
Nano-Hydroxyapatite and ZnO-NPs Mitigate Pb Stress in Maize


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
2.2. Measurements
2.2.1. Leaf Area
2.2.2. Total Chlorophyll (SPAD)
2.2.3. Gas Exchange Traits
2.2.4. Photosynthetic Pigments
2.2.5. Proline
2.2.6. Malonaldehyde (MDA)
2.2.7. Antioxidant Enzymes
2.2.8. Elemental Analysis
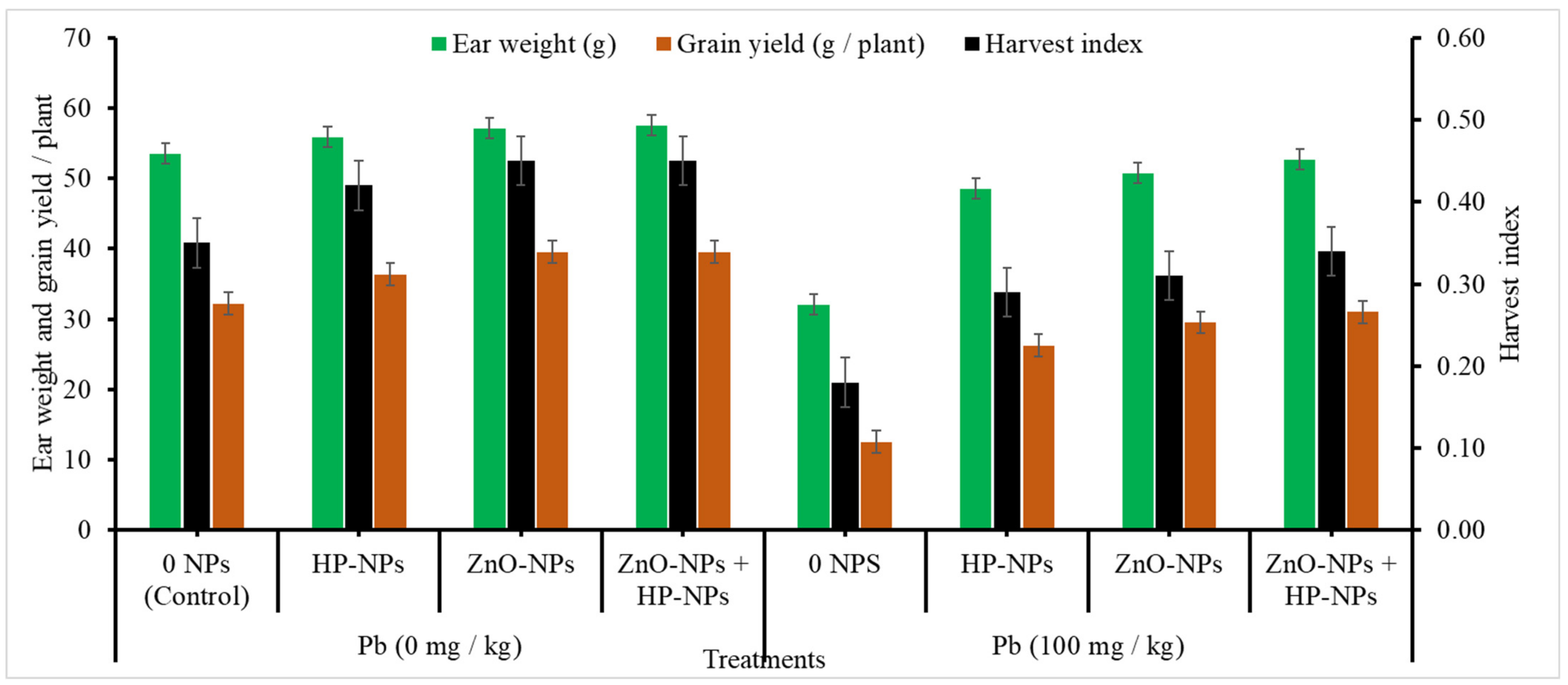
2.2.9. Yield Components
2.3. Statistical Analysis
3. Results
4. Discussion
- (a)
- Plant growth and physiological performance
- (b)
- Stress indicators and antioxidant enzymatic activity
- (c)
- Mineral ions uptake
- (d)
- Yield and yield attributes
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Waqas, M.A.; Kaya, C.; Riaz, A.; Farooq, M.; Nawaz, I.; Wilkes, A.; Li, Y. Potential mechanisms of abiotic stress tolerance in crop plants induced by thiourea. Front. Plant Sci. 2019, 10, 1336. [Google Scholar] [CrossRef] [PubMed]
- Taha, R.S.; Seleiman, M.F.; Shami, A.; Alhammad, B.A.; Mahdi, A.H.A. Integrated application of selenium and silicon enhances growth and anatomical structure, antioxidant defense system and yield of wheat grown in salt-stressed soil. Plants 2021, 10, 1040. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Ko, M.-S.; Kim, J.-Y.; Park, H.-S.; Kim, K.-W. Field assessment of arsenic immobilization in soil amended with iron rich acid mine drainage sludge. J. Clean. Prod. 2015, 108, 1073–1080. [Google Scholar] [CrossRef]
- Su, R.; Wang, Y.; Huang, S.; Chen, R.; Wang, J. Application for ecological restoration of contaminated soil: Phytoremediation. Int. J. Environ. Res. Public Health 2022, 19, 13124. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Alotaibi, M.A.; Alhammad, B.A.; Alharbi, B.M.; Refay, Y.; Badawy, S.A. Effects of ZnO nanoparticles and biochar of rice straw and cow manure on characteristics of contaminated soil and sunflower productivity, oil quality, and heavy metals uptake. Agronomy 2020, 10, 790. [Google Scholar] [CrossRef]
- Burakova, E.A.; Dyachkova, T.P.; Rukhov, A.V.; Tugolukov, E.N.; Galunin, E.V.; Tkachev, A.G.; Ali, I. Novel and economic method of carbon nanotubes synthesis on a nickel magnesium oxide catalyst using microwave radiation. J. Mol. Liq. 2018, 253, 340–346. [Google Scholar] [CrossRef]
- Abdel-Salam, A.; Salem, H.M.; Abdel-Salam, M.A.; Seleiman, M.F. Phyto and chemical removal of heavy metal-contaminated soils. In Heavy Metal Contamination of Soils: Monitoring and Remediation; Sherameti, I., Varma, A., Eds.; Series Soil Biology; Springer: Cham, Switzerland, 2015; Volume 44, pp. 299–308. [Google Scholar]
- Seleiman, M.F.; Santanen, A.; Mäkelä, P. Recycling sludge on cropland as fertilizer-Advantages and risks. Resour. Conserv. Recycl. 2020, 155, 104647. [Google Scholar] [CrossRef]
- Salem, H.S.; Abdel-Salam, A.; Abdel-Salam, M.A.; Seleiman, M.F. Phytoremediation of metal and metalloids from contaminated soil. In Plants under Metal and Metalloid Stress: Responses, Tolerance and Remediation; Hasanuzzaman, M., Nahar, K., Fujita, M., Eds.; Series Soil Biology; Springer Nature Pte Ltd.: Singapore, 2018; Chapter 9; pp. 249–262. [Google Scholar]
- Seneviratne, M.; Rajakaruna, N.; Rizwan, M.; Madawala, H.; Ok, Y.S.; Vithanage, M. Heavy metal-induced oxidative stress on seed germination and seedling development: A critical review. Environ. Geochem. Health 2019, 41, 1813–1831. [Google Scholar] [CrossRef]
- Tripathi, R.; Gupta, R.; Sahu, M.; Srivastava, D.; Das, A.; Ambasta, R.K.; Kumar, P. Free radical biology in neurological manifestations: Mechanisms to therapeutics interventions. Environ. Sci. Pollut. Res. 2021, 29, 62160–62207. [Google Scholar] [CrossRef]
