
Identification of Two Novel Loci Underlying Tolerance to Clavibacter michiganensis Originating from Solanum arcanum LA2157
 , , , and
, , , and
Abstract
:1. Introduction

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tolerance Source | Susceptible Parent | Population | Tolerance/Resistance Type | Reference |
|---|---|---|---|---|
| S. habrochaites LA407 | S. lycopersicum cv. Ohio 86120 | F2 | QTLs Rcm 2.0 on chromosome 2 (Chr2) and Rcm 5.1 on Chr5 | [13] |
| S. arcanum LA2157 | S. lycopersicum cv. Solentos | F2 | QTLs on Chr5, Chr7 and Chr9 | [19] |
| S. arcanum LA2157 | S. lycopersicum cv. MM | Recombinant inbred lines (RILs) | QTLs on Chr5, Chr7, Chr6 and, Chr11 | [21] |
| S. arcanum LA2157 | S. lycopersicum cv. MM | Recombinant inbred lines (RILs) | ~211 kb introgression on Chr7 | [22] |
| S. pimpinellifolium GI. 1554 | S. lycopersicum cv. MM | Recombinant inbred lines (RILs) | QTLs on Chr1, Chr2, Chr7, Chr8, and Chr12 | [24] |
| S. arcanum LA2157 | S. arcanum LA2172 | Backcross (BC) of intraspecific cross | QTLs on Chr1, Chr6, Chr7, Chr8, and Chr10 | [20] |
| S. arcanum var. humifusum | S. lycopersicum × S. chilense LA460 | F2BC population of three genome hybrid S. lycopersicum line Cm 180 | Dominant gene on Chr4 (with modifier genes) | [23] |
2. Materials and Methods
2.1. Plant Materials
2.2. Bacterial Strains and Growth Conditions
2.3. Disease Assays
2.4. Development of Cleaved Amplified Polymorphic Sequences (CAPS) Markers
2.5. Genomic DNA Isolation and Genotypic
2.6. DNA Isolation and Pooling
2.7. Comparative Subsequence Sets Analysis (CoSSA)
2.8. Protein-Protein Interaction Network Prediction
2.9. Protein Prediction of S. arcanum LA2157 Alleles
3. Results
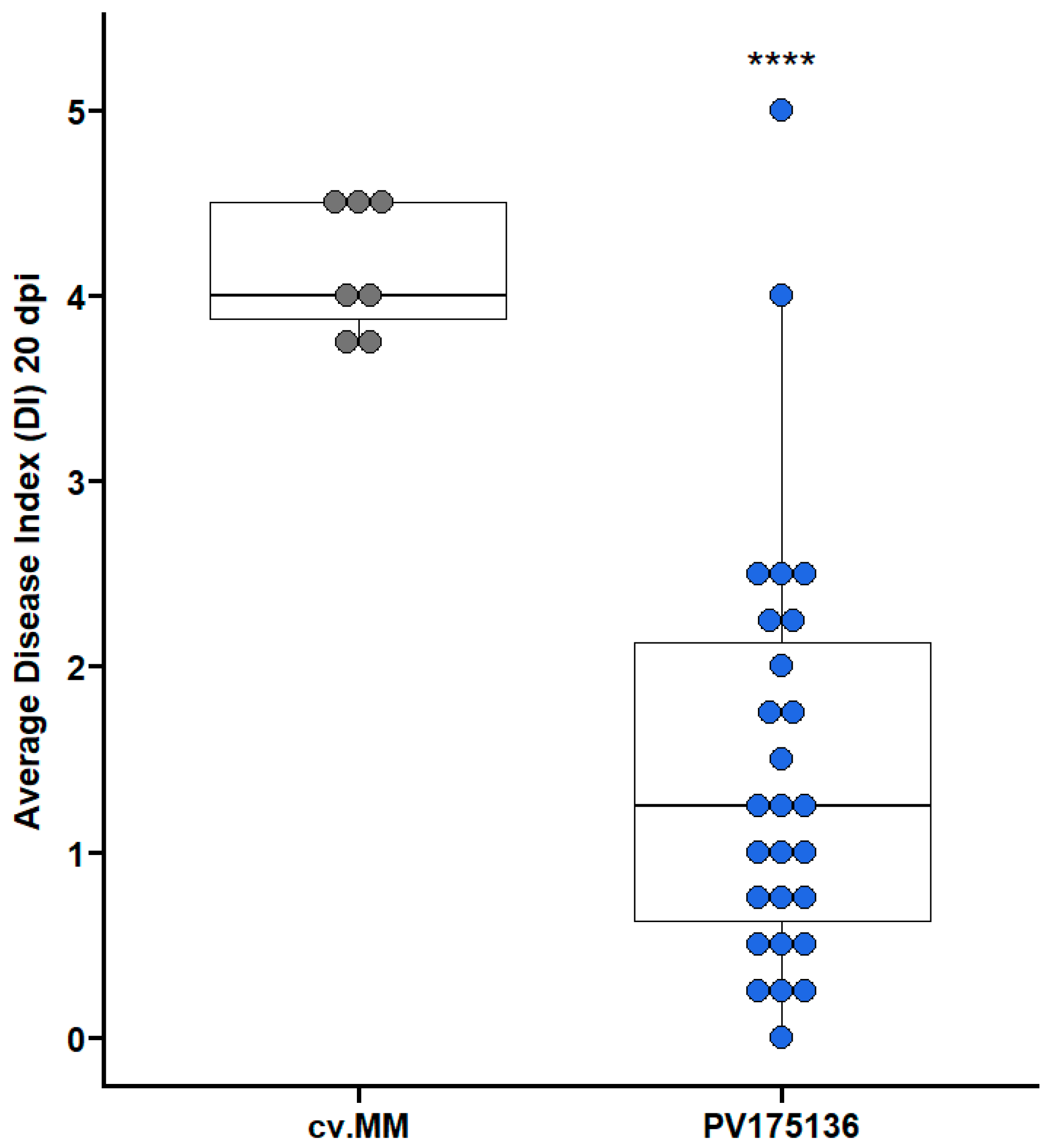
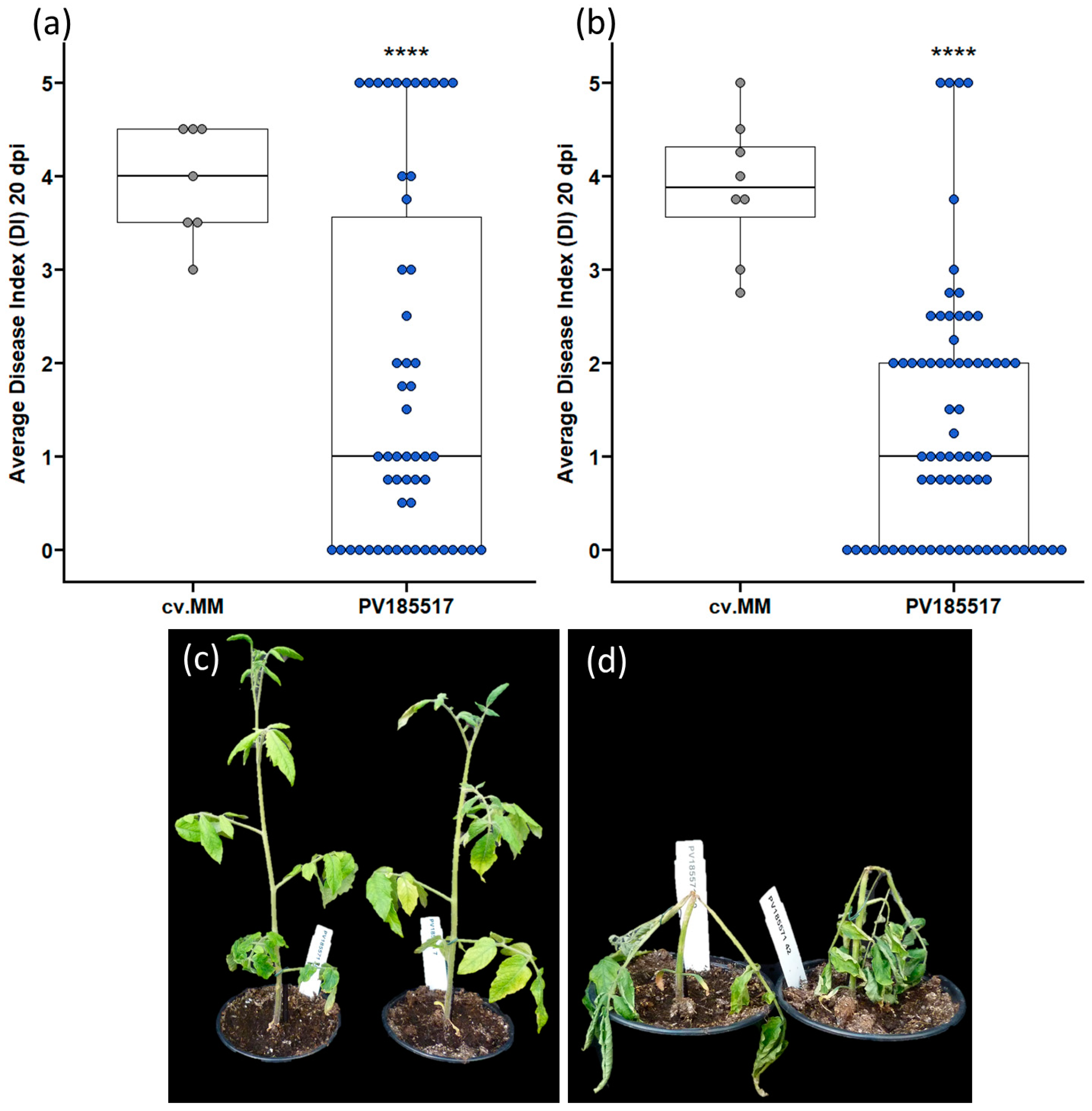
3.1. Phenotypic and Genotypic Evaluation of Lines PV175136 and PV185517
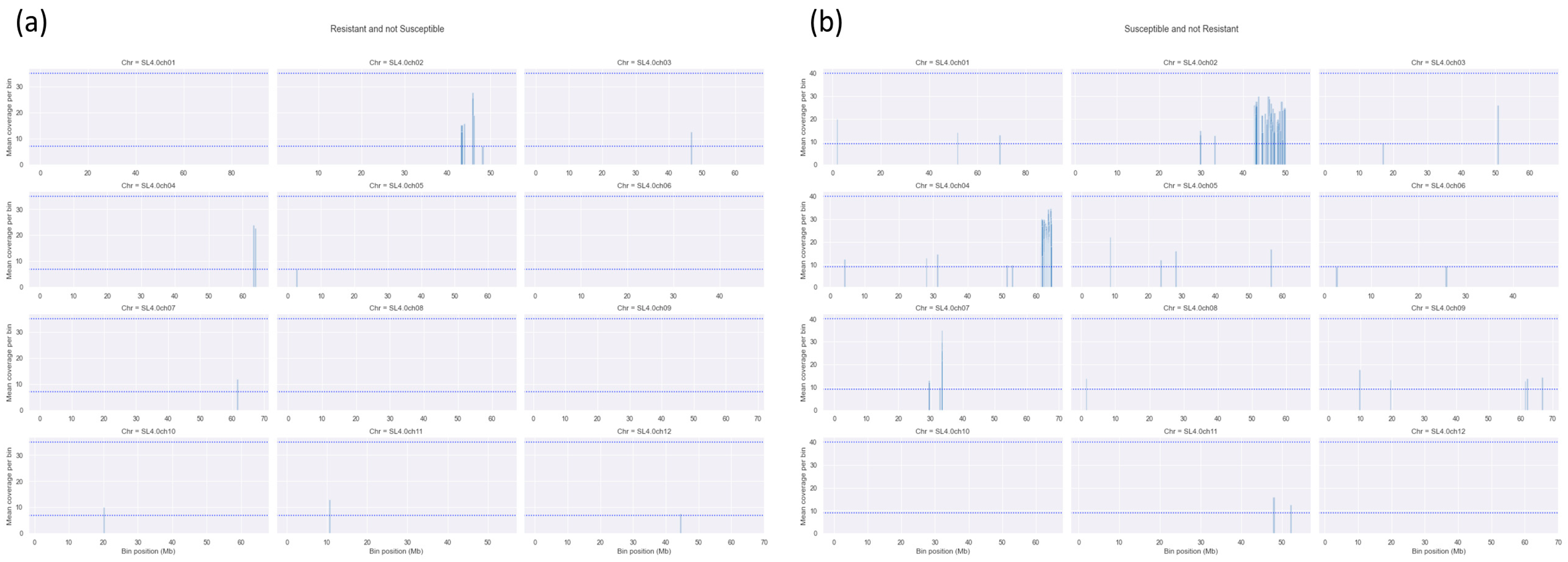
