
Differences in Homomorphic Sex Chromosomes Are Associated with Population Divergence in Sex Determination in Carinascincus ocellatus (Scincidae: Lygosominae)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Blood Culture and Metaphase Chromosome Preparations
2.3. Development of Sex-Linked Probe Set for Chromosome Mapping
2.4. C-Banding
2.5. Fluorescence In Situ Hybridisation (FISH) with Microsatellite Motif, Telomere and Y-Linked Probe Set
2.6. Microscopy and Image Analysis
2.7. Marker Homology
3. Results
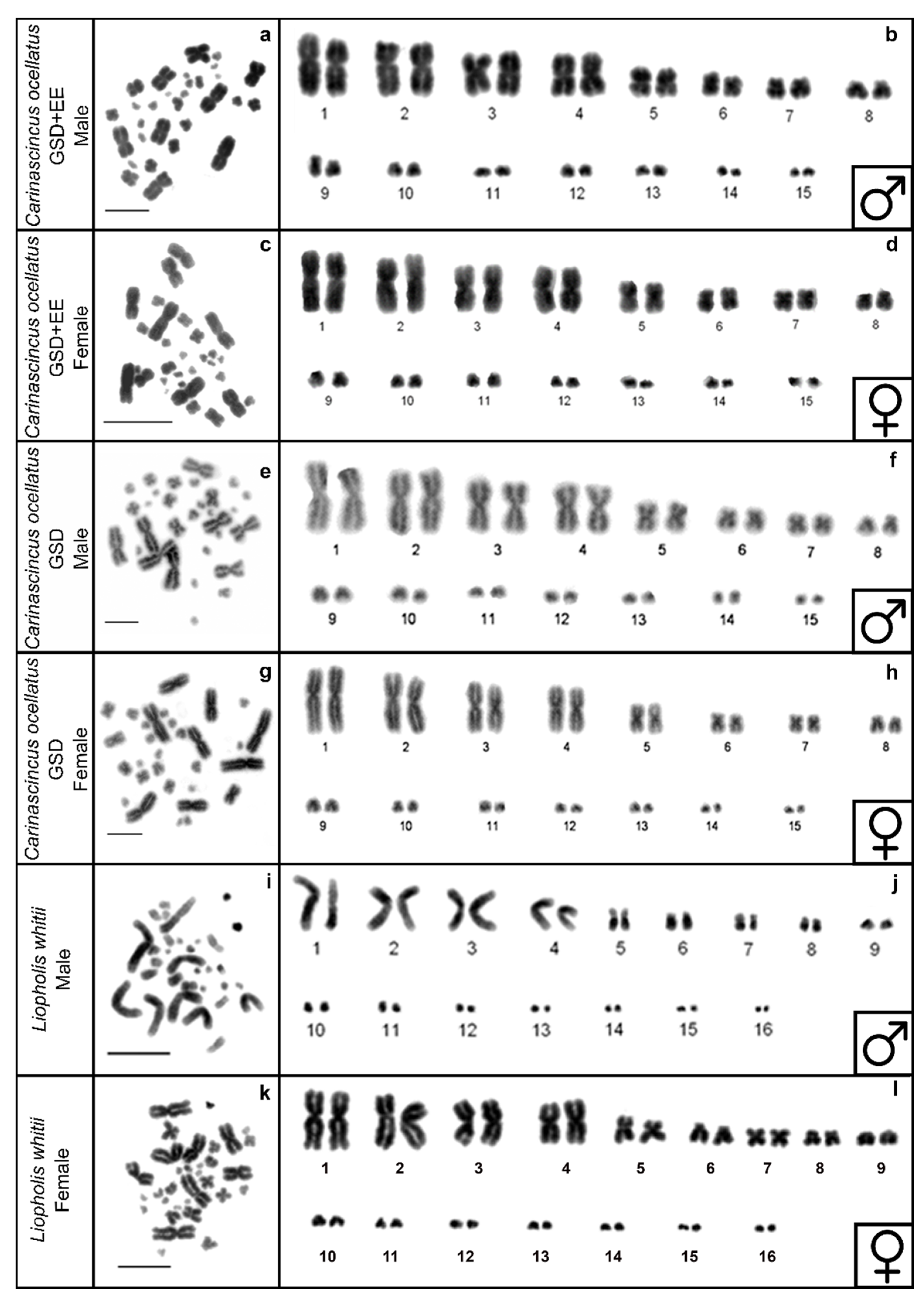
3.1. DAPI Karyotypes
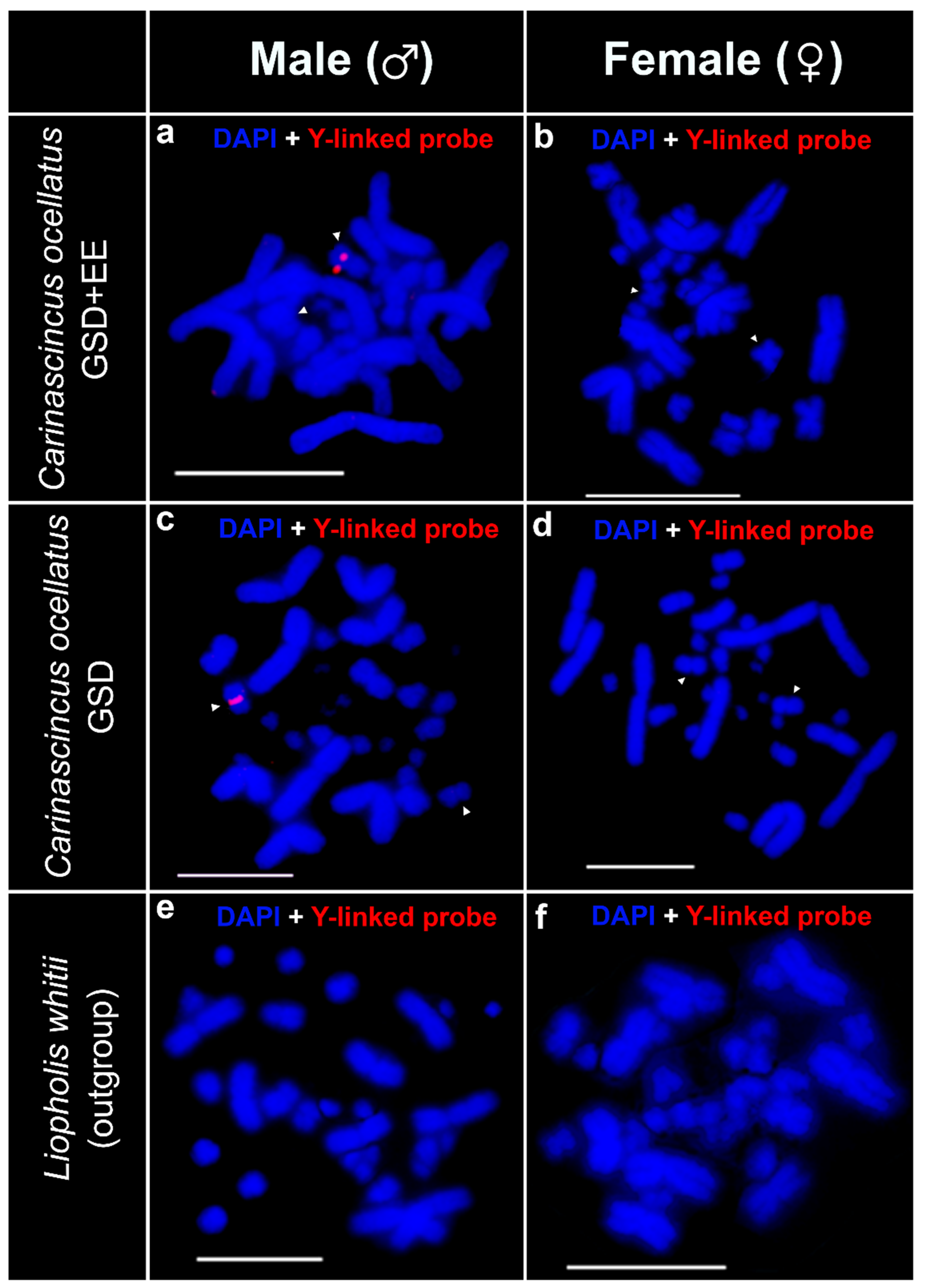
3.2. Custom Y-Linked C. ocellatus Probe Set
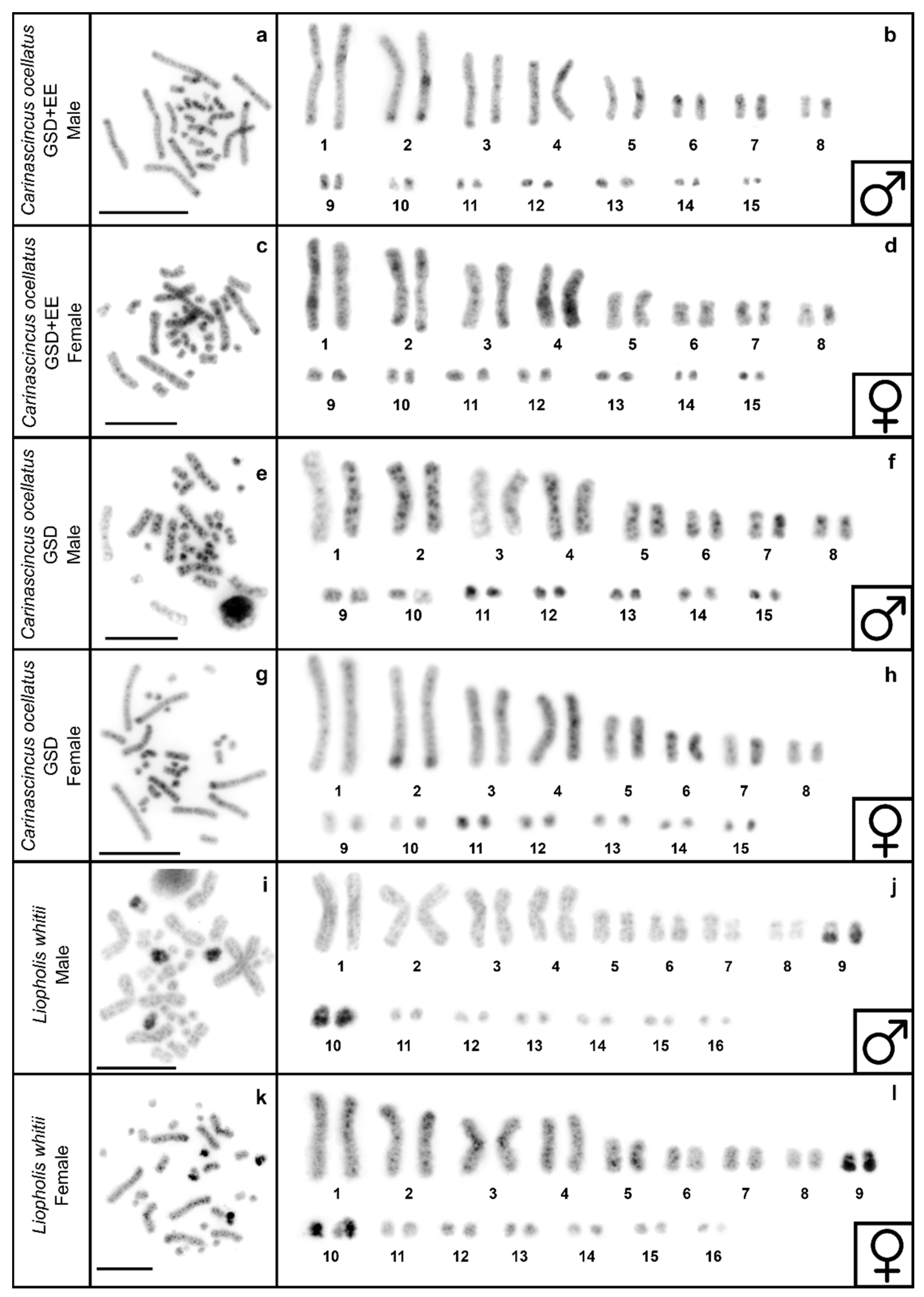
3.3. C-Banding
3.4. Telomere Repeats
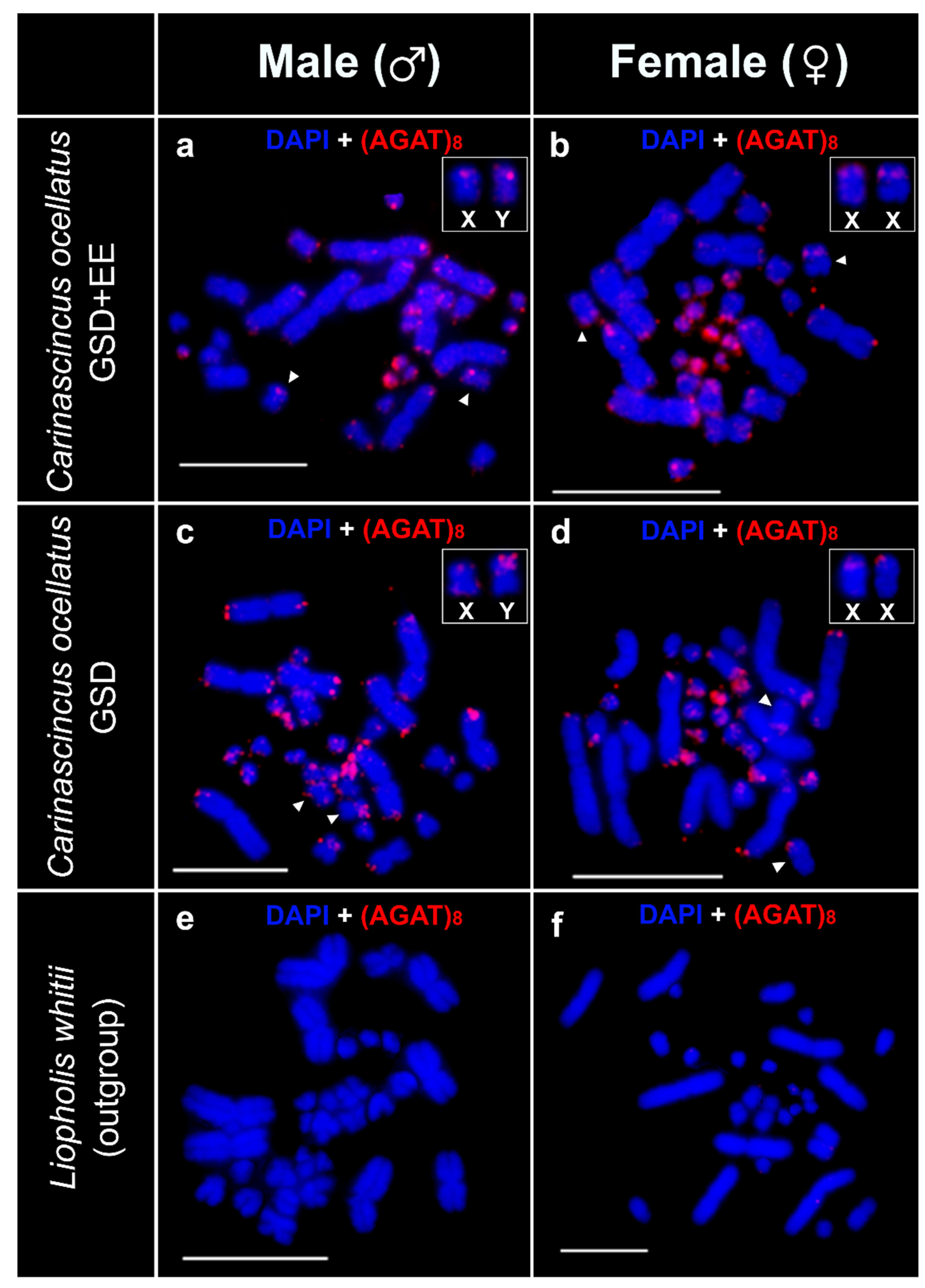
3.5. Microsatellite Motif (AGAT)8 Mapping
3.6. Sequential FISH of (AGAT)8 and Custom Y-Linked Probe Set
3.7. Marker Homology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bachtrog, D.; Mank, J.E.; Peichel, C.L.; Kirkpatrick, M.; Otto, S.P.; Ashman, T.L.; Hahn, M.W.; Kitano, J.; Mayrose, I.; Ming, R.; et al. Sex determination: Why so many ways of doing it? PLoS Biol. 2014, 12, e1001899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uller, T.; Pen, I.; Wapstra, E.; Beukeboom, L.W.; Komdeur, J. The evolution of sex ratios and sex-determining systems. Trends Ecol. Evol. 2007, 22, 292–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ezaz, T.; Stiglec, R.; Veyrunes, F.; Graves, J.A. Relationships between vertebrate ZW and XY sex chromosome systems. Curr. Biol. 2006, 16, R736–R743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capel, B. Vertebrate sex determination: Evolutionary plasticity of a fundamental switch. Nat. Rev. Genet. 2017, 18, 675–689. [Google Scholar] [CrossRef] [PubMed]
