

Diversity in Coffea arabica Cultivars in the Mountains of Gorongosa National Park, Mozambique, Regarding Bean and Leaf Nutrient Accumulation and Physical Fruit Traits
 ,
,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Brief Characterization of Plant Material, Experimental Design, and Plant Area
2.2. Fruit and Leaf Collection
2.3. Mineral Concentration in Fruit and Leaf Tissues
2.4. Statistical Analyses
3. Results
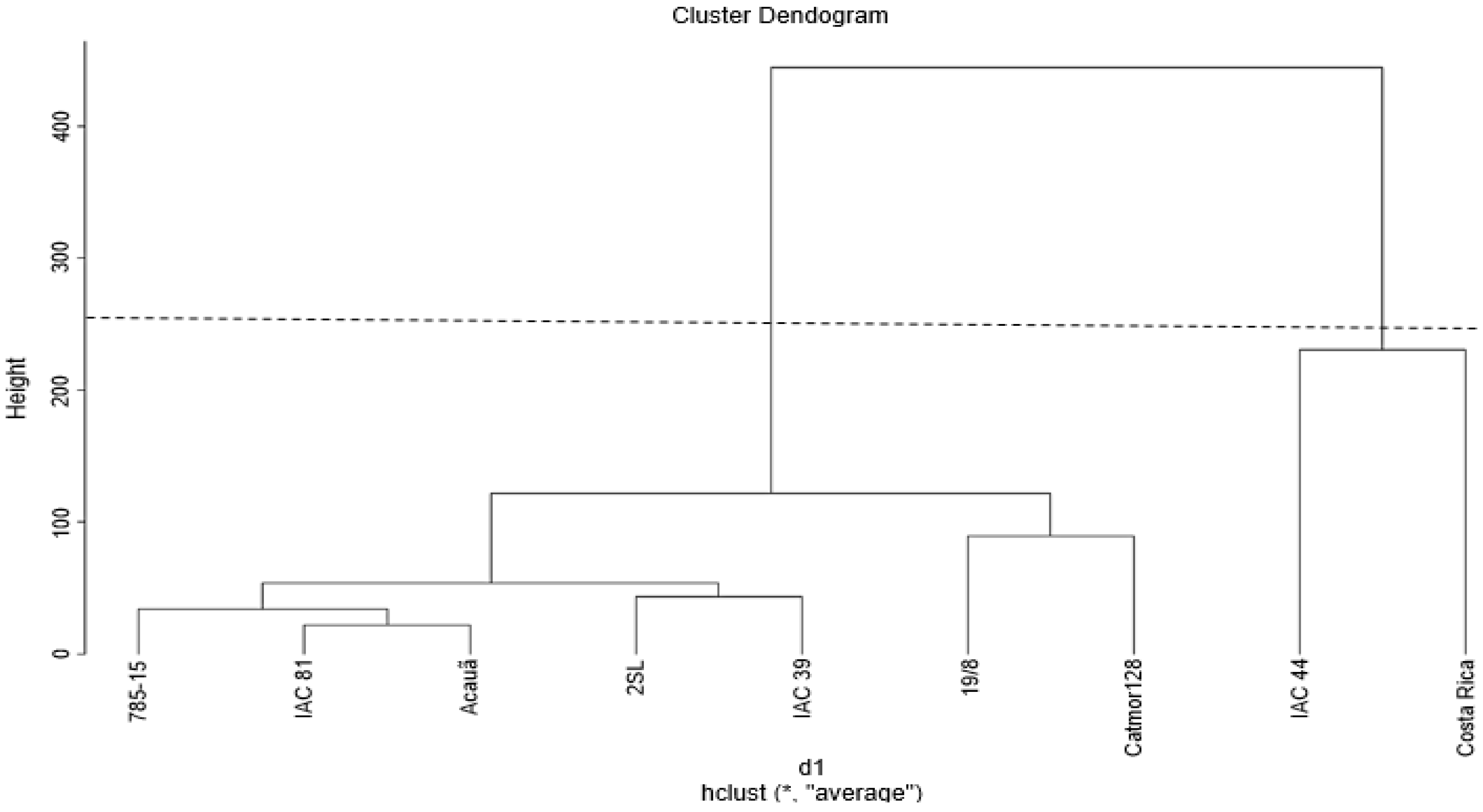
3.1. Dissimilarity between Genotypes and Genetic Contribution
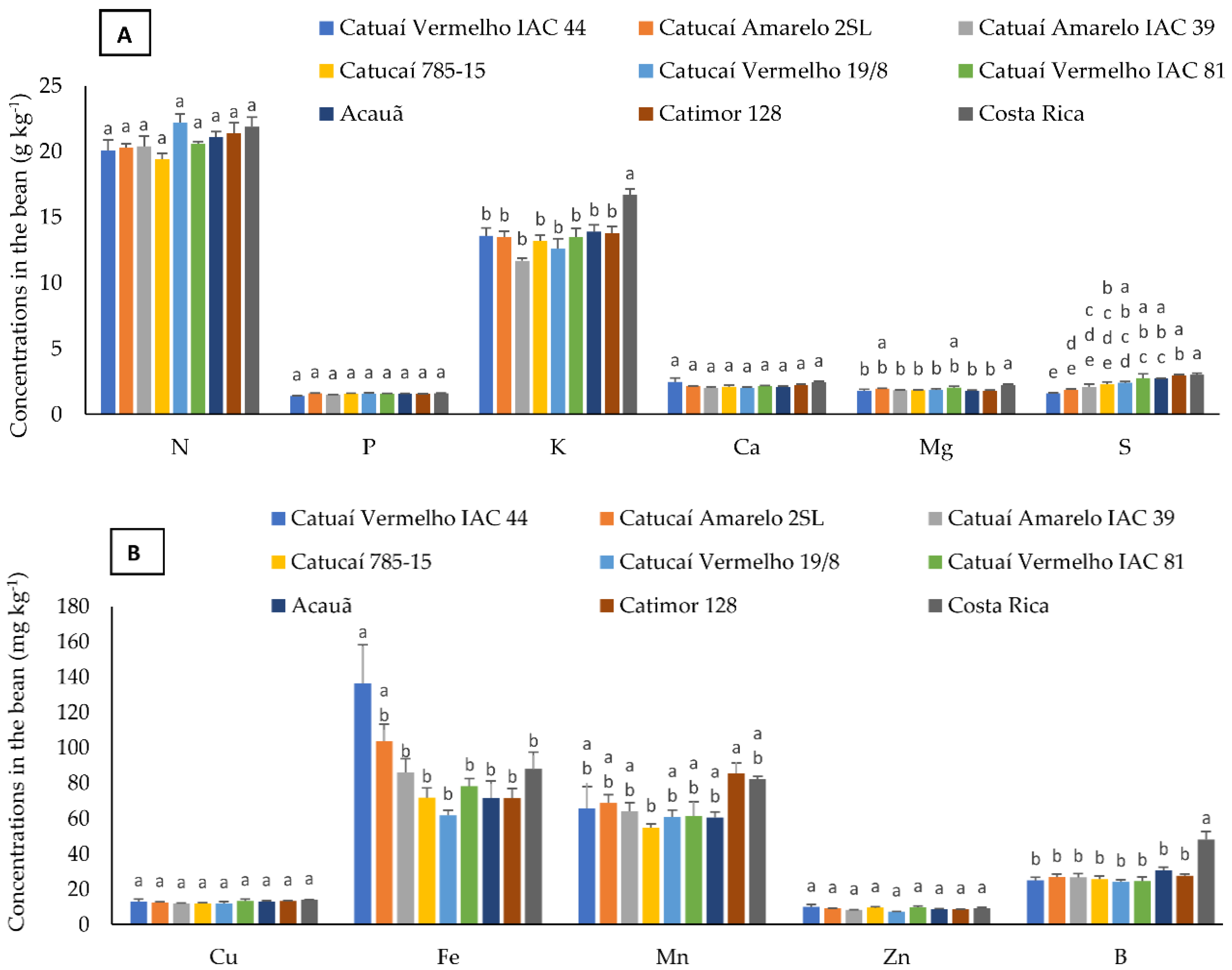
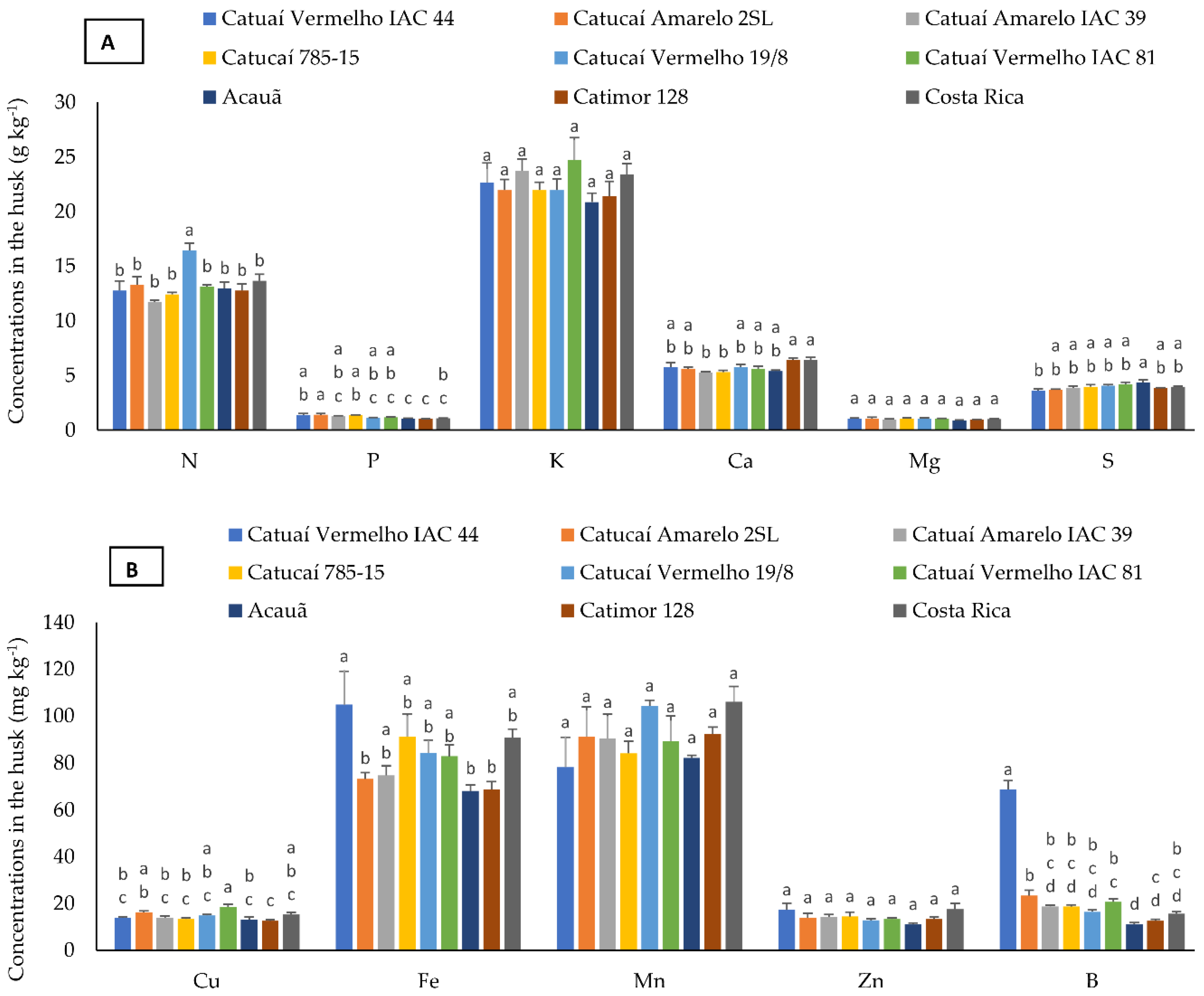
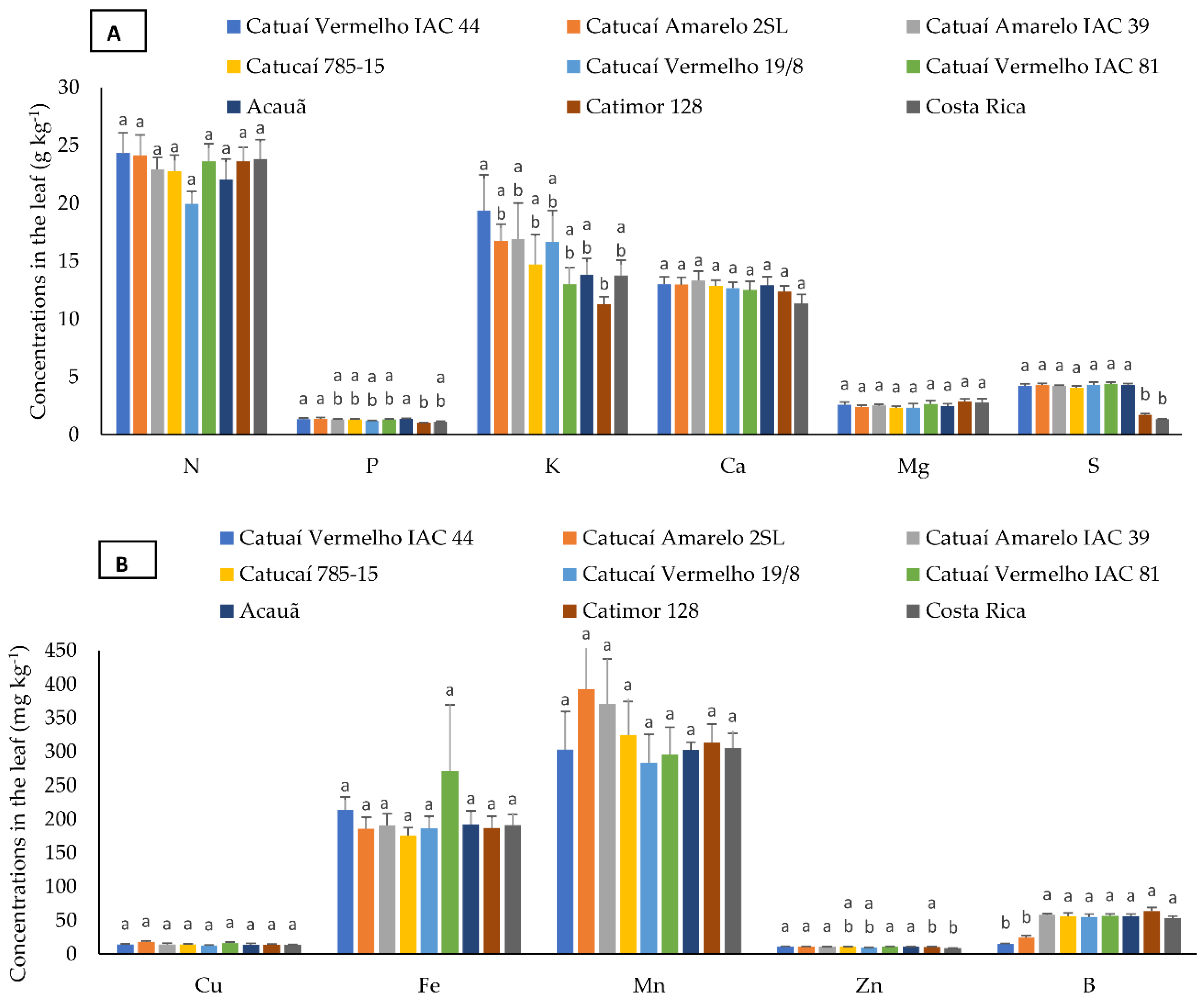
3.2. Nutrient Concentration in Bean, Husk, and Leaf
3.3. Nutrient Accumulation in the Fruit
3.4. Correlations between Cumulative Nutrient Concentrations in Bean, Husk, Fruit, and Husk Percentage, Bean Percentage, Bean Weight, and Fruit Weight
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- International Coffee Organization. Available online: https://www.ico.org/new_historical.asp?section=Statistics (accessed on 15 July 2022).