3.2. Additional QTLs on Chromosomes 5 and 9 Do Not Contribute to the Observed Tolerance
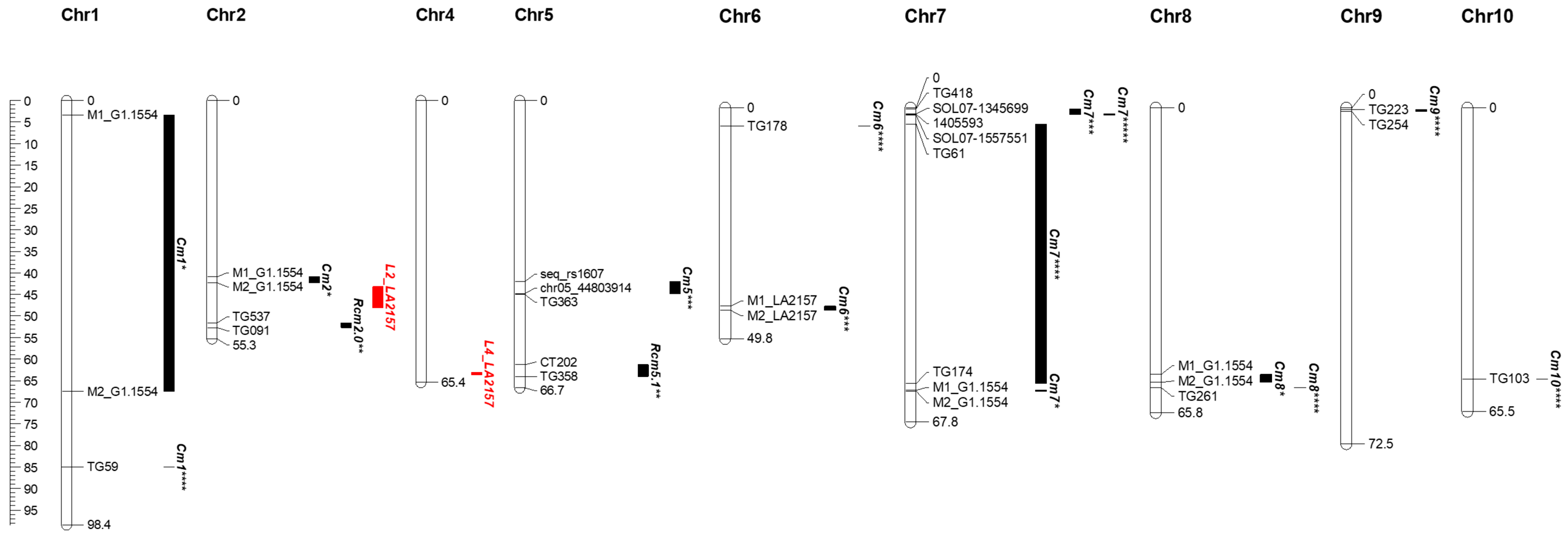
3.3. Tolerance to Cm Requires QTL7 in Combination with Two Additional Loci on Chromosomes 2 and 4
3.4. Changes in the Amino Acid Sequence of Produced by the S. arcanum LA2157 Allelic Variant Lead to Changed Proteins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pagán, I.; García-Arenal, F. Tolerance to Plant Pathogens: Theory and Experimental Evidence. Int. J. Mol. Sci. 2018, 19, 810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, B.A.; Kirchner, J.W. Evolutionary Dynamics of Pathogen Resistance and Tolerance. Evolution 2000, 54, 51–63. [Google Scholar] [CrossRef] [PubMed]
- Eichenlaub, R.; Gartemann, K.-H. The Clavibacter michiganensis Subspecies: Molecular Investigation of Gram-Positive Bacterial Plant Pathogens. Annu. Rev. Phytopathol. 2011, 49, 445–464. [Google Scholar] [CrossRef]
- Gartemann, K.-H.; Kirchner, O.; Engemann, J.; Gräfen, I.; Eichenlaub, R.; Burger, A. Clavibacter michiganensis subsp. michiganensis: First steps in the understanding of virulence of a Gram-positive phytopathogenic bacterium. J. Biotechnol. 2003, 106, 179–191. [Google Scholar] [CrossRef]
- Tancos, M.A.; Chalupowicz, L.; Barash, I.; Manulis-Sasson, S.; Smart, C.D. Tomato Fruit and Seed Colonization by Clavibacter michiganensis subsp. michiganensis through External and Internal Routes. Appl. Environ. Microbiol. 2013, 79, 6948–6957. [Google Scholar] [CrossRef] [Green Version]
- Sen, Y.; van der Wolf, J.; Visser, R.G.F.; van Heusden, S. Bacterial Canker of Tomato: Current Knowledge of Detection, Management, Resistance, and Interactions. Plant Dis. 2015, 99, 4–13. [Google Scholar] [CrossRef] [Green Version]
