

Applying the SIMPLE Crop Model to Assess Soybean (Glicine max. (L.) Merr.) Biomass and Yield in Tropical Climate Variation
 , , ,
, , ,
Abstract
:1. Introduction
2. Methods
2.1. Field Experiments
- -
- Sowing date, flowering date, maturity date, and growing time.
- -
- Plant height (cm), leaves number, total vegetative biomass (t ha−1) per 10 days. vegetative biomass (t ha−1), and grain yield.
2.2. Modeling of Soybean Yield
2.3. Model Parameters and Inputs
2.4. Model Assessment
2.5. Scenario Analysis
3. Results and Discussions
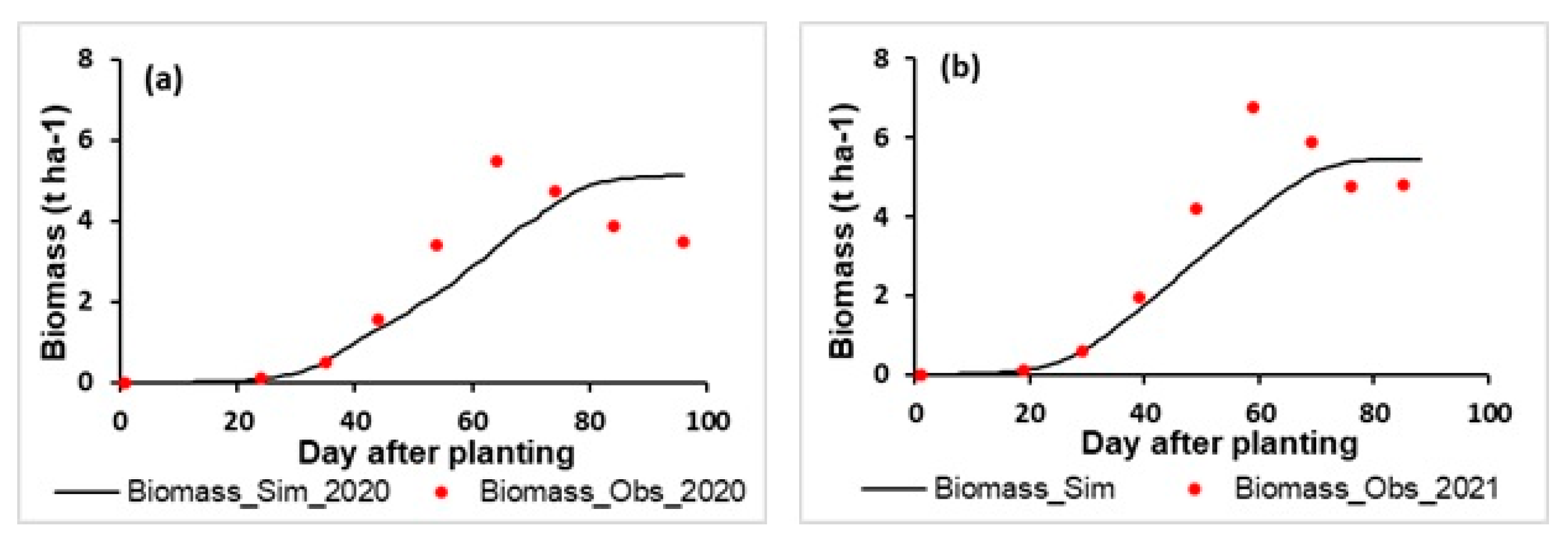
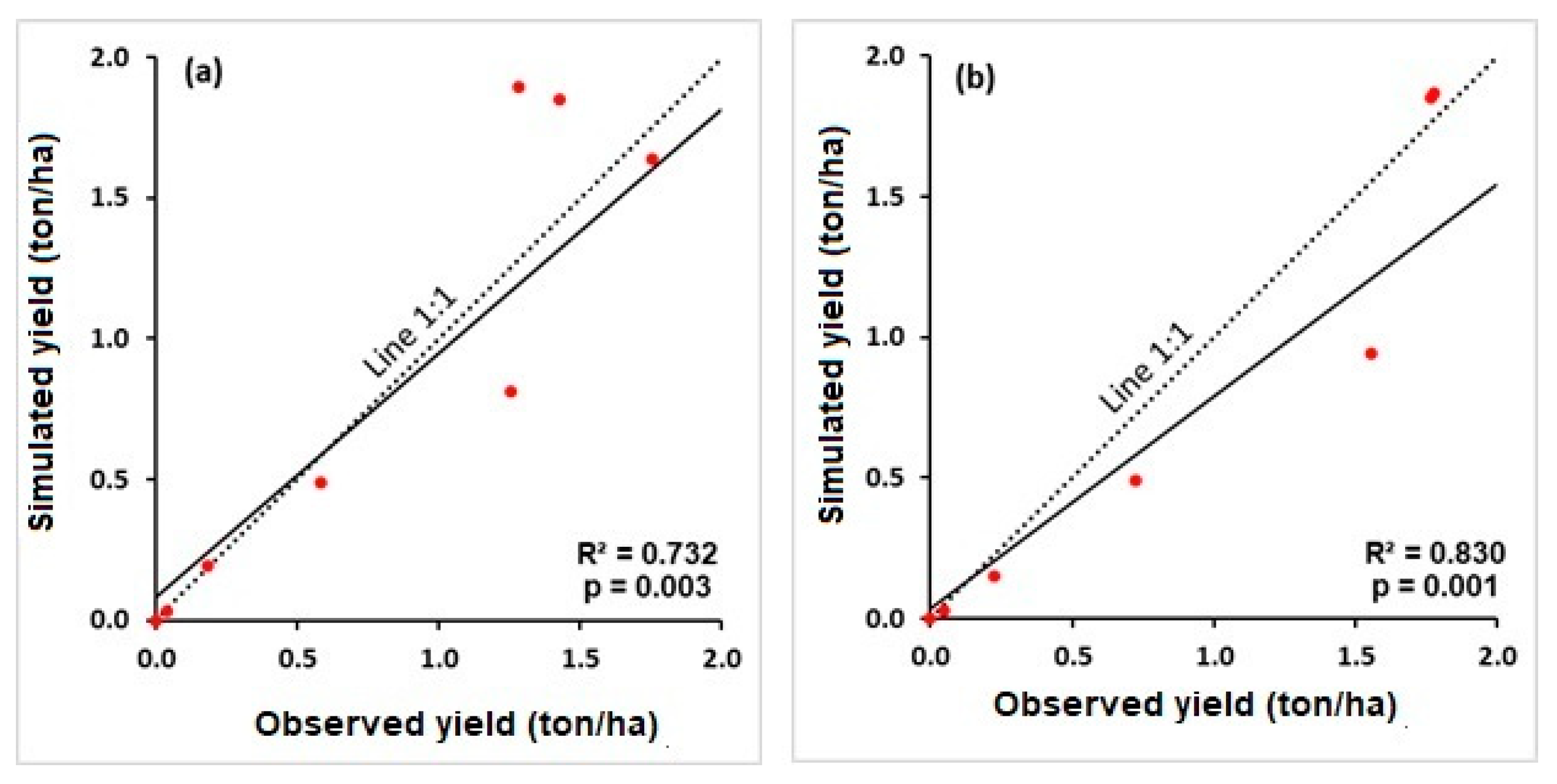
3.1. Model Assessment
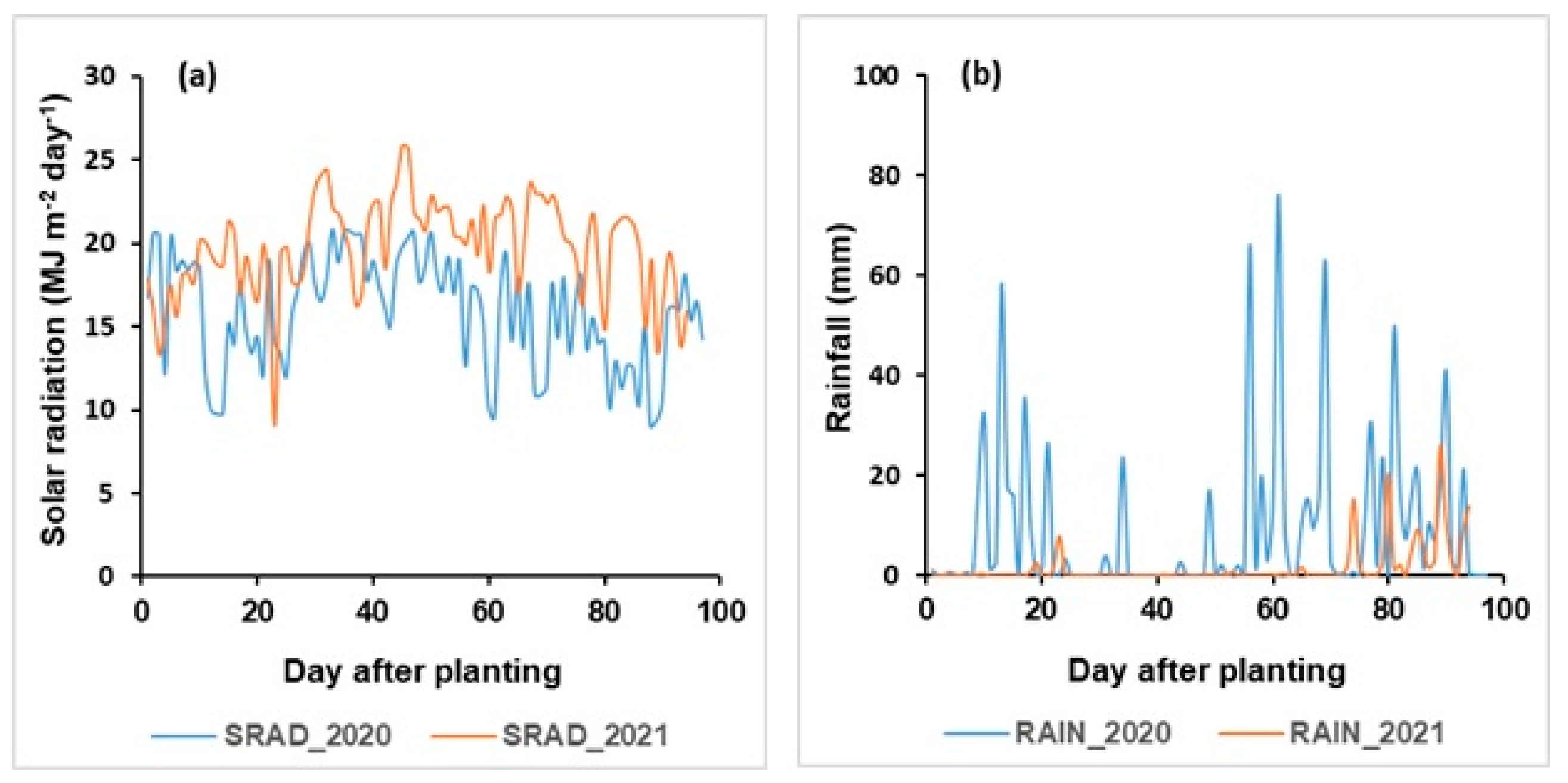
3.2. Effects of Climate Variation
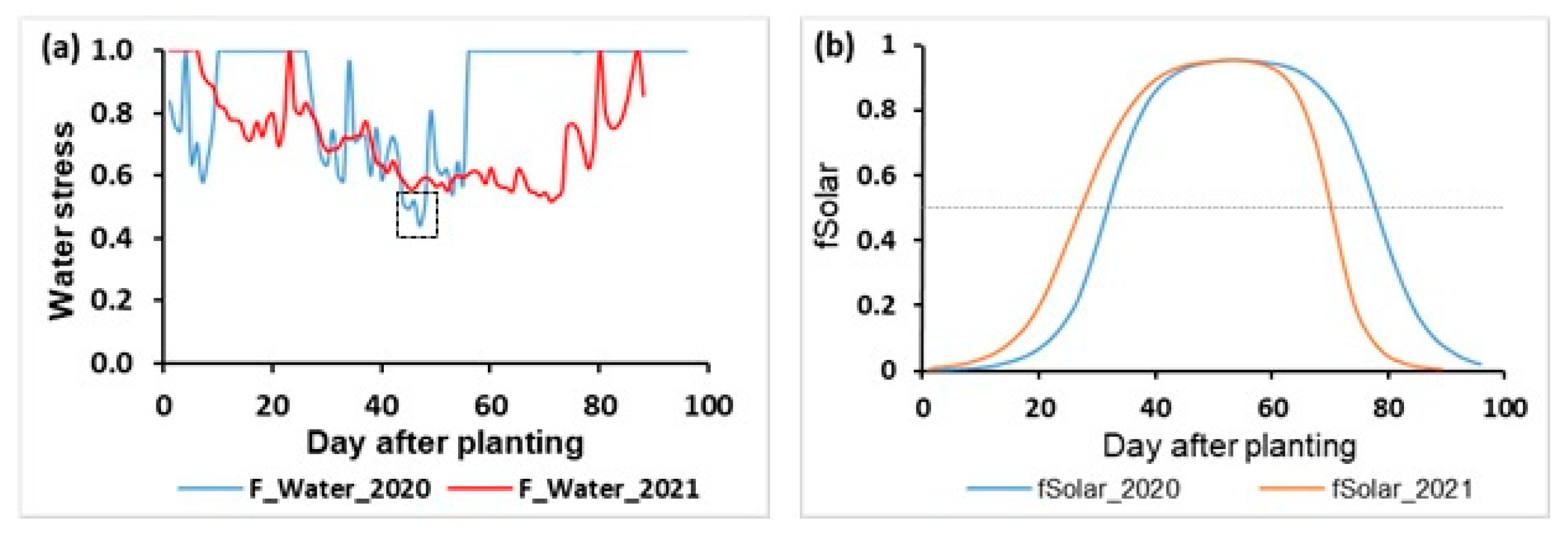
3.2.1. Growth Stages
3.2.2. Rainfall and Drought
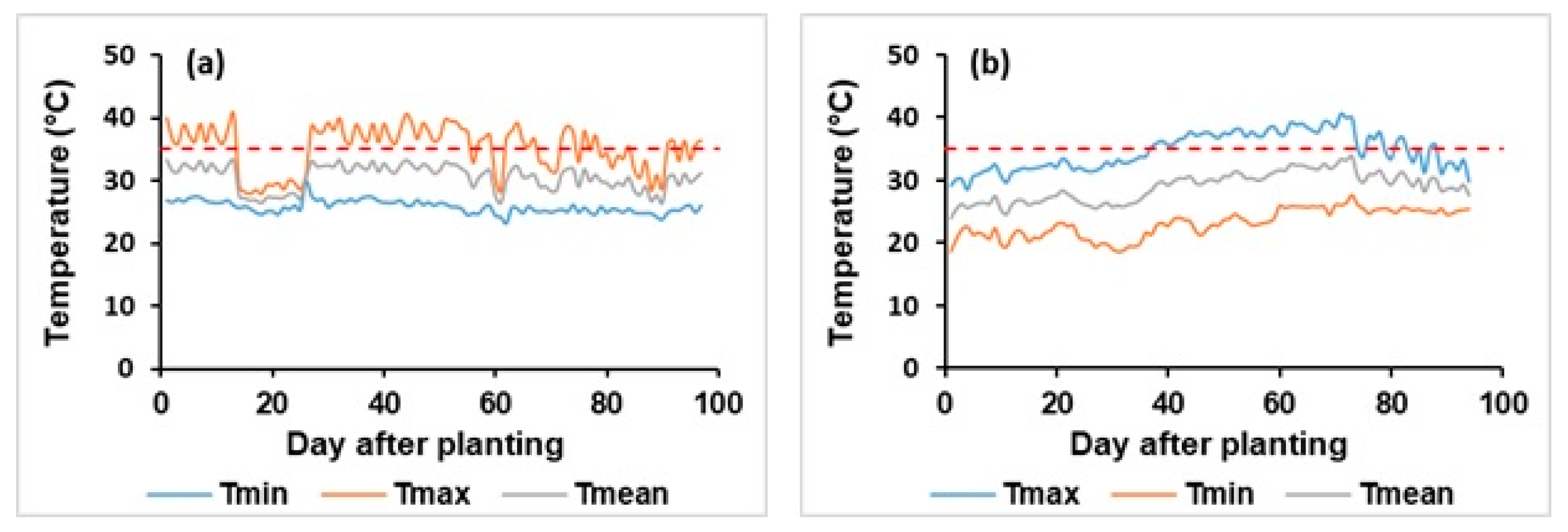
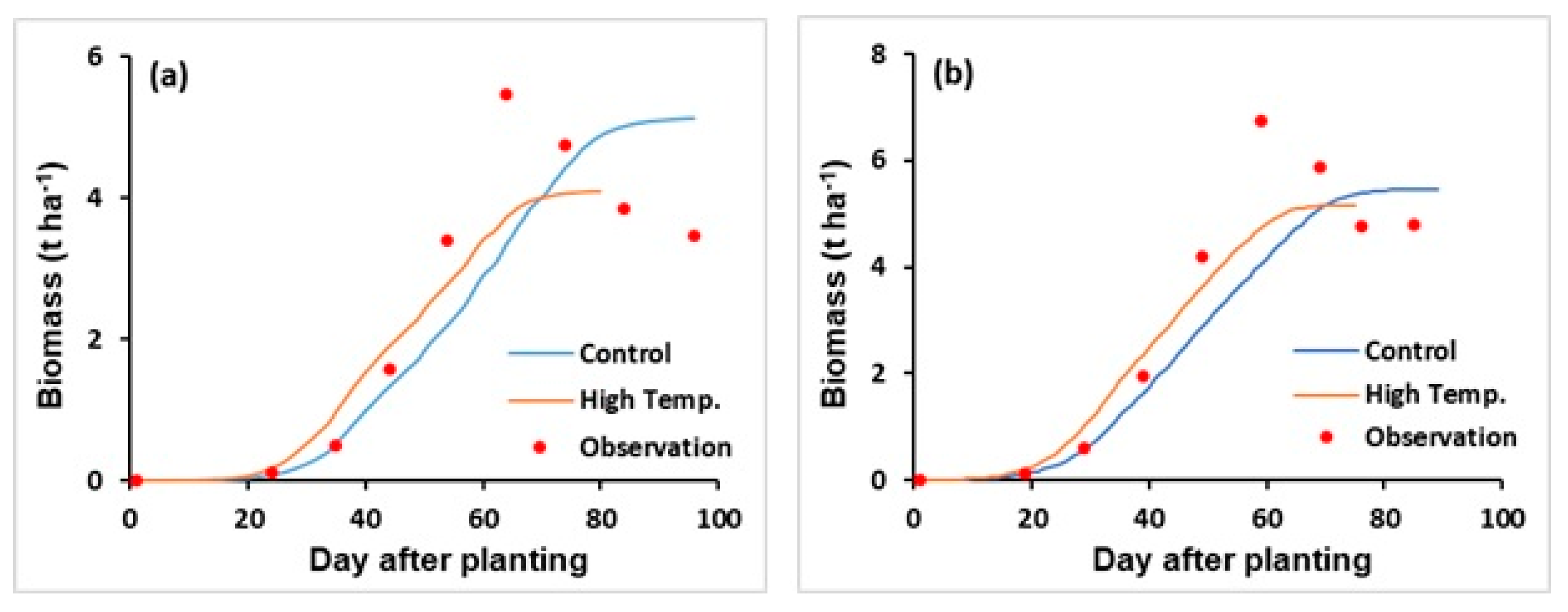
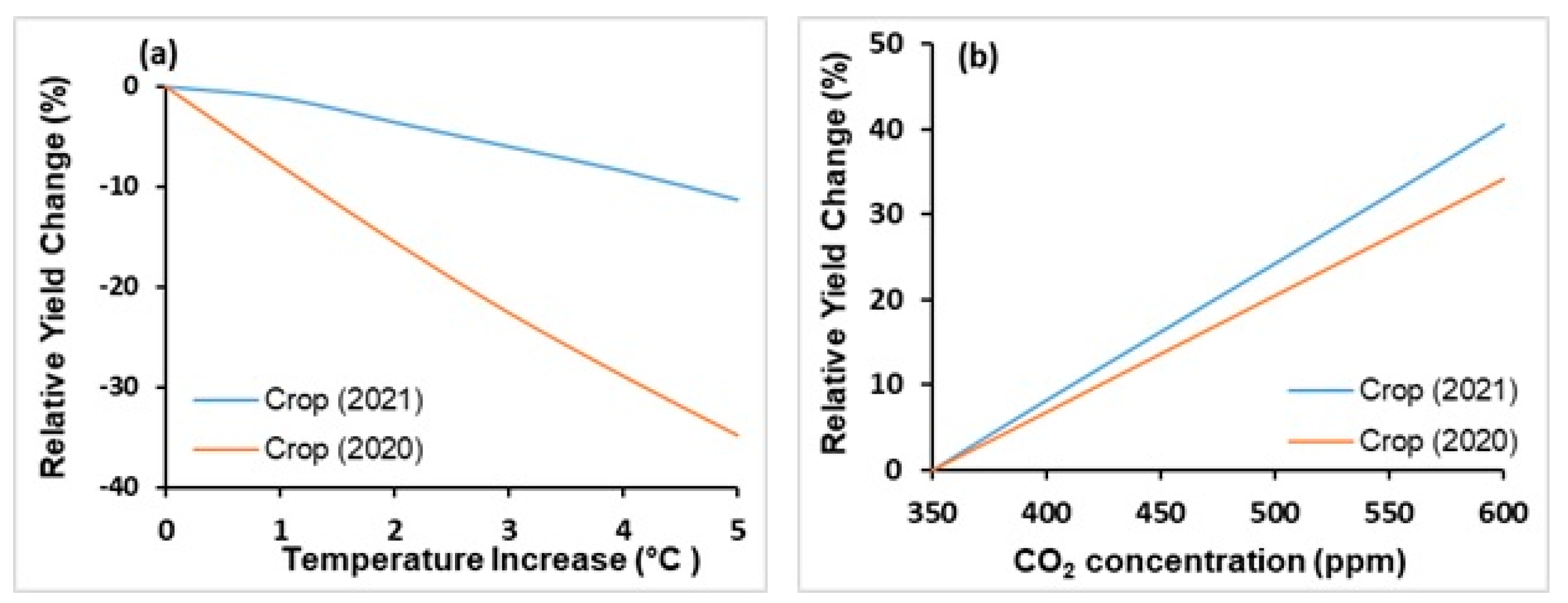
3.2.3. Temperature Effects
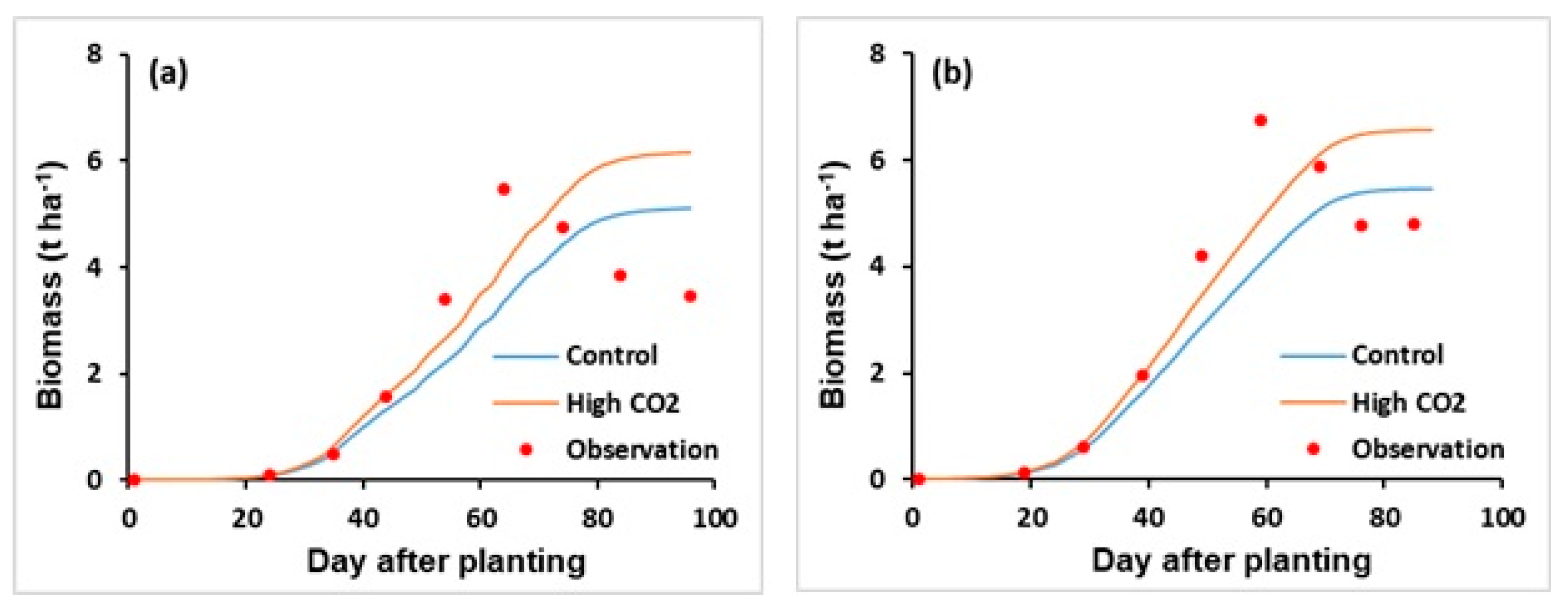
3.2.4. CO2 Fertilization Effect
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- El Bassam, N. Handbook of Bioenergy Crops: A Complete Reference to Species, Development and Applications; Routledge: Abingdon-on-Thames, UK, 2010. [Google Scholar]
- Saryoko, A.; Homma, K.; Lubis, I.; Shiraiwa, T. Plant development and yield components under a tropical environment in soybean cultivars with temperate and tropical origins. Plant Prod. Sci. 2017, 20, 375–383. [Google Scholar] [CrossRef]
- Kucharik, C.J.; Serbin, S.P. Impacts of recent climate change on Wisconsin corn and soybean yield trends. Environ. Res. Lett. 2008, 3, 034003. [Google Scholar] [CrossRef]
- Zanon, A.J.; Streck, N.A.; Grassini, P. Climate and Management Factors Influence Soybean Yield Potential in a Subtropical Environment. Agron. J. 2016, 108, 1447–1454. [Google Scholar] [CrossRef]
- Reis, L.; Santos e Silva, C.M.; Bezerra, B.; Mutti, P.; Spyrides, M.H.; Silva, P.; Magalhães, T.; Ferreira, R.; Rodrigues, D.; Andrade, L. Influence of climate variability on soybean yield in MATOPIBA, Brazil. Atmosphere 2020, 11, 1130. [Google Scholar] [CrossRef]
- Khojely, D.M.; Ibrahim, S.E.; Sapey, E.; Han, T. History, current status, and prospects of soybean production and research in sub-Saharan Africa. Crop J. 2018, 6, 226–235. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Xiao, L.; Hoogenboom, G.; Boote, K.J.; Kassie, B.T.; Pavan, W.; Shelia, V.; Kim, K.S.; Hernandez-Ochoa, I.M.; et al. A SIMPLE crop model. Eur. J. Agron. 2019, 104, 97–106. [Google Scholar] [CrossRef]