- Davis, A.P.; Rakotonasolo, F. Six new species of coffee (Coffea) from northern Madagascar. Kew Bull. 2021, 76, 497–511. [Google Scholar] [CrossRef]
- Loureiro, J. Flora Cochinchinensis; Ulyssipone: Typis, et Expensis Academicis: Lisbon, Portugal, 1790. [Google Scholar]
- Bridson, D.M.; Polhill, R.M.; Verdcourt, B. “Coffea” in Flora of Tropical East Africa, Rubiaceae; East African Governments: Rotterdam, The Netherlands, 1988; pp. 703–723. [Google Scholar]
- Bridson, D.M. “Coffea” in Flora Zambesiaca; Pope, G.V., Ed.; Royal Botanic Gardens, Kew: Richmond, UK, 2003; pp. 452–463. [Google Scholar]
- Davis, A.P.; Gargiulo, R.; Almeida, I.N.d.M.; Caravela, M.I.; Denison, C.; Moat, J. Hot Coffee: The Identity, Climate Profiles, Agronomy, and Beverage Characteristics of Coffea racemosa and C. zanguebariae. Front. Sustain. Food Syst. 2021, 5, 740137. [Google Scholar] [CrossRef]
- Cassamo, C.T.; Mangueze, A.V.J.; Leitão, A.E.; Pais, I.P.; Moreira, R.; Campa, C.; Chiulele, R.; Reis, F.O.; Marques, I.; Scotti-Campos, P.; et al. Shade and Altitude Implications on the Physical and Chemical Attributes of Green Coffee Beans from Gorongosa Mountain, Mozambique. Agronomy 2022, 12, 2540. [Google Scholar] [CrossRef]
- Flávio, F.S.; Júlio, C.F.S.; José, N.M.C.; Milton, M.S. Características das Principais Variedades de Café Cultivadas em Rondônia In Embrapa Centro de Pesquisa Agroflorestal de Rondônia Ministério da Agricultura, Pecuária e Abastecimento; Embrapa: Porto Velho, Brazil, 2004; ISSN 0103-9865. [Google Scholar]
- Nantawan, U.; Kanchana-Udomkan, C.; Drew, R.; Ford, R. Identification of genes related to sugar content inCarica papaya L.: Differential expression and candidate marker development. Acta Hortic. 2018, 203, 129–136. [Google Scholar] [CrossRef]
- Sakiyama, N.S.; Pereira, A.A.; Zambolim, L. Melhoramento de café arábica. In Melhoramento de Espécies Cultivadas; Borém, A., Ed.; UFV: Viçosa, Brazil, 1999; pp. 189–204. [Google Scholar]
- Sant’ana, G.C.; Pereira, L.F.P.; Pot, D.; Ivamoto, S.T.; Domingues, D.S.; Ferreira, R.V.; Pagiatto, N.F.; da Silva, B.S.R.; Nogueira, L.M.; Kitzberger, C.S.G.; et al. Genome-wide association study reveals candidate genes influencing lipids and diterpenes contents in Coffea arabica L. Sci. Rep. 2018, 8, 465. [Google Scholar] [CrossRef]
- Hu, F.; Bi, X.; Liu, H.; Fu, X.; Li, Y.; Yang, Y.; Zhang, X.; Wu, R.; Li, G.; Lv, Y.; et al. Transcriptome and carotenoid profiling of different varieties of Coffea arabica provides insights into fruit color formation. Plant Divers. 2021, 44, 322–334. [Google Scholar] [CrossRef]
- Ramalho, J.C.; Pais, I.P.; Leitão, A.E.; Guerra, M.; Reboredo, F.H.; Máguas, C.M.; Carvalho, M.L.; Scotti-Campos, P.; Ribeiro-Barros, A.I.; Lidon, F.J.C.; et al. Can Elevated Air [CO2] Conditions Mitigate the Predicted Warming Impact on the Quality of Coffee Bean? Front. Plant Sci. 2018, 9, 287. [Google Scholar] [CrossRef]
- Van Der Vossen, H.; Bertrand, B.; Charrier, A. Next generation variety development for sustainable production of arabica coffee (Coffea arabica L.): A review. Euphytica 2015, 204, 243–256. [Google Scholar] [CrossRef]
- Santana, L.S.; Ferraz, G.A.e.S.; Teodoro, A.J.d.S.; Santana, M.S.; Rossi, G.; Palchetti, E. Advances in Precision Coffee Growing Research: A Bibliometric Review. Agronomy 2021, 11, 1557. [Google Scholar] [CrossRef]
- Rodrigues, W.P.; Martins, M.Q.; Fortunato, A.S.; Rodrigues, A.P.; Semedo, J.N.; Simões-Costa, M.C.; Pais, I.P.; Leitão, A.E.; Colwell, F.; Goulao, L.; et al. Long-term elevated air [CO2] strengthens photosynthetic functioning and mitigates the impact of supra-optimal temperatures in tropical Coffea arabica and C. canephora species. Glob. Chang. Biol. 2016, 22, 415–431. [Google Scholar] [CrossRef]
- Machado, C.M.S.; Pimentel, N.S.; Golynsk, A.; Ferreira, A.; Vieira, H.D.; Partelli, F.L. Genetic diversity among 16 genotypes of Coffea arabica in the Brazilian cerrado. Genet. Mol. Res. 2017, 16, gmr16039794. [Google Scholar] [CrossRef] [PubMed]
- DaMatta, F.M.; Avila, R.; Cardoso, A.A.; Martins, S.C.V.; Ramalho, J.C. Physiological and Agronomic Performance of the Coffee Crop in the Context of Climate Change and Global Warming: A Review. J. Agric. Food Chem. 2018, 66, 5264–5274. [Google Scholar] [CrossRef] [PubMed]
- DaMatta, F.M.; Rahn, E.; Läderach, P.; Ghini, R.; Ramalho, J.C. Why could the coffee crop endure climate change and global warming to a greater extent than previously estimated? Clim. Chang. 2019, 152, 167–178. [Google Scholar] [CrossRef]
