



Integrated Transcriptome and Metabolome Analysis Reveal That Exogenous Gibberellin Application Regulates Lignin Synthesis in Ramie
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of the External Application of Gibberellin on Growth Indicators (Plant Height and Stem Thickness) of Forage Ramie in the Field
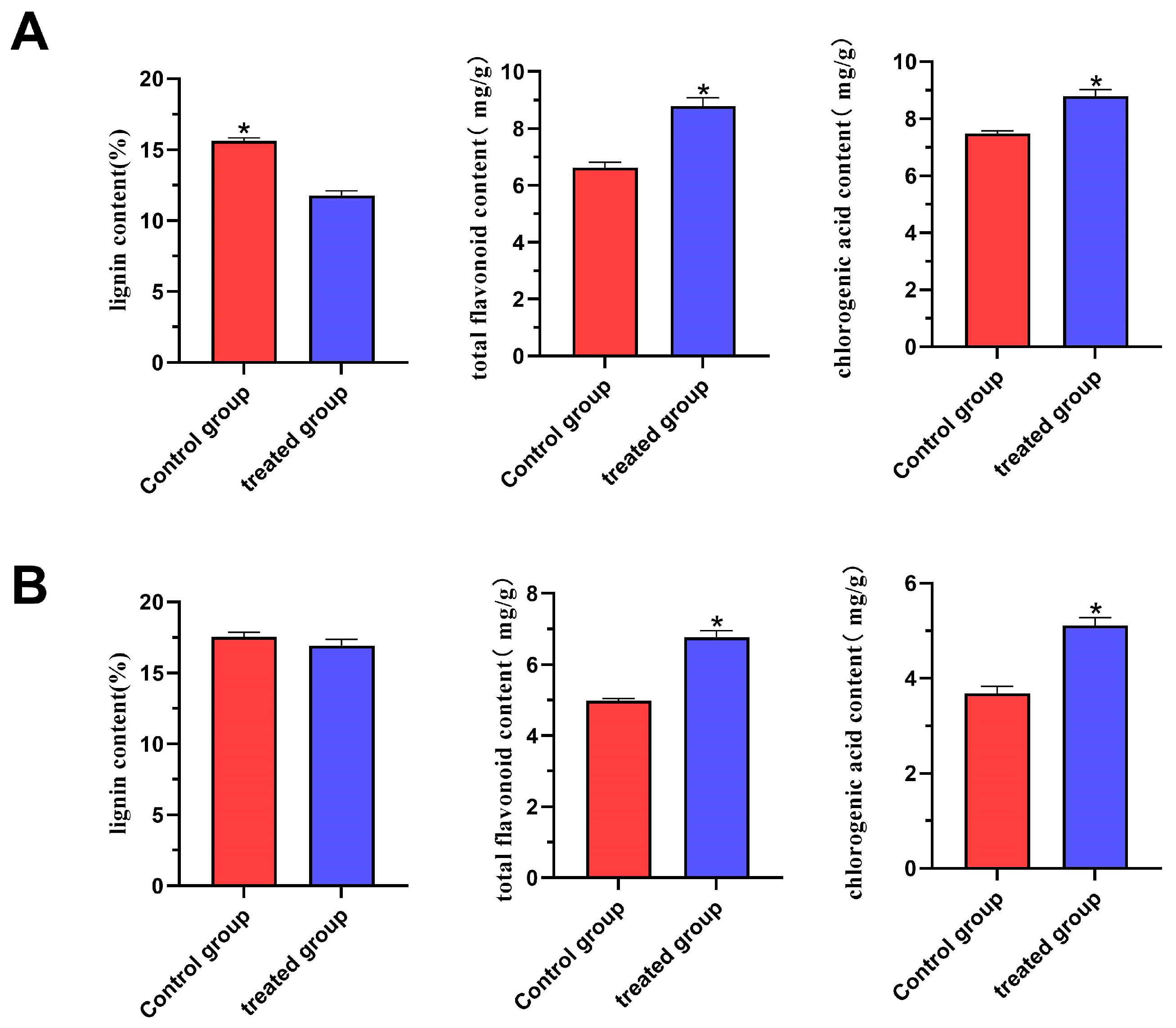
2.2. Effect of External Gibberellin Application on biochemical Parameters in Ramie Forage
2.3. Transcriptomic Analysis
2.3.1. Transcriptome Sequencing and Assembly
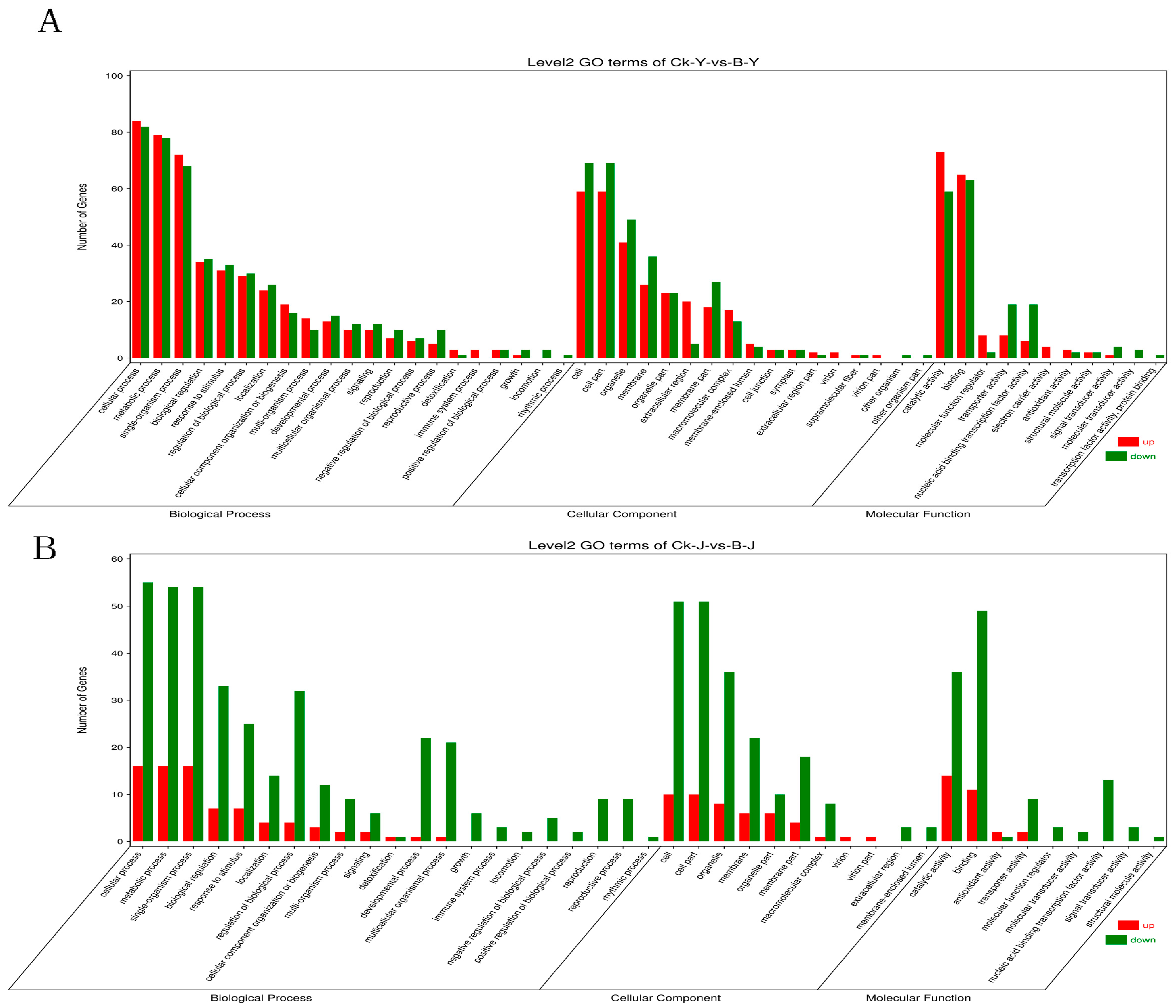
2.3.2. GO and KEGG Term Classification of DEGs
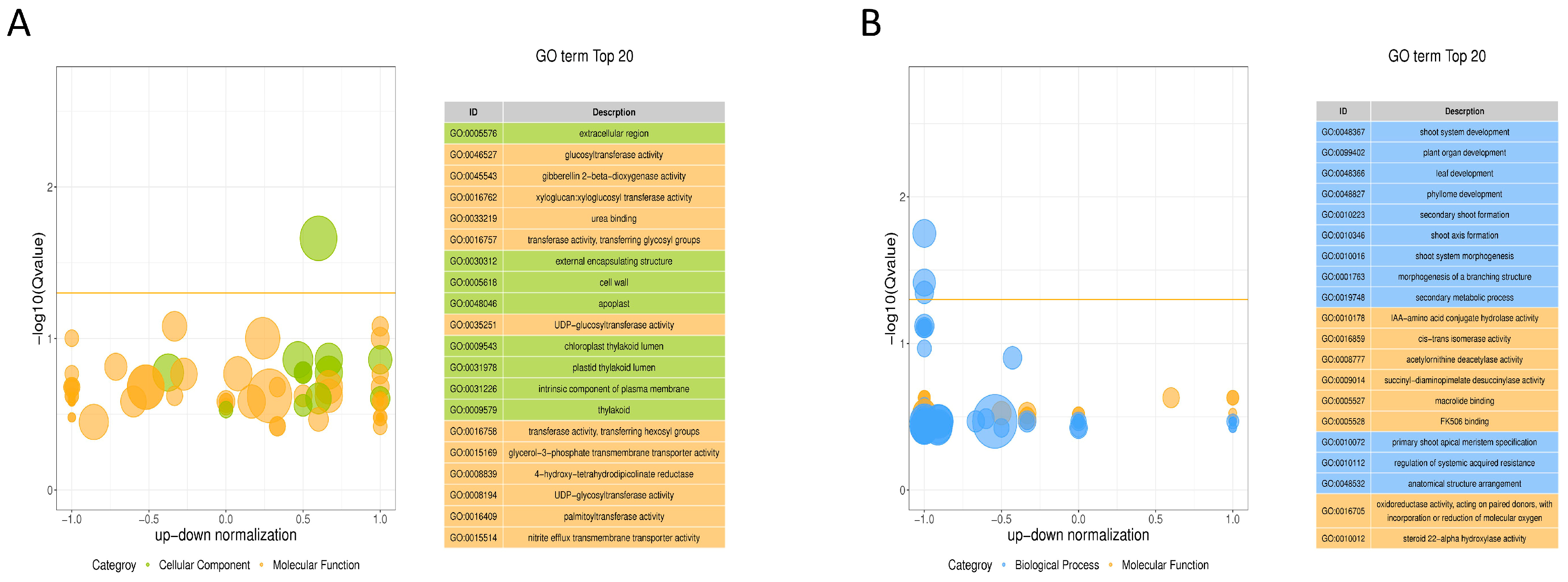
2.3.3. Enrichment Analysis of the DEG Functions in GO Annotation and KEGG Pathways
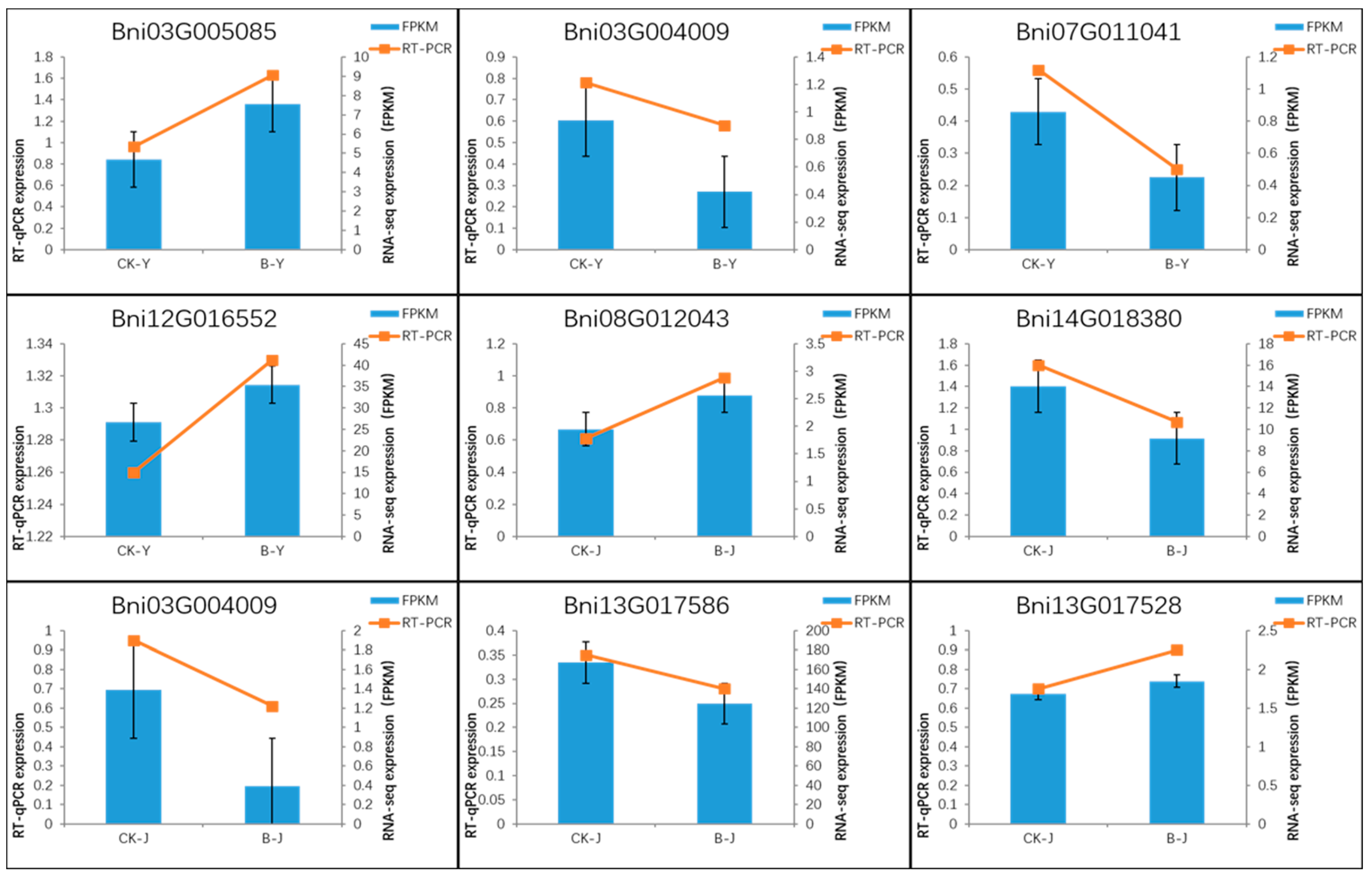
2.3.4. qRT-PCR Validation of DEGSs
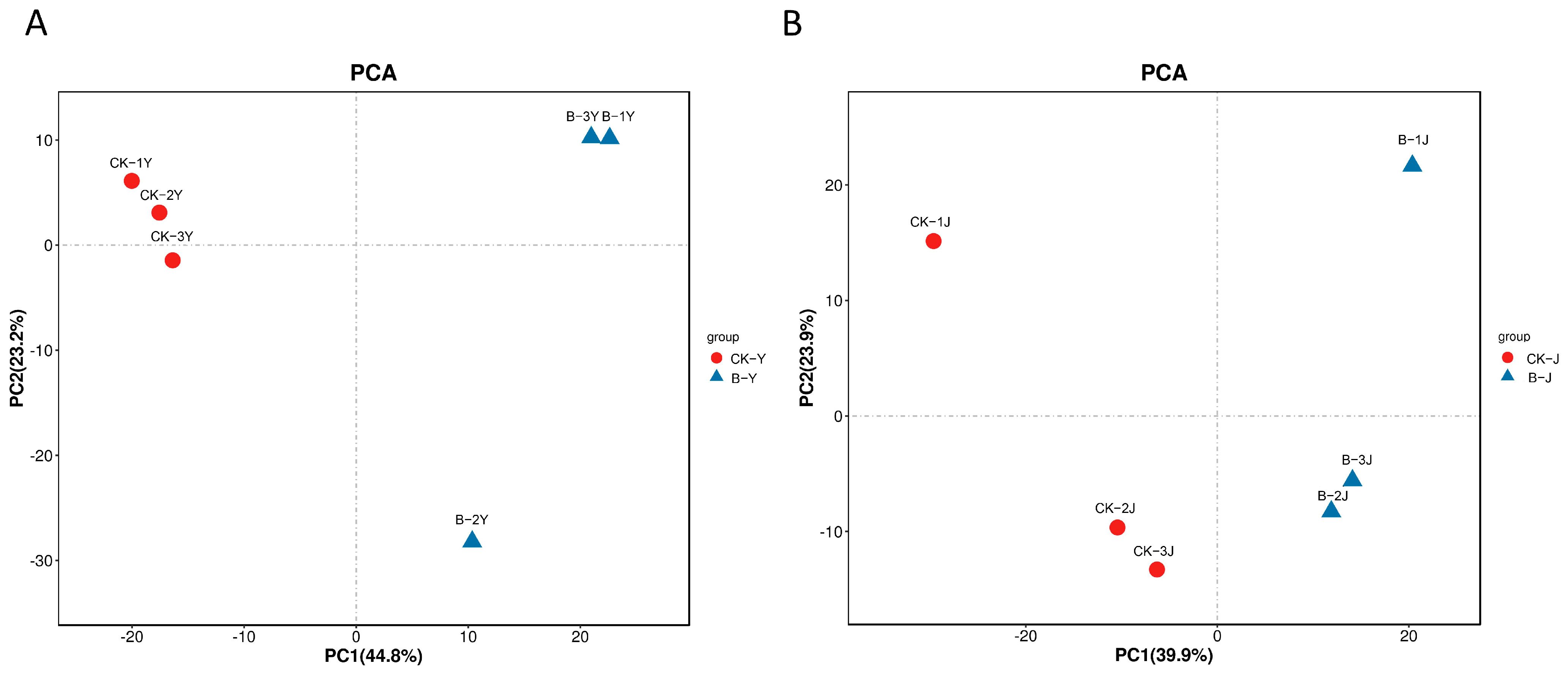
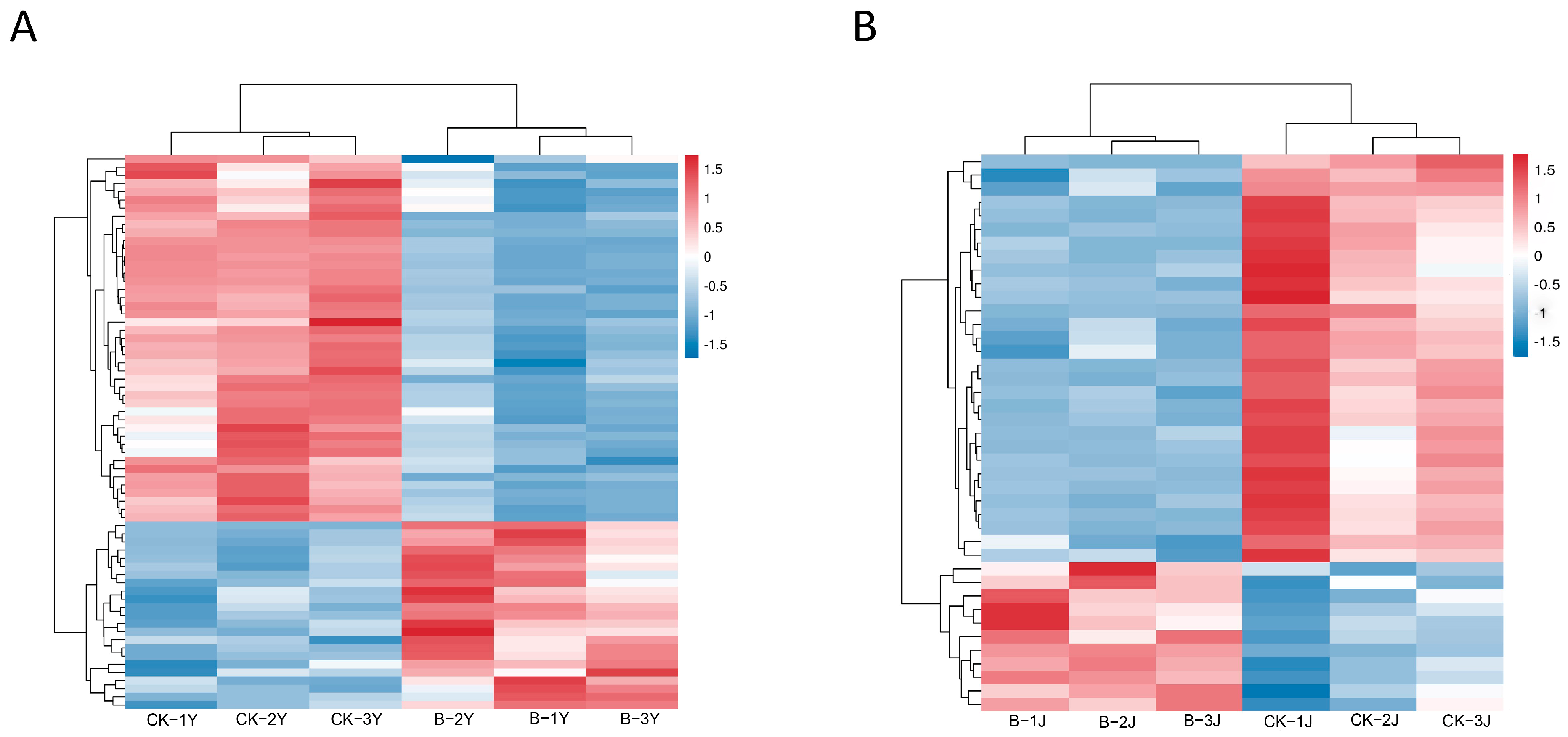
2.4. Metabolomics
2.4.1. Effects of Gibberellin on the Metabolites in Leaves and Stems of Forage Ramie
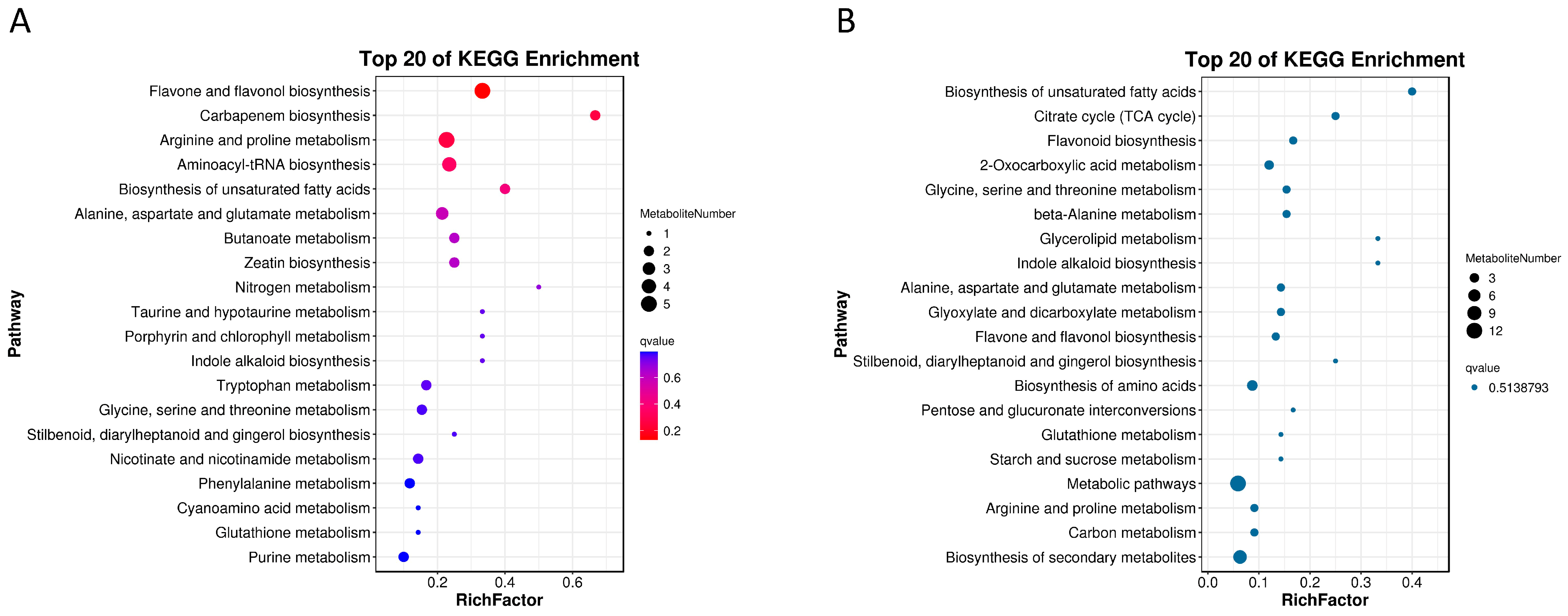
2.4.2. KEGG Pathway Enrichment Analysis of the Differential Metabolites
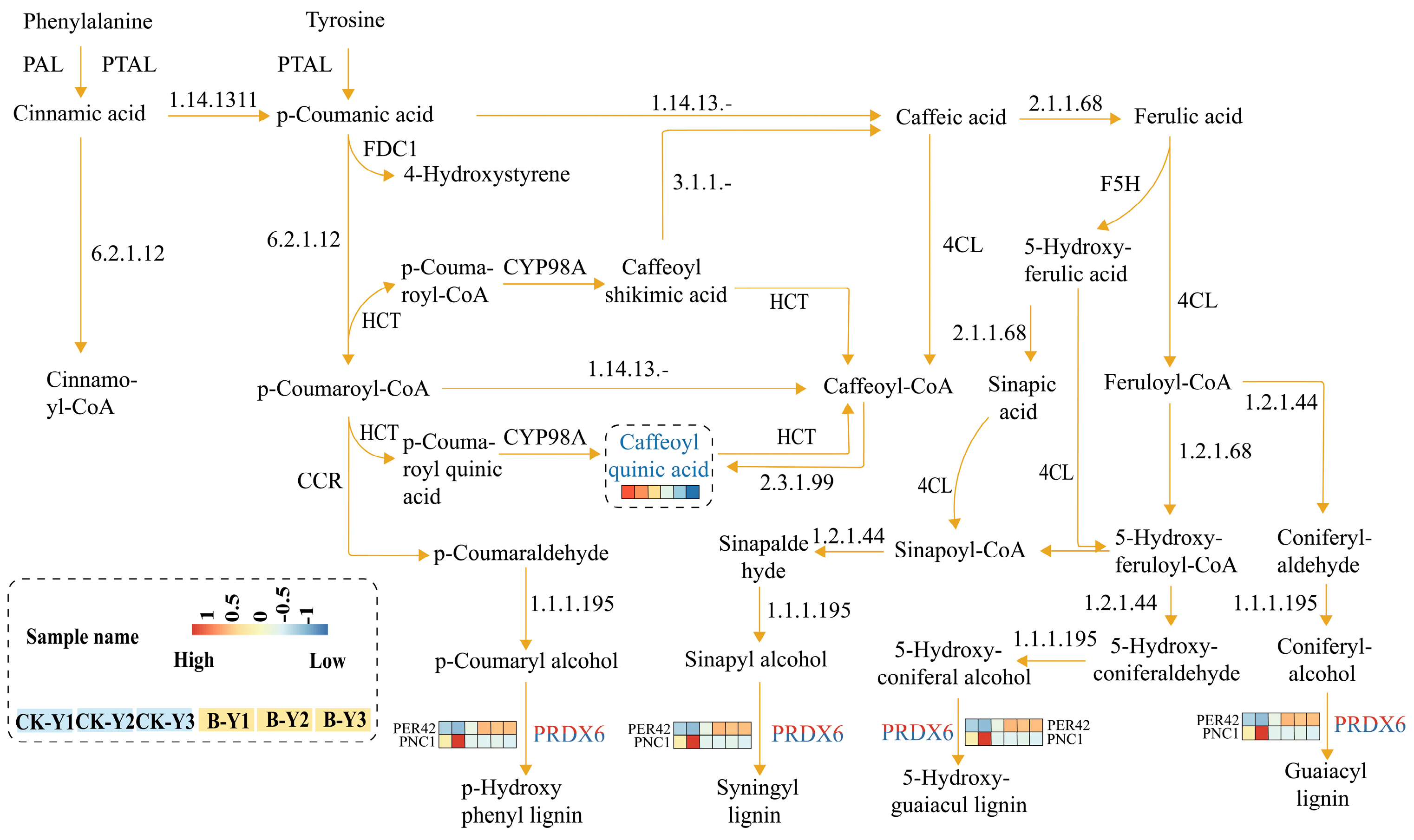
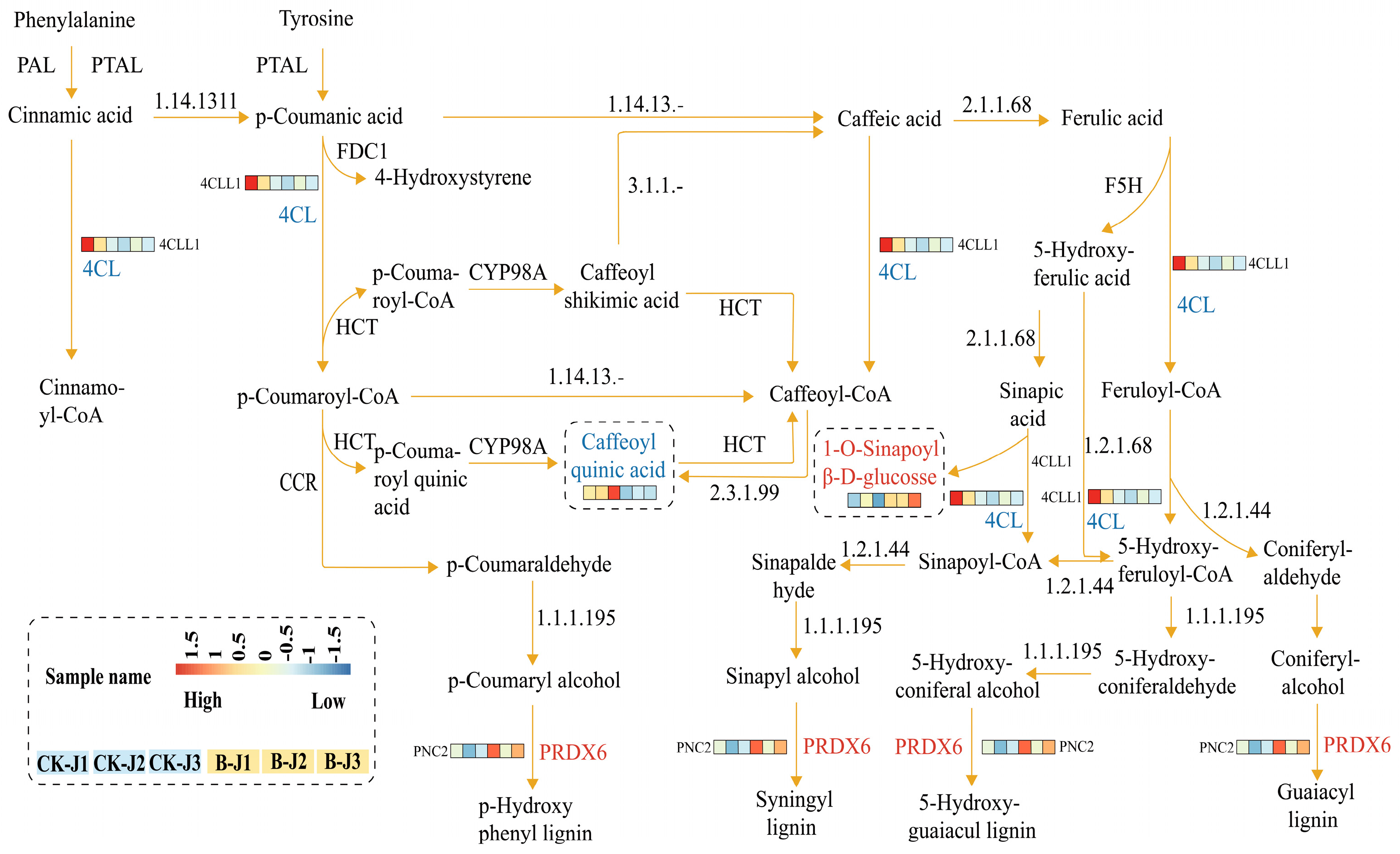
2.5. Combined Metabolomic and Transcriptomic Analysis
3. Discussion
4. Materials and Methods
4.1. Plant Material, Experimental Site, and Handling Methods
