Drivers of Sex Chromosome Evolution. Mol. Biol. Evol. 2020, 37, 799–810. [Google Scholar] [CrossRef]
- Augstenová, B.; Pensabene, E.; Kratochvíl, L.; Rovatsos, M. Cytogenetic Evidence for Sex Chromosomes and Karyotype Evolution in Anguimorphan Lizards. Cells 2021, 10, 1612. [Google Scholar] [CrossRef] [PubMed]
- Knytl, M.; Tlapakova, T.; Vankova, T.; Krylov, V. Silurana Chromosomal Evolution: A New Piece to the Puzzle. Cytogenet. Genome Res. 2018, 156, 223–228. [Google Scholar] [CrossRef]
- Gillberg, C. Chromosomal disorders and autism. J. Autism Dev. Disord. 1998, 28, 415–425. [Google Scholar] [CrossRef]
- Erkilic, N.; Gatinois, V.; Torriano, S.; Bouret, P.; Sanjurjo-Soriano, C.; Luca, V.D.; Damodar, K.; Cereso, N.; Puechberty, J.; Sanchez-Alcudia, R.; et al. A Novel Chromosomal Translocation Identified due to Complex Genetic Instability in iPSC Generated for Choroideremia. Cells 2019, 8, 1068. [Google Scholar] [CrossRef]
- Sember, A.; Bertollo, L.A.C.; Ráb, P.; Yano, C.F.; Hatanaka, T.; de Oliveira, E.A.; Cioffi, M.D.B. Sex Chromosome Evolution and Genomic Divergence in the Fish Hoplias malabaricus (Characiformes, Erythrinidae). Front. Genet. 2018, 9, 71. [Google Scholar] [CrossRef]
- Miura, I.; Shams, F.; Lin, S.M.; Cioffi, M.D.B.; Liehr, T.; Al-Rikabi, A.; Kuwana, C.; Srikulnath, K.; Higaki, Y.; Ezaz, T. Evolution of a Multiple Sex-Chromosome System by Three-Sequential Translocations among Potential Sex-Chromosomes in the Taiwanese Frog Odorrana swinhoana. Cells 2021, 10, 661. [Google Scholar] [CrossRef]
- Seroussi, E.; Knytl, M.; Pitel, F.; Elleder, D.; Krylov, V.; Leroux, S.; Morisson, M.; Yosefi, S.; Miyara, S.; Ganesan, S.; et al. Avian Expression Patterns and Genomic Mapping Implicate Leptin in Digestion and TNF in Immunity, Suggesting That Their Interacting Adipokine Role Has Been Acquired Only in Mammals. Int. J. Mol. Sci. 2019, 20, 4489. [Google Scholar] [CrossRef] [PubMed]
- Tymowska, J. Polyploidy and cytogenetic variation in frogs of the genus Xenopus. In Amphibian Cytogenetics and Evolution; Green, D.M., Sessions, S.K., Eds.; Academic Press: San Diego, CA, USA, 1991; pp. 259–297. [Google Scholar]
- Knytl, M.; Kalous, L.; Symonová, R.; Rylková, K.; Ráb, P. Chromosome studies of European cyprinid fishes: Cross-species painting reveals natural allotetraploid origin of a Carassius female with 206 chromosomes. Cytogenet. Genome Res. 2013, 139, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Kobayasi, H.; Kawashima, Y.; Takeuchi, N. Comparative Chromosome Studies in the Genus Carassius, Especially with a Finding of Polyploidy in the Ginbuna (C. auratus langsdorfii). Jpn. J. Ichthyol. 1970, 17, 153–160. [Google Scholar] [CrossRef]
- Ojima, Y.; Takai, A. Further cytogenetical studies on the origin of the gold-fish. Proc. Jpn. Academy. Ser. B Phys. Biol. Sci. 1979, 55, 346–350. [Google Scholar] [CrossRef]
- Yang, L.; Sado, T.; Vincent Hirt, M.; Pasco-Viel, E.; Arunachalam, M.; Li, J.; Wang, X.; Freyhof, J.; Saitoh, K.; Simons, A.M.; et al. Phylogeny and polyploidy: Resolving the classification of cyprinine fishes (Teleostei: Cypriniformes). Mol. Phylogenet. Evol. 2015, 85, 97–116. [Google Scholar] [CrossRef] [PubMed]
- Lusk, S.; Hanel, L.; Lojkásek, B.; Lusková, V.; Muška, M. The Red List of Lampreys and Fishes of the Czech Republic. In Red List of Threatened Species of the Czech Republic, Vertebrates; Němec, M., Chobot, K., Eds.; Příroda: Prague, Czech Republic, 2017; Chapter 34; pp. 51–82. [Google Scholar]
- Sayer, C.D.; Copp, G.H.; Emson, D.; Godard, M.J.; Ziȩba, G.; Wesley, K.J. Towards the conservation of crucian carp Carassius carassius: Understanding the extent and causes of decline within part of its native English range. J. Fish Biol. 2011, 79, 1608–1624. [Google Scholar] [CrossRef] [PubMed]
- Ohno, S.; Muramoto, J.; Christian, L.; Atkin, N.B. Diploid-tetraploid relationship among old-world members of the fish family Cyprinidae. Chromosoma 1967, 23, 1–9. [Google Scholar] [CrossRef]
- Balon, E.K. About the oldest domesticates among fishes. J. Fish Biol. 2004, 65, 1–27. [Google Scholar] [CrossRef]
- Lusková, V.; Lusk, S.; Halačka, K.; Vetešník, L. Carassius auratus gibelio—The most successful invasive fish in waters of the Czech Republic. Russ. J. Biol. Invasions 2010, 1, 176–180. [Google Scholar] [CrossRef]
- Tarkan, A.S.; Güler Ekmekçi, F.; Vilizzi, L.; Copp, G.H. Risk screening of non-native freshwater fishes at the frontier between Asia and Europe: First application in Turkey of the fish invasiveness screening kit. J. Appl. Ichthyol. 2014, 30, 392–398. [Google Scholar] [CrossRef]
- Card, J.T.; Hasler, C.T.; Ruppert, J.L.; Donadt, C.; Poesch, M.S. A Three-pass electrofishing removal strategy is not effective for eradication of prussian Carp in a North American stream network. J. Fish Wildl. Manag. 2020, 11, 485–493. [Google Scholar] [CrossRef]
- Haynes, G.D.; Gongora, J.; Gilligan, D.M.; Grewe, P.; Moran, C.; Nicholas, F.W. Cryptic hybridization and introgression between invasive Cyprinid species Cyprinus carpio and Carassius auratus in Australia: Implications for invasive species management. Anim. Conserv. 2012, 15, 83–94. [Google Scholar] [CrossRef]
- Kalous, L.; Knytl, M. Karyotype diversity of the offspring resulting from reproduction experiment between diploid male and triploid female of silver Prussian carp, Carassius gibelio (Cyprinidae, Actinopterygii). Folia Zool. 2011, 60, 115–121. [Google Scholar] [CrossRef]
- Xiao, J.; Zou, T.; Chen, Y.; Chen, L.; Liu, S.; Tao, M.; Zhang, C.; Zhao, R.; Zhou, Y.; Long, Y.; et al. Coexistence of diploid, triploid and tetraploid crucian carp (Carassius auratus) in natural waters. BMC Genet. 2011, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.F.; Wang, Z.W.; Zhou, L.; Jiang, L.; Zhang, X.J.; Apalikova, O.V.; Brykov, V.A.; Gui, J.F. High male incidence and evolutionary implications of triploid form in northeast Asia Carassius auratus complex. Mol. Phylogenet. Evol. 2013, 66, 350–359. [Google Scholar] [CrossRef]
- Knytl, M.; Kalous, L.; Rab, P. Karyotype and chromosome banding of endangered crucian carp, Carassius carassius (Linnaeus, 1758) (Teleostei, Cyprinidae). Comp. Cytogenet. 2013, 7, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Knytl, M.; Kalous, L.; Rylková, K.; Choleva, L.; Merilä, J.; Ráb, P. Morphologically indistinguishable hybrid Carassius female with 156 chromosomes: A threat for the threatened crucian carp, C. carassius, L. PLoS ONE 2018, 13, e0190924. [Google Scholar] [CrossRef] [PubMed]
- Bertollo, L.; Cioffi, M. Direct chromosome preparation from freshwater teleost fishes. In Fish Cytogenetic Techniques: Ray-Fin Fishes and Chondrichthyans; Ozouf-Costaz, C., Pisano, E., Foresti, F., Foresti, L.D.A.T., Eds.; CRC Press: Enfield, CT, USA, 2015; pp. 21–26. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020; Available online: https://www.R-project.org (accessed on 20 August 2021).
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, PBC.: Boston, MA, USA, 2019; Available online: http://www.rstudio.com (accessed on 20 August 2021).
- Naito, E.; Dewa, K.; Ymanouchi, H.; Kominami, R. Ribosomal ribonucleic acid (rRNA) gene typing for species identification. J. Forensic Sci. 1992, 37, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Sember, A.; Bohlen, J.; Šlechtová, V.; Altmanová, M.; Symonová, R.; Ráb, P. Karyotype differentiation in 19 species of river loach fishes (Nemacheilidae, Teleostei): Extensive variability associated with rDNA and heterochromatin distribution and its phylogenetic and ecological interpretation. BMC Evol. Biol. 2015, 15, 1–22. [Google Scholar] [CrossRef]
- Knytl, M.; Smolík, O.; Kubíčková, S.; Tlapáková, T.; Evans, B.J.; Krylov, V. Chromosome divergence during evolution of the tetraploid clawed frogs, Xenopus mellotropicalis and Xenopus epitropicalis as revealed by Zoo-FISH. PLoS ONE 2017, 12, e0177087. [Google Scholar] [CrossRef] [PubMed]
- Spoz, A.; Boron, A.; Porycka, K.; Karolewska, M.; Ito, D.; Abe, S.; Kirtiklis, L.; Juchno, D. Molecular cytogenetic analysis of the crucian carp, Carassius carassius (Linnaeus, 1758) (Teleostei, Cyprinidae), using chromosome staining and fluorescence in situ hybridisation with rDNA probes. Comp. Cytogenet. 2014, 8, 233–248. [Google Scholar] [CrossRef] [PubMed]
- Sember, A.; Pelikánová, Š.; de Bello Cioffi, M.; Šlechtová, V.; Hatanaka, T.; Do Doan, H.; Knytl, M.; Ráb, P. Taxonomic Diversity Not Associated with Gross Karyotype Differentiation: The Case of Bighead Carps, Genus Hypophthalmichthys (Teleostei, Cypriniformes, Xenocyprididae). Genes 2020, 11, 479. [Google Scholar] [CrossRef]
- Makino, S. Notes on the chromosomes of some fresh-water Teleosts. Jpn. J. Genet. 1934, 9, 100–103. [Google Scholar]
- Cherfas, N.B. Natural triploidy in females of the unisexual form of silver crucian carp (Carassius auratus gibelio Bloch). Genetika 1966, 2, 16–24. [Google Scholar]
- Chiarelli, B.; Ferrantelli, O.; Cucchi, C. The caryotype of some teleostea fish obtained by tissue culture in vitro. Experientia 1969, 25, 426–427. [Google Scholar] [CrossRef]
- Raicu, P.; Taisescu, E.; Banarescu, P. Carassius carassius and Carassius auratus, a pair of diploid and tetraploid representative species (Pices, Cyprinidae). Cytologia 1981, 46, 233–240. [Google Scholar] [CrossRef]
- Fister, S.; Soldatovic, B. Karyotype analysis of male and diploid female Carassius auratus gibelio, Bloch (Pisces, Cyprinidae) caught in the Danube at Belgrade, evidence for the existence of a bisexual population. Acta Vet. Beogr. 1991, 41, 81–90. [Google Scholar]
- Makino, S. A Karyological Study of Gold-fish of Japan. Cytologia 1941, 12, 96–111. [Google Scholar] [CrossRef]
- Ohno, S.; Atkin, N.B. Comparative DNA values and chromosome complements of eight species of fishes. Chromosoma 1966, 18, 455–466. [Google Scholar] [CrossRef]
- Ojima, Y.; Hitotsumachi, S.; Makino, S. Cytogenetic Studies in Lower Vertebrates. I A Preliminary Report on the Chromosomes of the Funa (Carassius auratus) and Gold-fish (A Revised Study). Proc. Jpn. Acad. 1966, 42, 62–66. [Google Scholar] [CrossRef]
- Ojima, Y.; Hitotsumachi, S. Cytogenetic studies in lower vertebrates. IV. a note on the chromosomes of the carp (cyprinus carpio) in comparison with those of the funa and the goldfish (Carassius auratus). Jpn. J. Genet. 1967, 42, 163–167. [Google Scholar] [CrossRef]
- Arai, R.; Fujiki, A. Chromosomes of three races of Goldfish, Kuro-demekin, Sanshiki- demekin and Ranchu. Bull. Natl. Sci. Mus. Ser. A Zool. 1977, 3, 187–192. [Google Scholar]
- Ojima, Y.; Ueda, T.; Narikawa, T. A Cytogenetic Assessment on the Origin of the Gold-fish. Proc. Jpn. Acad. Ser. B 1979, 55, 58–63. [Google Scholar] [CrossRef]
- Ojima, Y.; Yamano, T. The assignment of the nucleolar organizer in the chromosomes of the funa (Carassius, cyprinidae, pisces). Proc. Jpn. Acad. Ser. B 1980, 56, 551–556. [Google Scholar] [CrossRef]
- Zan, R.G.; Song, Z. Analysis and comparison between the karyotypes of Cyprinus carpio and Carassius auratus as well as Aristichthys nobilis and Hypophthalmichthys molitrix. Acta Genet. Sin. 1980, 7, 72–77. [Google Scholar]
- Zan, R.G. Studies of sex chromosomes and C-banding karyotypes of two forms of Carassius auratus in Kunming Lake. Acta Genet. Sin. 1982, 9, 32–39. [Google Scholar]
- Wang, R.F.; Shi, L.M.; He, W.S. A Comparative Study of the Ag-NORS of Carassius auratus from Different Geographic Districts. Zool. Res. 1988, 9, 165–169. [Google Scholar]
- Kasama, M.; Kobayasi, H. Hybridization Experiment Between Female Crucian Carp and Male Grass Carp. Nippon. Suisan Gakkaishi Jpn. Ed. 1989, 55, 1001–1006. [Google Scholar] [CrossRef]
- Kasama, M.; Kobayasi, H. Hybridization experiment between Carassius carassius female and Gnathopogon elongatus elongatus male. Jpn. J. Ichthyol. 1990, 36, 419–426. [Google Scholar] [CrossRef]
- Kasama, M.; Kobayashi, H. Hybridization experiment between Gnathopogon elongatus elongatus female and Carassius carassius male. Jpn. J. Ichthyol. 1991, 38, 295–300. [Google Scholar] [CrossRef]
- Hafez, R.; Labat, R.; Quillier, R. Cytogenetic study of some species of Cyprinidae from the Midi-Pyrenees region. Bull. Soc. D’Hist. Nat. Toulouse 1978, 114, 122–159. [Google Scholar]
- Sofradžija, A.; Berberović, L.; Hadžiselimović, R. Hromosomske garniture karaša (Carassius carassius) i babuške (Carassius auratus gibelio). Ichthyologia 1978, 10, 135–148. [Google Scholar]
- Mayr, B.; Ráb, P.; Kalat, M. NORs and counterstain-enhanced fluorescence studies in Cyprinidae of different ploidy level. Genetica 1986, 69, 111–118. [Google Scholar] [CrossRef]
- Kobayasi, H.; Ochi, H.; Takeuchi, N. Chromosome studies in the genus Carassius: Comparison of C. auratus grandoculis, C. auratus buergeri, and C. auratus langsdorfii. Jpn. J. Ichthyol. 1973, 20, 6. [Google Scholar] [CrossRef]
- Boron, A. Karyotypes of diploid and triploid silver crucian carp Carassius auratus gibelio (Bloch). Cytobios 1994, 80, 117–124. [Google Scholar]
- Boron, A.; Szlachciak, J.; Juchno, D.; Grabowska, A.; Jagusztyn, B.; Porycka, K. Karyotype, morphology, and reproduction ability of the Prussian carp, Carassius gibelio (Actinopterygii: Cypriniformes: Cyprinidae), from unisexual and bisexual populations in Poland. Acta Ichthyol. Piscat. 2011, 41, 19–28. [Google Scholar] [CrossRef]
- Kalous, L.; Bohlen, J.; Rylková, K.; Petrtýl, M. Hidden diversity within the Prussian carp and designation of a neotype for Carassius gibelio (Teleostei: Cyprinidae). Ichthyol. Explor. Freshwaters 2012, 23, 11–18. [Google Scholar]
- Papoušek, I.; Vetešník, L.; Halačka, K.; Lusková, V.; Humpl, M.; Mendel, J. Identification of natural hybrids of gibel carp Carassius auratus gibelio (Bloch) and crucian carp Carassius carassius (L.) from lower Dyje River floodplain (Czech Republic). J. Fish Biol. 2008, 72, 1230–1235. [Google Scholar] [CrossRef]
- Wouters, J.; Janson, S.; Lusková, V.; Olsén, K.H. Molecular identification of hybrids of the invasive gibel carp Carassius auratus gibelio and crucian carp Carassius carassius in Swedish waters. J. Fish Biol. 2012, 80, 2595–2604. [Google Scholar] [CrossRef] [PubMed]
- King, M. Species Evolution: The Role of Chromosome Change; Cambridge University Press: Cambridge, UK, 1993; p. 336. [Google Scholar]
- Cremer, T.; Cremer, C. Chromosome territories, nuclear architecture and gene regulation in mammalian cells. Nat. Rev. Genet. 2001, 2, 292–301. [Google Scholar] [CrossRef] [PubMed]
- Chevret, E.; Volpi, E.; Sheer, D. Mini review: Form and function in the human interphase chromosome. Cytogenet. Genome Res. 2000, 90, 13–21. [Google Scholar] [CrossRef]
- Claussen, U.; Michel, S.; Mhlig, P.; Westermann, M.; Grummt, U.W.; Kromeyer-Hauschild, K.; Liehr, T. Demystifying chromosome preparation and the implications for the concept of chromosome condensation during mitosis. Cytogenet. Genome Res. 2002, 98, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Tjio, J.H.; Levan, A. The chromosome number of man. Hereditas 1956, 42, 1–6. [Google Scholar] [CrossRef]
- Zhu, H.P.; Ma, D.M.; Gui, J.F. Triploid origin of the gibel carp as revealed by 5S rDNA localization and chromosome painting. Chromosome Res. 2006, 14, 767–776. [Google Scholar] [CrossRef]
- Mantovani, M.; Abel, L.D.D.S.; Moreira-Filho, O. Conserved 5S and variable 45S rDNA chromosomal localisation revealed by FISH in Astyanax scabripinnis (Pisces, Characidae). Genetica 2005, 123, 211–216. [Google Scholar] [CrossRef]
- Gromicho, M.; Coutanceau, J.P.; Ozouf-Costaz, C.; Collares-Pereira, M.J. Contrast between extensive variation of 28S rDNA and stability of 5S rDNA and telomeric repeats in the diploid-polyploid Squalius alburnoides complex and in its maternal ancestor Squalius pyrenaicus (Teleostei, Cyprinidae). Chromosome Res. 2006, 14, 297–306. [Google Scholar] [CrossRef]
- Schmid, M.; Vitelli, L.; Batistoni, R. Chromosome banding in amphibia. XI. Constitutive heterochromatin, nucleolus organizers, 18S + 28S and 5S ribosomal RNA genes in Ascaphidae, Pipidae, Discoglossidae and Pelobatidae. Chromosoma 1987, 95, 271–284. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, Y.; Gui, J.F. Genetic Evidence for Gonochoristic Reproduction in Gynogenetic Silver Crucian Carp (Carassius auratus gibelio Bloch) as Revealed by RAPD Assays. J. Mol. Evol. 2000, 51, 498–506. [Google Scholar] [CrossRef]
- Daněk, T.; Kalous, L.; Veselý, T.; Krásová, E.; Reschová, S.; Rylková, K.; Kulich, P.; Petrtýl, M.; Pokorová, D.; Knytl, M. Massive mortality of Prussian carp Carassius gibelio in the upper Elbe basin associated with herpesviral hematopoietic necrosis (CyHV-2). Dis. Aquat. Org. 2012, 102, 87–95. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Centromeric Position | Arm Ratio | Centromeric Index | Chromosome Category |
|---|---|---|---|
| median sensu stricto | 1.00 | 50 | M (metacentric sensu stricto) |
| median | 1.01–1.70 | 49.9–37.51 | m (metacentric) |
| submedian | 1.71–3.00 | 37.50–25.01 | sm (submetacentric) |
| subterminal | 3.01–7.00 | 25.00–12.51 | st (subtelocentric) |
| terminal | >7.01 | 12.50–0.01 | a/t (acro-/telocentric) |
| terminal sensu stricto | ∞ | 0 | T (telocentric sensu stricto) |
| CAU_median_i | CCA_median_i | CGI_median_i | CAU_category | CCA_category | CGI_category | |
|---|---|---|---|---|---|---|
| chromosome1 | 47.87 | 48.28 | 48.55 | m | m | m |
| chromosome2 | 47.07 | 45.68 | 46.12 | m | m | m |
| chromosome3 | 45.74 | 44.74 | 44.99 | m | m | m |
| chromosome4 | 44.81 | 43.52 | 43.47 | m | m | m |
| chromosome5 | 43.46 | 42.11 | 42.53 | m | m | m |
| chromosome6 | 42.43 | 41.08 | 40.89 | m | m | m |
| chromosome7 | 41.63 | 39.58 | 39.89 | m | m | m |
| chromosome8 | 40.84 | 37.90 | 38.42 | m | m | m |
| chromosome9 | 40.13 | 36.81 | 36.91 | m | sm | sm |
| chromosome10 | 39.67 | 36.17 | 35.84 | m | sm | sm |
| chromosome11 | 38.75 | 35.79 | 35.41 | m | sm | sm |
| chromosome12 | 37.65 | 35.19 | 34.73 | m | sm | sm |
| chromosome13 | 36.77 | 34.86 | 34.25 | sm | sm | sm |
| chromosome14 | 36.19 | 34.36 | 33.36 | sm | sm | sm |
| chromosome15 | 35.88 | 33.66 | 32.61 | sm | sm | sm |
| chromosome16 | 35.71 | 32.70 | 31.90 | sm | sm | sm |
| chromosome17 | 35.48 | 32.22 | 28.62 | sm | sm | sm |
| chromosome18 | 35.21 | 31.23 | 26.86 | sm | sm | sm |
| chromosome19 | 34.87 | 30.54 | 26.07 | sm | sm | sm |
| chromosome20 | 34.33 | 29.11 | 24.42 | sm | sm | st |
| chromosome21 | 33.45 | 28.54 | 23.45 | sm | sm | st |
| chromosome22 | 33.03 | 27.41 | 22.83 | sm | sm | st |
| chromosome23 | 32.78 | 26.86 | 21.37 | sm | sm | st |
| chromosome24 | 31.95 | 26.25 | 16.52 | sm | sm | st |
| chromosome25 | 31.41 | 25.22 | 9.19 | sm | sm | a |
| chromosome26 | 31.08 | 24.00 | 0.00 | sm | st | T |
| chromosome27 | 30.70 | 22.57 | 0.00 | sm | st | T |
| chromosome28 | 30.11 | 20.62 | 0.00 | sm | st | T |
| chromosome29 | 29.75 | 18.13 | 0.00 | sm | st | T |
| chromosome30 | 28.96 | 0.00 | 0.00 | sm | T | T |
| chromosome31 | 28.17 | 0.00 | 0.00 | sm | T | T |
| chromosome32 | 26.87 | 0.00 | 0.00 | sm | T | T |
| chromosome33 | 24.41 | 0.00 | 0.00 | st | T | T |
| chromosome34 | 11.19 | 0.00 | 0.00 | a | T | T |
| chromosome35 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome36 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome37 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome38 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome39 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome40 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome41 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome42 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome43 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome44 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome45 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome46 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome47 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome48 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome49 | 0.00 | 0.00 | 0.00 | T | T | T |
| chromosome50 | 0.00 | 0.00 | 0.00 | T | T | T |
| Karyotype | Sex | Locality | References |
|---|---|---|---|
| C. auratus | |||
| NA | Japan | [42,47] | |
| –104 | F, M | NA | [48] |
| – | F, M | Japan | [49,50] |
| F, M | NA | [22] | |
| – | NA | NA | [44] |
| F, M | NA | [17] | |
| – | NA | NA | [51] |
| – | F, M | China | [18,52,53] |
| – | F, M | China | [54,55,56] |
| C. carassius | |||
| – | NA | NA | [44] |
| F, M | Netherlands | [17,57,58,59] | |
| NA | France | [60] | |
| F, M | Bosnia | [61] | |
| – | NA | Romania | [45] |
| F, M | Czech Republic | [62] | |
| – | F, M | Czech Republic | [16,31] |
| – | F, M | Poland | [40] |
| – | M | Finland | [32] |
| C. gibelio | |||
| F, M | Belarus | [43] | |
| NA | River Amur | [63] | |
| – | NA | Romania | [45] |
| – | F | Yugoslavia | [46] |
| – | M | Yugoslavia | [46] |
| – | F, M | Poland | [64] |
| – | F, M | Poland | [65] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Knytl, M.; Fornaini, N.R. Measurement of Chromosomal Arms and FISH Reveal Complex Genome Architecture and Standardized Karyotype of Model Fish, Genus Carassius. Cells 2021, 10, 2343. https://doi.org/10.3390/cells10092343
Knytl M, Fornaini NR. Measurement of Chromosomal Arms and FISH Reveal Complex Genome Architecture and Standardized Karyotype of Model Fish, Genus Carassius. Cells. 2021; 10(9):2343. https://doi.org/10.3390/cells10092343
Chicago/Turabian StyleKnytl, Martin, and Nicola Reinaldo Fornaini. 2021. "Measurement of Chromosomal Arms and FISH Reveal Complex Genome Architecture and Standardized Karyotype of Model Fish, Genus Carassius" Cells 10, no. 9: 2343. https://doi.org/10.3390/cells10092343