- Pennell, M.W.; Mank, J.E.; Peichel, C.L. Transitions in sex determination and sex chromosomes across vertebrate species. Mol. Ecol. 2018, 27, 3950–3963. [Google Scholar] [CrossRef] [Green Version]
- Janzen, F.J.; Phillips, P.C. Exploring the evolution of environmental sex determination, especially in reptiles. J. Evol. Biol. 2006, 19, 1775–1784. [Google Scholar] [CrossRef]
- Pokorna, M.; Kratochvil, L. Phylogeny of sex-determining mechanisms in squamate reptiles: Are sex chromosomes an evolutionary trap? Zool. J. Linn. Soc. 2009, 156, 168–183. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, D.; Mank, J.E. The birds and the bees and the flowers and the trees: Lessons from genetic mapping of sex determination in plants and animals. Genetics 2010, 186, 9–31. [Google Scholar] [CrossRef] [Green Version]
- Sigeman, H.; Ponnikas, S.; Chauhan, P.; Dierickx, E.; Brooke, M.D.; Hansson, B. Repeated sex chromosome evolution in vertebrates supported by expanded avian sex chromosomes. Proc. R. Soc. B Biol. Sci. 2019, 286. [Google Scholar] [CrossRef]
- Ezaz, T.; Deakin, J.E. Repetitive sequence and sex chromosome evolution in vertebrates. Adv. Evol. Biol. 2014, 2014, 9. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, B. The evolution of sex chromosomes. Science 1991, 251, 1030–1033. [Google Scholar] [CrossRef]
- Hooper, D.M.; Griffith, S.C.; Price, T.D. Sex chromosome inversions enforce reproductive isolation across an avian hybrid zone. Mol. Ecol. 2019, 28, 1246–1262. [Google Scholar] [CrossRef] [PubMed]