4.2. Determination of Lignin, Chlorogenic Acid, and Total Flavonoid Contents
4.2.1. Determination of Lignin Content
4.2.2. Determination of Total Flavonoids
4.2.3. Determination of Chlorogenic Acid Content
4.3. RNA Extraction, Library Construction, and Sequencing
4.4. Metabolite Analysis
4.4.1. Sample Preparation and Extraction
4.4.2. Data Preprocessing and Metabolite Identification
4.5. Validation of Real-Time quantitativePCR (qRT-PCR)
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kipriotis, E.; Heping, X.; Vafeiadakis, T.; Kiprioti, M.; Alexopoulou, E. Ramie and kenaf as feed crops. Ind. Crops Prod. 2015, 68, 126–130. [Google Scholar] [CrossRef]
- Pandey, S.N. Ramie fibre: Part I. Chemical composition and chemical properties. A critical review of recent developments. Text. Prog. 2007, 39, 1–66. [Google Scholar] [CrossRef]
- Wang, X.; Peng, S.-M.; Liu, Y.; Liao, S.; Zhao, H.-H.; Duan, G.-Y.; Wu, Y.-M.; Liu, C.-J.; Wang, Y.-Z.; Liu, T.-M.; et al. Effect of Ramie on the Production Performance of Laying Hens, and the Quality, Nutrient Composition, Antioxidation of the Eggs. Front. Physiol. 2022, 13, 854760. [Google Scholar] [CrossRef]
- Vanholme, R.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin engineering. Curr. Opin. Plant Biol. 2008, 11, 278–285. [Google Scholar] [CrossRef]