- Metcalfe, N.B.; Olsson, M. How telomere dynamics are influenced by the balance between mitochondrial efficiency, reactive oxygen species production and DNA damage. Mol. Ecol. 2022, 31, 6040–6052. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.Y.; Roohi, M.; Iqbal, Z.; Ashraf, M.; Öztürk, M.; Gücel, S. Cadmium (Cd) and lead (Pb) induced changes in growth, some biochemical attributes, and mineral accumulation in two cultivars of mung bean [Vigna radiata (L.) Wilczek]. Commun. Soil Sci. Plant Anal. 2016, 47, 405–413. [Google Scholar]
- Shahid, M.J.; Ali, S.; Shabir, G.; Siddique, M.; Rizwan, M.; Seleiman, M.F.; Afzal, M. Comparing the performance of four macrophytes in bacterial assisted floating treatment wetlands for the removal of trace metals (Fe, Mn, Ni, Pb, and Cr) from polluted river water. Chemosphere 2020, 243, 125353. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Dubey, R.S. Lead toxicity in plants. Braz. J. Plant Physiol. 2005, 17, 35–52. [Google Scholar] [CrossRef]
- Hadi, F.; Aziz, T. A mini review on lead (Pb) toxicity in plants. J. Biol. Life Sci. 2015, 6, 91–101. [Google Scholar] [CrossRef]
- Ali, S.; Abbas, Z.; Seleiman, M.F.; Rizwan, M.; YavaŞ, İ.; Alhammad, B.A.; Shami, A.; Hasanuzzaman, M.; Kalderis, D. Glycine betaine accumulation, significance and interests for heavy metal tolerance in plants. Plants 2020, 9, 896. [Google Scholar] [CrossRef] [PubMed]
- Hossain, A.; Ahmad, Z.; Adeel, M.; Rahman, M.A.; Alam, M.J.; Ahmed, S.; Aftab, T. Emerging roles of osmoprotectants in heavy metal stress tolerance in plants. In Heavy Metal Toxicity in Plants; CRC Press: Boca Raton, FL, USA, 2021; pp. 95–110. ISBN 1003155081. [Google Scholar]
- Seleiman, M.F.; Selim, S.; Jaakkola, S.; Mäkelä, P.S.A. Chemical composition and in vitro digestibility of whole-crop maize fertilized with synthetic fertilizer or digestate and harvested at two maturity stages in boreal growing conditions. Agric. Food Sci. 2017, 26, 47–55. [Google Scholar] [CrossRef]
- Saleem, M.H.; Ali, S.; Seleiman, M.F.; Rizwan, M.; Rehman, M.; Aisha Akram, N.; Liu, L.; Alotaibi, M.; Al-Ashkar, I.; Mubushar, M. Assessing the correlations between different traits in copper-sensitive and copper-resistant varieties of jute (Corchorus capsularis L.). Plants 2019, 8, 545. [Google Scholar] [CrossRef]
- Riyazuddin, R.; Nisha, N.; Ejaz, B.; Khan, M.I.R.; Kumar, M.; Ramteke, P.W.; Gupta, R. A comprehensive review on the heavy metal toxicity and sequestration in plants. Biomolecules 2022, 12, 43. [Google Scholar] [CrossRef]
- Ganie, A.S.; Bano, S.; Khan, N.; Sultana, S.; Rehman, Z.; Rahman, M.M.; Sabir, S.; Coulon, F.; Khan, M.Z. Nanoremediation technologies for sustainable remediation of contaminated environments: Recent advances and challenges. Chemosphere 2021, 275, 130065. [Google Scholar] [CrossRef]
- Seewagen, C.L. The threat of global mercury pollution to bird migration: Potential mechanisms and current evidence. Ecotoxicology 2020, 29, 1254–1267. [Google Scholar] [CrossRef] [PubMed]
- Alazaiza, M.Y.D.; Albahnasawi, A.; Ali, G.A.M.; Bashir, M.J.K.; Copty, N.K.; Amr, S.S.A.; Abushammala, M.F.M.; Al Maskari, T. Recent advances of nanoremediation technologies for soil and groundwater remediation: A review. Water 2021, 13, 2186. [Google Scholar] [CrossRef]
- Ghani, M.I.; Saleem, S.; Rather, S.A.; Rehmani, M.S.; Alamri, S.; Rajput, V.D.; Kalaji, H.M.; Saleem, N.; Sial, T.A.; Liu, M. Foliar application of zinc oxide nanoparticles: An effective strategy to mitigate drought stress in cucumber seedling by modulating antioxidant defense system and osmolytes accumulation. Chemosphere 2022, 289, 133202. [Google Scholar] [CrossRef] [PubMed]