- Martins, L.D.; Tomaz, M.A.; Lidon, F.C.; DaMatta, F.M.; Ramalho, J.C. Combined effects of elevated [CO2] and high temperature on leaf mineral balance in Coffea spp. plants. Clim. Chang. 2014, 126, 365–379. [Google Scholar] [CrossRef]
- Partelli, F.L.; Gomes, W.R.; Oliveira, M.G.; Dias, J.R.M.; Espindula, M.C. Normas foliares e diagnóstico nutricional do cafeeiro na pré-florada e granação. Espirito Santo. Coffee Sci. 2016, 11, 544–554. [Google Scholar] [CrossRef]
- Partelli, F.L.; Oliveira, M.G.; Covre, A.M.; Vieira, H.D.; Dias, J.R.M.; Braun, H. Nutritional standards and nutritional diagnosis of the Conilon coffee plants in phenological growth stages. J. Plant Nutr. 2018, 41, 2536–2546. [Google Scholar] [CrossRef]
- Oliosi, G.; Partelli, F.L.; Da Silva, C.A.; Dubberstein, D.; Gontijo, I.; Tomaz, M.A. Seasonal variation in leaf nutrient concentration of conilon coffee genotypes. J. Plant Nutr. 2020, 44, 74–85. [Google Scholar] [CrossRef]
- Campos, R.C.; Pinto, V.R.A.; Melo, L.F.; da Rocha, S.J.S.S.; Coimbra, J.S. New sustainable perspectives for “coffee wastewater” and other by-products: A critical review. Futur. Foods 2021, 4, 100058. [Google Scholar] [CrossRef]
- Bragança, S.M.; Martinez, H.E.P.; Leite, H.G.; Santos, L.P.; Sediyama, C.S.; Alvarez, V.V.H.; Lani, J.A. B, Cu, Fe, Mn & Zn accumulation by Conilon coffee plant. Rev. Ceres 2007, 54, 398–404. [Google Scholar]
- Partelli, F.L.; Espindula, M.C.; Marré, W.B.; Vieira, H.D. Dry matter and macronutrient accumulation in fruits of Conilon coffee with different ripening cycles. Rev. Bras. Ciência Solo 2014, 38, 214–222. [Google Scholar] [CrossRef]
- Covre, A.M.; Rodrigues, W.P.; Vieira, H.D.; Braun, H.; Ramalho, J.; Partelli, F.L. Nutrient accumulation in bean and fruit from irrigated and non-irrigated Coffea canephora cv. Conilon. Emir. J. Food Agric. 2016, 28, 402–409. [Google Scholar] [CrossRef]
- Covre, A.M.; Partelli, F.L.; Bonomo, R.; Gontijo, I. Micronutrients in the fruits and leaves of irrigated and non-irrigated coffee plants. J. Plant Nutr. 2018, 41, 1119–1129. [Google Scholar] [CrossRef]
- Larcher, W. Ecologia Vegetal; RiMa Artes e Textos: São Carlos, Brazil, 2000; 531p. [Google Scholar]
- Martins, L.; Rodrigues, W.; Machado, S.; Brinate, S.; Colodetti, T.; Amaral, J.; Tomaz, M. Evidence of genetic tolerance to low availability of phosphorus in the soil among genotypes of Coffea canephora. Genet. Mol. Res. 2015, 14, 10576–10587. [Google Scholar] [CrossRef]
- Gomes, W.R.; Rodrigues, W.P.; Vieira, H.D.; Oliveira, M.G.; Dias, J.R.M.; Partelli, F.L. Genetic diversity of standard leaf nutrients in Coffea canephora genotypes during phenological phases. Genet. Mol. Res. 2016, 15, gmr.15048839. [Google Scholar] [CrossRef]
- Marraccini, P.; Vinecky, F.; Alves, G.S.C.; Ramos, H.J.O.; Elbelt, S.; Vieira, N.G.; Carneiro, F.A.; Sujii, P.S.; Alekcevetch, J.C.; Silva, V.A.; et al. Differentially expressed genes and proteins upon drought acclimation in tolerant and sensitive genotypes of Coffea canephora. Ind. J. Gen. Plant Breed. 2012, 63, 4191–4212. [Google Scholar] [CrossRef] [PubMed]
- Santos, H.G.; Jacomine, P.K.T.; Anjos, L.H.C.; Oliveira, V.A.; Lumbreras, J.F.; Coelho, M.R.; Almeida, J.A.; Araujo Filho, J.C.; Oliveira, J.B.; Cunha, T.J.F. Sistema Brasileiro de Classificação de Solos, 5th ed.; Embrapa: Brasília, Brazil, 2018. [Google Scholar]
- Silva, F.C. Manual de Análises Químicas de Solos, Plantas e Fertilizantes, 2nd ed.; Embrapa Informação Tecnológica: Brasília, Brazil, 2009; pp. 191–233. [Google Scholar]
- Peters, J.B. Wisconsin Procedures for Soil Testing, Plant Analysis and Feed & Forage Analysis: Plant Analysis; Department of Soil Science, College of Agriculture and Life Sciences, University of Wisconsin-Extension: Madison, WI, USA, 2005; Available online: https://uwlab.soils.wisc.edu/files/procedures/plant_icp.pdf (accessed on 20 February 2014).
- Martins, M.Q.; Partelli, F.L.; Ferreira, A.; Bernardes, C.D.O.; Golynski, A.; Vieira, H.D.; Freitas, M.S.M.; Ramalho, J.C. Genetic variability on nutrient contents in Coffea canephora genotypes cultivated at 850 meters of altitude in two crop seasons. Funct. Plant Breed. J. 2019, 1, 1–12. [Google Scholar] [CrossRef]
- dos Santos, M.M.; da Silva, C.A.; Oza, E.F.; Gontijo, I.; Amaral, J.F.T.D.; Partelli, F.L. Concentration of Nutrients in Leaves, Flowers, and Fruits of Genotypes of Coffea canephora. Plants 2021, 10, 2661. [Google Scholar] [CrossRef]
- Brazil Ministério da Agricultura, Pecuária e Abastecimento. Regras Para Análise de Sementes. 2009. Available online: https://www.gov.br/agricultura/pt-br/assuntos/insumos-agropecuarios/arquivos-publicacoes-insumos/2946regrasanalisesementes.pdf/view (accessed on 17 April 2023).