- Banterng, P.; Hoogenboom, G.; Patanothai, A.; Singh, P.; Wani, S.; Pathak, P.; Tongpoonpol, S.; Atichart, S.; Srihaban, P.; Buranaviriyakul, S. Application of the Cropping System Model (CSM)-CROPGRO-Soybean for Determining Optimum Management Strategies for Soybean in Tropical Environments. J. Agron. Crop Sci. 2010, 196, 231–242. [Google Scholar] [CrossRef]
- Wu, X.; Vuichard, N.; Ciais, P.; Viovy, N.; de Noblet-Ducoudré, N.; Wang, X.; Magliulo, V.; Wattenbach, M.; Vitale, L.; Di Tommasi, P. ORCHIDEE-CROP (v0), a new process-based agro-land surface model: Model description and evaluation over Europe. Geosci. Model Dev. 2016, 9, 857–873. [Google Scholar] [CrossRef]
- Li, Z.; Zhan, C.; Hu, S.; Ning, L.; Wu, L.; Guo, H. Evaluation of global gridded crop models (GGCMs) for the simulation of major grain crop yields in China. Hydrol. Res. 2022, 53, 353–369. [Google Scholar] [CrossRef]
- Schauberger, B.; Archontoulis, S.; Arneth, A.; Balkovic, J.; Ciais, P.; Deryng, D.; Elliott, J.; Folberth, C.; Khabarov, N.; Müller, C.; et al. Consistent negative response of US crops to high temperatures in observations and crop models. Nat. Commun. 2017, 8, 13931. [Google Scholar] [CrossRef]
- Goulart, H.M.; van der Wiel, K.; Folberth, C.; Boere, E.; van den Hurk, B. Increase of simultaneous soybean failures due to climate change. Earth’s Future 2023, 11, e2022EF003106. [Google Scholar] [CrossRef]
- Sun, Q.; Zhang, Y.; Che, X.; Chen, S.; Ying, Q.; Zheng, X.; Feng, A. Coupling Process-Based Crop Model and Extreme Climate Indicators with Machine Learning Can Improve the Predictions and Reduce Uncertainties of Global Soybean Yields. Agriculture 2022, 12, 1791. [Google Scholar] [CrossRef]
- Kaul, M.; Hill, R.L.; Walthall, C. Artificial neural networks for corn and soybean yield prediction. Agric. Syst. 2005, 85, 1–18. [Google Scholar] [CrossRef]
- Dasgupta, P. Chapter 5 Population, poverty, and the natural environment. In Handbook of Environmental Economics; Mäler, K.-G., Jeffrey, R.V., Eds.; Elsevier: Amsterdam, The Netherlands, 2003; Volume 1, pp. 191–247. [Google Scholar]
- Phung, D.; Nguyen-Huy, T.; Tran, N.N.; Tran, D.N.; Nghiem, S.; Nguyen, N.H.; Nguyen, T.H.; Bennett, T. Hydropower dams, river drought and health effects: A detection and attribution study in the lower Mekong Delta Region. Clim. Risk Manag. 2021, 32, 100280. [Google Scholar] [CrossRef]