- Rajput, V.D.; Minkina, T.; Upadhyay, S.K.; Kumari, A.; Ranjan, A.; Mandzhieva, S.; Sushkova, S.; Singh, R.K.; Verma, K.K. Nanotechnology in the Restoration of Polluted Soil. Nanomaterials 2022, 12, 769. [Google Scholar] [CrossRef]
- Alhammad, B.A.; Ahmad, A.; Seleiman, M.F.; Tola, E. Seed priming with nanoparticles and 24-epibrassinolide improved seed germination and enzymatic performance of Zea mays L. in salt-stressed soil. Plants 2023, 12, 690. [Google Scholar] [CrossRef]
- Zhao, L.; Lu, L.; Wang, A.; Zhang, H.; Huang, M.; Wu, H.; Xing, B.; Wang, Z.; Ji, R. Nano-biotechnology in agriculture: Use of nanomaterials to promote plant growth and stress tolerance. J. Agric. Food Chem. 2020, 68, 1935–1947. [Google Scholar] [CrossRef]
- Zhao, L.; Zhu, Y.-H.; Wang, M.; Ma, L.-G.; Han, Y.-G.; Zhang, M.-J.; Li, X.-C.; Feng, W.-S.; Zheng, X.-K. Comparative transcriptome analysis of the hyperaccumulator plant Phytolacca americana in response to cadmium stress. 3 Biotech 2021, 11, 327. [Google Scholar] [CrossRef]
- Rhaman, M.S.; Tania, S.S.; Imran, S.; Rauf, F.; Kibria, M.G.; Ye, W.; Hasanuzzaman, M.; Murata, Y. Seed priming with nanoparticles: An emerging technique for improving plant growth, development, and abiotic stress tolerance. J. Soil Sci. Plant Nutr. 2022, 22, 4047–4062. [Google Scholar] [CrossRef]
- Ahmad, A.; Tola, E.; Alshahrani, T.S.; Seleiman, M.F. Enhancement of Morphological and Physiological Performance of Zea mays L. under Saline Stress Using ZnO Nanoparticles and 24-Epibrassinolide Seed Priming. Agronomy 2023, 13, 771. [Google Scholar] [CrossRef]
- Venkatachalam, P.; Jayaraj, M.; Manikandan, R.; Geetha, N.; Rene, E.R.; Sharma, N.C.; Sahi, S.V. Zinc oxide nanoparticles (ZnONPs) alleviate heavy metal-induced toxicity in Leucaena leucocephala seedlings: A physiochemical analysis. Plant Physiol. Biochem. 2017, 110, 59–69. [Google Scholar] [CrossRef]
- Majeed, J.; Ramkumar, J.; Chandramouleeswaran, S.; Tyagi, A.K. Role of surface functionalization of ZnO nanoparticles as sorbents for heavy metal ions. Sep. Sci. Technol. 2020, 55, 1922–1931. [Google Scholar] [CrossRef]
- Karn, B.; Kuiken, T.; Otto, M. Nanotechnology and in situ remediation: A review of the benefits and potential risks. Environ. Health Perspect. 2009, 117, 1813–1831. [Google Scholar] [CrossRef] [PubMed]
- Le, A.T.; Pung, S.-Y.; Sreekantan, S.; Matsuda, A. Mechanisms of removal of heavy metal ions by ZnO particles. Heliyon 2019, 5, e01440. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Al-Selwey, W.A.; Ibrahim, A.A.; Shady, M.; Alsadon, A.A. Foliar applications of zno and sio2 nanoparticles mitigate water deficit and enhance potato yield and quality traits. Agronomy 2023, 13, 466. [Google Scholar] [CrossRef]
- Khattak, A.; Ullah, F.; Shinwari, Z.K.; Mehmood, S. The effect of titanium dioxide nanoparticles and salicylic acid on growth and biodiesel production potential of sunflower (Helianthus annuus L.) under water stress. Pak. J. Bot. 2021, 53, 1987–1995. [Google Scholar] [CrossRef]
- Cui, H.; Yi, Q.; Yang, X.; Wang, X.; Wu, H.; Zhou, J. Effects of hydroxyapatite on leaching of cadmium and phosphorus and their availability under simulated acid rain. J. Environ. Chem. Eng. 2017, 5, 3773–3779. [Google Scholar] [CrossRef]
- Zou, T.; Pu, L.; Lin, R.; Mo, H.; Wang, Z.; Jian, S.; Zhang, M. Roles of Canavalia rosea metallothioneins in metal tolerance and extreme environmental adaptation to tropical coral reefs. J. Plant Physiol. 2022, 268, 153559. [Google Scholar] [CrossRef]
- Fiol, N.; Tarrés, Q.; Vásquez, M.G.; Pereira, M.A.; Mendonça, R.T.; Mutjé, P.; Delgado-Aguilar, M. Comparative assessment of cellulose nanofibers and calcium alginate beads for continuous Cu (II) adsorption in packed columns: The influence of water and surface hydrophobicity. Cellulose 2021, 28, 4327–4344. [Google Scholar] [CrossRef]