- Frenkel, O.; Bornestein, M.; Shulhani, R.; Sharabani, G.; Sofer, M.; Abo-Moch, F.; Lofthouse, M.; Manulis-Sasson, S.; Shtienberg, D. Secondary spread of Clavibacter michiganensis subsp. michiganensis in nurseries and the conditions leading to infection of tomato seedlings. Eur. J. Plant Pathol. 2016, 144, 569–579. [Google Scholar] [CrossRef]
- Sharabani, G.; Manulis-Sasson, S.; Borenstein, M.; Shulhani, R.; Lofthouse, M.; Chalupowicz, L.; Shtienberg, D. The significance of guttation in the secondary spread of Clavibacter michiganensis subsp.michiganensis in tomato greenhouses. Plant Pathol. 2013, 62, 578–586. [Google Scholar] [CrossRef]
- Sharabani, G.; Shtienberg, D.; Borenstein, M.; Shulhani, R.; Lofthouse, M.; Sofer, M.; Chalupowicz, L.; Barel, V.; Manulis-Sasson, S. Effects of plant age on disease development and virulence of Clavibacter michiganensis subsp.michiganensis on tomato. Plant Pathol. 2013, 62, 1114–1122. [Google Scholar] [CrossRef]
- Sharabani, G.; Manulis-Sasson, S.; Chalupowicz, L.; Borenstein, M.; Shulhani, R.; Lofthouse, M.; Sofer, M.; Frenkel, O.; Dror, O.; Shtienberg, D. Temperature at the early stages of Clavibacter michiganensis subsp.michiganensis infection affects bacterial canker development and virulence gene expression. Plant Pathol. 2014, 63, 1119–1129. [Google Scholar] [CrossRef]
- Sen, Y.; Feng, Z.; Vandenbroucke, H.; Van Der Wolf, J.; Visser, R.G.F.; Van Heusden, A.W. Screening for new sources of resistance to Clavibacter michiganensis subsp. michiganensis (Cmm) in tomato. Euphytica 2013, 190, 309–317. [Google Scholar] [CrossRef]
- Medina-Mora, C.M.; Hausbeck, M.K.; Fulbright, D.W. Bird’s Eye Lesions of Tomato Fruit Produced by Aerosol and Direct Application of Clavibacter michiganensis subsp. michiganensis. Plant Dis. 2001, 85, 88–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalupowicz, L.; Barash, I.; Reuven, M.; Dror, O.; Sharabani, G.; Gartemann, K.-H.; Eichenlaub, R.; Sessa, G.; Manulis-Sasson, S. Differential contribution of Clavibacter michiganensis ssp. michiganensis virulence factors to systemic and local infection in tomato. Mol. Plant Pathol. 2017, 18, 336–346. [Google Scholar] [CrossRef] [PubMed]