- Vásquez-Garay, F.; Carrillo-Varela, I.; Vidal, C.; Reyes-Contreras, P.; Faccini, M.; Mendonça, R.T. A Review on the Lignin Biopolymer and Its Integration in the Elaboration of Sustainable Materials. Sustainability 2021, 13, 2697. [Google Scholar] [CrossRef]
- Sellami, K.; Couvert, A.; Nasrallah, N.; Maachi, R.; Abouseoud, M.; Amrane, A. Peroxidase enzymes as green catalysts for bioremediation and biotechnological applications: A review. Sci. Total Environ. 2022, 806, 150500. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Parvathi, K.; Dixon, R.A. Substrate Preferences of Caffeic Acid/5-Hydroxyferulic Acid 3/5-O-Methyltransferases in Developing Stems of Alfalfa (Medicago sativa L.). Arch. Biochem. Biophys. 2000, 375, 175–182. [Google Scholar] [CrossRef]
- Sederoff, R.R.; MacKay, J.J.; Ralph, J.; Hatfield, R.D. Unexpected variation in lignin. Curr. Opin. Plant Biol. 1999, 2, 145–152. [Google Scholar] [CrossRef]
- Richard, S.; Lapointe, G.; Rutledge, R.G.; Séguin, A. Induction of Chalcone Synthase Expression in White Spruce by Wounding and Jasmonate. Plant Cell Physiol. 2000, 41, 982–987. [Google Scholar] [CrossRef]
- Carpin, S.; Crèvecoeur, M.; Greppin, H.; Penel, C. Molecular Cloning and Tissue-Specific Expression of an Anionic Peroxidase in Zucchini1. Plant Physiol. 1999, 120, 799–810. [Google Scholar] [CrossRef]
