
Effects of Fluctuating Thermal Regimes and Pesticides on Egg Hatching of a Natural Enemy Harmonia axyridis (Coleoptera Coccinellidae)

Abstract
:1. Introduction
2. Materials and Methods
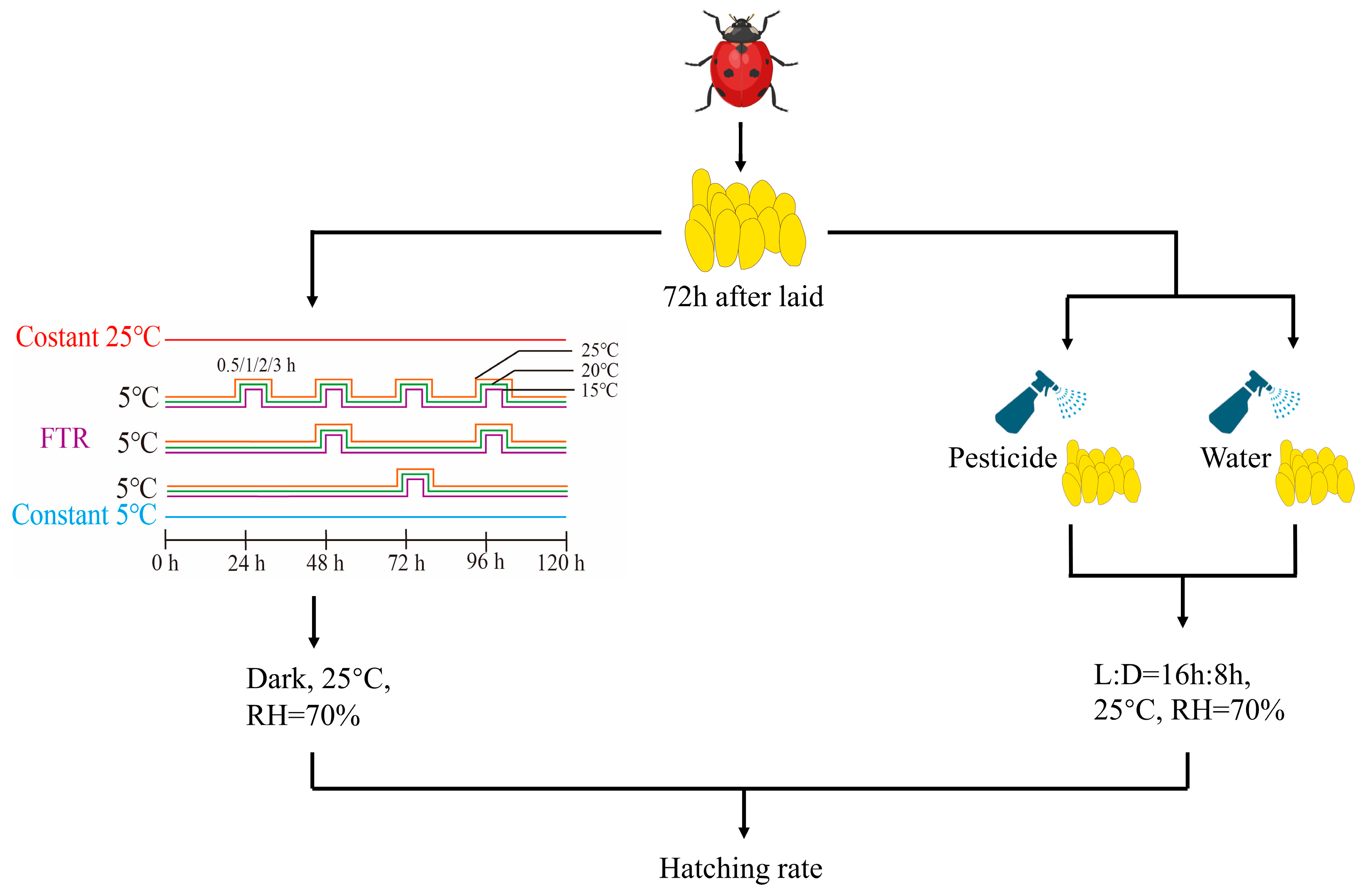
2.1. Insect Rearing and Egg Collection
2.2. Effects of Fluctuating Thermal Regimes on Egg Hatching Rate
2.3. Effects of Pesticides on Egg Hatching Rate
2.4. Statistical Analysis
3. Results
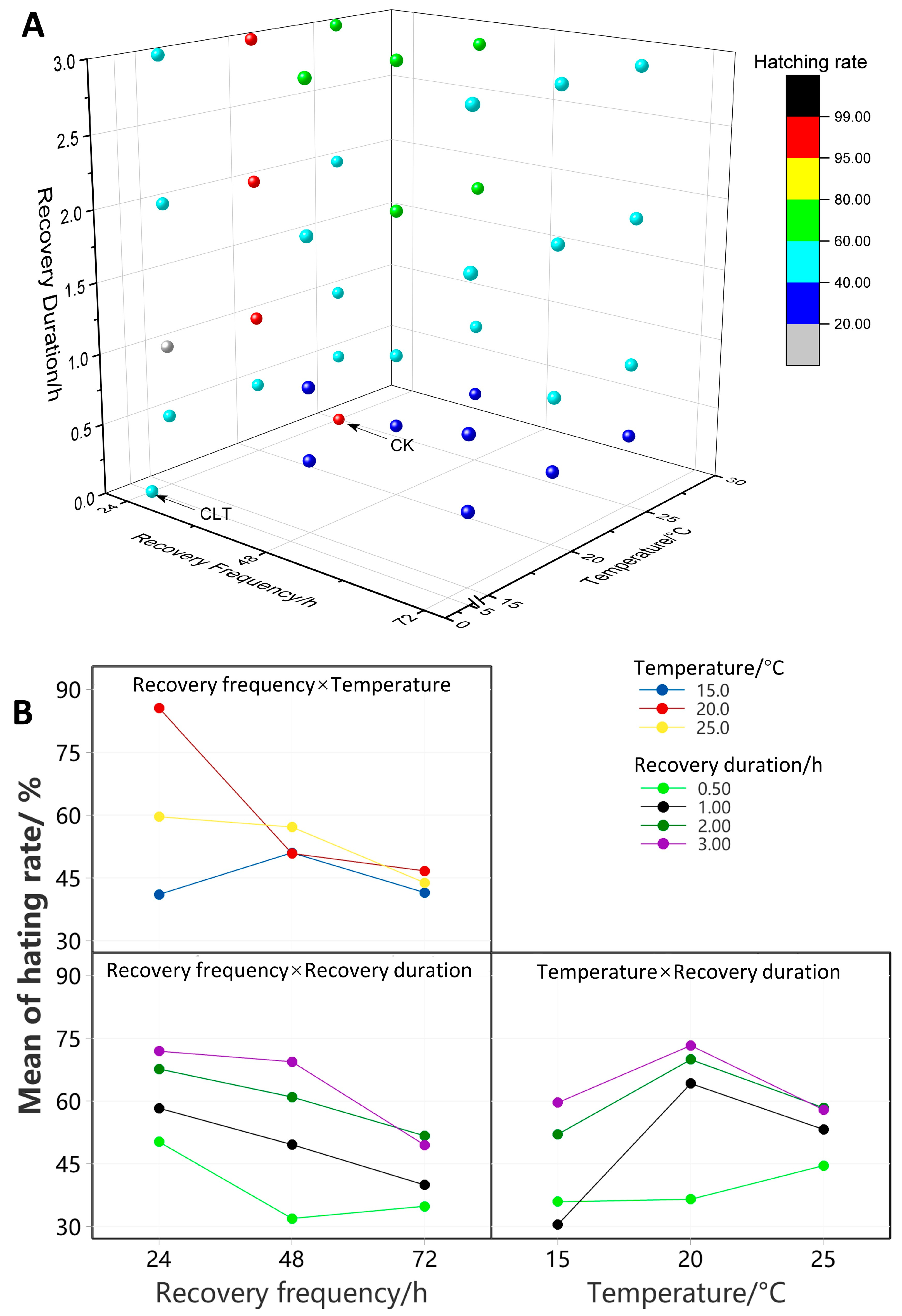
3.1. The Influence of Fluctuating Thermal Regimes on the Hatching Rate of Eggs
3.2. Effects of Seven Pesticides on the Hatching Rate of H. axyridis
4. Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, Y.; Wang, H.; Dang, L.M.; Sadeghi-Niaraki, A.; Moon, H. Crop pest recognition in natural scenes using convolutional neural networks. Comput. Electron. Agric. 2020, 169, 105174. [Google Scholar] [CrossRef]
- Gangola, S.; Bhatt, P.; Kumar, A.J.; Bhandari, G.; Joshi, S.; Punetha, A.; Bhatt, K.; Rene, E.R. Biotechnological tools to elucidate the mechanism of pesticide degradation in the environment. Chemosphere 2022, 296, 133916. [Google Scholar] [CrossRef]
- Helepciuc, F.E.; Todor, A. EU microbial pest control: A revolution in waiting. Pest Manag. Sci. 2022, 78, 1314–1325. [Google Scholar] [CrossRef]
- Wu, Y.; Xu, L.; Chang, L.; Ma, M.; You, L.; Jiang, C.; Li, S.; Zhang, J. Bacillus thuringiensis cry1C expression from the plastid genome of poplar leads to high mortality of leaf-eating caterpillars. Tree Physiol. 2019, 39, 1525–1532. [Google Scholar] [CrossRef]
- Tu, C.; Zhang, Y.; Zhu, P.; Sun, L.; Xu, P.; Wang, T.; Luo, J.; Yu, J.; Xu, L. Enhanced toxicity of entomopathogenic fungi Beauveria bassiana with bacteria expressing immune suppressive dsRNA in a leaf beetle. Pestic. Biochem. Physiol. 2023, 193, 105431. [Google Scholar] [CrossRef]
