
The Impact of Rotational Pasture Management for Farm-Bred Fallow Deer (Dama dama) on Fodder Quality in the Context of Animal Welfare
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
2.3. Mineral Concentration Analysis in Plants
2.4. Animal Weight Measurement
2.5. Statistical Analysis
3. Results
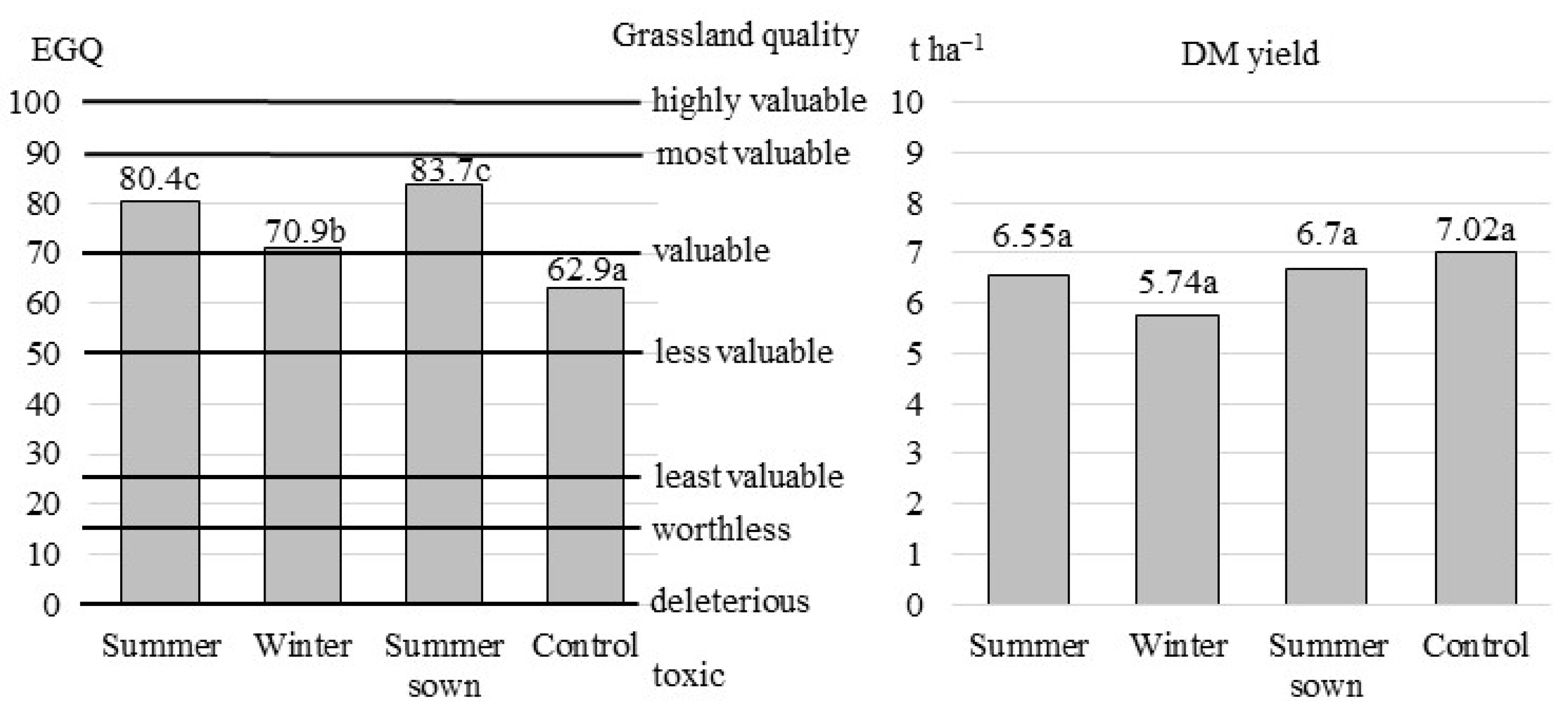
3.1. Species Composition vs. Grassland Quality
3.2. The Nutritional Value of Green Fodder
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wardle, D.A.; Bardgett, R.D.; Klironomos, J.N.; Setälä, H.; Van der Putten, W.H.; Wall, D.H. Ecological linkages between aboveground and belowground biota. Science 2004, 304, 1629–1633. [Google Scholar] [CrossRef]
- Pastor, J.; Naiman, R.J. Selective foraging and ecosystem processes in boreal forests. Am. Nat. 1992, 139, 690–705. [Google Scholar] [CrossRef]
- Pastor, J.; Dewey, B.; Naiman, R.J.; McInnes, P.F.; Cohen, Y. Moose browsing and soil fertility in the boreal forests of Isle Royale National Park. Ecology 1993, 74, 467–480. [Google Scholar] [CrossRef]
- Bruno, E. Microhistological analysis of rumen contents to evaluate the diet of adult male fallow deer (Dama dama) in a submediterranean costal area. In Ungulates/Ongules; S.F.E.P.M.: Toulouse, France, 1991; Volume 91, pp. 2–6, 15. [Google Scholar]
- Burke, K.M. Seasonal Diets and Foraging Selectivity of White-Tailed Deer in the Rolling Plains Ecological Region. M.S. Thesis, Southwest Texas State University, San Marcos, TX, USA, 2003. [Google Scholar]
- Estévez, J.A.; Landete-Castillejos, T.; García, A.J.; Ceacero, F.; Martinez, A.; Gaspar-López, E.; Calatayud, A.; Gallego, L. Seasonal variations in plant mineral content and free-choice minerals consumed by deer. Anim. Prod. Sci. 2010, 50, 177–185. [Google Scholar] [CrossRef]
- Kielland, K.; Bryant, J.P.; Ruess, R.W. Moose herbivory and carbon turnover of early successional stands in interior Alaska. Oikos 1997, 80, 25–30. [Google Scholar] [CrossRef]
- Futa, B.; Patkowski, K.; Bielińska, E.J.; Gruszecki, T.M.; Pluta, M.; Kulik, M.; Chmielewski, S. Sheep and horse grazing in a large-scale protection area and its positive impact on chemical and biological soil properties. Pol. J. Soil Sci. 2016, 49, 111–122. [Google Scholar] [CrossRef]