- Monteith, J.L. Light Distribution and Photosynthesis in Field Crops. Ann. Bot. 1965, 29, 17–37. [Google Scholar] [CrossRef]
- Ahmed, M.; Ahmad, S. Systems Modeling; Spinger: Singapore, 2020; pp. 1–44. [Google Scholar]
- Ross, J. The Radiation Regime and Architecture of Plant Stands; Springer Science & Business Media: Berlin, Germany, 1981. [Google Scholar]
- Asseng, S.; Foster, I.A.N.; Turner, N.C. The impact of temperature variability on wheat yields. Glob. Chang. Biol. 2011, 17, 997–1012. [Google Scholar] [CrossRef]
- Bindi, M.; Fibbi, L.; Gozzini, B.; Orlandini, S.; Miglietta, F. Modelling the impact of future climate scenarios on yield and yield variability of grapevine. Clim. Res. 1996, 7, 213–224. [Google Scholar] [CrossRef]
- Ewert, F.; Rodriguez, D.; Jamieson, P.; Semenov, M.A.; Mitchell, R.A.C.; Goudriaan, J.; Porter, J.R.; Kimball, B.A.; Pinter, P.J.; Manderscheid, R.; et al. Effects of elevated CO2 and drought on wheat: Testing crop simulation models for different experimental and climatic conditions. Agric. Ecosyst. Environ. 2002, 93, 249–266. [Google Scholar] [CrossRef]
- Priestley, C.H.B.; Taylor, R.J.F. On the Assessment of Surface Heat Flux and Evaporation Using Large-Scale Parameters. Mon. Weather. Rev. 1972, 100, 81–92. [Google Scholar] [CrossRef]
- Ritchie, J.; Godwin, D.; Otter-Nacke, S. CERES-Wheat: A User-Oriented Wheat Yield Model. Preliminary Documentation. AGRISTARS Publication No.: YM-U3-04442-JSC-18892; Michigan State University: East Lansing, MI, USA, 1985. [Google Scholar]
- van Ittersum, M.K.; Howden, S.M.; Asseng, S. Sensitivity of productivity and deep drainage of wheat cropping systems in a Mediterranean environment to changes in CO2, temperature and precipitation. Agric. Ecosyst. Environ. 2003, 97, 255–273. [Google Scholar] [CrossRef]
- Woli, P.; Jones, J.W.; Ingram, K.T.; Fraisse, C.W. Agricultural Reference Index for Drought (ARID). Agron. J. 2012, 104, 287–300. [Google Scholar] [CrossRef]
- Sinclair, T.R.; Tanner, C.B.; Bennett, J.M. Water-Use Efficiency in Crop Production. BioScience 1984, 34, 36–40. [Google Scholar] [CrossRef]
- Steduto, P.; Hsiao, T.C.; Raes, D.; Fereres, E. AquaCrop—The FAO Crop Model to Simulate Yield Response to Water: I. Concepts and Underlying Principles. Agron. J. 2009, 101, 426–437. [Google Scholar] [CrossRef]
