
Evaluation of Inoculation Methods for Determination of Winter Wheat Resistance to Fusarium Head Blight
1
Faculty of Agriculture, University of Zagreb, Svetošimunska cesta 25, 10000 Zagreb, Croatia
2
Centre of Excellence for Biodiversity and Molecular Plant Breeding (CroP-BioDiv), Svetošimunska cesta 25, 10000 Zagreb, Croatia
3
Elsoms Ackermann Barley Ltd., Spalding PE11 1QG, Lincolnshire, UK
4
Bc Institute for Breeding and Production of Field Crops, Rugvica, Dugoselska 7, 10370 Dugo Selo, Croatia
*
Author to whom correspondence should be addressed.
Agronomy 2023, 13(4), 1175; https://doi.org/10.3390/agronomy13041175
Submission received: 9 March 2023
/
Revised: 27 March 2023
/
Accepted: 17 April 2023
/
Published: 20 April 2023
(This article belongs to the Special Issue Treatment and Management of Fusarium Disease in Wheat)
Abstract
:One of the most severe winter wheat (Triticum aestivum L.) diseases is Fusarium head blight (FHB). It is believed that selection for resistance to FHB is better in high disease pressure environments, for which various methods of artificial inoculation are used. The standard spray method of artificial inoculation is believed to be technically demanding and labour intensive. Therefore, scattering Fusarium-infected maize stalks onto trial plots after wheat emergence is suggested as a suitable alternative. The aim of this study was to compare the mean values and heritability of the visual rating index (VRI) and the percentage of Fusarium-damaged kernels (FDK) between the two abovementioned methods of artificial inoculation and natural infection, and to determine the phenotypic correlations between the three methods for the studied traits. The achieved levels of VRI and FDK were comparable for the two methods of artificial inoculation and considerably lower under natural conditions. Heritability for VRI ranged over four years from 0.68 to 0.91 for the spray method, from 0.73 to 0.95 for the infected maize stalks, and from 0.26 to 0.65 for natural infection, whereas for FDK it ranged from 0.56 to 0.85, 0.38 to 0.83, and 0.11 to 0.44 for the three inoculation methods, respectively. The strong positive correlation between the two applied methods of artificial inoculation for studied traits suggests that scattering infected maize stalks could serve as a reliable supplement for the technically and labor-intensive spray method of artificial inoculation.
1. Introduction
Fusarium head blight (FHB), caused by Fusarium graminearum (teleomorph Gibberella zeae (Schw.) Petch) and other Fusarium species, is currently one of the most devastating wheat diseases in the world. Symptoms of FHB infection typically include premature bleaching of the entire spike or just a few spikelets, pinkish-red mycelium and spores on infected spikelets, inhibited grain formation, and the development of shriveled, light-weighted, and discolored grain (from white to pink) as a result of mycelial outgrowth from Fusarium-colonized grain [1]. Yield reductions due to formation of shriveled grain, reductions in baking and seed quality, and mycotoxin contamination are major threats posed by FHB [2,3,4,5,6]. The frequency of FHB epidemics has increased in recent years in most of the world’s major wheat-growing regions [7]. Among different species causing FHB, F. graminearum is considered to be the most important globally, due to its widespread occurrence and aggressiveness [8]. It is ranked fourth among plant fungal pathogens based on its scientific and economic importance [9]. The major causal agents of FHB in Europe are F. graminearum, F. culmorum, F. avenaceum, and F. poae [1]. However, the prevalence of certain species varies from country to country and depends on meteorological conditions as well as agricultural practices, such as crop rotation, previous crop, tillage, and the susceptibility of cultivars [1,10,11,12,13].
Central to FHB infection and development are the abundance and aggressiveness of the inoculum during the vulnerable plant growth stage, which essentially spans several days around anthesis, environmental conditions during this critical period, and the susceptibility or resistance status of the plant [14]. Fungicide applications have been shown to be only partially effective in controlling FHB [4,15,16,17], and are only effective within a narrow time frame [18]. Therefore, host resistance has long been considered the most practical and effective means to control the disease [14,17,19]. However, breeding wheat for durable resistance to FHB has been hindered by a lack of effective resistance genes and by the complexity of resistance in identified sources. FHB resistance is a quantitative trait controlled by multiple genes with either major or minor effects and is significantly influenced by genotype-environment interactions [4,14,19,20]. The combined effect of multiple genes interacting in a complex manner in resistant germplasm and their often poor agronomic and quality performance complicates the utilization of this germplasm [17,21]. According to Mesterhazy et al. [5], despite a 100-year tradition of FHB resistance research and breeding, most cultivated varieties are susceptible or highly susceptible to the disease.
The development of resistant cultivars requires the identification of both resistant sources and resistant lines within breeding populations. Although no source of complete resistance is known and current sources provide only partial resistance, most breeders have found genetic variability for FHB resistance in their existing germplasm. The level of resistance in the adapted germplasm pool will increase if programs actively screen for FHB resistance [22]. Practical breeders are trying to improve FHB resistance by recombining different resistance sources and types while selecting for resistance and desirable agronomic performance. Their ultimate goal is to develop productive cultivars with low disease symptoms and low mycotoxin contamination despite high infection pressures [3].
Resistance testing is best performed under uniform and moderate to high disease pressure, which is typically achieved through artificial inoculation [14]. Common inoculation methods include point inoculation, conidia spray, and the grain-spawn method [22,23,24]. These methods, while effective in quantifying resistance, are time and labor intensive. As a result, programs often receive data on resistance in breeding material at only one location per year. In addition, some material, often less advanced, may remain untested due to limited resources for FHB resistance screening [23]. Alternatively, sowing in fields with maize stubble on the soil surface or sowing trials in natural hot spots for Fusarium infection have been used to provoke infection [3,25]. However, screening for host resistance through natural infection is difficult because disease incidence and severity are inconsistent due to changes in environmental conditions that are difficult to control, such as temperature and precipitation [23,26]. Epidemics of Fusarium head blight have long been considered to originate from an inoculum associated with non-decomposed residues from the previous crop, particularly of small-grain cereals and maize, which provide a site for abundant sporulation in the next growing season [27,28]. Field observations have shown that the severity of FHB and deoxynivalenol (DON) contamination of wheat grains is positively correlated with increasing quantities of maize residues [28]. However, there are limited studies reporting the effects of crop residues as an inoculum source in FHB resistance testing [25,29]. Xue et al. [29] investigated the influence of inoculum sources (conidial suspension, infested barley and maize kernels, or infested wheat debris) on FHB development and DON content. Their results showed that inoculation with infested debris as a natural source of inoculum caused fewer FHB symptoms and lower DON contamination of grains compared to the conidial suspension or infested kernels. In the study by Mesterhazy et al. [25], the spray inoculation and polyethylene bag cover method showed better differentiation among wheat genotypes in terms of visual FHB rating, Fusarium-damaged kernels (FDK), and DON content in grain than method of spray inoculation and mist irrigation or the maize stalks method supported by mist irrigation. Regarding visual FHB ratings, all three methods showed similar ranking of genotypes, while the results for FDK and DON content were less consistent among methods.