- EFSA. Scientific Opinion on the pest categorisation of Clavibacter michiganensis subsp. michiganensis (Smith) Davis et al. EFSA J. 2014, 12, 3721. [Google Scholar] [CrossRef]
- Thyr, B.D. Inheritance of Resistance to Corynebacterium michiganense in Tomato. Phytopathology 1976, 66, 1116–1119. [Google Scholar] [CrossRef]
- Francis, D.M.; Kabelka, E.; Bell, J.; Franchino, B.; St Clair, D. Resistance to Bacterial Canker in Tomato (Lycopersicon hirsutum LA407) and its Progeny Derived from Crosses to L. esculentum. Plant Dis. 2001, 85, 1171–1176. [Google Scholar] [CrossRef] [Green Version]
- Coaker, G.L.; Francis, D.M. Mapping, genetic effects, and epistatic interaction of two bacterial canker resistance QTLs from Lycopersicon hirsutum. Theor. Appl. Genet. 2004, 108, 1047–1055. [Google Scholar] [CrossRef]
- Kabelka, E.; Franchino, B.; Francis, D.M. Two Loci from Lycopersicon hirsutum LA407 Confer Resistance to Strains of Clavibacter michiganensis subsp.michiganensis. Phytopathology 2002, 92, 504–510. [Google Scholar] [CrossRef] [Green Version]
- Van Heusden, A.W.; Koornneef, M.; Voorrips, R.E.; Brüggemann, W.; Pet, G.; Vrielink-van Ginkel, R.; Chen, X.; Lindhout, P. Three QTLs from Lycopersicon peruvianum confer a high level of resistance to Clavibacter michiganensis ssp. michiganensis. Theor. Appl. Genet. 1999, 99, 1068–1074. [Google Scholar] [CrossRef]
- Sandbrink, J.M.; Van Ooijen, J.W.; Purimahua, C.C.; Vrielink, M.; Verkerk, R.; Zabel, P.; Lindhout, P. Localization of genes for bacterial canker resistance in Lycopersicon peruvianum using RFLPs. Theor. Appl. Genet. 1995, 90, 444–450. [Google Scholar] [CrossRef]
- Sen, Y. Bacterial Canker Resistance in Tomato. Ph.D. Thesis, Wageningen University & Research, Wageningen, The Netherlands, 2014; p. 140. [Google Scholar]