- Biemelt, S.; Tschiersch, H.; Sonnewald, U. Impact of Altered Gibberellin Metabolism on Biomass Accumulation, Lignin Biosynthesis, and Photosynthesis in Transgenic Tobacco Plants. Plant Physiol. 2004, 135, 254–265. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Xu, X.; Cai, Z.; Yan, B. Effects of ethylene and 1-methylcyclopropene (1-MCP) on lignification of postharvest bamboo shoot. Food Chem. 2007, 105, 521–527. [Google Scholar] [CrossRef]
- Aloni, R.; Tollier, M.T.; Monties, B. The Role of Auxin and Gibberellin in Controlling Lignin Formation in Primary Phloem Fibers and in Xylem of Coleus blumei Stems. Plant Physiol. 1990, 94, 1743–1747. [Google Scholar] [CrossRef] [PubMed]
- Duan, A.-Q.; Feng, K.; Wang, G.-L.; Liu, J.-X.; Xu, Z.-S.; Xiong, A.-S. Elevated gibberellin enhances lignin accumulation in celery (Apium graveolens L.) leaves. Protoplasma 2019, 256, 777–788. [Google Scholar] [CrossRef]
- Rachappanavar, V.; Padiyal, A.; Sharma, J.K.; Gupta, S.K. Plant hormone-mediated stress regulation responses in fruit crops—A review. Sci. Hortic. 2022, 304, 111302. [Google Scholar] [CrossRef]
- Weiss, D.; van Tunen, A.J.; Halevy, A.H.; Mol, J.N.M.; Gerats, A.G.M. Stamens and Gibberellic Acid in the Regulation of Flavonoid Gene Expression in the Corolla of Petunia hybrida. Plant Physiol. 1990, 94, 511–515. [Google Scholar] [CrossRef]