In the Bc Institute’s winter wheat breeding program, screening of elite breeding lines for FHB resistance using the spray method of inoculation with Fusarium isolates serves as efficient criterion for final selection for registration purposes [30,31,32]. In the present study, an alternative inoculation method for screening FHB resistance in wheat, known as deployment of Fusarium-infected maize stalks, was compared with the standard spray method and natural infection. The objectives of the study were (1) to compare three inoculum sources in terms of their efficacy in inducing FHB symptoms, (2) to determine the relationship between FHB scores within the methods, and (3) to estimate the heritability of investigated traits.
2. Materials and Methods
2.1. Description of Field Experiments for Evaluation of Fusarium Head Blight Resistance
Field experiments including 25 winter wheat genotypes were conducted over four growing seasons (2011–2014) at the Bc Institute’s winter wheat breeding nursery in Botinec (Zagreb), Croatia. Each year, a different set of the 18 most promising elite breeding lines and seven controls with varying levels of FHB resistance were evaluated in separate experiments conducted using two methods of artificial inoculation with Fusarium graminearum and natural Fusarium infection. Among the controls, the cultivars Renan, Roazon, Poncheau, Žitarka, and Lucija were repeated throughout all years of the study, while the control cultivars Apache, Soissons, and resistant breeding lines (D48X42X6)2 and K9_21_AB.14 were included in some years. The soil type at the experimental location was loamy clay. In all four years, sowing was done in late October. The experiments were set up as randomized complete blocks with four replicates. The experimental plots consisted of three 1 m long rows with 25 cm of in-between row spacing. The sowing density was 80 seeds per row. Each year, standard agronomic practices for intensive winter wheat production were used. In the autumn before ploughing, nitrogen (N), phosphorus, and potassium (7:20:30) in the amount of 300 kg ha−1 and UREA (46% N) at a rate of 150 kg ha−1 were added to the soil. Calcium ammonium nitrate (CAN, 27% N) was applied as a top-dressing at the beginning of wheat tillering (185 kg ha−1) and in the phase of intensive stem elongation, when the first or second node was detectable on the main stem in most genotypes (110 kg ha−1). For weed control, 0.8 g L−1 of Pinoxaden (50 g L−1) and Fluroxypyr (360 g L−1) were applied when the majority of genotypes developed first node on the main stem. Fungal diseases were treated with Chlorothalonil (400 g L−1) and Azoxystrobin (80 g L−1) at a rate of 2.5 L ha−1 at the time of the emergence of flag leaves in most genotypes.
2.2. Inoculum Production and Inoculation Procedure
The first method of inoculation was the spray method, for which the Fusarium inoculum was developed using the “bubble breeding” method proposed by Mesterházy in 1977 [33]. Each year, new isolates were prepared and used for artificial inoculation by the spray method. The fungus F. graminearum was isolated from infected wheat grains from the previous year, which came from wheat genotypes with a high FDK score. Different strains of Fusarium spp. were isolated from 36 infected wheat grains on PDA medium. The 12 best isolates were selected visually, and based on the appearance of spores under the microscope, their membership to the species F. graminearum was confirmed according to the identification keys of Nelson et al. [34]. The aggressiveness test according to Mesterházy [35] was performed on these 12 isolates (Figure S1). The four most aggressive isolates were selected, and liquid inoculum was prepared from them for the spray method immediately before starting artificial inoculation. The concentrations of the inoculum were adjusted to 500,000 spores per ml using a hemocytometer. Equal volumes of liquid inoculum from four isolates were mixed. The liquid inoculum was stored at 4 °C during the inoculation period. The first inoculation of each plot was performed when 50% of the plants were at anthesis. The second inoculation followed two days later. The wheat spikes were sprayed using a backpack-carried manual sprayer early in the morning, and 40 mL of inoculum was applied to each plot in both sprays. The second method of inoculation was conducted using infected maize stalks collected in an infected maize field from a maize inbred line susceptible to F. graminearum. The stalks were cut into 20–30 cm long pieces and scattered on the soil surface (15–20 pieces per experimental plot) in late autumn when the first two leaves emerged on plants (Figure 1). The third type of infection occurred under natural conditions, i.e., no artificial inoculation was performed. In this study, no mist irrigation was applied for either artificial inoculation or natural infection.
2.3. Fusarium Head Blight Evaluation
The percentage of visually infected spikelets, referred to as the visual rating index (VRI), was estimated on a sample of approximately 100 spikes according to a linear scale from 0 to 100% (Table S1). Disease symptoms were assessed 21 and 25 days after spray inoculation of each genotype and finally expressed as the mean of the two readings. The percentage of Fusarium-damaged kernels (FDK) was determined on ten randomly selected spikes taken from each experimental plot after harvest. The spikes were threshed by hand and the Fusarium-damaged and normal kernels were counted. Only the pinkish white-colored grains along with the slightly infected whitish powdered kernels were considered as Fusarium-damaged, while the normally colored but shriveled kernels were not considered [25].
2.4. Statistical Analysis
An analysis of variance (ANOVA) was performed across inoculation methods for each year as well as for each year × inoculation method combination for two FHB ratings (VRI and FDK) using the PROC GLM of SAS/STAT [36]. The components of variance for the second ANOVA were obtained by equating the observed mean squares from the ANOVA to their expectations and solving for the desired variance components. Genotypic variance (σ2G) was calculated as (genotype mean square—error mean square)/r, where r is the number of replicates, and the error variance (σ2ε) is equal to the error mean square. Heritability on a plot mean basis was estimated using the equation: h2 = σ2G/(σ2G + σ2ε). Spearman’s rank correlation coefficients between inoculation methods for two FHB ratings (VRI and FDK) as well as between VRI and FDK within each of the three inoculation methods were calculated using a PROC CORR of the SAS/STAT [36].
3. Results
3.1. Analysis of Variance and Heritability
An analysis of variance across 25 wheat genotypes and three inoculation methods (Table 1) revealed a significant effect of inoculation method (IM) and genotype (G), and a significant IM × G interaction for visual rating index (VRI) and Fusarium-damaged kernels (FDK) in all four years. For the VRI, IM and G had similar effects in 2011, 2012 and 2013, explaining 34 to 39% and 32 to 38% of the total sum of squares (SS) respectively, while the IM × G interaction accounted for 18 to 24% of the total SS. In 2014, variation in VRI was predominantly explained by G (66% of SS), while IM and the IM × G interaction accounted for only 15 and 10% of the total SS, respectively. The variation of FDK in all years was explained primarily by IM, which contributed between 36 and 56% of the total SS, whereas G and IM × G interactions contributed from 20 to 26% and 14 to 19%, respectively. Considering the two methods of artificial inoculation (spray and maize stalk), the ANOVA showed a significant effect of all sources of variation for VRI and FDK, except for IM × G for VRI in 2011 and for FDK in 2014 (Table 1). However, the effect of G, which explained 69 to 88% of SS for VRI and 51 to 70% of SS for FDK, was much larger than in an analysis of all three inoculation methods. Consequently, the contribution of IM for both traits and the IM × G interaction for FDK was of a much smaller magnitude.