- Muniroh binti Mohd Nadzir, M. Different Approaches of Combating Bacterial Canker in Tomato: In Pursuit of Resistance. Ph.D. Thesis, Wageningen University & Research, Wageningen, The Netherlands, 2018; p. 149. [Google Scholar]
- Vulkova, Z.V.; Sotirova, V.G. Study of the three-genome hybrid Lycopersicon esculentum Mill.—L. chilense Dun.—L. peruvianum var ‘humifusum’ Mill. and its use as a source for resistance. Theor. Appl. Genet. 1993, 87, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Sen, Y.; Manrique, M.J.; Kabaş, A.; Visser, R.G.F.; Heusden, A.W. Qtl Mapping of Clavibacter Michiganensis Subsp. Michiganensis (cmm) Resistance Originating from Solanum Pimpinellifolium g1.1554. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Aflitos, S.; Schijlen, E.; De Jong, H.; De Ridder, D.; Smit, S.; Finkers, R.; Wang, J.; Zhang, G.; Li, N.; Mao, L.; et al. Exploring genetic variation in the tomato (Solanum section Lycopersicon) clade by whole-genome sequencing. Plant J. 2014, 80, 136–148. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J. DNA Protocols for Plants; Springer: Berlin/Heidelberg, Germany, 1991; pp. 283–293. [Google Scholar]
- Prodhomme, C.; Esselink, D.; Borm, T.; Visser, R.G.F.; Van Eck, H.J.; Vossen, J.H. Comparative Subsequence Sets Analysis (CoSSA) is a robust approach to identify haplotype specific SNPs; mapping and pedigree analysis of a potato wart disease resistance gene Sen3. Plant Methods 2019, 15, 60. [Google Scholar] [CrossRef]
- Ohnishi, T.; Szatmari, A.M.; Watanabe, B.; Fujita, S.; Bancos, S.; Koncz, C.; Lafos, M.; Shibata, K.; Yokota, T.; Sakata, K.; et al. C-23 hydroxylation by Arabidopsis CYP90C1 and CYP90D1 reveals a novel shortcut in brassinosteroid biosynthesis. Plant Cell 2006, 18, 3275–3288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peritore-Galve, F.C.; Miller, C.; Smart, C.D. Characterizing Colonization Patterns of Clavibacter michiganensis during Infection of Tolerant Wild Solanum Species. Phytopathology 2020, 110, 574–581. [Google Scholar] [CrossRef] [Green Version]