- Jetter, K.; Paine, T.D. Consumer preferences and willingness to pay for biological control in the urban landscape. Biol. Control 2004, 30, 312–322. [Google Scholar] [CrossRef]
- Camacho-Cervantes, M.; Mendoza-Arroyo, W.; Arellano-Sanchez, D.; Del-Val, E. Exotic predators are not better biocontrol agents: The harlequin ladybird is not the most voracious in Mexico. PeerJ 2021, 9, e12503. [Google Scholar] [CrossRef]
- Masetti, A.; Magagnoli, S.; Lami, F.; Lanzoni, A.; Burgio, G. Long term changes in the communities of native ladybirds in Northern Italy: Impact of the invasive species Harmonia axyridis (Pallas). BioControl 2018, 63, 665–675. [Google Scholar] [CrossRef]
- Tun, K.M.; Clavijo McCormick, A.; Jones, T.; Minor, M. The potential of harlequin ladybird beetle Harmonia axyridis as a predator of the giant willow aphid Tuberolachnus salignus: Voracity, life history and prey preference. BioControl 2020, 65, 313–321. [Google Scholar] [CrossRef]
- LaMana, M.L.; Miller, J.C. Field Observations onHarmonia axyridisPallas (Coleoptera: Coccinellidae) in Oregon. Biol. Control 1996, 6, 232–237. [Google Scholar] [CrossRef]
- Yu, W.; Liu, J. Predation and Control of Harmonia axyridis against Tinocallis kahawaluokalani. J. Zhejiang For. Sci. Technol. 2019, 39, 55–59. [Google Scholar]
- Tayeh, A.; Estoup, A.; Laugier, G.; Loiseau, A.; Turgeon, J.; Toepfer, S.; Facon, B. Evolution in biocontrol strains: Insight from the harlequin ladybird Harmonia axyridis. Evol. Appl. 2012, 5, 481–488. [Google Scholar] [CrossRef]
- Yocum, G.D.; Rajamohan, A.; Rinehart, J.P. Comparison of Fluctuating Thermal Regimes and Commercially Achievable Constant-Temperature Regimes for Short-Term Storage of the Alfalfa Leafcutting Bee (Hymenoptera: Megachilidae). J. Econ. Entomol. 2021, 114, 530–537. [Google Scholar] [CrossRef]