Heritability estimates for the two FHB scores under different inoculation methods for each respective set of 25 genotypes evaluated in 2011, 2012, 2013, and 2014 are shown in Table 2. For VRI, heritability over four years varied from 0.68 to 0.91 for the spray method, and from 0.73 to 0.95 for the infected maize stalk method. The estimated heritability in 2011 and 2013 was similar between the two methods. In 2012, it was higher for the spray method, and in 2014, it was higher for infected maize stalks. The heritabilities for VRI under natural infection were much lower compared to the heritabilities of the two methods of artificial inoculation, ranging from 0.26 to 0.65. For FDK, the heritability varied from 0.56 to 0.85 for the spray method, from 0.38 to 0.83 for the infected maize stalks, and from 0.11 to 0.44 for the natural infection. Heritability estimates in 2011 and 2012 were comparable between VRI and FDK for the three methods, and in 2013 and 2014 they were considerably lower for FDK, especially under the maize stalk inoculation and natural infection.
3.2. FHB Scores under Different Inoculation Methods
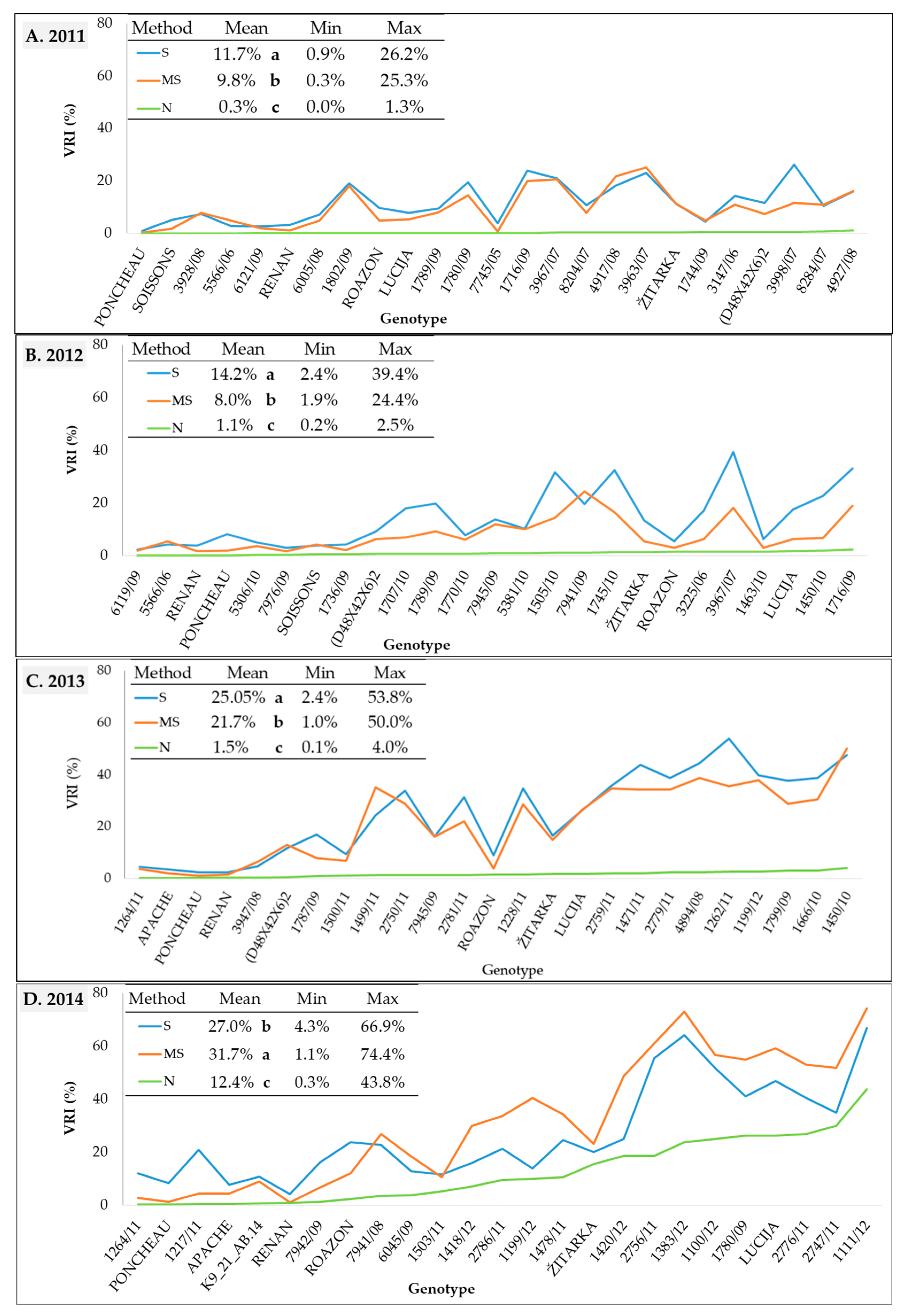
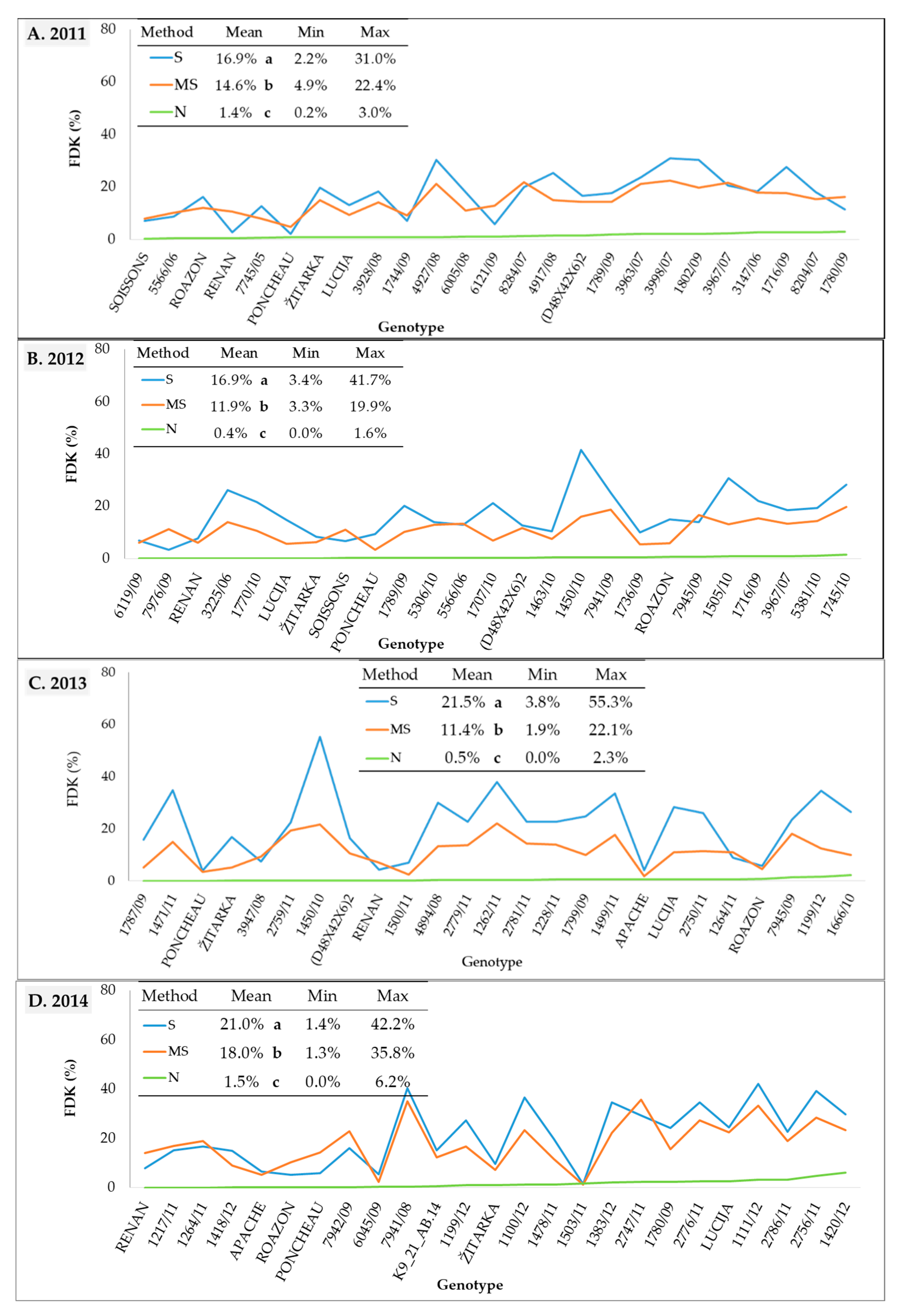
The visual rating index (VRI) varied significantly among genotypes for all three inoculation methods in all four growing seasons (Figure 2, Table S2). The highest mean VRI values were obtained using spray inoculation (S) in all years, except in 2014 when inoculation with maize stalk (MS) resulted in a significantly higher VRI value. Under natural conditions of infection (N), VRI values were, as expected, the lowest. Despite significant differences between mean VRI values for spray and maize stalk inoculation, VRI values of the two methods were comparable in 2011, 2013, and 2014, with respective mean values of 11.7 and 9.8%, 25.05 and 21.7%, and 27.0 and 31.7%. In the same years, the range of VRI scores was also similar for the two methods of artificial inoculation. The higher mean VRI in 2014 for the maize stalk inoculation method compared to the spray inoculation method primarily resulted from the higher VRI values observed in less resistant genotypes (Figure 2D). In the same year, VRIs under natural conditions was at least ten times higher than in the three previous growing seasons (12.4% in 2014, 1.5% in 2013, 1.1% in 2012, and 0.3% in 2011). Mean FDK values in all four years were significantly higher for the spray method compared to maize stalk method (Figure 3, Table S3). The difference between the mean FDK scores of the spray inoculation and maize stalk inoculation methods, as well as the difference between their ranges was most pronounced in 2013, with means of 21.5 and 11.4% and ranges from 3.8 to 55.3% and 1.9 to 22.1%, respectively. In 2011, 2012, and 2014, the spray and maize stalk inoculations produced comparable FDKs with mean values of 16.9 and 14.6%, 16.9 and 11.9%, and 21.0 and 18.0%. Under natural conditions (N), FDK values were considerably lower than for the two methods of artificial inoculation.
3.3. Correlations
The correlations between inoculation methods for visual rating index (VRI) and Fusarium-damaged kernels (FDK) in the four years of the study are shown in Table 3. The VRIs of the two methods of artificial inoculation (spray and maize stalk) were strongly positively correlated with coefficients of correlation ranging from 0.88 in 2012 to 0.94 in 2013. On the other hand, the correlation between VRI under natural infection and VRI under two types of artificial inoculation was moderately positive in 2011 and 2012, and strong and positive in 2013 and 2014. In 2012, for VRI, the correlation between natural infection and spray inoculation was slightly higher than between natural infection and maize stalk inoculation (0.72 vs. 0.57), whereas in 2014 the corresponding correlation coefficients were 0.83 and 0.93.
The FDK values for the two methods of artificial inoculation were strongly positively correlated, with the correlation coefficients ranging from 0.60 to 0.87. In 2011, the correlation of FDK scores between natural infection and maize stalk inoculation was slightly higher than the corresponding correlation between natural infection and spray inoculation (0.74 vs. 0.54), the correlations of FDK scores were similar in 2012, and the opposite was true in 2014 (0.49 vs. 0.62). In 2013, the correlations of FDK scores between natural infection and both methods of artificial inoculation were not significant.