- Payseur, B.A.; Presgraves, D.C.; Filatov, D.A. Sex chromosomes and speciation. Mol. Ecol. 2018, 27, 3745–3748. [Google Scholar] [CrossRef] [PubMed]
- Coyne, J.A. “Two Rules of Speciation” revisited. Mol. Ecol. 2018, 27, 3749–3752. [Google Scholar] [CrossRef] [PubMed]
- Haldane, J.B.S. Sex ratio and unisexual sterility in hybrid animals. J. Genet. 1922, 12, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Johnson, N.A.; Lachance, J. The genetics of sex chromosomes: Evolution and implications for hybrid incompatibility. Ann. N. Y. Acad. Sci. 2012, 1256, E1–E22. [Google Scholar] [CrossRef] [Green Version]
- Mank, J.E. Small but mighty: The evolutionary dynamics of W and Y sex chromosomes. Chromosome Res. 2012, 20, 21–33. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, D.; Charlesworth, B.; Marais, G. Steps in the evolution of heteromorphic sex chromosomes. Heredity 2005, 95, 118–128. [Google Scholar] [CrossRef] [Green Version]
- Bachtrog, D. Y-chromosome evolution: Emerging insights into processes of Y-chromosome degeneration. Nat. Rev. Genet. 2013, 14, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Reichwald, K.; Petzold, A.; Koch, P.; Downie, B.R.; Hartmann, N.; Pietsch, S.; Baumgart, M.; Chalopin, D.; Felder, M.; Bens, M.; et al. Insights into sex chromosome evolution and aging from the genome of a short-lived fish. Cell 2015, 163, 1527–1538. [Google Scholar] [CrossRef] [Green Version]
- Furman, B.L.S.; Metzger, D.C.H.; Darolti, I.; Wright, A.E.; Sandkam, B.A.; Almeida, P.; Shu, J.J.; Mank, J.E. Sex chromosome evolution: So many exceptions to the rules. Genome Biol. Evol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ezaz, T.; Sarre, S.D.; O’Meally, D.; Graves, J.A.; Georges, A. Sex chromosome evolution in lizards: Independent origins and rapid transitions. Cytogenet. Genome Res. 2009, 127, 249–260. [Google Scholar] [CrossRef] [PubMed]