- Kim, G.-T.; Tsukaya, H.; Uchimiya, H. The ROTUNDIFOLIA3 gene of Arabidopsis thaliana encodes a new member of the cytochrome P-450 family that is required for the regulated polar elongation of leaf cells. Genes Dev. 1998, 12, 2381–2391. [Google Scholar] [CrossRef] [Green Version]
- Oh, M.-H.; Honey, S.H.; Tax, F.E. The Control of Cell Expansion, Cell Division, and Vascular Development by Brassinosteroids: A Historical Perspective. Int. J. Mol. Sci. 2020, 21, 1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Fu, F.; Zhang, H.; Song, F. Genome-wide systematic characterization of the bZIP transcriptional factor family in tomato (Solanum lycopersicum L.). BMC Genom. 2015, 16, 771. [Google Scholar] [CrossRef] [Green Version]
- Ringli, C.; Keller, B. Specific interaction of the tomato bZIP transcription factor VSF-1 with a non-palindromic DNA sequence that controls vascular gene expression. Plant Mol. Biol. 1998, 37, 977–988. [Google Scholar] [CrossRef]
- Torres-Schumann, S.; Ringli, C.; Heierli, D.; Amrhein, N.; Keller, B. In vitro binding of the tomato bZIP transcriptional activator VSF-1 to a regulatory element that controls xylem-specific gene expression. Plant J. 1996, 9, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Dai, S.; Petruccelli, S.; Ordiz, M.I.; Zhang, Z.; Chen, S.; Beachy, R.N. Functional Analysis of RF2a, a Rice Transcription Factor. J. Biol. Chem. 2003, 278, 36396–36402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, S.; Srivastava, A.K.; Schwartz, C.E.; Alexov, E.; Wang, L. Structural assessment of the effects of Amino Acid Substitutions on protein stability and protein protein interaction. Int. J. Comput. Biol. Drug Des. 2010, 3, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]