The correlations between VRI and FDK were moderate to strong for the two artificial inoculation methods in the four years of study, with coefficients ranging from 0.77 to 0.89 and 0.55 to 0.86 for spray inoculation and maize stalk inoculation, respectively (Table 4). The correlation coefficients were similar for the two methods of artificial inoculation in 2011, 2012, and 2013, while the correlation coefficient was slightly stronger for the spray method in 2014. In natural infection compared to the two artificial inoculation methods, the correlation between VRI and FDK was generally weaker, except in 2014, when it was at the level observed for spray inoculation.
4. Discussion
For routine screening of FHB resistance in large breeding populations, the faster, cheaper, and more reliable inoculation method is preferable [2]. In the present study, an alternative inoculation method for screening FHB resistance in wheat, known as deployment of Fusarium-infected maize stalks, was evaluated over four consecutive years. The standard spray method and natural infection were used as controls. As expected, considerably more FHB symptoms in terms of both VRI and FDK were observed under the two artificial inoculation methods compared to natural infection. Although mean VRI and FDK values were generally significantly higher for spray than for maize stalk inoculation, the two inoculation methods resulted in comparable mean values and ranges for both FHB-related traits in three of four study years (Figure 1 and Figure 2). Similar to our study, Mesterhazy et al. [25] compared two variants of spray inoculation with inoculation using maize stalks and reported slightly higher mean FHB scores for the maize stalk method than for the standard spray method, similar mean FDK scores for the two spray methods, and considerably lower deoxynivalenol (DON) grain content for maize stalk method. In their study, the spray method with bags resulted in higher levels of symptoms for all three studied traits compared to the standard spray method and maize stalk method. The authors pointed out that differentiation of genotypes in resistance was more secure at higher levels of FHB symptoms. In this sense, the similar levels of FHB symptoms for VRI and FDK observed for standard spray inoculation and maize stalk inoculation in the present study indicate the suitability of both methods for screening FHB resistance. An analysis of variance in the present study showed that the inoculation method, genotype, and the inoculation method × genotype interaction had a significant effect on VRI and FDK. Mesterhazy et al. [25] also reported a significant inoculation method × genotype interaction for the FHB score and the FDK and DON content in grain, although the interaction effect was significantly weaker than the main effect of genotype for all of the traits studied. In the present study, the contribution of the inoculation method × genotype interaction effect to the total phenotypic variability for VRI was also much smaller compared to the main genotype effect (Table 1). On the other hand, the contributions of genotype and inoculation method × genotype interaction to the total phenotypic variability for FDK were similar. However, when considering only spray and maize stalk inoculations, the effect of genotype for both VRI and FDK was much larger than the effect of the method × genotype interaction. This suggests that the observed magnitude of the inoculation method × genotype interaction in the present study was mainly due to the method of natural infection. Engle et al. [37] also found a significant effect of the interaction between the genotype and inoculation technique for FHB severity in an experiment in which four methods of artificial inoculation of ears with F. graminearum were applied to seven wheat genotypes. Similarly, Miedaner et al. [2] found a significant inoculation × genotype interaction for visual FHB symptoms when comparing point and spray inoculation in wheat and concluded that the genotype-specific response to a particular method may be the result of different contributions of individual components (such as type I and II resistance) to the overall FHB resistance. The available literature shows that not only FHB resistance itself, but also individual FHB resistance components are quantitatively inherited and are often under the control of resistance component-specific QTLs [3,14,20,38,39,40,41].