- Wnuk, A.; Górny, A.; Bocianowski, J.; Kozak, M. Visualizing harvest index in crops. Commun. Biometry Crop Sci. 2013, 8, 48–59. [Google Scholar]
- Milton, E.P.-F.; Flávio, B.J. Yield Components and Biomass Partition in Soybean: Climate Change Vision. In Soybean-Biomass, Yield and Productivity; Intechopen: London, UK, 2019. [Google Scholar]
- Whigham, D.K.; Minor, H.C. 4-Agronomic Characteristics and Environmental Stress. In Soybean Physiology, Agronomy, and Utilization; Norman, A.G., Ed.; Academic Press: Cambridge, MA, USA, 1978; pp. 77–118. [Google Scholar]
- Ries, L.L.; Purcell, L.C.; Carter, T.E., Jr.; Edwards, J.T.; King, C.A. Physiological Traits Contributing to Differential Canopy Wilting in Soybean under Drought. Crop Sci. 2012, 52, 272–281. [Google Scholar] [CrossRef]
- Munoz, G.; Maraux, F.; Wahaj, R. Actual Crop Water Use in Project Countries a Synthesis at the Regional Level; The World Bank: Washington, DC, USA, 2007. [Google Scholar]
- Jamieson, P.D.; Porter, J.R.; Wilson, D.R. A test of the computer simulation model ARCWHEAT1 on wheat crops grown in New Zealand. Field Crops Res. 1991, 27, 337–350. [Google Scholar] [CrossRef]
- Adeboye, O.B.; Schultz, B.; Adekalu, K.O.; Prasad, K.C. Performance evaluation of AquaCrop in simulating soil water storage, yield, and water productivity of rainfed soybeans (Glycine max L. merr) in Ile-Ife, Nigeria. Agric. Water Manag. 2019, 213, 1130–1146. [Google Scholar] [CrossRef]
- Liu, S.; Yang, J.Y.; Zhang, X.Y.; Drury, C.F.; Reynolds, W.D.; Hoogenboom, G. Modelling crop yield, soil water content and soil temperature for a soybean–maize rotation under conventional and conservation tillage systems in Northeast China. Agric. Water Manag. 2013, 123, 32–44. [Google Scholar] [CrossRef]
- Gaso, D.V.; de Wit, A.; Berger, A.G.; Kooistra, L. Predicting within-field soybean yield variability by coupling Sentinel-2 leaf area index with a crop growth model. Agric. For. Meteorol. 2021, 308–309, 108553. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, K.; Kang, J.; Liang, F. Effects of rainfall, temperature and illumination on outcrossing rate of male sterile line in soybean. Oil Crop Sci. 2020, 5, 17–21. [Google Scholar] [CrossRef]
- Matthews, E.R.; Mazer, S.J. Historical changes in flowering phenology are governed by temperature × precipitation interactions in a widespread perennial herb in western North America. New Phytol. 2016, 210, 157–167. [Google Scholar] [CrossRef]