- Bielińska, E.J.; Gruszecki, T.M. Influence of the extensive sheep pasturage on enzymatic activity of soils in selected natural habitats “Natura 2000”. Zesz. Probl. Postęp. Nauk Rol. 2011, 567, 11–19. [Google Scholar]
- Patkowski, K.; Pluta, M.; Lipiec, A.; Greguła-Kania, M.; Gruszecki, T.M. Foraging behavior patterns of sheep and horses under a mixed species grazing system. J. Appl. Anim. Welf. Sci. 2019, 22, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Chabuz, W.; Kulik, M.; Sawicka-Zugaj, W.; Żółkiewski, P.; Warda, M.; Pluta, M.; Lipiec, A.; Bochniak, A.; Zdulski, J. Impact of the type of use of permanent grasslands areas in mountainos regions on the floristic diversity of habitats and animal welfare. Glob. Ecol. Conserv. 2019, 19, e00629. [Google Scholar] [CrossRef]
- Janiszewski, P.; Bogdaszewski, M.; Murawska, D.; Tajchman, K. Welfare of farmed deer-practical aspects. Pol. J. Nat. Sci. 2016, 31, 345–361. [Google Scholar]
- Nazaruk, M.; Jankowska-Huflejt, H.; Wróbel, B. Evaluation of the nutritional value of fodder from permanent grassland in the ecological farms under study. Woda-Sr.-Obsz. Wiej. 2009, 9, 61–76. [Google Scholar]
- Janiszewski, P.; Bogdaszewska, Z.; Bogdaszewski, M.; Bogdaszewski, P.; Cilulko-Dołęga, J.; Nasiadka, P.; Steiner, Ż. Breeding and Farm Breeding of Deer; Publishing House of the University of Warmia and Mazury in Olsztyn: Olsztyn, Poland, 2014. (In Polish) [Google Scholar]
- Richardson, C.; Lionberger, J.; Miller, G. White-Tailed Deer Management in the Rolling Plains of Texas; Wildlife Biologists Texas Parks and Wildlife Department: Austin, TX, USA, 2008.
- Litherland, A.J.; Woodward, S.J.R.; Stevens, D.R.; McDougal, D.B.; Boom, C.J.; Knight, T.L.; Lambert, M.G. Seasonal Variations in Pasture Quality on New Zealand Sheep and Beef Farms. Proc. N. Z. Soc. Anim. Prod. 2002, 62, 138–142. [Google Scholar]
- ARC. The Nutrient Requirements of Ruminant Livestock; Agricultural Research Council: Farnham Royal, UK, 1980.