The magnitude of the method × genotype interaction effects over the four years of the present study was reflected in the strength of phenotypic correlations found between the inoculation methods studied. The VRI scores of the two methods of artificial inoculation (spray and maize stalk) were strongly positively correlated with correlation coefficients ranging from 0.88 to 0.94, whereas the correlations of FDK scores were somewhat lower, ranging from 0.60 to 0.87. The correlations of the two methods of artificial inoculation and natural infection were strong and positive for VRI in 2013 and 2014, and were moderate and positive in 2011 and 2012. For FDK, the respective correlations over four years of the study were weak to moderate. In agreement with our results, Mesterhazy et al. [25] also found a higher positive correlation between the standard spray method and the maize stalk method for the FHB score than for FDK (0.73 vs. 0.63, respectively). Miedaner et al. [2] reported lower phenotypic correlation coefficients between spray and point inoculation methods for the percentage of infected spikelets (0.40) and the relative spike weight (0.52). A wide range in the strength of correlations between different inoculation methods for FHB traits observed in the present and previous studies is reflected in the agreement or disagreement in the ranking of genotypes, and suggests that certain genotypes have a specific type of resistance, while others combine different levels of multiple types of resistance. Therefore, the temporal and/or spatial combination of different inoculation methods could provide complementary information on genotype resistance to FHB. However, environmental factors such as temperature, precipitation, and relative humidity cannot be ruled out, as they may affect the development of disease symptoms to different degrees for different inoculation methods.
Another important aspect in evaluating the reliability of an inoculation method is the correlation between the various FHB-associated traits and their heritability for a given method. From a practical point of view, visual evaluation of FHB symptoms on spikes is less laborious and time-consuming than evaluation of Fusarium-damaged kernels (FDK) and is preferred by breeders. In the present study, the correlation coefficients between VRI and FDK were moderately to strongly positive for the two methods of artificial inoculation, ranging from 0.77 to 0.89 for spray inoculation and 0.55 to 0.86 for maize stalk inoculation, whereas they were much lower for natural infection in three of four study years. Consistent with our results, correlations between FHB severity and FDK in the study of Kubo et al. [42] ranged from 0.78 to 0.81 over a three-year field experiment including 31 wheat cultivars inoculated with F. graminearum. Similar values of correlation coefficients between the two FHB ratings were reported by Mesterhazy [43], who examined the responses of 19 wheat genotypes to seven isolates of F. graminearum and F. culmorum (0.74), and Goral et al. [44], who evaluated 27 wheat lines inoculated with F. culmorum (0.78). On the other hand, He et al. [39] found a much lower correlation between FHB symptoms on spikes and FDK in a RIL population of wheat inoculated with F. graminearum, ranging from 0.29 to 0.30 over three years. In addition to the two visual assessments of FHB symptoms, the content of DON is commonly used as an indicator of mycotoxin contamination of grains, but measurement of DON is expensive and impractical for routine breeding practices. Therefore, the extent of correlations between visual FHB ratings and DON content is critical to know for the use of visual FHB ratings as indirect criteria in selecting for low DON content. In several studies, FDK has been shown to be a better predictor of grain DON contamination than visual symptoms on spikes [5,20,25,39,43,45], making FDK a preferred indirect trait when selecting for low DON content.
In the present study, heritability for VRI over four years ranged from 0.68 to 0.91 for the spray method and from 0.73 to 0.95 for the infected maize stalks, whereas it was much lower for natural infection, ranging from 0.26 to 0.65. On the other hand, heritability estimates for FDK compared to VRI were slightly lower for spray inoculation (0.56 to 0.85), moderately lower for maize stalk inoculation (0.38 to 0.83), and much lower for natural infection (0.11 to 0.44). Miedaner et al. [2] reported that the heritability estimates were higher for spray inoculation than for point inoculation, as measured by the percentage of infected spikelets (0.81 vs. 0.77) and relative head weight (0.77 vs. 0.52). These findings are consistent with our study because we found that the differences between methods were greater for the heritability of FHB symptoms estimated on kernels than for the FHB symptoms associated with spikes. Zhang et al. [20] found similar ranges of heritability for FHB symptoms on spikes, FDK, and DON content, ranging from 0.55 to 0.85 in different environments. Moderate to high heritability of FHB resistance components was also reported in some other studies [39,46,47], suggesting that genetic variation plays a major role in the phenotypic variation of FHB-related traits.
5. Conclusions
The inoculation method using the infected maize stalks resulted in fewer FHB symptoms than the spray inoculation method, but from a practical point of view, the differentiation of genotypes according to their resistance was successful, especially for the more practical and quicker visual evaluation. This inoculation method does not require laboratory equipment or inoculum production. Resistance trials can be conducted at locations which are more distant from the laboratory, and flowering time recordings are not required. In this context, maize stalks can also be used as an alternative source of inoculum in early generations of selection when a large number of genotypes with a wide range of flowering times are evaluated. This method also simulates frequent production practice, especially on small family husbandries, where winter wheat is grown after maize and where large amounts of maize debris on the soil surface serves as a good source of inoculation. FHB severity under natural conditions was significantly lower than the FHB severity observed under the two methods of artificial inoculation analysed in the present study, and was largely dependent on the environmental conditions. Nevertheless, the resistance levels determined under natural conditions served as a good control for the resistance levels determined by the two methods of artificial inoculation.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/agronomy13041175/s1, Table S1: Scale for the assessment of visual rating index (% of diseased spikelets per plot); Table S2: Visual rating index (VRI, %) for 25 wheat genotypes under three inoculation methods (spray, maize stalk and natural) in 2011, 2012, 2013, and 2014; Table S3: Fusarium-damaged kernels (FDK, %) for 25 wheat genotypes under three inoculation methods (spray, maize stalk and natural) in 2011, 2012, 2013, and 2014; Figure S1. Aggressiveness test with 12 different isolates of F. graminearum.