- Smith, J. Crop, Pasture and Timber Yield Index. Cedara Report N/A/94/4; Natal Agricultural Research Institute: Cedara, South Africa, 1994. [Google Scholar]
- Schlenker, W.; Roberts, M.J. Nonlinear temperature effects indicate severe damages to US crop yields under climate change. Proc. Natl. Acad. Sci. USA 2009, 106, 15594–15598. [Google Scholar] [CrossRef]
- Wiebbecke, C.E.; Graham, M.A.; Cianzio, S.R.; Palmer, R.G. Day temperature influences the male-sterile locus ms9 in soybean. Crop Sci. 2012, 52, 1503–1510. [Google Scholar] [CrossRef]
- Caviness, C.; Fagala, B. Influence of Temperature on a Partially Male-Sterile Soybean Strain 1. Crop Sci. 1973, 13, 503–504. [Google Scholar] [CrossRef]
- Allen, L.H.; Zhang, L.; Boote, K.J.; Hauser, B.A. Elevated temperature intensity, timing, and duration of exposure affect soybean internode elongation, mainstem node number, and pod number per plant. Crop J. 2018, 6, 148–161. [Google Scholar] [CrossRef]
- Liu, Y.; Dai, L. Modelling the impacts of climate change and crop management measures on soybean phenology in China. J. Clean. Prod. 2020, 262, 121271. [Google Scholar] [CrossRef]
- Wang, X.; Zhao, C.; Müller, C.; Wang, C.; Ciais, P.; Janssens, I.; Peñuelas, J.; Asseng, S.; Li, T.; Elliott, J. Emergent constraint on crop yield response to warmer temperature from field experiments. Nat. Sustain. 2020, 3, 908–916. [Google Scholar] [CrossRef]
- Burroughs, C.H.; Montes, C.M.; Moller, C.A.; Mitchell, N.G.; Michael, A.M.; Peng, B.; Kimm, H.; Pederson, T.L.; Lipka, A.E.; Bernacchi, C.J.; et al. Reductions in leaf area index, pod production, seed size, and harvest index drive yield loss to high temperatures in soybean. J. Exp. Bot. 2022, 74, 1629–1641. [Google Scholar] [CrossRef] [PubMed]
- Leng, G.; Hall, J. Crop yield sensitivity of global major agricultural countries to droughts and the projected changes in the future. Sci. Total Environ. 2019, 654, 811–821. [Google Scholar] [CrossRef]
- Fernandes, R.D.M.; de Melo, D.M.; Elli, E.F.; Battisti, R. Climate change impacts on rainfed and irrigated soybean yield in Brazil’s new agricultural frontier. Theor. Appl. Climatol. 2022, 147, 803–816. [Google Scholar] [CrossRef]
- Alsajri, F.A.; Wijewardana, C.; Bheemanahalli, R.; Irby, J.T.; Krutz, J.; Golden, B.; Reddy, V.R.; Reddy, K.R. Morpho-physiological, yield, and transgenerational seed germination responses of soybean to temperature. Front. Plant Sci. 2022, 13, 839270. [Google Scholar] [CrossRef]