- NRC. Nutrient Requirements of Small Ruminants. Sheep, Goats, Cervids, and New World Camelids; National Academy Press: Washington, DC, USA, 2007. [Google Scholar]
- Asher, G.W.; Stevens, D.R.; Archer, J.A.; Barrell, G.K.; Scott, I.C.; Ward, J.F.; Littlejohn, R.P. Energy and protein as nutritional drivers of lactation and calf growth of farmed red deer. Livest. Sci. 2011, 140, 8–16. [Google Scholar] [CrossRef]
- Molvar, E.M.; Bowyer, R.T.; Ballenberghe Van, V. Moose herbivory, browse quality, and nutrient cycling in an Alaskan treeline community. Oecologia 1993, 94, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.A.; Groffman, P.M. Ungulate vs. landscape control of soil C and N processes in grasslands of Yellowstone National Park. Ecology 1998, 79, 2229–2241. [Google Scholar] [CrossRef]
- Franzluebbers, A.J.; Stuedemann, J.A.; Schomberg, H.H.; Wilkinson, S.R. Soil organic C and N pools under long-term pasture management in the Southern Piedmont USA. Soil Biol. Biochem. 2000, 32, 469–478. [Google Scholar] [CrossRef]
- Sirotnak, J.M.; Huntly, N.J. Direct and indirect effects of herbivores on nitrogen dynamics: Voles in riparian areas. Ecology 2000, 81, 78–87. [Google Scholar] [CrossRef]
- Wardle, D.A.; Barker, G.M.; Yeates, G.W.; Bonner, K.I.; Ghani, A. Introduced browsing mammals in New Zealand natural forests: Aboveground and belowground consequences. Ecol. Monogr. 2001, 71, 587–614. [Google Scholar] [CrossRef]
- Stark, S.; Mannisto, M.K.; Smolander, A. Multiple effects of reindeer grazing on the soil processes in nutrient-poor nothern boreal forests. Soil Biol. Biochem. 2010, 42, 2068–2077. [Google Scholar] [CrossRef]
- Kulik, M.; Bochniak, A.; Chabuz, W.; Żółkiewski, P.; Rysiak, A. Is Grazing Good for Wet Meadows? Vegetation Changes Caused by White-Backed Cattle. Agriculture 2023, 13, 261. [Google Scholar] [CrossRef]
- Gómez, J.A.; Ceacero, F.; Landete-Castillejos, T.; Gaspar-López, E.; García, A.J.; Gallego, L. Factors affecting antler investment in Iberian red deer. Anim. Prod. Sci. 2012, 52, 867–873. [Google Scholar] [CrossRef]
- Gómez, J.A.; Landete-Castillejos, T.; García, A.J.; Gaspar-López, E.; Estévez, J.A.; Gallego, L. Lactation growth influences mineral composition of first antler in Iberian red deer (Cervus elaphus hispanicus). Wildl. Biol. 2008, 14, 331–338. [Google Scholar] [CrossRef]
- Landete-Castillejos, T.; Estevez, J.A.; Ceacero, F.; García, A.J.; Gallego, L. A review of factors affecting antler composition and mechanics. Front. Biosci. 2012, E4, 2328–2339. [Google Scholar] [CrossRef]
- Olguin, C.A.; Landete-Castillejos, T.; Ceacero, F.; García, A.J.; Gallego, L. Effects of Feed Supplementation on Mineral Composition, Mechanical Properties and Structure in Femurs of Iberian Red Deer Hinds (Cervus elaphus hispanicus). PLoS ONE 2013, 8, e65461. [Google Scholar] [CrossRef]
- Ceacero, F. Lon or heavy? Physiological constraints in the evolution of antlers. J. Mammal. Evol. 2015, 23, 2209–2216. [Google Scholar] [CrossRef]
- Dryden, G.M. Nutrition of antler growth in deer. Anim. Prod. Sci. 2016, 56, 962–970. [Google Scholar] [CrossRef]
- Tajchman, K.; Bogdaszewski, M.; Kowalczuk-Vasilev, E. Effects of supplementation with different levels of calcium and phosphorus on mineral content of first antler, bone, muscle, and liver of farmed fallow deer (Dama dama). Can. J. Anim. Sci. 2020, 100, 17–26. [Google Scholar] [CrossRef]
- Steiner-Bogdaszewska, Ż.; Tajchman, K.; Ukalska-Jaruga, A.; Florek, M.; Pecio, M. The Mineral Composition of Bone Marrow, Plasma, Bones and the First Antlers of Farmed Fallow Deer. Animals 2022, 12, 2764. [Google Scholar] [CrossRef]
- Gaspar-López, E.; García, A.J.; Landete-Castillejos, T.; Carrion, D.; Estévez, J.A.; Gallego, L. Growth of the first antler in Iberian red deer (Cervus elaphus hispanicus). Eur. J. Wildl. Res. 2008, 54, 1–5. [Google Scholar] [CrossRef]
- Zannèse, A.; Morellet, N.; Targhetta, C.; Coulon, A.; Fuser, S.; Hewison, A.J.M.; Ramanzin, M. Spatial structure of roe deer populations: Towards defining management units at a landscape scale. J. Appl. Ecol. 2006, 43, 1087–1097. Available online: https://www.jstor.org/stable/4123801 (accessed on 1 April 2023). [CrossRef]