Author Contributions
Conceptualization, M.M., M.B. and H.Š.; methodology, M.M. and A.L.; software, M.B. and H.Š.; validation, H.Š., M.M. and M.B.; formal analysis, M.B. and H.Š.; investigation, M.M. and A.L.; resources, M.M.; data curation, M.M., M.B. and H.Š.; writing—original draft preparation, H.Š.; writing—review and editing, M.B., M.M. and A.L.; visualization, H.Š. and M.B.; supervision, H.Š. and M.B.; project administration, M.M. and A.L.; funding acquisition, M.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the program for the development of new wheat cultivars at the Bc Institute for Breeding and Production of Field Crops.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Birr, T.; Hasler, M.; Verreet, J.A.; Klink, H. Composition and predominance of Fusarium species causing Fusarium head blight in winter wheat grain depending on cultivar susceptibility and meteorological factors. Microorganisms 2020, 8, 617. [Google Scholar] [CrossRef] [PubMed]
- Miedaner, T.; Moldovan, M.; Ittu, M. Comparison of spray and point inoculation to assess resistance to Fusarium head blight in a multienvironment wheat trial. Phytopathology 2003, 93, 1068–1072. [Google Scholar] [CrossRef] [PubMed]
- Buerstmayr, H.; Ban, T.; Anderson, J.A. QTL mapping and marker assisted selection for Fusarium head blight resistance in wheat: A review. Plant Breed. 2009, 128, 1–26. [Google Scholar] [CrossRef]
- McMullen, M.; Bergstrom, G.; De Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A Unified effort to fight an enemy of wheat and barley: Fusarium head blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef]
- Mesterházy, A.; Varga, M.; György, G.; Lehoczki-Krsjak, S.; Tóth, B. The role of adapted and non-adapted resistance sources in breeding resistance of winter wheat to Fusarium head blight and deoxynivalenol contamination. World Mycotoxin J. 2018, 11, 539–557. [Google Scholar] [CrossRef]
- Martínez, M.; Ramírez Albuguerque, L.; Arata, F.A.; Biganzoli, F.; Fernández Pinto, V.; Stenglein, S.A. Effects of Fusarium graminearumand Fusarium poae on disease parameters, grain quality and mycotoxins contamination in bread wheat (Part I). J. Sci. Food Agric. 2020, 100, 863–873. [Google Scholar] [CrossRef]
- Steiner, B.; Buerstmayr, M.; Michel, S.; Schweiger, W.; Lemmens, M.; Buerstmayr, H. Breeding strategies and advances in line selection for Fusarium head blight resistance in wheat. Trop. Plant Pathol. 2017, 42, 165–174. [Google Scholar] [CrossRef]
- Beccari, G.; Arellano, C.; Covarelli, L.; Tini, F.; Sulyok, M.; Cowger, C. Effect of wheat infection timing on Fusarium head blight causal agents and secondary metabolites in grain. Int. J. Food Microbiol. 2019, 290, 214–225. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J. The top 10 fungal pathogens in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Waalwijk, C.; Kastelein, P.; Vries, I.; Kerény, Z.; van der Lee, T.; Hesselnik, T.; Köhl, J.; Kema, G. Major changes in Fusarium spp. in wheat in the Netherlands. Eur. J. Plant Pathol. 2003, 109, 743–754. [Google Scholar] [CrossRef]
- Xu, X.M.; Parry, D.W.; Nicholson, P.; Thomsett, M.A.; Simpson, D.; Edwards, S.G.; Cooke, B.M.; Doohan, F.M.; Brennan, J.M.; Moretti, A.; et al. Predominance and association of pathogenic fungi causing Fusarium ear blightin wheat in four European countries. Eur. J. Plant Pathol. 2005, 112, 143–154. [Google Scholar] [CrossRef]
- Cosic, J.; Jurkovic, D.; Vrandecic, K.; Simic, B. Pathogenicity of Fusarium species to wheat and barley ears. Cereal Res. Commun. 2007, 35, 529–532. [Google Scholar] [CrossRef]
- Spanic, V.; Lemmens, M.; Drezner, G. Morphological and molecular identification of Fusarium species associated with head blight on wheat in east Croatia. Eur. J. Plant Pathol. 2010, 128, 511–516. [Google Scholar] [CrossRef]
- Buerstmayr, M.; Steiner, B.; Buerstmayr, H. Breeding for Fusarium head blight resistance in wheat-progress and challenges. Plant Breed. 2020, 139, 429–454. [Google Scholar] [CrossRef]
- Mesterházy, A.; Toth, B.; Varga, M.; Bartok, T.; Szabo-Hever, A.; Farady, L.; Lehoczki-Krsjak, S. Role of fungicides, application of nozzle types, and the resistance level of wheat varieties in the control of Fusarium head blight and deoxynivalenol. Toxins 2011, 3, 1453–1483. [Google Scholar] [CrossRef]
- Willyerd, K.T.; Li, C.; Madden, L.V.; Bradley, C.A.; Bergstrom, G.C.; Sweets, L.E.; Paul, P.A. Efficacy and stability of integrating fungicide and cultivar resistance to manage Fusarium head blight and deoxynivalenol in wheat. Plant Dis. 2012, 96, 957–967. [Google Scholar] [CrossRef]
- Gilbert, J.; Haber, S. Overview of some recent research developments in Fusarium head blight of wheat. Can. J. Plant Pathol. 2013, 35, 149–174. [Google Scholar] [CrossRef]
- Paul, P.A.; McMullen, M.P.; Hershman, D.E.; Madden, L.V. Meta-analysis of the effects of triazole-based fungicides on wheat yield and test weight as influenced by Fusarium head blight intensity. Phytopathology 2010, 100, 160–171. [Google Scholar] [CrossRef]
- Bai, G.; Shaner, G. Scab of wheat: Prospects for control. Plant Dis. 1994, 78, 760–766. [Google Scholar] [CrossRef]
- Zhang, W.; Boyle, K.; Brûlé-Babel, A.L.; Fedak, G.; Gao, P.; Robleh Djama, Z.; Polley, B.; Cuthbert, R.D.; Randhawa, H.S.; Jiang, F.; et al. Genetic characterization of multiple components contributing to Fusarium head blight resistance of FL62R1, a Canadian bread wheat developed using systemic breeding. Front. Plant Sci. 2020, 11, 580833. [Google Scholar] [CrossRef]
- Miedaner, T.; Korzun, V. Marker-assisted selection for disease resistance in wheat and barley breeding. Phytopathology 2012, 102, 560–566. [Google Scholar] [CrossRef] [PubMed]
- Rudd, J.C.; Horsley, R.D.; McKendry, A.L.; Elias, E.M. Host plant resistance genes for Fusarium head blight: Sources, mechanisms and utility in conventional breeding systems. Crop Sci. 2001, 41, 620–627. [Google Scholar] [CrossRef]
- Thomson, C.J. Evaluation of An Inoculation Method and Quantitative Trait Loci for Fusarium Head Blight Resistance in Wheat. Master’s Thesis, University of Illinois at Urbana-Champaign, Champaign, IL, USA, 2010. [Google Scholar]
- Imathiu, M.; Edwards, S.G.; Ray, R.V.; Back, M. Artificial inoculum and inoculation techniques commonly used in the investigation of Fusarium head blight in cereals. Acta Phytopathol. Hung. 2014, 49, 129–139. [Google Scholar] [CrossRef]
- Mesterházy, A.; Lehoczki-Krsjak, S.; Varga, M.; Szabo-Hever, A.; Tóth, B.; Lemmens, M. Breeding for FHB resistance via Fusarium damaged kernels and deoxynivalenol accumulation as well as inculation methods in winter wheat. Agric. Sci. 2015, 6, 970–1002. [Google Scholar] [CrossRef]
- Mesterházy, A.; Bartók, T.; Lamper, C.S. Influence of cultivar resistance, epidemic severity, and Fusarium species on the efficacy of fungicide control of Fusarium head blight in wheat and deoxynivalent (DON) contamination of grain. Plant Dis. 2003, 87, 1107–1115. [Google Scholar] [CrossRef]
- Maiorano, A.; Blandino, M.; Reyneri, A.; Vanara, F. Effects of maize residues on the Fusarium spp. infection and deoxynivalenol (DON) contamination of wheat grain. Crop. Prot. 2008, 27, 182–188. [Google Scholar] [CrossRef]
- Blandino, M.; Pilati, A.; Reyneri, A.; Scudellari, D. Effect of maize crop residue density on Fusarium head blight and on deoxynivalenol contamination of common wheat grains. Cereal Res. Commun. 2010, 38, 550–559. [Google Scholar] [CrossRef]
- Xue, A.G.; Butler, G.; Voldeng, H.D.; Fedak, G.; Savard, M.E. Comparison of the influence of inoculum sources on the development of Fusarium head blight and the deoxynivalenol content in spring wheat in a disease nursery. Can. J. Plant Pathol. 2006, 28, 152–159. [Google Scholar] [CrossRef]
- Tomasović, S.; Palaveršić, B.; Ikić, I. Study of wheat lines resistance to Fusarium head blight. Cereal Res. Commun. 2005, 33, 583–588. [Google Scholar] [CrossRef]
- Tomasovic, S.; Palaversic, B.; Mlinar, R.; Ikic, I.; Sarcevic, H. Breeding winter wheat for yield and Fusarium head blight resistance in the Zagreb Bc Institute. Cereal Res. Commun. 2008, 36 (Suppl. B), 179–180. [Google Scholar]
- Tomasović, S.; Palaveršić, B.; Ikić, I.; Mlinar, R.; Šarčević, H.; Jukić, K.; Ivanušić, T. Latest results in breeding winter wheat for resistance to Fusarium head blight in the Zagreb Bc Institute. Plant Breed. Seed Sci. 2011, 64, 141–145. [Google Scholar] [CrossRef]
- Mesterházy, Á. The effect of inoculation method on the expression of symptoms caused by Fusarium graminearum Schwabe on wheat in seedling stage. In Symposium on Current Topics in Plant Pathology; Kiadó, A., Ed.; Academic Publishing House of the Hungarian Academy of Sciences: Budapest, Hungary, 1975; issued 1977; pp. 223–232. [Google Scholar]
- Nelson, P.E.; Toussoun, T.A.; Marasas, W.F.O. Section Discolor. In Fusarium Species: An Illustrated Manual for Identification; Pennsylvania State University Press: University Park, PA, USA, 1983; pp. 107–123. [Google Scholar]
- Mesterházy, Á. A laboratory method to predict pathogenicity of Fusarium graminearum in field and resistance of wheat to scab. Acta Phytopathol. Hung. 1984, 19, 205–218. [Google Scholar]
- SAS Institute Inc. SAS/STAT 9.4 Users Guide; SAS Inc.: Cary, NC, USA, 2013. [Google Scholar]
- Engle, J.S.; Madden, L.V.; Lipps, P.E. Evaluation of inoculation methods to determine resistance reactions of wheat to Fusarium graminearum. Plant Dis. 2003, 87, 1530–1535. [Google Scholar] [CrossRef] [PubMed]
- Buerstmayr, H.; Lemmens, M.; Hartl, L.; Doldi, L.; Steiner, B.; Stierschneider, M.; Ruckenbauer, P. Molecular mapping of QTLs for Fusarium head blight resistance in spring wheat. I. Resistance to fungal spread (Type II resistance). Theor. Appl. Genet. 2002, 104, 84–91. [Google Scholar] [CrossRef]
- He, X.; Dreisigacker, S.; Singh, R.P.; Singh, P.K. Genetics for low correlation between Fusarium head blight disease and deoxynivalenol (DON) content in a bread wheat mapping population. Theor. Appl. Genet. 2019, 132, 2401–2411. [Google Scholar] [CrossRef]
- Mesterházy, A. Updating the breeding philosophy of wheat to Fusarium head blight (FHB): Resistance Components. Plants 2020, 9, 1702. [Google Scholar] [CrossRef]
- Goddard, R.; Steed, A.; Scheeren, P.L.; Maciel, J.L.N.; Caierão, E.; Torres, G.A.M.; Consoli, L.; Santana, F.M.; Fernandes, J.M.C.; Simmonds, J.; et al. Identification of Fusarium head blight resistance loci in two Brazilian wheat mapping populations. PLoS ONE 2021, 16, e0248184. [Google Scholar] [CrossRef]
- Kubo, K.; Kawada, N.; Nakajima, T.; Kazuyuhi, H.; Fujita, M. Field evaluation of resistance to kernel infection and mycotoxin accumulation caused by Fusarium head blight in western Japanese wheat (Triticum aestivum L.) cultivars. Euphytica 2014, 200, 81–93. [Google Scholar] [CrossRef]
- Mesterházy, A.; Bartók, T.; Mirocha, C.M.; Komoróczy, R. Nature of resistance of wheat to Fusarium head blight and deoxynivalenol contamination and their consequences for breeding. Plant Breed. 1999, 118, 97–110. [Google Scholar] [CrossRef]
- Goral, T.; Wisniewska, H.; Ochodzki, P.; Nielsen, L.K.; Walentyn-Goral, D.; Stepien, L. Relationship between Fusarium head blight, kernel damage, concentration of Fusarium biomass, and Fusarium toxins in grain of winter wheat inoculated with Fusarium culmorum. Toxins 2018, 11, 2. [Google Scholar] [CrossRef]
- Bai, G.; Plattner, R.; Desjardins, A.; Kolb, F.; McIntosh, R.A. Resistance to Fusarium head blight and deoxynivalenol accumulation in wheat. Plant Breed. 2001, 120, 1–6. [Google Scholar] [CrossRef]
- Buerstmayr, H.; Steiner, B.; Lemmens, M.; Ruckenbauer, P. Resistance to Fusarium head blight in winter wheat: Heritability and trait associations. Crop Sci. 2000, 40, 1012–1018. [Google Scholar] [CrossRef]
- Lu, Q.; Lillemo, M.; Skinnes, H.; He, X.; Shi, J.; Ji, F.; Dong, Y.; Bjørnstad, Å. Anther extrusion and plant height are associated with Type I resistance to Fusarium head blight in bread wheat line ‘Shanghai-3/Catbird’. Theor. Appl. Genet. 2013, 126, 317–334. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Maize stalk residue deployed in the field at the location of Botinec in 2011.

Figure 2.
Visual rating index (VRI) in 25 wheat genotypes under spray inoculation (S), maize stalk inoculation (MS), and natural infection (N) in the (A) 2011, (B) 2012, (C) 2013, and (D) 2014 growing seasons. Means of the inoculation methods followed by the different letter are significantly different according to LSD test at p < 0.05.
Figure 2.
Visual rating index (VRI) in 25 wheat genotypes under spray inoculation (S), maize stalk inoculation (MS), and natural infection (N) in the (A) 2011, (B) 2012, (C) 2013, and (D) 2014 growing seasons. Means of the inoculation methods followed by the different letter are significantly different according to LSD test at p < 0.05.

Figure 3.
Fusarium-damaged kernels (FDK) in 25 wheat genotypes under spray inoculation (S), maize stalk inoculation (MS), and natural infection (N) in the (A) 2011, (B) 2012, (C) 2013, and (D) 2014 growing seasons. Means of the inoculation methods followed by the different letter are significantly different according to LSD test at p < 0.05.