- Graves, J.A. Sex chromosome specialization and degeneration in mammals. Cell 2006, 124, 901–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graves, J.A. Avian sex, sex chromosomes, and dosage compensation in the age of genomics. Chromosome Res. 2014, 22, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Vicoso, B.; Kaiser, V.B.; Bachtrog, D. Sex-biased gene expression at homomorphic sex chromosomes in emus and its implication for sex chromosome evolution. Proc. Natl. Acad. Sci. USA 2013, 110, 6453–6458. [Google Scholar] [CrossRef] [Green Version]
- Bracewell, R.R.; Bentz, B.J.; Sullivan, B.T.; Good, J.M. Rapid neo-sex chromosome evolution and incipient speciation in a major forest pest. Nat. Commun. 2017, 8, 1593. [Google Scholar] [CrossRef] [Green Version]
- Lande, R.; Seehausen, O.; van Alphen, J.J. Mechanisms of rapid sympatric speciation by sex reversal and sexual selection in cichlid fish. Genetica 2001, 112–113, 435–443. [Google Scholar] [CrossRef]
- Ming, R.; Bendahmane, A.; Renner, S.S. Sex chromosomes in land plants. Annu. Rev. Plant. Biol 2011, 62, 485–514. [Google Scholar] [CrossRef] [Green Version]
- Pennell, M.W.; Kirkpatrick, M.; Otto, S.P.; Vamosi, J.C.; Peichel, C.L.; Valenzuela, N.; Kitano, J. Y fuse? Sex chromosome fusions in fishes and reptiles. PLoS Genet. 2015, 11, e1005237. [Google Scholar] [CrossRef] [Green Version]