- Akhtar, N.; Khan, S.; Jamil, M.; Rehman, S.U.; Rehman, Z.U.; Rha, E.S. Combine effect of ZnO NPs and bacteria on protein and gene’s expression profile of rice (Oryza sativa L.) plant. Toxics 2022, 10, 305. [Google Scholar] [CrossRef]
- Agarwal, P.; Khurana, P. Characterization of a novel zinc finger transcription factor (TaZnF) from wheat conferring heat stress tolerance in Arabidopsis. Cell Stress Chaperones 2018, 23, 253–267. [Google Scholar] [CrossRef]
- Rossi, L.; Sharifan, H.; Zhang, W.; Schwab, A.P.; Ma, X. Mutual effects and in planta accumulation of co-existing cerium oxide nanoparticles and cadmium in hydroponically grown soybean (Glycine max (L.) Merr.). Environ. Sci. Nano 2018, 5, 150–157. [Google Scholar] [CrossRef]
- Shaheen, S.M.; Rinklebe, J. Impact of emerging and low cost alternative amendments on the (im) mobilization and phytoavailability of Cd and Pb in a contaminated floodplain soil. Ecol. Eng. 2015, 74, 319–326. [Google Scholar] [CrossRef]
- Shen, M.; Liu, W.; Zeb, A.; Lian, J.; Wu, J.; Lin, M. Bioaccumulation and Phytotoxicity of ZnO Nanoparticles in Soil-Grown Brassica chinensis L. and Potential risks. J. Environ. Manag. 2022, 306, 114454. [Google Scholar] [CrossRef] [PubMed]
- Youssef, M.S.; Elamawi, R.M. Evaluation of Phytotoxicity, Cytotoxicity, and Genotoxicity of ZnO Nanoparticles in Vicia faba. Environ. Sci. Pollut. Res. 2020, 27, 18972–18984. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Li, J.; Liu, W.; Zuo, Q.; Liang, S. Influence of nano-hydroxyapatite on the metal bioavailability, plant metal accumulation and root exudates of ryegrass for phytoremediation in lead-polluted soil. Int. J. Environ. Res. Public Health 2017, 14, 532. [Google Scholar] [CrossRef]
- Yan, Y.; Qi, F.; Zhao, S.; Luo, Y.; Gu, S.; Li, Q.; Zhang, L.; Zhou, S.; Bolan, N. A new low-cost hydroxyapatite for efficient immobilization of lead. J. Colloid Interface Sci. 2019, 553, 798–804. [Google Scholar] [CrossRef]
- Feng, Y.; Yang, J.; Liu, W.; Yan, Y.; Wang, Y. Hydroxyapatite as a passivator for safe wheat production and its impacts on soil microbial communities in a Cd-contaminated alkaline soil. J. Hazard. Mater. 2021, 404, 124005. [Google Scholar] [CrossRef]
- Liu, C.; Wang, L.; Yin, J.; Qi, L.; Feng, Y. Combined amendments of nano-hydroxyapatite immobilized cadmium in contaminated soil-potato (Solanum tuberosum L.) system. Bull. Environ. Contam. Toxicol. 2018, 100, 581–587. [Google Scholar] [CrossRef]
- Jin, Y.; Liu, W.; Li, X.; Shen, S.; Liang, S.; Liu, C.; Shan, L. Nano-hydroxyapatite immobilized lead and enhanced plant growth of ryegrass in a contaminated soil. Ecol. Eng. 2016, 95, 25–29. [Google Scholar] [CrossRef]
- Wang, P.; Zhang, S.; Wang, C.; Lu, J. Effects of Pb on the oxidative stress and antioxidant response in a Pb bioaccumulator plant Vallisneria natans. Ecotoxicol. Environ. Saf. 2012, 78, 28–34. [Google Scholar] [CrossRef]
- Li, Z.; Huang, J. Effects of nanoparticle hydroxyapatite on growth and antioxidant system in pakchoi (Brassica chinensis L.) from cadmium-contaminated soil. J. Nanomater. 2014, 2014, 470962. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Al-Suhaibani, N.; El-Hendawy, S.; Abdella, K.; Alotaibi, M.; Alderfasi, A. Impacts of long- and short-term of irrigation with treated wastewater and synthetic fertilizers on the growth, biomass, heavy metal content, and energy traits of three potential bioenergy crops in arid regions. Energies 2021, 14, 3037. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Santanen, A.; Jaakkola, S.; Ekholm, P.; Hartikainen, H.; Stoddard, F.L.; Mäkelä, P. Biomass yield and quality of bioenergy crops grown with synthetic and organic fertilizers. Biomass-Bioenergy 2013, 59, 477–485. [Google Scholar] [CrossRef]