Figure 3.
Fusarium-damaged kernels (FDK) in 25 wheat genotypes under spray inoculation (S), maize stalk inoculation (MS), and natural infection (N) in the (A) 2011, (B) 2012, (C) 2013, and (D) 2014 growing seasons. Means of the inoculation methods followed by the different letter are significantly different according to LSD test at p < 0.05.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Analysis of variance for the visual rating index and Fusarium-damaged kernels in four years across 25 genotypes and three inoculation methods (A) and two inoculation methods (B).
Table 1.
Analysis of variance for the visual rating index and Fusarium-damaged kernels in four years across 25 genotypes and three inoculation methods (A) and two inoculation methods (B).
| A. Three Inoculation Methods | B. Two Inoculation Methods a | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Year | Inoculation Method (IM) | Genotype (G) | IM × G | Error | Inoculation Method (IM) | Genotype (G) | IM × G | Error | ||||||
| Df | 2 | 24 | 48 | 216 | 1 | 24 | 24 | 144 | ||||||
| SS% | Sig | SS% | Sig | SS% | Sig | SS% | SS% | Sig | SS% | Sig | SS% | Sig | SS% | |
| Visual rating index (VRI) | ||||||||||||||
| 2011 | 36 | ** | 32 | ** | 18 | ** | 14 | 1 | ** | 72 | ** | 4 | ns | 22 |
| 2012 | 34 | ** | 35 | ** | 24 | ** | 7 | 10 | ** | 69 | ** | 12 | ** | 9 |
| 2013 | 39 | ** | 38 | ** | 18 | ** | 5 | 1 | ** | 88 | ** | 3 | ** | 8 |
| 2014 | 15 | ** | 66 | ** | 10 | ** | 9 | 1 | ** | 86 | ** | 6 | ** | 7 |
| Fusarium-damaged kernels (FDK) | ||||||||||||||
| 2011 | 56 | ** | 23 | ** | 14 | ** | 7 | 2 | ** | 70 | ** | 12 | ** | 15 |
| 2012 | 56 | ** | 20 | ** | 19 | ** | 5 | 13 | ** | 57 | ** | 20 | ** | 10 |
| 2013 | 47 | ** | 22 | ** | 19 | ** | 12 | 17 | ** | 51 | ** | 13 | ** | 19 |
| 2014 | 36 | ** | 26 | ** | 14 | ** | 24 | 1 | * | 57 | ** | 5 | ns | 37 |
* and ** F test significant (Sig) at p < 0.05 and p < 0.01, respectively; SS% percent of the total sum of squares; a only spray and maize stalk inoculation were included in the ANOVA.
Table 2.
Heritability estimates for the visual rating index (VRI) and the percentage of Fusarium-damaged kernels (FDK) in 25 wheat genotypes evaluated under three inoculation methods (spray, maize stalk and natural) in 2011, 2012, 2013, and 2014.
Table 2.
Heritability estimates for the visual rating index (VRI) and the percentage of Fusarium-damaged kernels (FDK) in 25 wheat genotypes evaluated under three inoculation methods (spray, maize stalk and natural) in 2011, 2012, 2013, and 2014.
| Year | Inoculation Method | Inoculation Method | ||||
|---|---|---|---|---|---|---|
| Spray | Maize Stalk | Natural | Spray | Maize Stalk | Natural | |
| Visual Rating Index (VRI) | Fusarium-Damaged Kernels (FDK) | |||||
| 2011 | 0.68 | 0.73 | 0.26 | 0.85 | 0.65 | 0.29 |
| 2012 | 0.91 | 0.77 | 0.46 | 0.85 | 0.83 | 0.44 |
| 2013 | 0.90 | 0.89 | 0.65 | 0.80 | 0.38 | 0.11 |
| 2014 | 0.83 | 0.95 | 0.63 | 0.56 | 0.43 | 0.12 |
Table 3.
Spearman’s rank correlation coefficients between inoculation methods for visual rating index and Fusarium-damaged kernels in 2011, 2012, 2013, and 2014.
Table 3.
Spearman’s rank correlation coefficients between inoculation methods for visual rating index and Fusarium-damaged kernels in 2011, 2012, 2013, and 2014.
| Visual Rating Index | Fusarium-Damaged Kernels | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Year | Inoculation Method | Spray | Maize Stalk | Spray | Maize Stalk | ||||
| 2011 | Maize stalk | 0.92 | ** | 0.84 | ** | ||||
| Natural | 0.60 | ** | 0.55 | ** | 0.54 | ** | 0.74 | ** | |
| 2012 | Maize stalk | 0.88 | ** | 0.60 | ** | ||||
| Natural | 0.72 | ** | 0.57 | ** | 0.53 | ** | 0.52 | ** | |
| 2013 | Maize stalk | 0.94 | ** | 0.77 | ** | ||||
| Natural | 0.88 | ** | 0.82 | ** | 0.19 | ns | 0.06 | ns | |
| 2014 | Maize stalk | 0.90 | ** | 0.87 | ** | ||||
| Natural | 0.83 | ** | 0.93 | ** | 0.62 | ** | 0.49 | * | |
* and ** correlation coefficient significant at p < 0.05 and p < 0.01, respectively.
Table 4.
Spearman’s rank correlation coefficients between visual rating index (VRI) and Fusarium-damaged kernels (FDK) under spray inoculation, maize stalk inoculation, and natural infection in 2011, 2012, 2013, and 2014.
Table 4.
Spearman’s rank correlation coefficients between visual rating index (VRI) and Fusarium-damaged kernels (FDK) under spray inoculation, maize stalk inoculation, and natural infection in 2011, 2012, 2013, and 2014.
| Inoculation Method | ||||||
|---|---|---|---|---|---|---|
| Year | Spray | Maize Stalk | Natural | |||
| VRI Versus FDK | ||||||
| 2011 | 0.83 | ** | 0.86 | ** | 0.43 | * |
| 2012 | 0.81 | ** | 0.72 | ** | 0.46 | * |
| 2013 | 0.89 | ** | 0.77 | ** | 0.16 | ns |
| 2014 | 0.77 | ** | 0.55 | ** | 0.82 | ** |
* and ** correlation coefficient significant at p < 0.05 and p < 0.01, respectively.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Šarčević, H.; Bukan, M.; Lovrić, A.; Maričević, M. Evaluation of Inoculation Methods for Determination of Winter Wheat Resistance to Fusarium Head Blight. Agronomy 2023, 13, 1175. https://doi.org/10.3390/agronomy13041175
AMA Style
Šarčević H, Bukan M, Lovrić A, Maričević M. Evaluation of Inoculation Methods for Determination of Winter Wheat Resistance to Fusarium Head Blight. Agronomy. 2023; 13(4):1175. https://doi.org/10.3390/agronomy13041175
Chicago/Turabian StyleŠarčević, Hrvoje, Miroslav Bukan, Ana Lovrić, and Marko Maričević. 2023. "Evaluation of Inoculation Methods for Determination of Winter Wheat Resistance to Fusarium Head Blight" Agronomy 13, no. 4: 1175. https://doi.org/10.3390/agronomy13041175
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.