- Colinet, H.; Rinehart, J.P.; Yocum, G.D.; Greenlee, K.J. Mechanisms underpinning the beneficial effects of fluctuating thermal regimes in insect cold tolerance. J. Exp. Biol. 2018, 221, jeb164806. [Google Scholar] [CrossRef]
- Rinehart, J.P.; Yocum, G.D.; Kemp, W.P.; Bowsher, J.H. Optimizing Fluctuating Thermal Regime Storage of Developing Megachile rotundata (Hymenoptera: Megachilidae). J. Econ. Entomol. 2016, 109, 993–1000. [Google Scholar] [CrossRef]
- Torson, A.S.; Yocum, G.D.; Rinehart, J.P.; Nash, S.A.; Bowsher, J.H. Fluctuating thermal regimes prevent chill injury but do not change patterns of oxidative stress in the alfalfa leafcutting bee, Megachile rotundata. J. Insect Physiol. 2019, 118, 103935. [Google Scholar] [CrossRef]
- Youn, Y.N.; Seo, M.J.; Shin, J.G.; Jang, C.; Yu, Y.M. Toxicity of greenhouse pesticides to multicolored Asian lady beetles, Harmonia axyridis (Coleoptera: Coccinellidae). Biol. Control 2003, 28, 164–170. [Google Scholar] [CrossRef]
- Galvan, T.L.; Koch, R.L.; Hutchison, W.D. Toxicity of indoxacarb and spinosad to the multicolored Asian lady beetle, Harmonia axyridis (Coleoptera: Coccinellidae), via three routes of exposure. Pest Manag. Sci. 2006, 62, 797–804. [Google Scholar] [CrossRef]
- Rico, A.; Arenas-Sánchez, A.; Pasqualini, J.; García-Astillero, A.; Cherta, L.; Nozal, L.; Vighi, M. Effects of imidacloprid and a neonicotinoid mixture on aquatic invertebrate communities under Mediterranean conditions. Aquat. Toxicol. 2018, 204, 130–143. [Google Scholar] [CrossRef]
- Serrao, J.E.; Plata-Rueda, A.; Martinez, L.C.; Zanuncio, J.C. Side-effects of pesticides on non-target insects in agriculture: A mini-review. Naturwissenschaften 2022, 109, 17. [Google Scholar] [CrossRef]
- Whitehorn, P.R.; O’Connor, S.; Wackers, F.L.; Goulson, D. Neonicotinoid pesticide reduces bumble bee colony growth and queen production. Science 2012, 336, 351–352. [Google Scholar] [CrossRef] [PubMed]