- Waters, P.D.; Duffy, B.; Frost, C.J.; Delbridge, M.L.; Graves, J.A. The human Y chromosome derives largely from a single autosomal region added to the sex chromosomes 80-130 million years ago. Cytogenet. Cell Genet. 2001, 92, 74–79. [Google Scholar] [CrossRef]
- Meisel, R.P. Evolution of sex determination and sex chromosomes: A novel alternative paradigm. Bioessays News Rev. Mol. Cell. Dev. Biol. 2020, 42, 1900212. [Google Scholar] [CrossRef] [PubMed]
- Quinn, A.E.; Sarre, S.D.; Ezaz, T.; Graves, J.A.; Georges, A. Evolutionary transitions between mechanisms of sex determination in vertebrates. Biol. Lett. 2011, 7, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Holleley, C.E.; O’Meally, D.; Sarre, S.D.; Graves, J.A.; Ezaz, T.; Matsubara, K.; Azad, B.; Zhang, X.; Georges, A. Sex reversal triggers the rapid transition from genetic to temperature-dependent sex. Nature 2015, 523, 79–82. [Google Scholar] [CrossRef]
- Gamble, T.; Coryell, J.; Ezaz, T.; Lynch, J.; Scantlebury, D.P.; Zarkower, D. Restriction site-associated DNA sequencing (RAD-seq) reveals an extraordinary number of transitions among gecko sex-determining systems. Mol. Biol. Evol. 2015, 32, 1296–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornejo-Paramo, P.; Dissanayake, D.S.B.; Lira-Noriega, A.; Martinez-Pacheco, M.L.; Acosta, A.; Ramirez-Suastegui, C.; Mendez-de-la-Cruz, F.R.; Szekely, T.; Urrutia, A.O.; Georges, A.; et al. Viviparous reptile regarded to have temperature-dependent sex determination has old XY chromosomes. Genome Biol. Evol. 2020, 12, 924–930. [Google Scholar] [CrossRef]
- Dissanayake, D.S.B.; Holleley, C.E.; Hill, L.; O’Meally, D.; Deakin, J.; Georges, A. Identification of Y chromosome markers in the eastern three-lined skink (Bassiana duperreyi) using in-silico whole genome subtraction. BMC Genom. 2020, 21, 667. [Google Scholar] [CrossRef]