- Zhang, X.; Zhu, J.; Cheng, S.; Zhang, W.-W.; Xu, F.; Liao, Y. Effect of exogenous gibberellin on endogenous hormone and ginkgolide content in Ginkgo leaves. Not. Bot. Horti Agrobot. 2020, 48, 140–149. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, Q.; Liu, S.; Ma, P.; Jia, Z.; Xie, Y.; Bian, X. Effects of exogenous phytohormones on chlorogenic acid accumulation and pathway-associated gene expressions in sweetpotato stem tips. Plant Physiol. Biochem. 2021, 164, 21–26. [Google Scholar] [CrossRef]
- Xie, J.; Li, J.; Jie, Y.; Xie, D.; Yang, D.; Shi, H.; Zhong, Y. Comparative transcriptomics of stem bark reveals genes associated with bast fiber development in Boehmeria nivea L. gaud (ramie). BMC Genom. 2020, 21, 1–17. [Google Scholar] [CrossRef]
- Karakoti, N.; Shankhdhar, S.C.; Shankhdhar, D. Enhanced phenolic and flavonoid content in callus of Withania somnifera (Dunal. L). Progress. Hortic. 2012, 44, 194–200. [Google Scholar]
- Li, J.; Yang, Y.; Chai, M.; Ren, M.; Yuan, J.; Yang, W.; Dong, Y.; Liu, B.; Jian, Q.; Wang, S.; et al. Gibberellins modulate local auxin biosynthesis and polar auxin transport by negatively affecting flavonoid biosynthesis in the root tips of rice. Plant Sci. 2020, 298, 110545. [Google Scholar] [CrossRef] [PubMed]
- Saure, M.C. External control of anthocyanin formation in apple. Sci. Hortic. 1990, 42, 181–218. [Google Scholar] [CrossRef]