- Jumrani, K.; Bhatia, V.S. Interactive effect of temperature and water stress on physiological and biochemical processes in soybean. Physiol. Mol. Biol. Plants 2019, 25, 667–681. [Google Scholar] [CrossRef] [PubMed]
- Casali, L.; Herrera, J.M.; Rubio, G. Resilient soybean and maize production under a varying climate in the semi-arid and sub-humid Chaco. Eur. J. Agron. 2022, 135, 126463. [Google Scholar] [CrossRef]
- Sakurai, G.; Iizumi, T.; Nishimori, M.; Yokozawa, M. How much has the increase in atmospheric CO2 directly affected past soybean production? Sci. Rep. 2014, 4, 4978. [Google Scholar] [CrossRef]
- Hao, X.; Gao, J.; Han, X.; Ma, Z.; Merchant, A.; Ju, H.; Li, P.; Yang, W.; Gao, Z.; Lin, E. Effects of open-air elevated atmospheric CO2 concentration on yield quality of soybean (Glycine max (L.) Merr). Agric. Ecosyst. Environ. 2014, 192, 80–84. [Google Scholar] [CrossRef]
- Lal, M.; Singh, K.K.; Srinivasan, G.; Rathore, L.S.; Naidu, D.; Tripathi, C.N. Growth and yield responses of soybean in Madhya Pradesh, India to climate variability and change. Agric. For. Meteorol. 1999, 93, 53–70. [Google Scholar] [CrossRef]
- Haskett, J.D.; Pachepsky, Y.A.; Acock, B. Increase of CO2 and Climate Change Effects on Iowa Soybean Yield, Simulated Using GLYCIM. Agron. J. 1997, 89, 167–176. [Google Scholar] [CrossRef]
- Srivastava, A.; McGhee, R.P.; Flanagan, D.C.; Frankenberger, J.R.; Engel, B.A. Modeling the Impacts of Climate change on Crop Yield, Runoff, and Soil Loss Using the Water Erosion Prediction Project (WEPP) Model. In Soil Erosion Research Under a Changing Climate, January 8–13, 2023, Aguadilla, Puerto Rico; ASABE: St. Joseph, MI, USA, 2023. [Google Scholar]
- Thomey, M.L.; Slattery, R.A.; Köhler, I.H.; Bernacchi, C.J.; Ort, D.R. Yield response of field-grown soybean exposed to heat waves under current and elevated [CO2]. Glob. Chang. Biol. 2019, 25, 4352–4368. [Google Scholar] [CrossRef]
- Li, B.; Feng, Y.; Zong, Y.; Zhang, D.; Hao, X.; Li, P. Elevated CO2-induced changes in photosynthesis, antioxidant enzymes and signal transduction enzyme of soybean under drought stress. Plant Physiol. Biochem. 2020, 154, 105–114. [Google Scholar] [CrossRef]
- Köhler, I.H.; Huber, S.C.; Bernacchi, C.J.; Baxter, I.R. Increased temperatures may safeguard the nutritional quality of crops under future elevated CO2 concentrations. Plant J. 2019, 97, 872–886. [Google Scholar] [CrossRef]
- Qiao, Y.; Miao, S.; Li, Q.; Jin, J.; Luo, X.; Tang, C. Elevated CO2 and temperature increase grain oil concentration but their impacts on grain yield differ between soybean and maize grown in a temperate region. Sci. Total Environ. 2019, 666, 405–413. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Climatic Features | Year | |
|---|---|---|
| 2020 | 2021 | |
| Total rainfall (mm) | 874.1 | 89.1 |
| Number of rainy days | 62.0 | 38.0 |
| Mean temperature (°C) * | 30.5 | 29.0 |
| Mean radiation (MJ m−2d−1) ** | 15.8 | 19.7 |
| Items | Characteristics |
|---|---|
| Soil texture | |
| Sand (%) | 30 |
| Silt (%) | 42 |
| Clay (%) | 28 |
| Soil properties or soil parameters | |
| Total nitrogen (%) | 0.17 |
| Total phosphorus (%P2O5) | 0.03 |
| Exchangeable potassium (cmol kg−1) | 0.23 |
| Field capacity (cm3 cm−3) | 0.18 |
| Available water capacity—AWC (cm3 cm−3) | 0.14 |
| Runoff curve number—RCN | 84 |
| Deep drainage coefficient—DDC | 0.18 |
| Root zone—RZD (mm) | 329 |
| Ground water level (cm) | 35.8 |
| Fertilizer applied | |
| N (kg) | 55 |
| P (kg) | 75 |
| K (kg) | 80 |
| Name | Description | Range | Note |
|---|---|---|---|
| Cultivars parameters | |||
| Tsum | Thermal time requirement from sowing to maturity in daily mean (°C days). | 2070–2300 | * |
| HI | Harvest index. | 0.30–0.37 | * |
| I50A | Thermal time requirement after sowing fraction of light interception to reach 50% (°C days). | 500–710 | * |
| I50B | Represents natural senescence. Thermal time requirement from maturity backwards for light interception to reach 50% (°C days). | 200–250 | * |
| Species parameters | |||
| Tbase | Base temperature (daily mean T) for phenology development and growth (°C). | 6–10 | [30] |
| Topt | Optimal temperature (daily mean T) for biomass growth (°C). | 25–30 | [31] |
| RUE | Radiation use efficiency (above ground biomass and below ground, if harvestable, product is below ground) (g MJ−1m−2). | 0.85–1.60 | [32] |
| I50maxH | The maximum daily increase in I50B due to heat stress (°C d). | 120 | [7] |
| I50maxW | The maximum daily increase in I50B due to water stress (°C d). | 20 | [7] |
| MaxT | Threshold for daily Tmax to start accelerating senescence due to heat stress (°C). | 40 | [33] |
| ExtremeT | Daily Tmax threshold when RUE becomes 0 due to heat stress (°C). | 25–40 | [33] |