- Rovatsos, M.; Vukic, J.; Kratochvil, L. Mammalian X homolog acts as sex chromosome in lacertid lizards. Heredity 2016, 117, 8–13. [Google Scholar] [CrossRef]
- Rovatsos, M.; Altmanova, M.; Pokorna, M.; Kratochvil, L. Conserved sex chromosomes across adaptively radiated Anolis lizards. Evolution 2014, 68, 2079–2085. [Google Scholar] [CrossRef]
- Iannucci, A.; Altmanova, M.; Ciofi, C.; Ferguson-Smith, M.; Milan, M.; Pereira, J.C.; Pether, J.; Rehak, I.; Rovatsos, M.; Stanyon, R.; et al. Conserved sex chromosomes and karyotype evolution in monitor lizards (Varanidae). Heredity 2019, 123, 215–227. [Google Scholar] [CrossRef]
- Patawang, I.; Chuaynkern, Y.; Supanuam, P.; Maneechot, N.; Pinthong, K.; Tanomtong, A. Cytogenetics of the skinks (Reptilia, Scincidae) from Thailand; IV: Newly investigated karyotypic features of Lygosoma quadrupes and Scincella melanosticta. Caryologia 2018, 71, 29–34. [Google Scholar] [CrossRef]
- Donnellan, S.C. Chromosomes of Australian lygosomine skinks (Lacertilia: Scincidae) I the Egernia group. Genetica 1991, 83, 207–222. [Google Scholar] [CrossRef]
- Donnellan, S.C. Chromosomes of Australian lygosomine skinks (Lacertilia: Scincidae) II The genus Lampropholis. Genetica 1991, 83, 223–234. [Google Scholar] [CrossRef]
- Olmo, E.; Signorino, G. Chromorep: A Reptiles Chromosomes Database. 2005. Available online: https://www.semanticscholar.org/paper/Chromorep%3A-A-reptiles-chromosomes-database-Olmo-Sgnorino/1effb2cb3d047b791266cab434ffb26af0bd357c (accessed on 20 October 2020).
- Shine, R.; Elphick, M.J.; Donnellan, S. Co-occurrence of multiple, supposedly incompatible modes of sex determination in a lizard population. Ecol. Lett. 2002, 5, 486–489. [Google Scholar] [CrossRef]
- Radder, R.S.; Quinn, A.E.; Georges, A.; Sarre, S.D.; Shine, R. Genetic evidence for co-occurrence of chromosomal and thermal sex-determining systems in a lizard. Biol. Lett. 2008, 4, 176–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wapstra, E.; Olsson, M.; Shine, R.; Edwards, A.; Swain, R.; Joss, J.M. Maternal basking behaviour determines offspring sex in a viviparous reptile. Biol. Lett. 2004, 271, S230–S232. [Google Scholar] [CrossRef] [Green Version]
- Quinn, A.E.; Georges, A.; Sarre, S.D.; Guarino, F.; Ezaz, T.; Graves, J.A. Temperature sex reversal implies sex gene dosage in a reptile. Science 2007, 316, 411. [Google Scholar] [CrossRef] [Green Version]
- Hill, P.L.; Burridge, C.P.; Ezaz, T.; Wapstra, E. Conservation of sex-linked markers among conspecific populations of a viviparous skink, Niveoscincus ocellatus, exhibiting genetic and temperature-dependent sex determination. Genome Biol. Evol. 2018, 10, 1079–1087. [Google Scholar] [CrossRef] [Green Version]
- Robert, K.A.; Thompson, M.B. Sex determination. Viviparous lizard selects sex of embryos. Nature 2001, 412, 698–699. [Google Scholar] [CrossRef]
- Wapstra, E.; Uller, T.; Sinn, D.L.; Olsson, M.; Mazurek, K.; Joss, J.; Shine, R. Climate effects on offspring sex ratio in a viviparous lizard. J. Anim. Ecol. 2009, 78, 84–90. [Google Scholar] [CrossRef]
- Cunningham, G.D.; While, G.M.; Wapstra, E. Climate and sex ratio variation in a viviparous lizard. Biol. Lett. 2017, 13. [Google Scholar] [CrossRef]
- Pen, I.; Uller, T.; Feldmeyer, B.; Harts, A.; While, G.M.; Wapstra, E. Climate-driven population divergence in sex-determining systems. Nature 2010, 468, 436–438. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, N.; Adams, D.C.; Janzen, F.J. Pattern does not equal process: Exactly when is sex environmentally determined? Am. Nat. 2003, 161, 676–683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnellan, S.C. The Evolution of Sex Chromosomes in Scincid Lizards. Ph.D. Thesis, Macquarie University, Sydney, Australia, 1985. [Google Scholar]
- Cliff, H.B.; Wapstra, E.; Burridge, C.P. Persistence and dispersal in a Southern Hemisphere glaciated landscape: The phylogeography of the spotted snow skink (Niveoscincus ocellatus) in Tasmania. BMC Evol. Biol. 2015, 15, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A resource for timelines, timetrees, and divergence times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef]
- Matsubara, K.; O’Meally, D.; Azad, B.; Georges, A.; Sarre, S.D.; Graves, J.A.; Matsuda, Y.; Ezaz, T. Amplification of microsatellite repeat motifs is associated with the evolutionary differentiation and heterochromatinization of sex chromosomes in Sauropsida. Chromosoma 2016, 125, 111–123. [Google Scholar] [CrossRef]
- Matsubara, K.; Uno, Y.; Srikulnath, K.; Matsuda, Y.; Miller, E.; Olsson, M. No interstitial telomeres on autosomes but remarkable amplification of telomeric repeats on the W sex chromosome in the sand lizard (Lacerta agilis). J. Hered. 2015, 106, 753–757. [Google Scholar] [CrossRef] [Green Version]
- Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol 2013, 13, 93. [Google Scholar] [CrossRef] [Green Version]
- Wapstra, E.; Swain, R.; Jones, S.M.; O’Reilly, J. Geographic and annual variation in reproductive cycles in the Tasmanian spotted snow skink, Niveoscincus ocellatus (Squamata: Scincidae). Aust. J. Zool. 1999, 47, 539–550. [Google Scholar] [CrossRef]
- Ezaz, T.; Quinn, A.E.; Miura, I.; Sarre, S.D.; Georges, A.; Graves, J.A. The dragon lizard Pogona vitticeps has ZZ/ZW micro-sex chromosomes. Chromosome Res. 2005, 13, 763–776. [Google Scholar] [CrossRef]
- Kilian, A.; Wenzl, P.; Huttner, E.; Carling, J.; Xia, L.; Blois, H.; Caig, V.; Heller-Uszynska, K.; Jaccoud, D.; Hopper, C.; et al. Diversity Arrays Technology: A generic genome profiling technology on open platforms. In Data Production and Analysis in Population Genomics: Methods and Protocols; Pompanon, F., Bonin, A., Eds.; Humana Press: Totowa, NJ, USA, 2012; pp. 67–89. [Google Scholar] [CrossRef]