- Dustgeer, Z.; Seleiman, M.F.; Khan, I.; Chattha, M.U.; Ali, E.F.; Alhammad, B.A.; Jalal, R.S.; Refay, Y.; Hassan, M.U. Glycine-betaine induced salinity tolerance in maize by regulating the physiological attributes, antioxidant defense system and ionic homeostasis. Not. Bot. Horti Agrobot. Cluj Napoca 2021, 49, 12248. [Google Scholar] [CrossRef]
- Erenstein, O.; Jaleta, M.; Sonder, K.; Mottaleb, K.; Prasanna, B.M. Global maize production, consumption and trade: Trends and R & D implications. Food Secur. 2022, 14, 1295–1319. [Google Scholar]
- Poole, N.; Donovan, J.; Erenstein, O. Agri-nutrition research: Revisiting the contribution of maize and wheat to human nutrition and health. Food Policy 2021, 100, 101976. [Google Scholar] [CrossRef]
- Sofy, M.R.; Seleiman, M.F.; Alhammad, B.A.; Alharbi, B.M.; Mohamed, H.I. Minimizing adverse effects of pb on maize plants by combined treatment with jasmonic, salicylic acids and proline. Agronomy 2020, 10, 699. [Google Scholar] [CrossRef]
- Abedi, T.; Gavanji, S.; Mojiri, A. Lead and zinc uptake and toxicity in maize and their management. Plants 2022, 11, 1922. [Google Scholar] [CrossRef]
- Chapman, H.D.; Pratt, P.F. Methods of analysis for soils, plants and waters. Soil Sci. 1962, 93, 68. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 1987; Volume 148, pp. 350–382. ISBN 0076-6879. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Cakmak, I.; Horst, W.J. Effect of aluminium on lipid peroxidation, superoxide dismutase, catalase, and peroxidase activities in root tips of soybean (Glycine max). Physiol. Plant. 1991, 83, 463–468. [Google Scholar] [CrossRef]
- García-Limones, C.; Hervás, A.; Navas-Cortés, J.A.; Jiménez-Díaz, R.M.; Tena, M. Induction of an antioxidant enzyme system and other oxidative stress markers associated with compatible and incompatible interactions between chickpea (Cicer arietinum L.) and Fusarium oxysporum f. sp. ciceris. Physiol. Mol. Plant Pathol. 2002, 61, 325–337. [Google Scholar] [CrossRef]
- Collin, S.; Baskar, A.; Geevarghese, D.M.; Ali, M.N.V.S.; Bahubali, P.; Choudhary, R.; Lvov, V.; Tovar, G.I.; Senatov, F.; Koppala, S. Bioaccumulation of Lead (Pb) and Its Effects in Plants: A Review. J. Hazard. Mater. Lett. 2022, 3, 100064. [Google Scholar] [CrossRef]
- Zhou, J.; Zhang, Z.; Zhang, Y.; Wei, Y.; Jiang, Z. Effects of Lead Stress on the Growth, Physiology, and Cellular Structure of Privet Seedlings. PLoS ONE 2018, 13, e0191139. [Google Scholar] [CrossRef] [PubMed]
- Akinci, I.E.; Akinci, S.; Yilmaz, K. Response of Tomato (Solanum lycopersicum L.) to Lead Toxicity: Growth, Element Uptake, Chlorophyll and Water Content. Afr. J. Agric. Res. 2010, 5, 416–423. [Google Scholar]
- Seregin, I.V.; Kozhevnikova, A.D. Distribution of Cadmium, Lead, Nickel, and Strontium in Imbibing Maize Caryopses. Russ. J. Plant Physiol. 2005, 52, 565–569. [Google Scholar] [CrossRef]
- Khan, A.; Khan, S.; Khan, M.A.; Qamar, Z.; Waqas, M. The Uptake and Bioaccumulation of Heavy Metals by Food Plants, Their Effects on Plants Nutrients, and Associated Health Risk: A Review. Environ. Sci. Pollut. Res. 2015, 22, 13772–13799. [Google Scholar] [CrossRef]
- Malar, S.; Shivendra Vikram, S.; JC Favas, P.; Perumal, V. Lead Heavy Metal Toxicity Induced Changes on Growth and Antioxidative Enzymes Level in Water Hyacinths [Eichhornia crassipes (Mart.)]. Bot. Stud. 2016, 55, 54. [Google Scholar] [CrossRef]