- Fogel, M.N.; Schneider, M.I.; Desneux, N.; González, B.; Ronco, A.E. Impact of the neonicotinoid acetamiprid on immature stages of the predator Eriopis connexa (Coleoptera: Coccinellidae). Ecotoxicology 2013, 22, 1063–1071. [Google Scholar] [CrossRef]
- Shi, Z.; Liu, X.; Xu, Q.; Qin, Z.; Wang, S.; Zhang, F.; Wang, S.; Tang, B. Two novel soluble trehalase genes cloned from Harmonia axyridis and regulation of the enzyme in a rapid changing temperature. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2016, 198, 10–18. [Google Scholar] [CrossRef]
- Grumiaux, C.; Andersen, M.K.; Colinet, H.; Overgaard, J. Fluctuating thermal regime preserves physiological homeostasis and reproductive capacity in Drosophila suzukii. J. Insect Physiol. 2019, 113, 33–41. [Google Scholar] [CrossRef]
- Enriquez, T.; Ruel, D.; Charrier, M.; Colinet, H. Effects of fluctuating thermal regimes on cold survival and life history traits of the spotted wing Drosophila (Drosophila suzukii). Insect Sci. 2020, 27, 317–335. [Google Scholar] [CrossRef]
- Lombaert, E.; Malausa, T.; Devred, R.; Estoup, A. Phenotypic variation in invasive and biocontrol populations of the harlequin ladybird, Harmonia axyridis. In From Biological Control to Invasion: The Ladybird Harmonia axyridis as a Model Species; Roy, H.E., Wajnberg, E., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 89–102. [Google Scholar]
- Ping, S.; Jian-hua, W.; Ping, L.; Xue-yi, Y.; Hu-ying, Z.; Yong-qiang, Z.; Gang, D. Screening of insecticides for the control of Aphis gossypil and the effect on Harmonia axyridis larvae. J. Gansu Agric. Univ. 2014, 49, 96–101. [Google Scholar] [CrossRef]
- GB/T31270-2014; Test Guidelines on Environmental Safety Assessment for Chemical Pesticides―Part 17: Trichogramma Acute Toxicity Test. China, Ministry of Agriculture of the People’s Republic of China: Beijing, China,, 2014.
- Hong, Z.G.Z.J.Z.C.Y. Toxicity Determination of 7 Pesticides to Myzus Persicae (Sulzer) and Orius Similis Zheng. Chin. Tob. Sci. 2018, 39, 59–65. [Google Scholar] [CrossRef]
- Akcay, M.; Arslan, H.; Durmus, N.; Mese, M.; Capar, I.D. Dentinal tubule penetration of AH Plus, iRoot SP, MTA fillapex, and guttaflow bioseal root canal sealers after different final irrigation procedures: A confocal microscopic study. Lasers Surg. Med. 2016, 48, 70–76. [Google Scholar] [CrossRef]
- McWhinnie, R.; Sckrabulis, J.; Raffel, T. Temperature and mass scaling affect cutaneous and pulmonary respiratory performance in a diving frog. Integr. Zool. 2021, 16, 712–728. [Google Scholar] [CrossRef]
- Lemoine, N.P.; Drews, W.A.; Burkepile, D.E.; Parker, J.D. Increased temperature alters feeding behavior of a generalist herbivore. Oikos 2013, 122, 1669–1678. [Google Scholar] [CrossRef]
- Leith, N.T.; Macchiano, A.; Moore, M.P.; Fowler-Finn, K.D. Temperature impacts all behavioral interactions during insect and arachnid reproduction. Curr. Opin. Insect Sci. 2021, 45, 106–114. [Google Scholar] [CrossRef]
- Giroux, A.; Ortega, Z.; Bertassoni, A.; Desbiez, A.L.J.; Kluyber, D.; Massocato, G.F.; De Miranda, G.; Mourão, G.; Surita, L.; Attias, N.; et al. The role of environmental temperature on movement patterns of giant anteaters. Integr. Zool. 2022, 17, 285–296. [Google Scholar] [CrossRef]
- Hodek, I.; Emden, H.F.; Honěk, A. Ecology and Behaviour of the Ladybird Beetles (Coccinellidae); John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Ruan, C.-C.; Du, W.-M.; Wang, X.-M.; Zhang, J.-J.; Zang, L.-S. Effect of long-term cold storage on the fitness of pre-wintering Harmonia axyridis (Pallas). BioControl 2011, 57, 95–102. [Google Scholar] [CrossRef]
- Sweeney, B.W.; Schnack, J.A. Egg Development, Growth, and Metabolism of Sigara Alternata (Say) (Hemiptera: Corixidae) in Fluctuating Thermal Environments. Ecology 1977, 58, 265–277. [Google Scholar] [CrossRef]
- Koštál, V.; Renault, D.; Mehrabianová, A.; Bastl, J. Insect cold tolerance and repair of chill-injury at fluctuating thermal regimes: Role of ion homeostasis. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2007, 147, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Kültz, D. Molecular and evolutionary basis of the cellular stress response. Annu. Rev. Physiol. 2005, 67, 225–257. [Google Scholar] [CrossRef] [PubMed]
- Phillips, T.W.; Throne, J.E. Biorational Approaches to Managing Stored-Product Insects. Annu. Rev. Entomol. 2009, 55, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Rolim, G.D.S.; Plata-Rueda, A.; Martinez, L.C.; Ribeiro, G.T.; Serrao, J.E.; Zanuncio, J.C. Side effects of Bacillus thuringiensis on the parasitoid Palmistichus elaeisis (Hymenoptera: Eulophidae). Ecotoxicol. Environ. Saf. 2020, 189, 109978. [Google Scholar] [CrossRef]
- Barzman, M.; Bàrberi, P.; Birch, A.N.E.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P.; et al. Eight principles of integrated pest management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- Kunkel, B.A.; Held, D.W.; Potter, D.A. Lethal and sublethal effects of bendiocarb, halofenozide, and imidacloprid on Harpalus pennsylvanicus (Coleoptera: Carabidae) following different modes of exposure in turfgrass. J. Econ. Entomol. 2001, 94, 60–67. [Google Scholar] [CrossRef]
- Cabral, S.; Garcia, P.; Soares, A.O. Effects of pirimicarb, buprofezin and pymetrozine on survival, development and reproduction of Coccinella undecimpunctata (Coleoptera: Coccinellidae). Biocontrol Sci. Technol. 2008, 18, 307–318. [Google Scholar] [CrossRef]
- Yao, F.L.; Zheng, Y.; Zhao, J.W.; Desneux, N.; He, Y.X.; Weng, Q.Y. Lethal and sublethal effects of thiamethoxam on the whitefly predator Serangium japonicum (Coleoptera: Coccinellidae) through different exposure routes. Chemosphere 2015, 128, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Dai, P.; Jack, C.J.; Mortensen, A.N.; Ellis, J.D. Acute toxicity of five pesticides to Apis mellifera larvae reared in vitro. Pest Manag. Sci. 2017, 73, 2282–2286. [Google Scholar] [CrossRef]
- Bartlett, B.R. Toxicity of Some Pesticides to Eggs, Larvae, and Adults of the Green Lacewing, Chrysopa carnea1. J. Econ. Entomol. 1964, 57, 366–369. [Google Scholar] [CrossRef]
- Houbraken, M.; Spranghers, T.; De Clercq, P.; Cooreman-Algoed, M.; Couchement, T.; De Clercq, G.; Verbeke, S.; Spanoghe, P. Pesticide contamination of Tenebrio molitor (Coleoptera: Tenebrionidae) for human consumption. Food Chem. 2016, 201, 264–269. [Google Scholar] [CrossRef]
- Leopold, R. Cold Storage of Insects for Integrated Pest Management; CRC Press: Boca Raton, FL, USA, 2019; pp. 235–267. [Google Scholar]



{kind=link}
{kind=link}
| Insecticides | Formulation | Recommended Concentrations (mg/L) | Toxicity | Families | Initial Concentration (mg/L) | Final Concentration (mg/L) |
|---|---|---|---|---|---|---|
| 3% Emamectin benzoate | ME | 2000 | Low | Microbial pesticide | 125 | 8000 |
| 5% Abamectin | EW | 1000 | Moderate | Microbial pesticide | 50 | 3200 |
| 30% Thiamethoxam | SC | 666.67 | Low | Second generation nicotine | 62.5 | 4000 |
| 20% Acetamiprid | SL | 1000 | Low | Nicotine chloride compounds | 62.5 | 4000 |
| 20% Imidacloprid | SL | 1000 | Low | Neonicotinic insecticides | 84 | 5336 |
| 1.3% Matrine | EW | 1000 | Low | Alkaloid | 125 | 8000 |
| 0.3% Azadirachtin | SC | 1000 | Low | Biological insecticide | 125 | 8000 |
| Factors | df | MSE | F | p-Value |
|---|---|---|---|---|
| Recovery frequency (1) | 2 | 0.229 | 3.722 | 0.027 |
| Temperature (2) | 2 | 0.048 | 0.778 | 0.461 |
| Recovery duration (3) | 3 | 0.166 | 2.698 | 0.048 |
| Between (1) + (2) | 4 | 0.225 | 3.661 | 0.007 |
| Between (1) + (3) | 6 | 0.067 | 1.091 | 0.370 |
| Between (2) + (3) | 6 | 0.105 | 1.713 | 0.122 |
| Between (1) + (2) + (3) | 12 | 0.057 | 0.929 | 0.520 |
| Error value | 144 | 0.062 | ||
| Total | 180 |
| Recovery Temperature | Recovery Frequency | Recovery Duration | |||
|---|---|---|---|---|---|
| 0.5 h | 1 h | 2 h | 3 h | ||
| 15 °C | 24 h | 44.78 ± 7.34 | 19.69 ± 4.47 | 46.96 ± 13.80 a | 52.78 ± 20.10 |
| 48 h | 33.55 ± 4.09 Ab | 38.33 ± 10.46 AB | 56.43 ± 1.62 Bb | 75.70 ± 3.48 AB | |
| 72 h | 29.45 ± 9.53 | 33.27 ± 8.51 | 52.77 ± 7.91 a | 50.45 ± 10.54 | |
| 20 °C | 24 h | 48.86 ± 4.12 Aa | 97.49 ± 1.38 A | 97.30 ± 0.29 A | 98.41 ± 1.59 B |
| 48 h | 25.18 ± 5.64 Ab | 52.81 ± 3.33 B | 61.49 ± 9.32 AB | 63.86 ± 1.76 B | |
| 72 h | 35.51 ± 8.42 a | 42.36 ± 4.11 | 51.09 ± 9.02 | 57.70 ± 5.79 | |
| 25 °C | 24 h | 57.12 ± 3.90 A | 57.76 ± 14.60 AB | 58.85 ± 0.64 B | 64.70 ± 5.55 AB |
| 48 h | 37.07 ± 2.14 | 57.69 ± 12.31 | 65.09 ± 7.67 | 68.75 ± 4.93 | |
| 72 h | 39.59 ± 3.67 | 44.20 ± 13.15 | 51.26 ± 0.67 | 40.28 ± 15.24 | |
| Constant low temperatures | 54.29 ± 7.52 a | ||||
| Control group | 99.20 ± 0.72 b | ||||
| Insecticide | Regression Equation | LC50/mg/L | LC95/mg/L | Correlation Coefficient | Relative Safety Factor | Index of Relative Toxicity |
|---|---|---|---|---|---|---|
| 20% acetamiprid SL | Y = −1.411 + 0.009X | 158.35 | 342.89 | 0.914 | 0.16 | 67.90 |
| 20% imidacloprid SL | Y = −4.362 + 1.708X | 358.15 | 3290.17 | 0.923 | 0.36 | 30.02 |
| 5% abamectin EW | Y = −10.334 + 3.765X | 555.87 | 1520.19 | 0.948 | 0.56 | 19.34 |
| 0.3% azadirachtin SC | Y = −5.406 + 1.516X | 3679.67 | 44,744.10 | 0.971 | 3.68 | 2.92 |
| 3% abamectin-aminomethyl ME | Y = −8.773 + 2.339X | 5630.88 | 28,430.59 | 0.902 | 2.82 | 1.91 |
| 30% thiamethoxam SC | Y = −4.687 + 1.219X | 6969.76 | 155,604.83 | 0.904 | 10.45 | 1.54 |
| 1.3% matrine EW | Y = −4.870 + 1.180X | 107,52.25 | 263,162.094 | 0.911 | 10.75 | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Li, C.; Dong, L.; Mao, R.; Wang, Z.; Pei, Z.; Xu, L. Effects of Fluctuating Thermal Regimes and Pesticides on Egg Hatching of a Natural Enemy Harmonia axyridis (Coleoptera Coccinellidae). Agronomy 2023, 13, 1470. https://doi.org/10.3390/agronomy13061470
Yu J, Li C, Dong L, Mao R, Wang Z, Pei Z, Xu L. Effects of Fluctuating Thermal Regimes and Pesticides on Egg Hatching of a Natural Enemy Harmonia axyridis (Coleoptera Coccinellidae). Agronomy. 2023; 13(6):1470. https://doi.org/10.3390/agronomy13061470
Chicago/Turabian StyleYu, Jingya, Chong Li, Likun Dong, Runping Mao, Zhihua Wang, Zhangxin Pei, and Letian Xu. 2023. "Effects of Fluctuating Thermal Regimes and Pesticides on Egg Hatching of a Natural Enemy Harmonia axyridis (Coleoptera Coccinellidae)" Agronomy 13, no. 6: 1470. https://doi.org/10.3390/agronomy13061470