- Sumner, A.T. A simple technique for demonstrating centromeric heterochromatin. Exp. Cell Res. 1972, 75, 304–306. [Google Scholar] [CrossRef]
- Shams, F.; Dyer, F.; Thompson, R.; Duncan, R.P.; Thiem, J.D.; Majtánová, Z.; Ezaz, T. Karyotypes and sex chromosomes in two Australian native freshwater fishes, golden perch (Macquaria ambigua) and Murray cod (Maccullochella peelii) (Percichthyidae). Int. J. Mol. Sci. 2019, 20, 4244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsubara, K.; Knopp, T.; Sarre, S.D.; Georges, A.; Ezaz, T. Karyotypic analysis and FISH mapping of microsatellite motifs reveal highly differentiated XX/XY sex chromosomes in the pink-tailed worm-lizard (Aprasia parapulchella, Pygopodidae, Squamata). Mol. Cytogenet 2013, 6, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madden, T. The BLAST sequence analysis tool. In The NCBI Handbook, 2nd ed.; McEntyre, J., Ostell, J., Eds.; National Center for Biotechnology Information (US): Bethesda, MD, USA, 2013. [Google Scholar]
- Piskurek, O.; Austin, C.C.; Okada, N. Sauria SINEs: Novel short interspersed retroposable elements that are widespread in reptile genomes. J. Mol. Evol. 2006, 62, 630–644. [Google Scholar] [CrossRef] [PubMed]
- Hill, P.L.; Ezaz, T.; Wapstra, E.; Burridge, C.P. Pleistocene divergence in the absence of gene flow among populations of a viviparous reptile with intraspecific variation in sex determination. Ecol. Evol. 2021. in Press. [Google Scholar]
- Hutchinson, M.N.; Donnellan, S.C. Taxonomy and genetic variation in the Australian lizards of the genus Pseudemoia (Scincidae: Lygosominae). J. Nat. Hist. 1992, 26, 215–264. [Google Scholar] [CrossRef]
- Ezaz, T.; Srikulnath, K.; Graves, J.A. Origin of Amniote sex chromosomes: An ancestral super-sex chromosome, or common requirements? J. Hered. 2017, 108, 94–105. [Google Scholar] [CrossRef]
- Holmquist, G.; Dancis, B.M. A general model of karyotype evolution. Genetica 1984, 52, 151–163. [Google Scholar] [CrossRef]
- Srikulnath, K.; Azad, B.; Singchat, W.; Ezaz, T. Distribution and amplification of interstitial telomeric sequences (ITSs) in Australian dragon lizards support frequent chromosome fusions in Iguania. PLoS ONE 2019, 14, e0212683. [Google Scholar] [CrossRef] [Green Version]
- Alam, S.M.I.; Altmanová, M.; Prasongmaneerut, T.; Georges, A.; Sarre, S.D.; Nielsen, S.V.; Gamble, T.; Srikulnath, K.; Rovatsos, M.; Kratochvíl, L.; et al. Cross-species BAC mapping highlights conservation of chromosome synteny across dragon lizards (Squamata: Agamidae). Genes 2020, 11, 698. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GSD+EE C. ocellatus | GSD C. ocellatus | L. whitii | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Male | Female | Male | Female | Male | Female | |||||||
| Ind. | Cells | Ind. | Cells | Ind. | Cells | Ind. | Cells | Ind. | Cells | Ind. | Cells | |
| Karyotyping | 3 | 73 | 3 | 50 | 3 | 129 | 3 | 64 | 3 | 43 | 3 | 67 |
| c-banding | 1 | 4 | 2 | 6 | 2 | 18 | 2 | 31 | 1 | 3 | 1 | 7 |
| FISH | ||||||||||||
| Telomere | 3 | 15 | 2 | 20 | 2 | 36 | 2 | 18 | 3 | 17 | 3 | 24 |
| AGAT | 2 | 28 | 2 | 14 | 2 | 43 | 2 | 28 | 2 | 14 | 2 | 24 |
| Y-linked | 3 | 19 | 2 | 16 | 2 | 33 | 3 | 18 | 3 | 12 | 3 | 19 |
| Sequential | 1 | 11 | 1 | 17 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hill, P.; Shams, F.; Burridge, C.P.; Wapstra, E.; Ezaz, T. Differences in Homomorphic Sex Chromosomes Are Associated with Population Divergence in Sex Determination in Carinascincus ocellatus (Scincidae: Lygosominae). Cells 2021, 10, 291. https://doi.org/10.3390/cells10020291
Hill P, Shams F, Burridge CP, Wapstra E, Ezaz T. Differences in Homomorphic Sex Chromosomes Are Associated with Population Divergence in Sex Determination in Carinascincus ocellatus (Scincidae: Lygosominae). Cells. 2021; 10(2):291. https://doi.org/10.3390/cells10020291
Chicago/Turabian StyleHill, Peta, Foyez Shams, Christopher P. Burridge, Erik Wapstra, and Tariq Ezaz. 2021. "Differences in Homomorphic Sex Chromosomes Are Associated with Population Divergence in Sex Determination in Carinascincus ocellatus (Scincidae: Lygosominae)" Cells 10, no. 2: 291. https://doi.org/10.3390/cells10020291