- Altmann, M.; Altmann, S.; Rodriguez, P.A.; Weller, B.; Vergara, L.E.; Palme, J.; la Rosa, N.M.-D.; Sauer, M.; Wenig, M.; Villaécija-Aguilar, J.A.; et al. Extensive signal integration by the phytohormone protein network. Nature 2020, 583, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Fraser, C.M.; Chapple, C. The Phenylpropanoid Pathway in Arabidopsis. Arab. Book 2011, 9, e0152. [Google Scholar] [CrossRef]
- Dong, N.Q.; Lin, H.X. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, C.-J. Multifaceted Regulations of Gateway Enzyme Phenylalanine Ammonia-Lyase in the Biosynthesis of Phenylpropanoids. Mol. Plant 2015, 8, 17–27. [Google Scholar] [CrossRef]
- Liu, S.; Hu, Y.; Wang, X.; Han, L.; Song, S.; Cheng, H.; Lin, Z. Isolation and characterization of a gene encoding cinnamate 4-hydroxylase from Parthenocissus henryana. Mol. Biol. Rep. 2009, 36, 1605–1610. [Google Scholar] [CrossRef]
- Negishi, N.; Nanto, K.; Hayashi, K.; Onogi, S.; Kawaoka, A. Transcript abundances of LIM transcription factor, 4CL, CAld5H and CesAs affect wood properties in Eucalyptus globulus. Silvae Genet. 2011, 60, 288–296. [Google Scholar] [CrossRef]
- Yang, S.-L.; Zhang, X.-N.; Lu, G.-L.; Wang, C.-R.; Wang, R. Regulation of gibberellin on gene expressions related with the lignin biosynthesis in ‘Wangkumbae’ pear (Pyrus pyrifolia Nakai) fruit. Plant Growth Regul. 2015, 76, 127–134. [Google Scholar] [CrossRef]
- Blankenship, S.M.; Unrath, C.R. Pal and ethylene content during maturation of red and golden delicious apples. Phytochemistry 1988, 27, 1001–1003. [Google Scholar] [CrossRef]