- Kreulen, D. Wildebeest habitat selection in the Serengeti plains, Tanzania, inrelation to calcium and lactation: A preliminary report. East Afr. Wildl. J. 1975, 3, 297–304. [Google Scholar] [CrossRef]
- McNaughton, S.J. Mineral nutrition and spatial concentrations of African ungulates. Nature 1988, 334, 343–345. [Google Scholar] [CrossRef]
- McNaughton, S.J. Mineral nutrition and seasonal movements of African migratory ungulates. Nature 1990, 345, 613–615. [Google Scholar] [CrossRef]
- Alldredge, M.W.; Peek, J.M.; Wall, W.A. Nutritional quality of forages used by elk in northern Idaho. J. Range Manag. 2002, 55, 253–259. [Google Scholar] [CrossRef]
- Wilson, P.R.; Grace, N.D. A review of tissue reference values used to assess the trace element status of farmed red deer (Cervus elaphus). N. Z. Vet. J. 2001, 49, 126–132. [Google Scholar] [CrossRef]
- Ceacero, F.; Landete-Castillejos, T.; García, A.J.; Estévez, J.; Gallego, L. Can Iberian red deer (Cervus elaphus hispanicus) discriminate among essential minerals in their diet? Br. J. Nutr. 2010, 103, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Ceacero, F.; Landete-Castillejos, T.; García, A.J.; Estévez, J.A.; Gaspar-López, E.; Gallego, L. Effects of free-choice mineral consumption in Iberian red deer hinds and calves. Anim. Prod. Sci. 2010, 50, 37–44. [Google Scholar] [CrossRef]
- Tajchman, K.; Steiner-Bogdaszewska, Ż.; Żółkiewski, P. Requirements and role of selected micro and macro elements in nutrition of cervids (Cervidae)-Review. Appl. Ecol. Environ. Res. 2018, 16, 7669–7686. [Google Scholar] [CrossRef]
- Stacja Badawcza Instytutu Parazytologii PAN. Available online: https://kosewopan.pl/pl/home/ (accessed on 22 February 2023).
- Darmochwał, T.; Rumiński, M.J. Warmia Mazury Guide; TD Agency: Białystok, Poland, 1998. (In Polish) [Google Scholar]
- DEFRA. Code of Recommendations for the Welfare of Farmed Deer. IOP Publishing PhysicsWeb. 2023. Available online: http://www.defra.gov.uk/animalh/welfare/farmed/othersps/deer/pb0055/deercode.htm (accessed on 25 February 2023).
- FEDFA. The Federation of European Deer Farmers Associations. IOP Publishing PhysicsWeb. 2023. Available online: https://www.fedfa.com/en/fedfa-members/#1364 (accessed on 25 February 2023).
- Mattiello, S. Welfare issues of modern deer farming. Ital. J. Anim. Sci. 2009, 8, 205–217. [Google Scholar] [CrossRef]
- Tajchman, K.; Ukalska-Jaruga, A.; Ceacero, F.; Pecio, M.; Steiner-Bogdaszewska, Ż. Concentration of Macroelements and Trace elements in Farmed Fallow Deer Antlers Depending on Age. Animals 2022, 12, 3409. [Google Scholar] [CrossRef]
- Mirek, Z.; Piękoś-Mirkowa, H.; Zając, A.; Zając, M. Flowering Plants and Pteridophytes of Poland. A Checklist; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2002. [Google Scholar]
- Novák, J. Evaluation of grassland quality. Ekol. Bratisl. 2004, 232, 127–143. [Google Scholar]
- AOAC. Official Methods of Analysis of AOAC International, 18th ed.; Horwitz, W., Jr., Latimer, G.W., Eds.; Revision 4; AOAC International: Gaithersburg, MD, USA, 2011. [Google Scholar]
- Verite, R.; Journet, M.; Jarrige, R. A new system for the protein feeding of ruminants: The PDI system. Lives. Prod. Sci. 1979, 6, 349–367. [Google Scholar] [CrossRef]
- Ru, Y.J.; Fisher, M.; Glatz, P.C.; Wyatt, S.; Swanson, K.; Falkenberg, S. Forage Intakeand Nutrient Requirements of Fallow Weaner Deer in Southern Australia. Asian-Aust. J. Anim. Sci. 2003, 16, 685–692. [Google Scholar] [CrossRef]
- Ru, Y.J.; Fortune, J.A. The effect of grazing intensity and cultivars on morphology, phenology and nutritive value of subterranean clover. II. Nutritive value during the growing season. Aust. J. Agric. Res. 2000, 51, 1047–1055. [Google Scholar] [CrossRef]
- Karpowicz, A. Farm rearing of Deer and Fallow Deer; Malopolska Agricultural Advisory Centre: Karniowice, Poland, 2012. (In Polish)
- Luo, X.; