
Cytogenetic Evidence for Sex Chromosomes and Karyotype Evolution in Anguimorphan Lizards


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
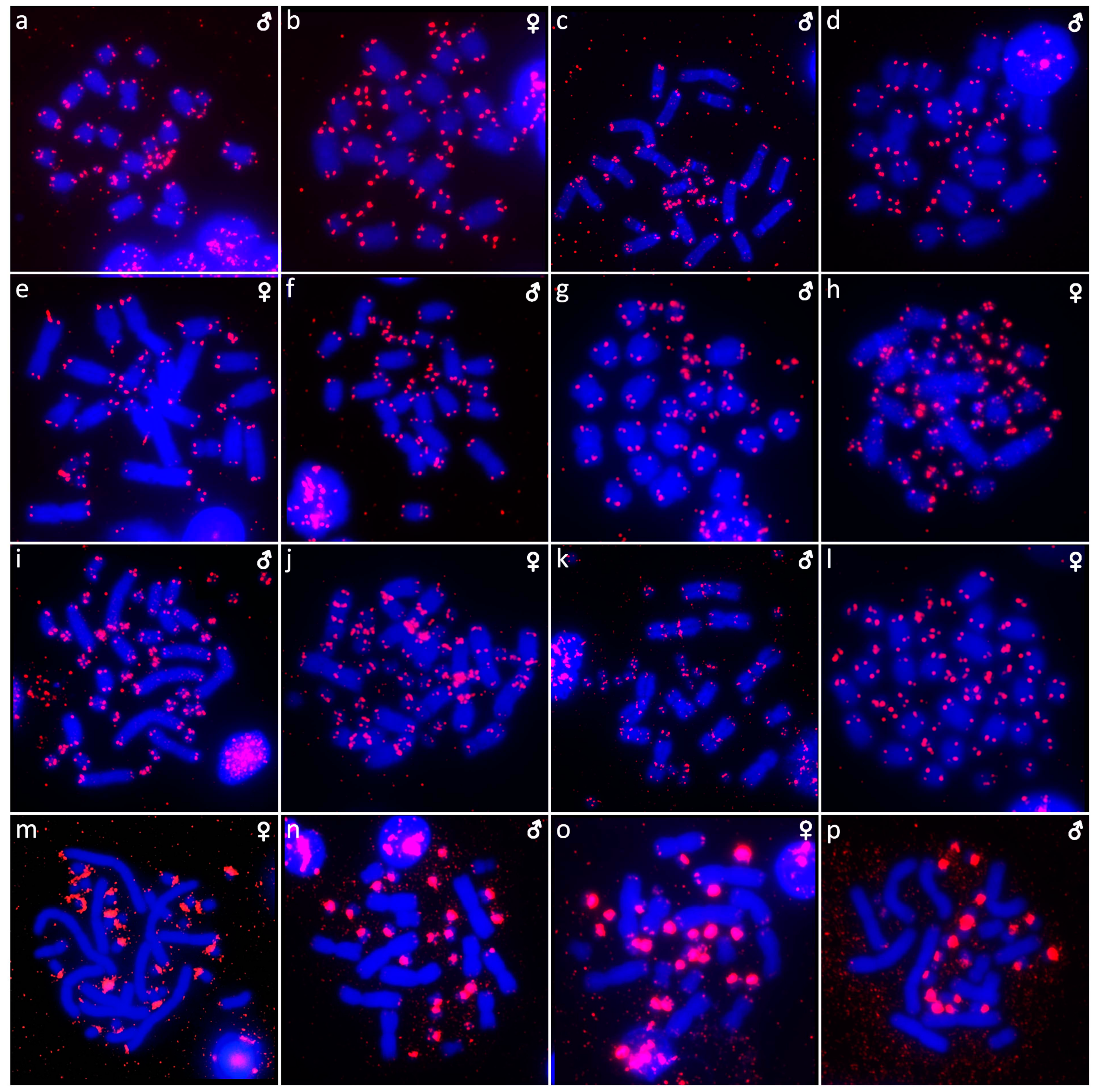{kind=link}
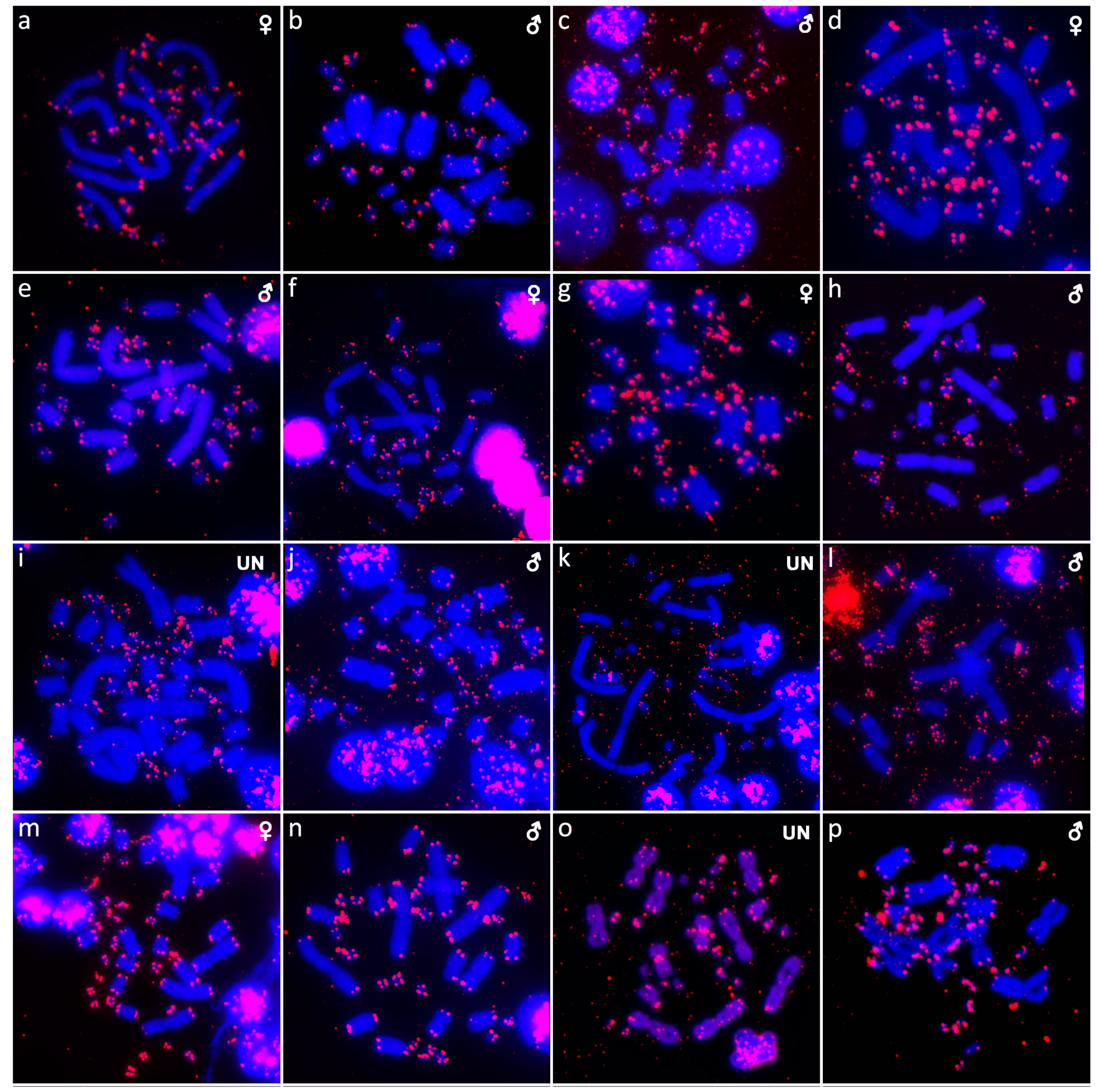{kind=link}
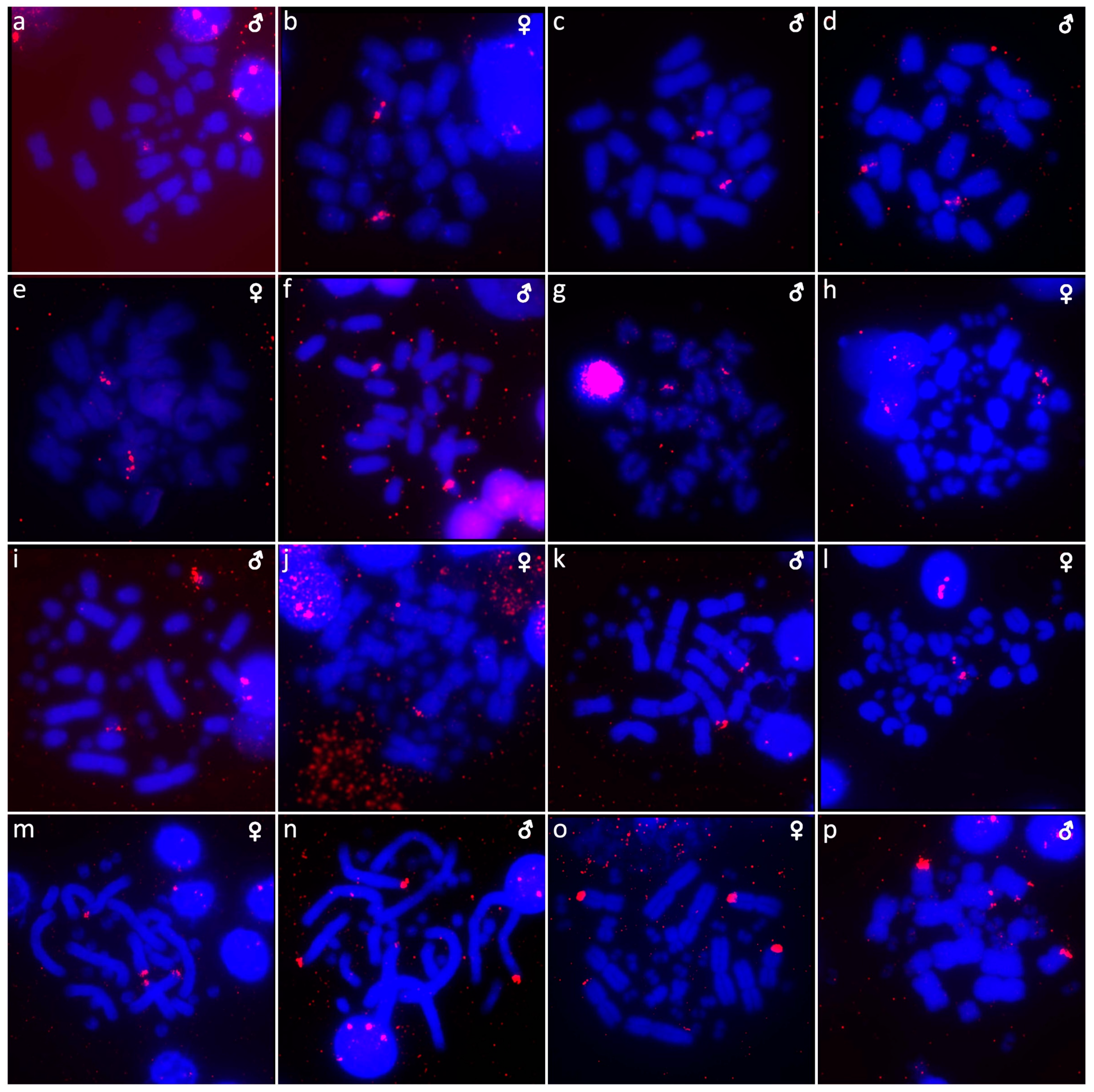{kind=link}
{kind=link}
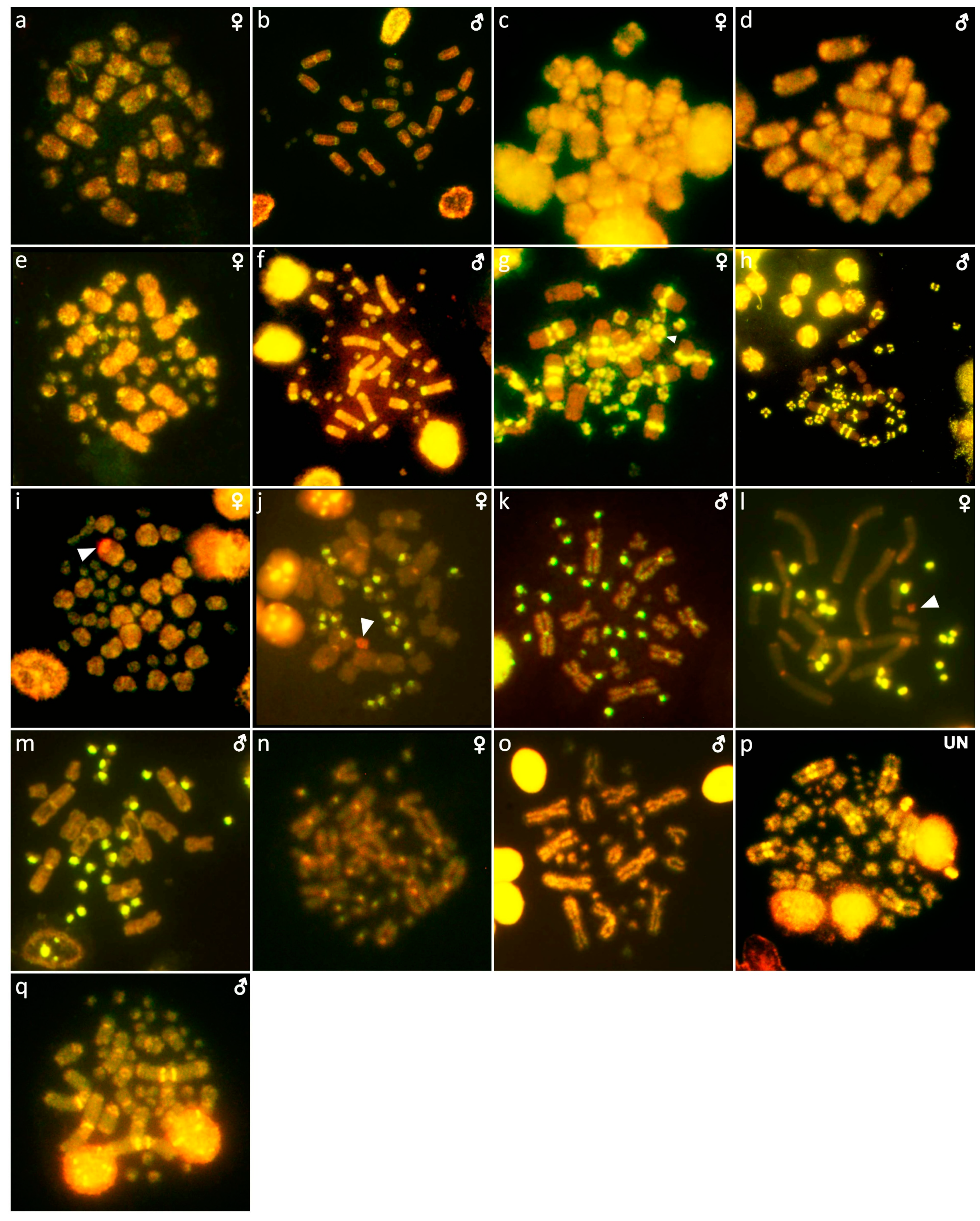{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Studied Material, Chromosome Preparations and Staining
2.2. Fluorescence In Situ Hybridization with Probe for Telomeric Sequences
2.3. Fluorescence In Situ Hybridization with Probe for 18S/28S rDNA Loci
2.4. Comparative Genome Hybridization
2.5. Microscopy and Image Analyses
2.6. qPCR Test for Sex Chromosome Constitution in the Triploid Varanus primordius
3. Results
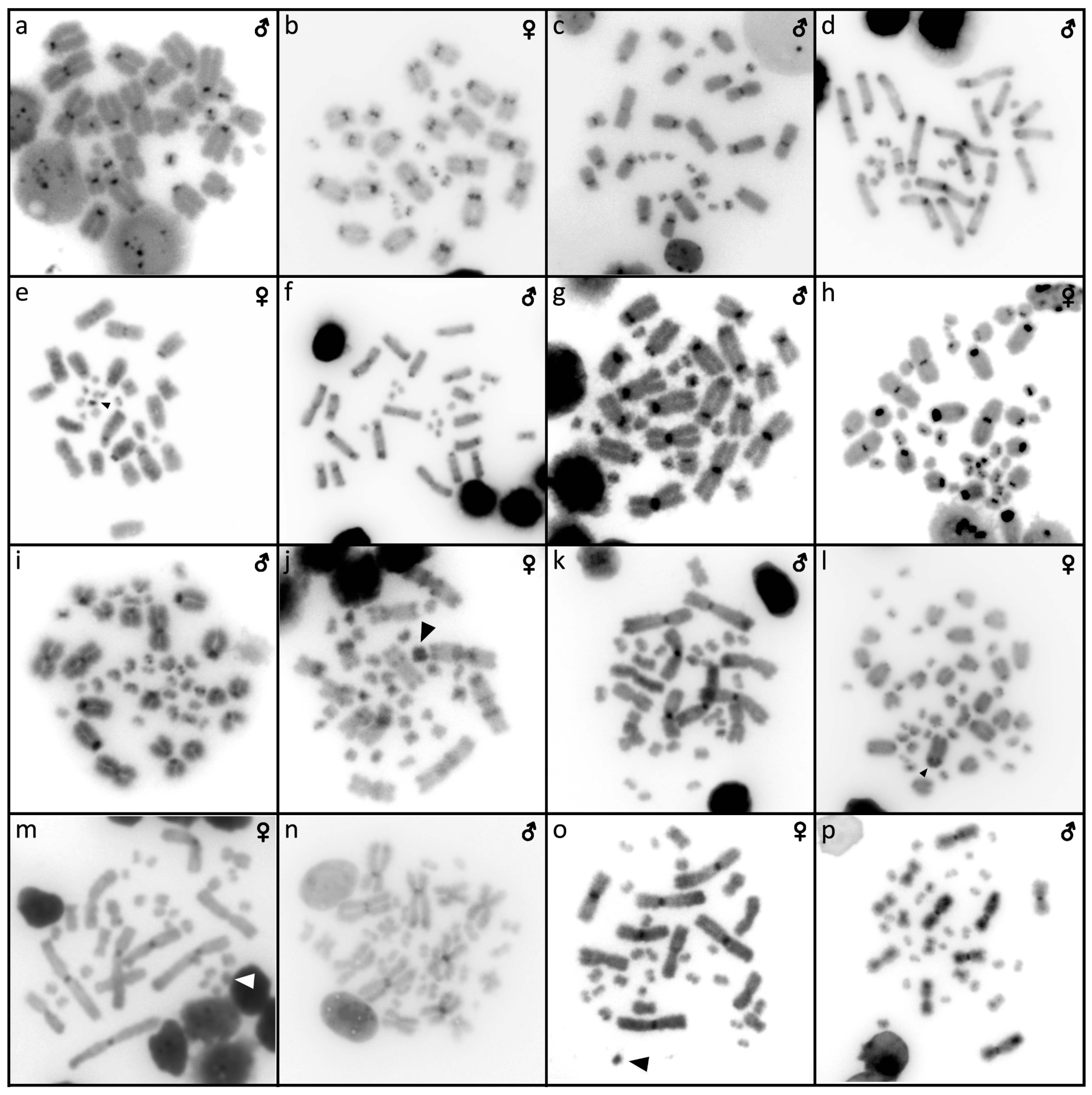
3.1. Abronia campbelli (Brodie & Savage, 1993)
3.2. Abronia deppii (Wiegmann, 1828)
3.3. Abronia graminea (Cope, 1864)
3.4. Abronia lythrochila Smith & Alvarez del Toro, 1963
3.5. Abronia mixteca Bogert & Porter, 1967
3.6. Abronia smithi Campbell & Frost, 1993
3.7. Abronia taeniata (Wiegmann, 1828)
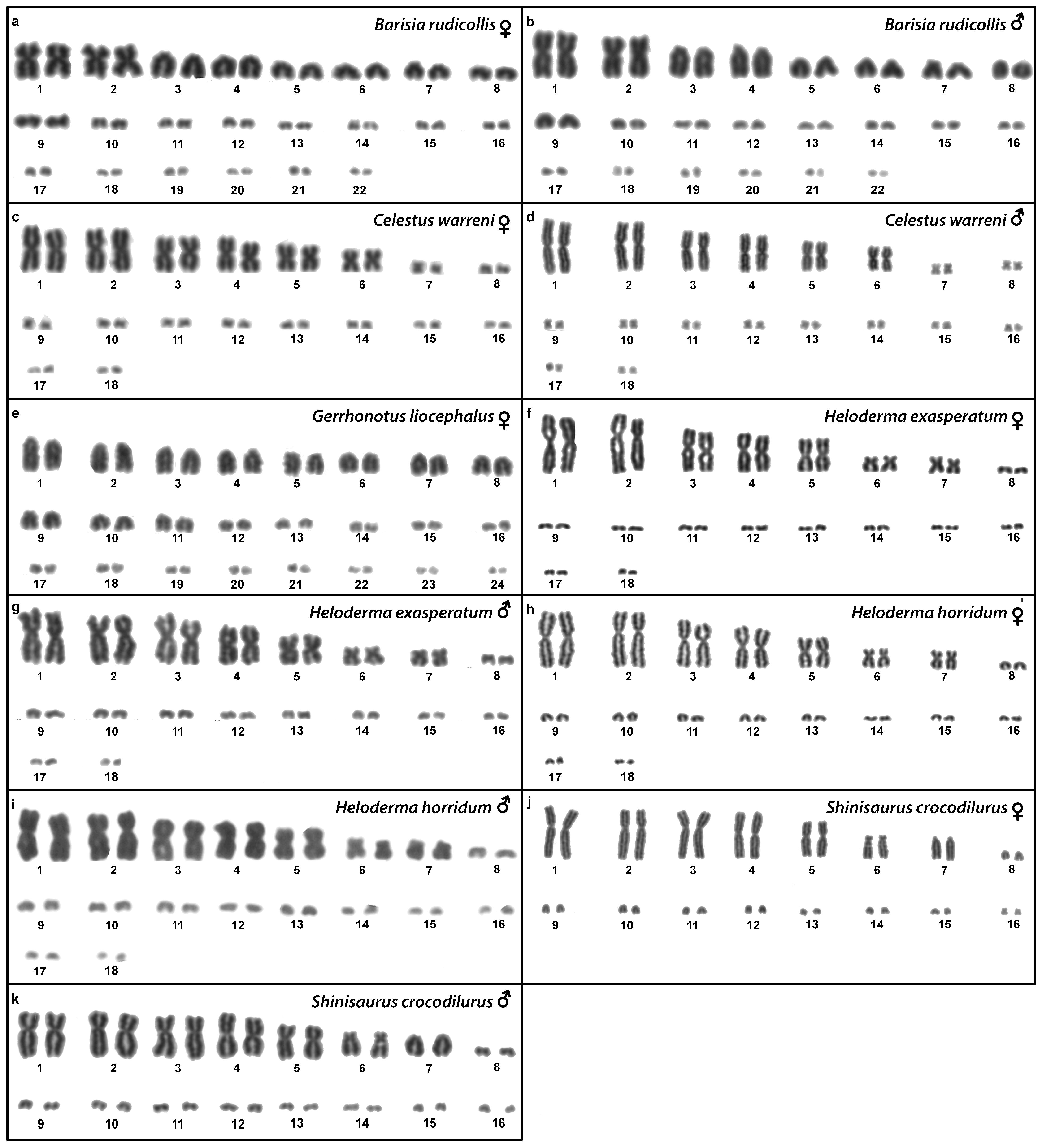
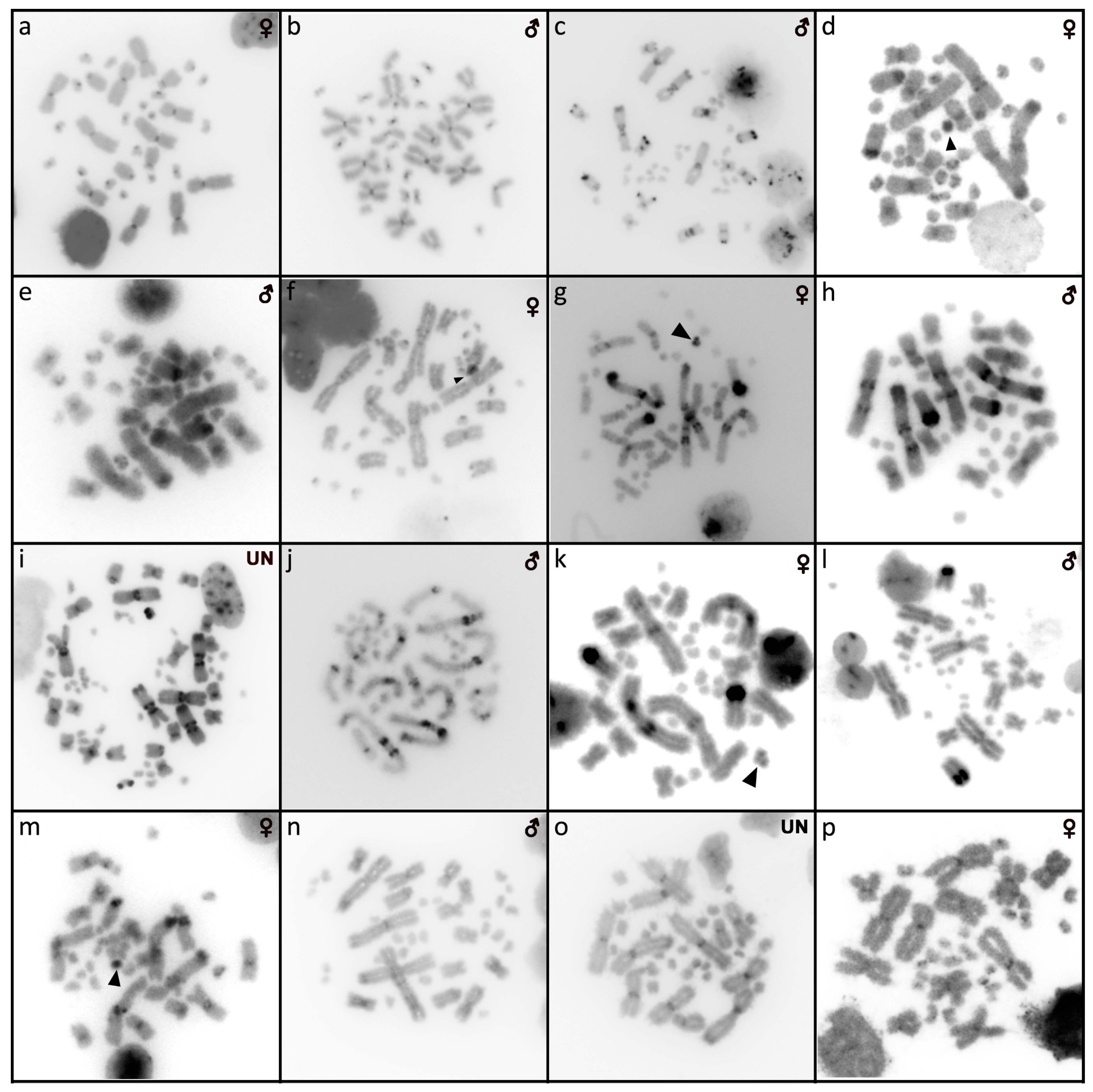
3.8. Barisia rudicollis (Wiegmann, 1828)
3.9. Celestus warreni (Schwartz, 1970)
3.10. Gerrhonotus liocephalus (Wiegmann, 1828)
3.11. Heloderma exasperatum Bogert & Martin del Campo, 1956
3.12. Heloderma horridum (Wiegmann, 1828)
3.13. Shinisaurus crocodilurus Ahl, 1930
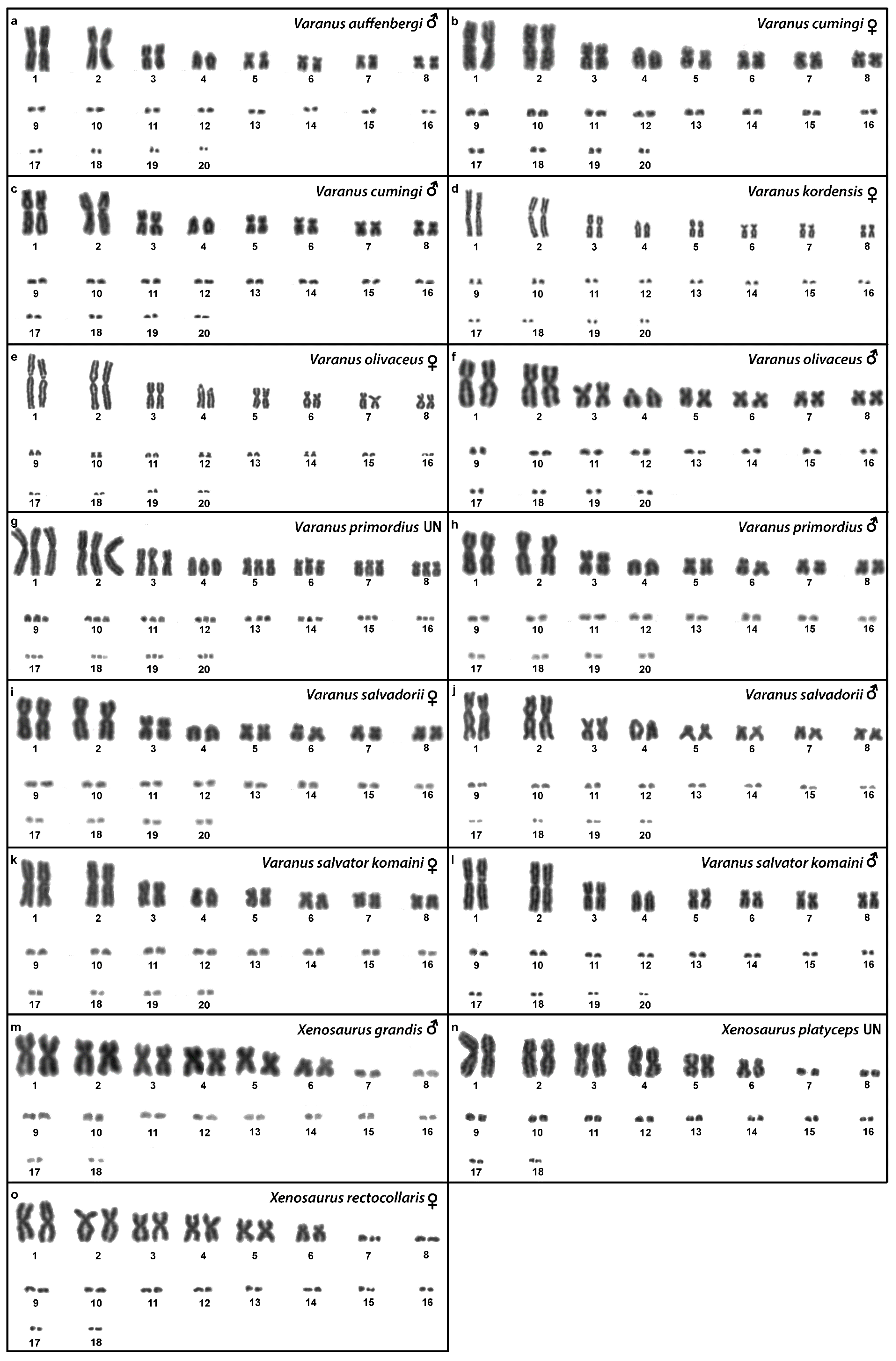
3.14. Varanus auffenbergi Sprackland, 1999
3.15. Varanus cumingi Martin, 1893
3.16. Varanus kordensis (Meyer, 1874)
3.17. Varanus olivaceus Hallowell, 1857
3.18. Varanus primordius Mertens, 1942
3.19. Varanus salvadorii (Peters & Doria, 1878)
3.20. Varanus salvator komaini Nutaphand 1987
3.21. Xenosaurus grandis (Gray, 1856)
3.22. Xenosaurus platyceps King & Thompson, 1968
3.23. Xenosaurus rectocollaris Smith & Iverson, 1993
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilson, M.A.; Makova, K.D. Genomic analyses of sex chromosome evolution. Rev. Genom. Hum. Genet. 2009, 10, 333–354. [Google Scholar] [CrossRef] [PubMed]
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Tree of sex consortium. Sex determination: Why so many ways of doing it? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef] [Green Version]
- Pennell, M.W.; Mank, J.E.; Peichel, C.L. Transitions in sex determination and sex chromosomes across vertebrate species. Mol. Ecol. 2018, 27, 3950–3963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, S. Sex Chromosomes and Sex-Linked Genes; Springer: Berlin/Heidelberg, Germany, 1967. [Google Scholar]
- Rice, W.R. The accumulation of sexually antagonistic genes as a selective agent promoting the evolution of reduced recombination between primitive sex chromosomes. Evolution 1987, 41, 911–914. [Google Scholar] [CrossRef] [PubMed]
- Charlesworth, B. The evolution of chromosomal sex determination and dosage compensation. Curr. Biol. 1996, 6, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Kostmann, A.; Kratochvíl, L.; Rovatsos, M. Poorly differentiated XX/XY sex chromosomes are widely shared across skink radiation. Proc. R. Soc. B 2021, 288, 20202139. [Google Scholar] [CrossRef]
- Rovatsos, M.; Johnson Pokorná, M.; Altmanová, M.; Kratochvíl, L. Female heterogamety in Madagascar chameleons (Squamata: Chamaeleonidae: Furcifer): Differentiation of sex and neo-sex chromosomes. Sci. Rep. 2015, 5, 13196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rovatsos, M.; Altmanová, M.; Augstenová, B.; Mazzoleni, S.; Velenský, P.; Kratochvíl, L. ZZ/ZW sex determination with multiple neo-sex chromosomes is common in Madagascan chameleons of the genus Furcifer (Reptilia: Chamaeleonidae). Genes 2019, 10, 1020. [Google Scholar] [CrossRef] [Green Version]
- Epplen, J.T.; McCarrey, J.R.; Sutou, S.; Ohno, S. Base sequence of a cloned snake W-chromosome 65DNA fragment and identification of a male-specific putative mRNA in the mouse. Proc. Natl. Acad. Sci. USA 1982, 79, 3798–3802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nanda, I.; Deubelbeiss, C.; Guttenbach, M.; Epplen, J.T.; Schmid, M. Heterogeneities in the distribution of (GACA)n simple repeats in the karyotypes of primates and mouse. Hum. Genet. 1990, 85, 187–194. [Google Scholar] [CrossRef]
- Steinemann, S.; Steinemann, M. Retroelements: Tools for sex chromosome evolution. Cytogenet. Genome Res. 2005, 110, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Kejnovsky, E.; Hobza, R.; Cermak, T.; Kubat, Z.; Vyskot, B. The role of repetitive DNA in structure and evolution of sex chromosomes in plants. Heredity 2009, 102, 533–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Meally, D.; Patel, H.R.; Stiglec, R.; Sarre, S.D.; Georges, A.; Marshall Graves, J.A.; Ezaz, T. Non-homologous sex chromosomes of birds and snakes share repetitive sequences. Chromosome Res. 2010, 18, 787–800. [Google Scholar] [CrossRef]
- Pokorná, M.; Kratochvíl, L.; Kejnovský, E. Microsatellite distribution on sex chromosomes at different stages of heteromorphism and heterochromatinization in two lizard species (Squamata: Eublepharidae: Coleonyx elegans and Lacertidae: Eremias velox). BMC Genet. 2011, 12, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsubara, K.; Sarre, S.D.; Georges, A.; Matsuda, Y.; Graves, J.A.M.; Ezaz, T. Highly differentiated ZW sex microchromosomes in the Australian Varanus species evolved through rapid amplification of repetitive sequences. PLoS ONE 2014, 9, e95226. [Google Scholar] [CrossRef]
- Matsubara, K.; O’Meally, D.; Azad, B.; Georges, A.; Sarre, S.D.; Graves, J.A.M.; Matsuda, Y.; Ezaz, T. Amplification of microsatellite repeat motifs is associated with the evolutionary differentiation and heterochromatinization of sex chromosomes in Sauropsida. Chromosoma 2016, 125, 111–123. [Google Scholar] [CrossRef]
- Augstenová, B.; Mazzoleni, S.; Kratochvíl, L.; Rovatsos, M. Evolutionary dynamics of the W chromosome in caenophidian snakes. Genes 2018, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Suwala, G.; Altmanová, M.; Mazzoleni, S.; Karameta, E.; Pafilis, P.; Kratochvíl, L.; Rovatsos, M. Evolutionary variability of W-linked repetitive content in Lacertid lizards. Genes 2020, 11, 531. [Google Scholar] [CrossRef]
- Uetz, P.; Freed, P.; Hošek, J. (Eds.) The Reptile Database. 2020. Available online: http://www.reptile-database.org (accessed on 5 March 2021).
- Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol. 2013, 13, 1–54. [Google Scholar] [CrossRef] [Green Version]
- Gorman, G.C.; Gress, F. Chromosome cytology of four boid snakes and a varanid lizard, with comments on the cytosystematics of primitive snakes. Herpetologica 1970, 26, 308–317. [Google Scholar]
- Singh, L.; Sharma, T.; Ray-Chaudhu, S.P. Chromosome numbers and sex chromosomes in few Indian species of amphibia and reptiles. Mamm. Chrom. News 1970, 11, 91–94. [Google Scholar]
- Singh, L. Study of mitotic and meiotic chromosomes in seven species of lizards. Proc. Zool. Soc. 1974, 27, 57–79. [Google Scholar]
- King, M.; King, D. Chromosomal evolution in the lizard genus Varanus (Reptilia). Aust. J. Biol. Sci. 1975, 28, 89–108. [Google Scholar] [CrossRef]
- De Smet, W.H.O. Description of the orsein stained karyotypes of 136 lizard species (Lacertilia, Reptilia) belonging to the families Teiidae, Scincidae, Lacertidae, Cordylidae and Varanidae (Autarchoglossa). Acta Zool. Pathol. Antverp. 1981, 76, 407–420. [Google Scholar]
- King, M.; Mengden, G.A.; King, D. A pericentric-inversion polymorphism and a ZZ/ZW sex-chromosome system in Varanus acanthurus Boulenger analysed by G- and C-banding and Ag staining. Genetica 1982, 58, 39–45. [Google Scholar] [CrossRef]
- Porter, C.; Haiduk, M.; De Queiroz, K. Evolution and phylogenetic significance of ribosomal gene location in chromosomes of squamate reptiles. Copeia 1994, 1994, 302–313. [Google Scholar] [CrossRef]
- Srikulnath, K.; Uno, Y.; Nishida, C.; Matsuda, Y. Karyotype evolution in monitor lizards: Cross-species chromosome mapping of cDNA reveals highly conserved synteny and gene order in the Toxicofera clade. Chromosome Res. 2013, 21, 805–819. [Google Scholar] [CrossRef]
- Johnson Pokorná, M.; Altmanová, M.; Rovatsos, M.; Velenský, P.; Vodička, R.; Rehák, I.; Kratochvíl, L. First description of the karyotype and sex chromosomes in the Komodo dragon (Varanus komodoensis). Cytogenet. Genome Res. 2016, 148, 284–291. [Google Scholar] [CrossRef]
- Patawang, I.; Tanomtong, A. Constitutive heterochromatin observed on metaphase chromosome of Varanus bengalensis by C-banding and DAPI methods. Cytologia 2017, 82, 1. [Google Scholar] [CrossRef]
- Patawang, I.; Tanomtong, A.; Getlekha, N.; Phimphan, S.; Pinthong, K.; Neeratanaphan, L. Standardized karyotype and idiogram of bengal monitor lizard, Varanus bengalensis (Squamata, Varanidae). Cytologia 2017, 82, 75–82. [Google Scholar] [CrossRef]
- Iannucci, A.; Altmanová, M.; Ciofi, C.; Ferguson-Smith, M.; Milan, M.; Pereira, J.C.; Pether, J.; Rehák, I.; Rovatsos, M.; Stanyon, R.; et al. Conserved sex chromosomes and karyotype evolution in monitor lizards (Varanidae). Heredity 2019, 123, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Rehák, I.; Velenský, P.; Kratochvíl, L. Shared ancient sex chromosomes in varanids, beaded lizards, and alligator lizards. Mol. Biol. Evol. 2019, 36, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Lind, A.L.; Lai, Y.Y.; Mostovoy, Y.; Holloway, A.K.; Iannucci, A.; Mak, A.C.Y.; Fondi, M.; Orlandini, V.; Eckalbar, W.L.; Milan, M.; et al. Genome of the Komodo dragon reveals adaptations in the cardiovascular and chemosensory systems of monitor lizards. Nat. Ecol. Evol. 2019, 3, 1241–1252. [Google Scholar] [CrossRef] [Green Version]
- Johnson Pokorná, M.; Rovatsos, M.; Kratochvíl, L. Sex chromosomes and karyotype of the (nearly) mythical creature, the Gila monster, Heloderma suspectum (Squamata: Helodermatidae). PLoS ONE 2014, 9, e104716. [Google Scholar]
- Bury, R.B.; Gorman, G.C.; Lynch, J.F. Karyotypic data for five species of anguid lizards. Experientia 1969, 25, 314–316. [Google Scholar] [CrossRef]
- Beçak, M.L.; Beçak, W.; Denaro, L. Chromosome polymorphism, geographical variation and karyotypes in Sauria. Caryologia 1972, 25, 313–326. [Google Scholar] [CrossRef]
- Stamm, B.; Gorman, G.C. Notes on the chromosomes of Anolis agassizi (Sauria: Iguanidae) and Diploglossus millepunctatus (Sauria: Anguidae). In The Biological Investigation of Malpelo Island; Graham, J.B., Ed.; Smithsonian Contributions to Zoology Colombia: Washington, DC, USA, 1975; Volume 176, pp. 52–56. [Google Scholar]
- Bezy, R.L.; Gorman, G.C.; Kim, Y.J.; Wright, J.W. Chromosomal and genetic divergence in the fossorial lizards of the family Anniellidae. Syst. Biol. 1977, 26, 57–71. [Google Scholar] [CrossRef]
- Olmo, E.; Signorino, G.G. Chromorep: A Reptile Chromosomes Database. 2005. Available online: http://chromorep.univpm.it (accessed on 4 March 2021 ).
- Mezzasalma, M.; Guarino, F.M.; Aprea, G.; Petraccioli, A.; Crottini, A.; Odierna, G. Karyological evidence for diversification of Italian slow worm populations (Squamata, Anguidae). Comp. Cytogenet. 2013, 7, 217. [Google Scholar] [CrossRef] [PubMed]
- Papenfuss, T.J.; Parham, J.F. Four new species of California legless lizards (Anniella). Breviora 2013, 536, 1–17. [Google Scholar] [CrossRef]
- Mazzoleni, S.; Augstenová, B.; Clemente, L.; Auer, M.; Fritz, U.; Praschag, P.; Protiva, T.; Velenský, P.; Kratochvíl, L.; Rovatsos, M. Turtles of the genera Geoemyda and Pangshura (Testudines: Geoemydidae) lack differentiated sex chromosomes: The end of a 40-year error cascade for Pangshura. PeerJ 2019, 7, e6241. [Google Scholar] [CrossRef] [Green Version]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Ijdo, J.W.; Baldini, A.; Ward, D.C.; Reeders, S.T.; Wells, R.A. Origin of human chromosome 2: An ancestral telomere-telomere fusion. Proc. Natl. Acad. Sci. USA 1991, 88, 9051–9055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endow, S.A. Polytenization of the ribosomal genes on the X and Y chromosomes of Drosophila melanogaster. Genetics 1982, 100, 375–385. [Google Scholar] [CrossRef]
- Oguiura, N.; Ferrarezzi, H.; Batistic, R.F. Cytogenetics and molecular data in snakes: A phylogenetic approach. Cytogenet. Genome Res. 2009, 127, 128–142. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Altmanová, M.; Johnson Pokorná, M.; Velenský, P.; Sanchez Baca, A.; Kratochvíl, L. Evolution of karyotypes in chameleons. Genes 2017, 8, 382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorman, G. The Chromosomes of the Reptilia, a Cytotaxonomic Interpretation. In Cytotaxonomy and Vertebrate Evolution; Chiarelli, A.B., Capanna, E., Eds.; Academic Press: Cambridge, MA, USA, 1973. [Google Scholar]
- Gao, J.; Li, Q.; Wang, Z.; Zhou, Y.; Martelli, P.; Li, F.; Xiong, Z.; Wang, J.; Yang, H.; Zhang, G. Sequencing, de novo assembling, and annotating the genome of the endangered Chinese crocodile lizard Shinisaurus crocodilurus. GigaScience 2017, 6, gix041. [Google Scholar] [CrossRef] [Green Version]
- Nanda, I.; Schrama, D.; Feichtinger, W.; Haaf, T.; Schartl, M.; Schmid, M. Distribution of telomeric (TTAGGG)n sequences in avian chromosomes. Chromosoma 2002, 111, 215–227. [Google Scholar] [CrossRef]
- Mazzoleni, S.; Augstenová, B.; Clemente, L.; Auer, M.; Fritz, U.; Praschag, P.; Protiva, T.; Velenský, P.; Kratochvíl, L.; Rovatsos, M. Sex is determined by XX/XY sex chromosomes in Australasian side-necked turtles (Testudines: Chelidae). Sci. Rep. 2020, 10, 4276. [Google Scholar] [CrossRef] [Green Version]
- Clemente, L.; Mazzoleni, S.; Pensabene Bellavia, E.; Augstenová, B.; Auer, M.; Praschag, P.; Protiva, T.; Velenský, P.; Wagner, P.; Fritz, U.; et al. Interstitial telomeric repeats are rare in turtles. Genes 2020, 11, 657. [Google Scholar] [CrossRef]
- Rovatsos, M.; Pokorná, M.J.; Kratochvíl, L. Differentiation of sex chromosomes and karyotype characterisation in the dragon snake Xenodermus javanicus (Squamata: Xenodermatidae). Cytogenet. Genome Res. 2015, 147, 48–54. [Google Scholar] [CrossRef]
- Burt, D.W. Origin and evolution of avian microchromosomes. Cytogenet. Genome Res. 2002, 96, 97–112. [Google Scholar] [CrossRef]
- International Chicken Genome Sequencing Consortium. Sequence and comparative analysis of the chicken genome provide unique perspectives on vertebrate evolution. Nature 2004, 432, 695–716. [Google Scholar] [CrossRef]
- Backström, N.; Forstmeier, W.; Schielzeth, H.; Mellenius, H.; Nam, K.; Bolund, E.; Webster, M.T.; Öst, T.; Schneider, M.; Kempenaers, B.; et al. The recombination landscape of the zebra finch Taeniopygia guttata genome. Genome Res. 2010, 20, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Melek, M.; Shippen, D.E. Chromosome healing: Spontaneous and programmed de novo telomere formation by telomerase. Bioessays 1996, 18, 301–308. [Google Scholar] [CrossRef]
- Shay, J.R.; Wright, W.E. Telomeres and telomerase: Three decades of progress. Nat. Rev. Genet. 2019, 20, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Bolzán, A.D. Interstitial telomeric sequences in vertebrate chromosomes: Origin, function, instability and evolution. Mutat. Res. 2017, 773, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Bolzán, A.D.; Bianchi, M.S. Telomeres, interstitial telomeric repeat sequences, and chromosomal aberrations. Mutat. Res. 2006, 612, 189–214. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Herrera, A.; Nergadze, S.G.; Santagostino, M.; Giulotto, E. Telomeric repeats far from the ends: Mechanisms of origin and role in evolution. Cytogenet. Genome Res. 2008, 122, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Birchler, J.A.; Presting, G.G. Retrotransposon insertion targeting: A mechanism for homogenization of centromere sequences on nonhomologous chromosomes. Genes. Dev. 2012, 26, 638–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rovatsos, M.T.; Marchal, J.A.; Romero-Fernández, I.; Fernández, F.J.; Giagia-Athanosopoulou, E.B.; Sánchez, A. Rapid, independent, and extensive amplification of telomeric repeats in pericen-tromeric regions in karyotypes of arvicoline rodents. Chromosome Res. 2011, 19, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Rovatsos, M.; Marchal, J.A.; Giagia-Athanasopoulou, E.; Sánchez, A. Molecular composition of heterochromatin and its contribution to chromosome variation in Microtus thomasi/Microtus atticus species complex. Genes 2021, 12, 807. [Google Scholar] [CrossRef] [PubMed]
- Augstenová, B.; Mazzoleni, S.; Kostmann, A.; Altmanová, M.; Frynta, D.; Kratochvíl, L.; Rovatsos, M. Cytogenetic analysis did not reveal differentiated sex chromosomes in ten species of boas and pythons (Reptilia: Serpentes). Genes 2019, 10, 934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gvoždík, V.; Benkovský, N.; Crottini, A.; Bellati, A.; Moravec, J.; Romano, A.; Sacchi, R.; Jandzik, D. An ancient lineage of slow worms, genus Anguis (Squamata: Anguidae), survived in the Italian Peninsula. Mol. Phylogenet. Evol. 2013, 69, 1077–1092. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.-H.; Wiens, J.J. Comparing macroecological patterns across continents: Evolution of climatic niche breadth in varanid lizards. Ecography 2017, 40, 960–970. [Google Scholar] [CrossRef]
- De Oca, A.N.M.; Barley, A.J.; Meza-Lázaro, R.N.; García-Vázquez, U.O.; Zamora-Abrego, J.G.; Thomson, R.C.; Leaché, A.D. Phylogenomics and species delimitation in the knob-scaled lizards of the genus Xenosaurus (Squamata: Xenosauridae) using ddRADseq data reveal a substantial underestimation of diversity. Mol. Phylogenet. Evol. 2017, 106, 241–253. [Google Scholar] [CrossRef]
- Brennan, I.G.; Lemmon, A.R.; Lemmon, E.M.; Portik, D.M.; Weijola, V.; Welton, L.; Donnellan, S.C.; Keogh, J.S. Phylogenomics of monitor lizards and the role of competition in dictating body size disparity. Syst. Biol. 2021, 70, 120–132. [Google Scholar] [CrossRef]
- Porter, C.A.; Hamilton, M.J.; Sites, J.W., Jr.; Baker, R.J. Location of ribosomal DNA in chromosomes of squamate reptiles: Systematic and evolutionary implications. Herpetologica 1991, 47, 271–280. [Google Scholar]
- Stults, D.M.; Killen, M.W.; Pierce, H.H.; Pierce, A.J. Genomic architecture and inheritance of human ribosomal RNA gene clusters. Genome Res. 2008, 18, 13–18. [Google Scholar] [CrossRef] [Green Version]
- Altmanová, M.; Rovatsos, M.; Kratochvíl, L.; Johnson Pokorná, M. Minute Y chromosomes and karyotype evolution in Madagascan iguanas (Squamata: Iguania: Opluridae). Biol. J. Linn. Soc. 2016, 118, 618–633. [Google Scholar] [CrossRef]
- Sochorová, J.; Garcia, S.; Gálvez, F.; Symonová, R.; Kovařík, A. Evolutionary trends in animal ribosomal DNA loci: Introduction to a new online database. Chromosoma 2017, 127, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Mazzoleni, S.; Rovatsos, M.; Schillaci, O.; Dumas, F. Evolutionary insight on localization of 18S, 28S rDNA genes on homologous chromosomes in Primates genomes. Comp. Cytogenet. 2018, 12, 27–40. [Google Scholar] [CrossRef] [Green Version]
- Micolino, R.; Cristiano, M.P.; Travenzoli, N.M.; Lopes, D.M.; Cardoso, D.C. Chromosomal dynamics in space and time: Evolutionary history of Mycetophylax ants across past climatic changes in the Brazilian Atlantic coast. Sci. Rep. 2019, 9, 18800. [Google Scholar] [CrossRef] [PubMed]
- Degrandi, T.M.; Gunski, R.J.; Garnero, A.D.V.; Oliveira, E.H.C.; Kretschmer, R.; Souza, M.S. Barcellos, S.A.; Hass, I. The distribution of 45S rDNA sites in bird chromosomes suggests multiple evolutionary histories. Genet. Mol. Biol. 2020, 43, e20180331. [Google Scholar] [CrossRef]
- Literman, R.; Badenhorst, D.; Valenzuela, N. qPCR-based molecular sexing by copy number variation in r RNA genes and its utility for sex identification in soft-shell turtles. Methods Ecol. Evol. 2014, 5, 872–880. [Google Scholar] [CrossRef] [Green Version]
- Literman, R.; Radhakrishnan, S.; Tamplin, J.; Burke, R.; Dresser, C.; Valenzuela, N. Development of sexing primers in Glyptemys insculpta and Apalone spinifera turtles uncovers an XX/XY sex-determining system in the critically-endangered bog turtle Glyptemys muhlenbergii. Conserv. Genet. Resour. 2017, 9, 651–658. [Google Scholar] [CrossRef]
- Rovatsos, M.; Praschag, P.; Fritz, U.; Kratochvíl, L. Stable Cretaceous sex chromosomes enable molecular sexing in softshell turtles (Testudines: Trionychidae). Sci. Rep. 2017, 7, 42150. [Google Scholar] [CrossRef] [PubMed]
- Kostmann, A.; Kratochvíl, L.; Rovatsos, M. First report of sex chromosomes in plated lizards (Squamata: Gerrhosauridae). Sex. Dev. 2021, 1–6. [Google Scholar] [CrossRef]
- Zurita, F.; Sánchez, A.; Burgos, M.; Jiménez, R.; de la Guardia, R.D. Interchromosomal, intercellular and interindividual variability of NORs studied with silver staining and in situ hybridization. Heredity 1997, 78, 229–234. [Google Scholar] [CrossRef]
- Zurita, F.; Jimenez, R.; Burgos, M.; de La Guardia, R.D. Sequential silver staining and in situ hybridization reveal a direct association between rDNA levels and the expression of homologous nucleolar organizing regions: A hypothesis for NOR structure and function. J. Cell Sci. 1998, 111, 1433–1439. [Google Scholar] [CrossRef]
- Nirchio, M.; Oliveira, C.; Ferreira, I.A.; Granado, A.; Ron, E. Extensive polymorphism and chromosomal characteristics of ribosomal DNA in the characid fish Triportheus venezuelensis (Characiformes, Characidae). Genet. Mol. Biol. 2007, 30, 25–30. [Google Scholar] [CrossRef]
- Miller, L.; Knowland, J. Reduction of ribosomal RNA synthesis and ribosomal RNA genes in a mutant of Xenopus laevis which organizes only a partial nucleolus: II. The number of ribosomal RNA genes in animals of different nucleolar types. J. Mol. Biol. 1970, 53, 329–338. [Google Scholar] [CrossRef]
- Gibbons, J.G.; Branco, A.T.; Godinho, S.A.; Yu, S.; Lemos, B. Concerted copy number variation balances ribosomal DNA dosage in human and mouse genomes. Proc. Natl. Acad. Sci. USA 2015, 112, 2485–2490. [Google Scholar] [CrossRef] [Green Version]
- Xu, B.; Li, H.; Perry, J.M.; Singh, V.P.; Unruh, J.; Yu, Z.; Zakari, M.; McDowell, W.; Li, L.; Gerton, J.L. Ribosomal DNA copy number loss and sequence variation in cancer. PLoS Genet. 2017, 13, e1006771. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T. How does genome instability affect lifespan? Roles of rDNA and telomeres. Genes Cells 2011, 16, 617–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rovatsos, M.; Augstenová, B.; Altmanová, M.; Sloboda, M.; Kodym, P.; Kratochvíl, L. Triploid colubrid snake provides insight into the mechanism of sex determination in advanced snakes. Sex. Dev. 2018, 12, 251–255. [Google Scholar] [CrossRef]
- Tiersch, T.R.; Figiel, C.R. A triploid snake. Copeia 1991, 1991, 838–841. [Google Scholar] [CrossRef]
- Peters, G. Die intragenerischen Gruppen und die Phylogenese der Schmetterlingsagamen (Agamidae: Leiolepis). Zool. Jahrb. Syst. 1971, 98, 11–130. [Google Scholar]
- Moritz, C. Parthenogenesis in the endemic Australian lizard Heteronotia binoei (Gekkonidae). Science 1983, 220, 735–737. [Google Scholar] [CrossRef]
- Moritz, C.; Case, T.J.; Bolger, D.T.; Donnellan, S. Genetic diversity and the history of pacific island house geckos (Hemidactylus and Lepidodactylus). Biol. J. Linn. Soc. 1993, 48, 113–133. [Google Scholar] [CrossRef]
- Darevsky, I.S.; Kupriyanova, L.A.; Roshchin, V.V. A new all-female triploid species of gecko and karyological data on the bisexual Hemidactylus frenatus from Vietnam. J. Herpetol. 1984, 18, 277–284. [Google Scholar] [CrossRef]
- Wynn, A.H.; Cole, C.J.; Gardner, A.L. Apparent triploidy in the unisexual Brahminy blind snake, Ramphotyphlops braminus. Am. Mus. Novit. 1987, 2868, 1–7. [Google Scholar]
- Adams, M.; Foster, R.; Hutchinson, M.N.; Hutchinson, R.G.; Donnellan, S.C. The Australian scincid lizard Menetia greyii: A new instance of widespread vertebrate parthenogenesis. Evolution 2003, 57, 2619–2627. [Google Scholar] [CrossRef] [PubMed]
- Lutes, A.A.; Baumann, D.P.; Neaves, W.B.; Baumann, P. Laboratory synthesis of an independently reproducing vertebrate species. Proc. Natl. Acad. Sci. USA 2011, 108, 9910–9915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vergun, A.A.; Martirosyan, I.A.; Semyenova, S.K.; Omelchenko, A.V.; Petrosyan, V.G.; Lazebny, O.E.; Tokarskaya, O.N.; Korchagin, V.I.; Ryskov, A.P. Clonal diversity and clone formation in the parthenogenetic Caucasian rock lizard Darevskia dahli. PLoS ONE 2014, 9, e91674. [Google Scholar] [CrossRef] [Green Version]
- Trifonov, V.A.; Paoletti, A.; Caputo Barucchi, V.; Kalinina, T.; O’Brien, P.C.M.; Ferguson-Smith, M.A.; Giovannotti, M. Comparative chromosome painting and NOR distribution suggest a complex hybrid origin of triploid Lepidodactylus lugubris (Gekkonidae). PLoS ONE 2015, 10, e0132380. [Google Scholar] [CrossRef]











Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Augstenová, B.; Pensabene, E.; Kratochvíl, L.; Rovatsos, M. Cytogenetic Evidence for Sex Chromosomes and Karyotype Evolution in Anguimorphan Lizards. Cells 2021, 10, 1612. https://doi.org/10.3390/cells10071612
Augstenová B, Pensabene E, Kratochvíl L, Rovatsos M. Cytogenetic Evidence for Sex Chromosomes and Karyotype Evolution in Anguimorphan Lizards. Cells. 2021; 10(7):1612. https://doi.org/10.3390/cells10071612
Chicago/Turabian StyleAugstenová, Barbora, Eleonora Pensabene, Lukáš Kratochvíl, and Michail Rovatsos. 2021. "Cytogenetic Evidence for Sex Chromosomes and Karyotype Evolution in Anguimorphan Lizards" Cells 10, no. 7: 1612. https://doi.org/10.3390/cells10071612