- Yang, Y.; Han, X.; Liang, Y.; Ghosh, A.; Chen, J.; Tang, M. The Combined Effects of Arbuscular Mycorrhizal Fungi (AMF) and Lead (Pb) Stress on Pb Accumulation, Plant Growth Parameters, Photosynthesis, and Antioxidant Enzymes in Robinia pseudoacacia L. PLoS ONE 2015, 10, 0145726. [Google Scholar] [CrossRef]
- Sidhu, G.P.S.; Singh, H.P.; Batish, D.R.; Kohli, R.K. Effect of lead on oxidative status, antioxidative response and metal accumulation in Coronopus didymus. Plant Physiol. Biochem. 2016, 105, 290–296. [Google Scholar] [CrossRef]
- Lamhamdi, M.; El Galiou, O.; Bakrim, A.; Nóvoa-Muñoz, J.C.; Arias-Estevez, M.; Aarab, A.; Lafont, R. Effect of Lead Stress on Mineral Content and Growth of Wheat (Triticum aestivum) and Spinach (Spinacia oleracea) Seedlings. Saudi J. Biol. Sci. 2013, 20, 29–36. [Google Scholar] [PubMed]
- Shu, X.; Yin, L.; Zhang, Q.; Wang, W. Effect of Pb Toxicity on Leaf Growth, Antioxidant Enzyme Activities, and Photosynthesis in Cuttings and Seedlings of Jatropha curcas L. Environ. Sci. Pollut. Res. 2012, 19, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Küpper, H.; Küpper, F.; Spiller, M. Environmental Relevance of Heavy Metal-Substituted Chlorophylls Using the Example of Water Plants. J. Exp. Bot. 1996, 47, 259–266. [Google Scholar] [CrossRef]
- Ashraf, U.; Kanu, A.S.; Deng, Q.; Mo, Z.; Pan, S.; Tian, H.; Tang, X. Lead (Pb) Toxicity; Physio-Biochemical Mechanisms, Grain Yield, Quality, and Pb Distribution Proportions in Scented Rice. Front. Plant Sci. 2017, 8, 259. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Almutairi, K.F.; Alotaibi, M.; Shami, A.; Alhammad, B.A.; Battaglia, M.L. Nano-fertilization as an emerging fertilization technique: Why can modern agriculture benefit from its use? Plants 2021, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Feleafel, M.N.; Mirdad, Z.M. Hazard and Effects of Pollution by Lead on Vegetable Crops. J. Agric. Environ. Ethics 2013, 26, 547–567. [Google Scholar] [CrossRef]
- Faiz, S.; Yasin, N.A.; Khan, W.U.; Shah, A.A.; Akram, W.; Ahmad, A.; Ali, A.; Naveed, N.H.; Riaz, L. Role of Magnesium Oxide Nanoparticles in the Mitigation of Lead-Induced Stress in Daucus carota: Modulation in Polyamines and Antioxidant Enzymes. Int. J. Phytoremediation 2022, 24, 364–372. [Google Scholar] [CrossRef]
- Tokarz, K.; Piwowarczyk, B.; Makowski, W. Mechanisms Involved in Photosynthetic Apparatus Protection against Lead Toxicity. In Lead in Plants and the Environment; Springer: Berlin/Heidelberg, Germany, 2020; pp. 117–128. [Google Scholar]
- Azmat, R.; Saleem, A.; Ahmed, W.; Qayyum, A.; El-Serehy, H.A.; Ali, S. The Investigation of the Impact of Toxicity of Metals on Oxygen-Evolving Complex in Spinacia oleracea. Antioxidants 2022, 11, 1802. [Google Scholar] [CrossRef]
- Ahmad, S.; Mfarrej, M.F.B.; El-Esawi, M.A.; Waseem, M.; Alatawi, A.; Nafees, M.; Saleem, M.H.; Rizwan, M.; Yasmeen, T.; Anayat, A. Chromium-Resistant Staphylococcus Aureus Alleviates Chromium Toxicity by Developing Synergistic Relationships with Zinc Oxide Nanoparticles in Wheat. Ecotoxicol. Environ. Saf. 2022, 230, 113142. [Google Scholar]
- Piotrowska, A.; Bajguz, A.; Godlewska-Żyłkiewicz, B.; Czerpak, R.; Kamińska, M. Jasmonic Acid as Modulator of Lead Toxicity in Aquatic Plant Wolffia arrhiza (Lemnaceae). Environ. Exp. Bot. 2009, 66, 507–513. [Google Scholar]
- Ashraf, M.; Foolad, M.R. Roles of Glycine Betaine and Proline in Improving Plant Abiotic Stress Resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of Proline under Changing Environments: A Review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [PubMed]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy Metal Stress and Some Mechanisms of Plant Defense Response. Sci. World J. 2015, 2015, 756120. [Google Scholar] [CrossRef] [PubMed]