| CO2_RUE | Relative increase in RUE per 1 ppm elevated CO2 above 350 ppm. | 350–600 | [7] |
| S_Water | Sensitivity of RUE to drought stress (ARID index). | 0.6–0.96 | [7] |
| Input Variables | Description | Unit |
|---|---|---|
| Crop management | Sowing date (SowingDate) | - |
| Harvesting date (HarvestDate) | - | |
| Irrigation depth (Irri) | m | |
| Initial | Biomass (InitialBio) | Ton |
| Cumulative temperature (InitialTT) | °C day | |
| Solar radiation interception (InitialFsolar) | - | |
| Soil characteristics | Atmospheric CO2 concentration | ppm |
| Soil water-holding capacity (AWC) | - | |
| Runoff curve number (RCN) | - | |
| Deep drainage coefficient (DDC) | - | |
| Root zone depth (RZD) | m | |
| Weather | Daily maximum temperature (TMAX) | °C |
| Daily minimum temperature (TMIN) | °C | |
| Daily rainfall amount (RAIN) | m | |
| Daily solar radiation (SRAD) | MJ m−2 day−1 |
| Season Crop | Observation (t ha−1) | Simulation (t ha−1) | ||
|---|---|---|---|---|
| Biomass | Yield | Biomass | Yield | |
| 2020 | 4.801 | 2.101 | 5.005 | 1.852 |
| 2021 | 4.739 | 2.225 | 5.466 | 2.023 |
| Stages | Year | |
|---|---|---|
| 2020 | 2021 | |
| Growing period | 21 July to 24 October | 20 January to 17 April |
| Bloom | 20 August (31 DAP) | 18 February (30 DAP) |
| Seed fill | 2 September (44 DAP) | 28 February (40 DAP) |
| Mat | 12 October (84 DAP) | 7 April (78 DAP) |
| Harvest date | 25 October (96 DAP) | 17 April (88 DAP) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pham, Q.V.; Nguyen, T.T.N.; Vo, T.T.X.; Le, P.H.; Nguyen, X.T.T.; Duong, N.V.; Le, C.T.S. Applying the SIMPLE Crop Model to Assess Soybean (Glicine max. (L.) Merr.) Biomass and Yield in Tropical Climate Variation. Agronomy 2023, 13, 1180. https://doi.org/10.3390/agronomy13041180
Pham QV, Nguyen TTN, Vo TTX, Le PH, Nguyen XTT, Duong NV, Le CTS. Applying the SIMPLE Crop Model to Assess Soybean (Glicine max. (L.) Merr.) Biomass and Yield in Tropical Climate Variation. Agronomy. 2023; 13(4):1180. https://doi.org/10.3390/agronomy13041180
Chicago/Turabian StylePham, Quang V., Tanh T. N. Nguyen, Tuyen T. X. Vo, Phuoc H. Le, Xuan T. T. Nguyen, Nha V. Duong, and Ca T. S. Le. 2023. "Applying the SIMPLE Crop Model to Assess Soybean (Glicine max. (L.) Merr.) Biomass and Yield in Tropical Climate Variation" Agronomy 13, no. 4: 1180. https://doi.org/10.3390/agronomy13041180