- Cruz, C.D. Programa Genes—Ampliado e integrado aos aplicativos R, Matlab e Selegen. Acta Sci. Agron. 2016, 38, 547–552. [Google Scholar] [CrossRef]
- Gu, Z.; Gu, L.; Eils, R.; Schlesner, M.; Brors, B. circlize implements and enhances circular visualization in R. Bioinformatics 2014, 30, 2811–2812. [Google Scholar] [CrossRef]
- Singh, D. The relative importance of characters affecting genetic divergence. Ind. J. Gen. Plant Breed. 1981, 41, 237–245. [Google Scholar]
- Schmidt, R.; da Silva, C.A.; Dubberstein, D.; Dias, J.R.M.; Vieira, H.D.; Partelli, F.L. Genetic Diversity Based on Nutrient Concentrations in Different Organs of Robusta Coffee. Agronomy 2022, 12, 640. [Google Scholar] [CrossRef]
- Yan, L.; Ogutu, C.; Huang, L.; Wang, X.; Zhou, H.; Lv, Y.; Long, Y.; Dong, Y.; Han, Y. Genetic Diversity and Population Structure of Coffee Germplasm Collections in China Revealed by ISSR Markers. Plant Mol. Biol. Rep. 2019, 37, 204–213. [Google Scholar] [CrossRef]
- Castro, R.D.; Marraccini, P. Cytology, biochemistry and molecular changes during coffee fruit development. Braz. J. Plant Physiol. 2006, 18, 175–199. [Google Scholar] [CrossRef]
- Partelli, F.L.; Oliosi, G.; Dalazen, J.R.; da Silva, C.A.; Vieira, H.D.; Espindula, M.C. Proportion of ripe fruit weight and volume to green coffee: Differences in 43 genotypes of Coffea canephora. Agron. J. 2021, 113, 1050–1057. [Google Scholar] [CrossRef]
- Prezotti, L.C.; Bragança, S.M. Accumulation of dry mass, N, P and K in different genetic sources of Conilon coffee. Coffee Sci. 2013, 8, 284–294. [Google Scholar]
- Gomes, P.F. Curso de Estatística Experimental, 12th ed.; Livraria Nobel: Piracicaba, Brazil, 1985; 467p. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Properties | Results | ||
|---|---|---|---|
| M.O. | Organic matter (Oxy-Red.) | dag/dm3 | 5.5 |
| Ph | (water—ratio 1:2.5) | unit | 5.1 |
| P | (Mehlich−1) | mg/dm3 | 8.4 |
| K | (Mehlich−1) | mg/dm3 | 80 |
| Ca | (Kcl−1 mol/L) | cmolc/dm3 | 1.2 |
| Mg | (Kcl−1 mol/L) | cmolc/dm3 | 0.4 |
| Al | (Kcl−1 mol/L) | cmolc/dm3 | 0.6 |
| H + Al | (Calcium acetate) | cmolc/dm3 | 11 |
| S.B. | (Sum of bases) | cmolc/dm3 | 1.81 |
| C.T.C. | (Cation exchange capacity at pH = 7) | cmolc/dm3 | 12.81 |
| V% | (Base saturation) | % | 14 |
| %K C.T.C | (%K at CTC) | % | 2 |
| %Ca C.T.C | (%Ca at CTC) | % | 9 |
| %Mg C.T.C. | (%Mg at CTC) | % | 3 |
| %Al C.T.C. | (%Al at CTC) | % | 4.7 |
| %H+Al C.T.C. | (%H + Al at C.T.C.) | % | 86 |
| P (Resin) | mg/dm3 | 13.6 | |
| P-rem | (Remaining phosphorus) | mg/L | 13 |
| Na | (Mehlich−1) | mg/dm3 | 3 |
| S | (Monocalcium acetic phosphate) | mg/dm3 | 22 |
| B | (Hot water) | mg/dm3 | 0.2 |
| Zn | (Mehlich−1) | mg/dm3 | 1.4 |
| Mn | (Mehlich−1) | mg/dm3 | 30.6 |
| Cu | (Mehlich−1) | mg/dm3 | 4.9 |
| Fe | (Mehlich−1) | mg/dm3 | 28 |
| Granulometric fractions (g kg−1) | |||
| Sand | |||
| Silt | |||
| Clay | |||
| Soil type | Clayish | ||
| Identification | Name | MAPA Registration Number | Maturation Season | Productivity |
|---|---|---|---|---|
| 1 | Catuaí Vermelho IAC 44 | 2929 | Medium and late | High |
| 2 | Catucaí Amarelo 2SL | 4915 | Medium | High |
| 3 | Catuaí Amarelo IAC 39 | 2937 | Late | High |
| 4 | Catucaí 785-15 | 4996 | Precocious | High |
| 5 | Catucaí Vermelho 19/8 | 4909 | Precocious | High/Medium |
| 6 | Catuaí Vermelho IAC 81 | 2932 | Medium | High |
| 7 | Acauã | 4995 | Late | High |
| 8 | Catimor 128 | |||
| 9 | Called Costa Rica |
| Variable | S.j | Value (%) | Cumulative Value (%) |
|---|---|---|---|
| B | 5559.12 | 44.45 | 44.45 |
| Fe | 2573.03 | 20.58 | 65.03 |
| S | 1310.84 | 10.48 | 75.51 |
| Bean weight | 839.46 | 6.71 | 82.22 |
| K | 614.21 | 4.91 | 87.13 |
| Mn | 482.87 | 3.86 | 91.00 |
| Mg | 278.87 | 2.23 | 93.23 |
| %Bean | 269.01 | 2.15 | 95.38 |
| ripe Kg | 214.37 | 1.71 | 97.09 |
| P | 145.69 | 1.17 | 98.26 |
| N | 101.21 | 0.81 | 99.07 |
| Ca | 70.37 | 0.56 | 99.63 |
| Cu | 23.63 | 0.19 | 99.82 |
| Zn | 20.87 | 0.17 | 99.98 |
| Fruit weight | 2.02 | 0.02 | 100.00 |
| Groups | Cultivars |
|---|---|
| I | Catuaí Vermelho IAC 81, Acauã, Catuaí Amarelo IAC 39, Catucaí 785-15, Catucaí Amarelo 2SL, Catucaí Vermelho 19/8, Catimor 128, Costa Rica |
| II | Catuaí Vermelho IAC 44 |
| Cultivars | B | H | DBW | RFW | DFW | RFW/ DFW | RFW/ DBW | Ripe Fruit Weight/ton of Bean | Ripe Fruit Weight/60 kg of Bean | Dry Fruit Weight/ton of Bean |
|---|---|---|---|---|---|---|---|---|---|---|
| % | % | g | g | G | kg−1 | kg−1 | ton−1 | |||
| Catuaí Vermelho IAC 44 | 62.2 a | 37.8 a | 0.17 ab | 1.60 ab | 0.50 ab | 3.21 b | 5.16 a | 5166 a | 310 a | 1608 a |
| Catucaí Amarelo 2SL | 59.9 a | 40.1 a | 0.13 c | 1.34 c | 0.41 c | 3.24 ab | 5.41 a | 5415 a | 325 a | 1671 a |
| Catuaí Amarelo IAC 39 | 61.5 a | 38.5 a | 0.15 abc | 1.50 bc | 0.47 ab | 3.13 b | 5.09 a | 5096 a | 306 a | 1628 a |
| Catucaí 785-15 | 61.5 a | 38.5 a | 0.15 bc | 1.46 bc | 0.44 bc | 3.24 ab | 5.28 a | 5282 a | 317 a | 1629 a |
| Catucaí Vermelho 19/8 | 60.9 a | 39.1 a | 0.15 bc | 1.54 abc | 0.47 abc | 3.27 ab | 5.39 a | 5398 a | 324 a | 1648 a |
| Catuaí Vermelho IAC 81 | 61.1 a | 38.9 a | 0.16 ab | 1.57 abc | 0.49 ab | 3.15 b | 5.18 a | 5182 a | 311 a | 1640 a |
| Acauã | 62.9 a | 37.1 a | 0.17 ab | 1.64 ab | 0.51 ab | 3.21 b | 5.11 a | 5111 a | 307 a | 1589 a |
| Catimor 128 | 62.5 a | 37.5 a | 0.18 a | 1.77 a | 0.52 a | 3.35 ab | 5.36 a | 5360 a | 322 a | 1601 a |
| Costa Rica | 61.7 a | 38.3 a | 0.15 bc | 1.61 ab | 0.46 abc | 3.44 a | 5.58 a | 5586 a | 335 a | 1623 a |
| CV (%) | 3.74 | 5.99 | 6.21 | 6.17 | 5.41 | 2.83 | 4.73 | 4.73 | 4.73 | 3.76 |
| Cultivars | Macronutrients | Micronutrients | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N | P | K | Ca | Mg | S | Cu | Fe | Mn | Zn | B | |
| (g kg−1) | (mg kg−1) | ||||||||||
| Fruit | |||||||||||
| Catuaí Vermelho IAC 44 | 27.3 b | 2.15 ab | 26.3 ab | 5.74 a | 2.40 ab | 3.62 b | 20.8 a | 195.5 a | 110.3 a | 19.9 a | 63.4 a |
| Catucaí Amarelo 2SL | 28.5 ab | 2.46 a | 27.1 ab | 5.63 a | 2.59 ab | 4.17 ab | 22.7 a | 149.2 ab | 125.1 a | 17.7 a | 41.3 b |
| Catuaí Amarelo IAC 39 | 27.2 b | 2.22 ab | 25.5 b | 5.06 a | 2.38 ab | 4.32 ab | 20.1 a | 129.2 b | 116.3 a | 16.7 a | 37.6 b |
| Catucaí 785-15 | 26.7 b | 2.35 ab | 26.0 ab | 5.19 a | 2.43 ab | 4.60 ab | 19.8 a | 124.8 b | 103.9 a | 17.8 a | 36.5 b |
| Catucaí Vermelho 19/8 | 32.1 a | 2.22 ab | 25.8 b | 5.48 a | 2.47 ab | 4.84 ab | 20.9 a | 112.8 b | 123.0 a | 15.0 a | 34.0 b |
| Catuaí Vermelho IAC 81 | 28.6 ab | 2.24 ab | 27.9 ab | 5.41 a | 2.63 ab | 5.24 a | 24.1 a | 127.8 b | 115.3 a | 17.7 a | 36.7 b |
| Acauã | 28.1 ab | 2.12 b | 25.3 b | 5.03 a | 2.26 b | 5.11 a | 20.4 a | 108.8 b | 105.3 a | 14.6 a | 36.8 b |
| Catmor 128 | 28.5 ab | 2.14 ab | 25.6 b | 5.81 a | 2.31 ab | 5.08 a | 20.2 a | 109.8 b | 137.1 a | 16.2 a | 34.7 b |
| Costa Rica | 29.7 ab | 2.21 ab | 30.1 a | 6.10 a | 2.82 a | 5.28 a | 22.8 a | 140.2 ab | 143.1 a | 19.2 a | 57.3 a |
| CV (%) | 6.41 | 6.09 | 6.52 | 9.4 | 8.65 | 10.98 | 8.96 | 18.4 | 18.0 | 14.2 | 10.5 |
| Variables | Nutrients | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N | P | K | Ca | Mg | S | Cu | Fe | Mn | Zn | B | |
| Bean × Husk | 0.64 * | −0.23 | −0.30 | 0.71 * | 0.38 | 0.54 | 0.15 | 0.38 | 0.50 | 0.6 * | −0.55 |
| Bean × NAB | 1 *** | 1 *** | 1 *** | 1 *** | 1 *** | 1 *** | 0.99 *** | 1 *** | 1 *** | 1 *** | 1 *** |
| Bean × NAH | 0.28 | −0.18 | −0.49 | 0.55 | 0.60 * | 0.27 | 0.00 | 0.42 | 0.22 | 0.48 | −0.56 |
| Bean × NAF | 0.82 ** | 0.18 | 0.23 | 0.80 ** | 0.96 *** | 0.95 *** | 0.57 | 0.93 *** | 0.83 ** | 0.83 ** | 0.35 |
| Bean × %Bean | 0.09 | −0.18 | 0.66 * | 0.30 | −0.69 * | 0.31 | 0.40 | −0.12 | 0.12 | 0.08 | 0.55 |
| Bean × %Husk | −0.09 | 0.18 | −0.66 * | −0.30 | 0.69 * | −0.31 | −0.40 | 0.12 | −0.12 | −0.08 | −0.54 |
| Bean × Fruit weight | 0.29 | −0.35 | 0.37 | 0.11 | −0.65 * | 0.29 | 0.33 | −0.36 | 0.12 | −0.06 | 0.05 |
| Bean × Bean weight | 0.14 | −0.31 | 0.47 | 0.29 | −0.66 * | 0.30 | 0.44 | −0.28 | 0.11 | 0.18 | 0.09 |
| Husk × %Husk | 0.39 | 0.60 * | 0.39 | −0.24 | 0.64 * | −0.06 | 0.75 ** | 0.18 | 0.34 | 0.04 | 0.54 |
| Husk × NAH | 0.85 ** | 0.99 *** | 0.83 ** | 0.80 ** | 0.86 ** | 0.85 ** | 0.98 *** | 0.98 *** | 0.87 ** | 0.92 *** | 1 *** |
| Husk × NAF | 0.91 *** | 0.75 ** | 0.56 | 0.90 *** | 0.55 | 0.71 * | 0.83 ** | 0.55 | 0.88 ** | 0.93 *** | 0.47 |
| Husk × %Bean | −0.39 | −0.6 * | −0.39 | 0.24 | −0.64 * | 0.06 | −0.75 ** | −0.18 | −0.34 | −0.04 | −0.54 |
| Husk × Bean weight | −0.29 | −0.60 * | −0.29 | 0.39 | −0.44 | 0.12 | −0.59 * | −0.20 | −0.39 | −0.31 | −0.33 |
| Husk × Fruit weight | −0.22 | −0.66 * | −0.27 | 0.34 | −0.52 | 0.16 | −0.54 | −0.35 | −0.29 | −0.47 | −0.37 |
| NAF × NAH | 0.73 * | 0.80 ** | 0.52 | 0.9 ** | 0.68 * | 0.47 | 0.77 ** | 0.58 | 0.68 * | 0.86 ** | 0.44 |
| NAF × %Bean | −0.3 | −0.90 *** | −0.43 | 0.03 | −0.65 * | 0.24 | −0.38 | −0.35 | −0.17 | −0.09 | 0.14 |
| NAF × %Husk | 0.3 | 0.80 ** | 0.43 | −0.03 | 0.65 * | −0.24 | 0.38 | 0.35 | 0.17 | 0.09 | −0.14 |
| NAF × Fruit weight | 0.01 | −0.83 ** | −0.49 | −0.03 | −0.64 * | 0.22 | −0.13 | −0.48 | −0.11 | −0.40 | −0.19 |
| NAF × Bean weight | −0.097 | −0.80 ** | −0.36 | 0.06 | −0.60 * | 0.22 | −0.18 | −0.42 | −0.19 | −0.19 | −0.14 |
| NAH × %Bean | −0.66 * | −0.63 * | −0.75 ** | −0.23 | −0.90 *** | −0.44 | −0.84 ** | −0.24 | −0.69 * | −0.26 | −0.55 |
| NAH × %Husk | 0.66 * | 0.63 * | 0.75 ** | 0.23 | 0.90 *** | 0.44 | 0.84 ** | 0.24 | 0.69 * | 0.26 | 0.55 |
| NAH × Fruit weight | −0.49 | −0.70 * | −0.54 | −0.18 | −0.70 * | −0.19 | −0.54 | −0.41 | −0.50 | −0.58 | −0.39 |
| NAH × Bean weight | −0.49 | −0.62 * | −0.62 * | −0.15 | −0.68 * | −0.25 | −0.62 * | −0.25 | −0.63 * | −0.49 | −0.33 |
| %Bean × %Husk | −1 * | ||||||||||
| %Bean × Fruit weight | 0.73 * | ||||||||||
| %Bean × Bean weight | 0.80 ** | ||||||||||
| %Husk × Fruit weight | −0.73 * | ||||||||||
| %Husk × Bean weight | −0.80 ** | ||||||||||
| Fruit weight × Bean weight | 0.96 *** | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alberto, N.J.; Ramalho, J.C.; Ribeiro-Barros, A.I.; Viana, A.P.; Krohling, C.A.; Moiane, S.S.; Alberto, Z.; Rodrigues, W.P.; Partelli, F.L. Diversity in Coffea arabica Cultivars in the Mountains of Gorongosa National Park, Mozambique, Regarding Bean and Leaf Nutrient Accumulation and Physical Fruit Traits. Agronomy 2023, 13, 1162. https://doi.org/10.3390/agronomy13041162
Alberto NJ, Ramalho JC, Ribeiro-Barros AI, Viana AP, Krohling CA, Moiane SS, Alberto Z, Rodrigues WP, Partelli FL. Diversity in Coffea arabica Cultivars in the Mountains of Gorongosa National Park, Mozambique, Regarding Bean and Leaf Nutrient Accumulation and Physical Fruit Traits. Agronomy. 2023; 13(4):1162. https://doi.org/10.3390/agronomy13041162
Chicago/Turabian StyleAlberto, Niquisse J., José C. Ramalho, Ana I. Ribeiro-Barros, Alexandre P. Viana, Cesar A. Krohling, Sional S. Moiane, Zito Alberto, Weverton P. Rodrigues, and Fábio L. Partelli. 2023. "Diversity in Coffea arabica Cultivars in the Mountains of Gorongosa National Park, Mozambique, Regarding Bean and Leaf Nutrient Accumulation and Physical Fruit Traits" Agronomy 13, no. 4: 1162. https://doi.org/10.3390/agronomy13041162