- Pinto, E.; Sigaud-kutner, T.C.S.; Leitao, M.A.S.; Okamoto, O.K.; Morse, D.; Colepicolo, P. Heavy Metal–Induced Oxidative Stress in Algae 1. J. Phycol. 2003, 39, 1008–1018. [Google Scholar] [CrossRef]
- Lozano-Rodriguez, E.; Hernandez, L.E.; Bonay, P.; Carpena-Ruiz, R.O. Distribution of Cadmium in Shoot and Root Tissues 1. J. Exp. Bot. 1997, 48, 123–128. [Google Scholar] [CrossRef]
- Walker, W.M.; Miller, J.E.; Hassett, J.J. Effect of Lead and Cadmium upon the Calcium, Magnesium, Potassium, and Phosphorus Concentration in Young Corn Plants. Soil Sci. 1977, 124, 145–151. [Google Scholar] [CrossRef]
- Xiong, Z.; Zhao, F.; Li, M. Lead Toxicity in Brassica pekinensis Rupr.: Effect on Nitrate Assimilation and Growth. Environ. Toxicol. Int. J. 2006, 21, 147–153. [Google Scholar] [CrossRef]
- Song, Y.; Jin, L.; Wang, X. Cadmium Absorption and Transportation Pathways in Plants. Int. J. Phytoremediation 2017, 19, 133–141. [Google Scholar] [CrossRef]
- Gao, F.; Zhang, X.; Zhang, J.; Li, J.; Niu, T.; Tang, C.; Wang, C.; Xie, J. Zinc Oxide Nanoparticles Improve Lettuce (Lactuca sativa L.) Plant Tolerance to Cadmium by Stimulating Antioxidant Defense, Enhancing Lignin Content and Reducing the Metal Accumulation and Translocation. Front. Plant Sci. 2022, 13, 1015745. [Google Scholar] [CrossRef]
- Palusińska, M.; Barabasz, A.; Kozak, K.; Papierniak, A.; Maślińska, K.; Antosiewicz, D.M. Zn/Cd Status-Dependent Accumulation of Zn and Cd in Root Parts in Tobacco Is Accompanied by Specific Expression of ZIP Genes. BMC Plant Biol. 2020, 20, 37. [Google Scholar] [CrossRef]
- Emamverdian, A.; Hasanuzzaman, M.; Ding, Y.; Barker, J.; Mokhberdoran, F.; Liu, G. Zinc Oxide Nanoparticles Improve Pleioblastus pygmaeus Plant Tolerance to Arsenic and Mercury by Stimulating Antioxidant Defense and Reducing the Metal Accumulation and Translocation. Front. Plant Sci. 2022, 13, 841501. [Google Scholar] [CrossRef] [PubMed]
- Kidwai, M.; Dhar, Y.V.; Gautam, N.; Tiwari, M.; Ahmad, I.Z.; Asif, M.H.; Chakrabarty, D. Oryza sativa Class III Peroxidase (OsPRX38) Overexpression in Arabidopsis thaliana Reduces Arsenic Accumulation Due to Apoplastic Lignification. J. Hazard. Mater. 2019, 362, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.Y.; Traina, S.J.; Logan, T.J.; Ryan, J.A. In Situ Lead Immobilization by Apatite. Environ. Sci. Technol. 1993, 27, 1803–1810. [Google Scholar] [CrossRef]
- Zhang, P.; Ryan, J.A. Transformation of Pb (II) from Cerrusite to Chloropyromorphite in the Presence of Hydroxyapatite under Varying Conditions of PH. Environ. Sci. Technol. 1999, 33, 625–630. [Google Scholar] [CrossRef]
- Zhang, P.; Ryan, J.A. Formation of Chloropyromorphite from Galena (PbS) in the Presence of Hydroxyapatite. Environ. Sci. Technol. 1999, 33, 618–624. [Google Scholar] [CrossRef]
- Takeuchi, Y.; Arai, H. Removal of Coexisting Pb2+, Cu2+ and Cd2+ Ions from Water by Addition of Hydroxyapatite Powder. J. Chem. Eng. Jpn. 1990, 23, 75–80. [Google Scholar] [CrossRef]
- Shashkova, I.L.; Rat’ko, A.I.; Kitikova, N.V. Removal of Heavy Metal Ions from Aqueous Solutions by Alkaline-Earth Metal Phosphates. Colloids Surf. A Physicochem. Eng. Asp. 1999, 160, 207–215. [Google Scholar] [CrossRef]
- Hussain, B.; Lin, Q.; Hamid, Y.; Sanaullah, M.; Di, L.; Khan, M.B.; He, Z.; Yang, X. Foliage Application of Selenium and Silicon Nanoparticles Alleviates Cd and Pb Toxicity in Rice (Oryza sativa L.). Sci. Total Environ. 2020, 712, 136497. [Google Scholar] [CrossRef]


{kind=link}
| Traits | Leaf Area (cm2 Plant−1) | |||
|---|---|---|---|---|
| Treatments | 40 DAS | 70 DAS | 100 DAS | |
| Pb (0 mg kg−1) | 0 NPs (Control) | 2751 b | 8259 b | 7752 a |
| HP-NPs (50 mg kg−1) | 2894 a | 8344 b | 7756 a | |