- Feng, X.; Gao, G.; Yu, C.; Zhu, A.; Chen, J.; Chen, K.; Wang, X.; Abubakar, A.S.; Chen, P. Transcriptome and metabolome analysis reveals anthocyanin biosynthesis pathway associated with ramie (Boehmeria nivea (L.) Gaud.) leaf color formation. BMC Genom. 2021, 22, 684. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, Y.; Zheng, K.; Xie, M.; Feng, K.; Jawdy, S.S.; Gunter, L.E.; Ranjan, P.; Singan, V.R.; Engle, N.; et al. Genome-wide association studies and expression-based quantitative trait loci analyses reveal roles of HCT 2 in caffeoylquinic acid biosynthesis and its regulation by defense-responsive transcription factors in Populus. New Phytol. 2018, 220, 502–516. [Google Scholar] [CrossRef]
- Hoffmann, L.; Besseau, S.; Geoffroy, P.; Ritzenthaler, C.; Meyer, D.; Lapierre, C.; Legrand, M. Si-lencing of hydroxycinnamoyl-coenzyme A shikimate/quinate hydroxycinnamoyltransferase affects phenylpropanoid biosynthesis. Plant Cell 2004, 16, 1446–1465. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, Y.; Zheng, K.; Xie, M.; Feng, K.; Jawdy, S.S.; Muchero, W.; Besseau, S.; Hoff-mann, L.; Geoffroy, P.; et al. Flavonoid accumulation in Arabidopsis repressed in lignin synthesis affects auxin transport and plant growth. Plant Cell 2007, 19, 148–162. [Google Scholar] [CrossRef]