| PV175136 Line | Q7M1 | Q7M7 | Q7M2 | Q7M3 | Q7M4 | Q7M5 | Q7M9 | Disease Index | Phenotype |
|---|---|---|---|---|---|---|---|---|---|
| PV175136_5 | M | A | A | A | A | M | M | 4 | S |
| PV175136_13 | M | A | A | A | A | M | M | 5 | S |
| PV175136_20 | M | A | A | A | A | M | M | 2.5 | S |
| PV175136_29 | M | A | A | A | A | M | M | 0 | T |
| PV175136_4 | M | A | A | A | A | M | M | 0.25 | T |
| PV175136_27 | M | A | A | A | A | M | M | 0.25 | T |
| PV185517 Line | Q7M1 | Q7M7 | Q7M2 | Q7M3 | Q7M4 | Q7M5 | Q7M9 | Disease Index | Phenotype |
|---|---|---|---|---|---|---|---|---|---|
| PV185517_14 | M | A | A | A | A | M | M | 5 | S |
| PV185517_18 | M | A | A | A | A | M | M | 3.75 | S |
| PV185517_31 | M | A | A | A | A | M | M | 5 | S |
| PV185517_37 | M | A | A | A | A | M | M | 0 | T |
| PV185517_39 | M | A | A | A | A | M | M | 0 | T |
| PV185517_40 | M | A | A | A | A | M | M | 0 | T |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koseoglou, E.; Brouwer, M.; Mudadirwa, D.; Van der Wolf, J.M.; Visser, R.G.F.; Bai, Y. Identification of Two Novel Loci Underlying Tolerance to Clavibacter michiganensis Originating from Solanum arcanum LA2157. Agronomy 2023, 13, 953. https://doi.org/10.3390/agronomy13040953
Koseoglou E, Brouwer M, Mudadirwa D, Van der Wolf JM, Visser RGF, Bai Y. Identification of Two Novel Loci Underlying Tolerance to Clavibacter michiganensis Originating from Solanum arcanum LA2157. Agronomy. 2023; 13(4):953. https://doi.org/10.3390/agronomy13040953
Chicago/Turabian StyleKoseoglou, Eleni, Matthijs Brouwer, Derek Mudadirwa, Jan M. Van der Wolf, Richard G. F. Visser, and Yuling Bai. 2023. "Identification of Two Novel Loci Underlying Tolerance to Clavibacter michiganensis Originating from Solanum arcanum LA2157" Agronomy 13, no. 4: 953. https://doi.org/10.3390/agronomy13040953