| ZnO-NPs (50 mg L−1) | 2938 a | 8561 a | 7866 a | |
| ZnO-NPs + HP-NPs | 2953 a | 8592 a | 7884 a | |
| Pb (100 mg kg−1) | 0 NPs | 1350 e | 3677 f | 3554 d |
| HP-NPs (50 mg kg−1) | 1927 d | 6722 e | 6311 c | |
| ZnO-NPs (50 mg kg−1) | 2153 c | 7125 d | 6982 b | |
| ZnO-NPs + HP-NPs | 2745 b | 7850 c | 7733 a | |
| LSD 0.05 | 75 | 105 | 145 | |
| Traits | Total Chlorophyll (SPAD) | Pn (μmol CO2 m−2 s−1) | gs (mol H2O m−2 s−1) | |
|---|---|---|---|---|
| Treatments | ||||
| Pb (0 mg kg−1) | 0 NPs (Control) | 49.1 a | 15.56 c | 0.22 b |
| HP-NPs (50 mg kg−1) | 49.9 a | 18.54 b | 0.23 b | |
| ZnO-NPs (50 mg L−1) | 50.4 a | 20.54 a | 0.29 a | |
| ZnO-NPs + HP-NPs | 50.5 a | 20.61 a | 0.29 a | |
| Pb (100 mg kg−1) | 0 NPs | 32.3 e | 4.65 f | 0.05 d |
| HP-NPs (50 mg kg−1) | 40.9 d | 11.56 e | 0.18 bc | |
| ZnO-NPs (50 mg kg−1) | 43.7 c | 13.56 d | 0.21 b | |
| ZnO-NPs + HP-NPs | 47.5 ab | 16.51 c | 0.24 b | |
| LSD 0.05 | 2.1 | 1.15 | 0.04 |
| Traits | Proline (mg g−1 DW) | MDA (μM g−1 FW) | Carotenoids (mg g−1 FW) | |
|---|---|---|---|---|
| Treatments | ||||
| Pb (0 mg kg−1) | 0 NPs (Control) | 109.21 e | 8.07 f | 2.85 b |
| HP-NPs (50 mg kg−1) | 103.15 f | 8.91 ef | 2.82 b | |
| ZnO-NPs (50 mg L−1) | 99.04 f | 8.51 f | 3.02 a | |
| ZnO-NPs + HP-NPs | 97.58 f | 8.56 f | 3.08 a | |
| Pb (100 mg kg−1) | 0 NPs | 235.87 a | 19.62 a | 1.56 e |
| HP-NPs (50 mg kg−1) | 199.25 b | 12.58 b | 2.15 d | |
| ZnO-NPs (50 mg kg−1) | 162.58 c | 11.85 c | 2.39 c | |
| ZnO-NPs + HP-NPs | 121.98 d | 9.12 d | 2.79 b | |
| LSD 0.05 | 4.52 | 0.46 | 0.07 |
| Traits | SOD (unit g−1 FW) | POX (unit g−1 FW) | CAT (unit g−1 FW) | |
|---|---|---|---|---|
| Treatments | ||||
| Pb (0 mg kg−1) | 0 NPs (Control) | 33.56 e | 31.02 de | 2.65 c |
| HP-NPs (50 mg kg−1) | 34.89 de | 31.61 de | 2.81 c | |
| ZnO-NPs (50 mg L−1) | 33.97 e | 30.36 e | 2.75 c | |
| ZnO-NPs + HP-NPs | 34.90 de | 31.52 de | 2.80 c | |
| Pb (100 mg kg−1) | 0 NPs | 62.79 a | 59.19 a | 6.52 a |
| HP-NPs (50 mg kg−1) | 43.89 b | 45.34 b | 3.62 b | |
| ZnO-NPs (50 mg kg−1) | 39.46 c | 41.25 c | 3.41 b | |
| ZnO-NPs + HP-NPs | 36.21 de | 33.27 d | 2.92 c | |
| LSD 0.05 | 1.98 | 2.51 | 0.47 |
| Traits | N | P | K | Zn | Pb | Pb | |
|---|---|---|---|---|---|---|---|
| Treatments | In Biomass (Above Ground) | In Grains | |||||
| (g kg−1 DM) | (mg kg−1 DM) | ||||||
| Pb (0 mg kg−1) | 0 NPs (Control) | 16.39 a | 1.53 a | 7.84 a | 40.23 c | 0.00 d | 0.00 d |
| HP-NPs (50 mg kg−1) | 16.52 a | 1.75 a | 7.55 a | 42.09 c | 0.02 d | 0.00 d | |
| ZnO-NPs (50 mg L−1) | 16.71 a | 1.49 ab | 7.56 a | 51.21 a | 0.05 d | 0.00 d | |
| ZnO-NPs + HP-NPs | 16.74 a | 1.77 a | 7.56 a | 53.53 a | 0.04 d | 0.00 d | |
| Pb (100 mg kg−1) | 0 NPs | 10.05 d | 0.46 d | 4.17 c | 7.21 e | 13.47 a | 5.11 a |
| HP-NPs (50 mg kg−1) | 12.53 c | 1.21 c | 6.72 ab | 35.40 d | 7.25 b | 1.74 b | |
| ZnO-NPs (50 mg kg−1) | 12.99 c | 1.07 c | 6.99 a | 45.96 b | 6.26 b | 1.57 b | |
| ZnO-NPs + HP-NPs | 15.96 ab | 1.46 ab | 7.23 a | 47.02 b | 3.02 c | 0.05 c | |
| LSD 0.05 | 0.62 | 0.23 | 0.95 | 2.05 | 1.21 | 0.24 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alhammad, B.A.; Ahmad, A.; Seleiman, M.F. Nano-Hydroxyapatite and ZnO-NPs Mitigate Pb Stress in Maize. Agronomy 2023, 13, 1174. https://doi.org/10.3390/agronomy13041174
Alhammad BA, Ahmad A, Seleiman MF. Nano-Hydroxyapatite and ZnO-NPs Mitigate Pb Stress in Maize. Agronomy. 2023; 13(4):1174. https://doi.org/10.3390/agronomy13041174
Chicago/Turabian StyleAlhammad, Bushra Ahmed, Awais Ahmad, and Mahmoud F. Seleiman. 2023. "Nano-Hydroxyapatite and ZnO-NPs Mitigate Pb Stress in Maize" Agronomy 13, no. 4: 1174. https://doi.org/10.3390/agronomy13041174