- Li, X.; Bonawitz, N.D.; Weng, J.-K.; Chapple, C. The Growth Reduction Associated with Repressed Lignin Biosynthesis in Arabidopsis thaliana Is Independent of Flavonoids. Plant Cell 2010, 22, 1620–1632. [Google Scholar] [CrossRef]
- Zhu, Z.-J.; Schultz, A.W.; Wang, J.; Johnson, C.H.; Yannone, S.M.; Patti, G.J.; Siuzdak, G. Liquid chromatography quadrupole time-of-flight mass spectrometry characterization of metabolites guided by the METLIN database. Nat. Protoc. 2013, 8, 451–460. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2007, 36 (Suppl. S1), D480–D484. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jie, H.; Zhao, L.; Ma, Y.; Rasheed, A.; Jie, Y. Integrated Transcriptome and Metabolome Analysis Reveal That Exogenous Gibberellin Application Regulates Lignin Synthesis in Ramie. Agronomy 2023, 13, 1450. https://doi.org/10.3390/agronomy13061450
Jie H, Zhao L, Ma Y, Rasheed A, Jie Y. Integrated Transcriptome and Metabolome Analysis Reveal That Exogenous Gibberellin Application Regulates Lignin Synthesis in Ramie. Agronomy. 2023; 13(6):1450. https://doi.org/10.3390/agronomy13061450
Chicago/Turabian StyleJie, Hongdong, Long Zhao, Yushen Ma, Adnan Rasheed, and Yucheng Jie. 2023. "Integrated Transcriptome and Metabolome Analysis Reveal That Exogenous Gibberellin Application Regulates Lignin Synthesis in Ramie" Agronomy 13, no. 6: 1450. https://doi.org/10.3390/agronomy13061450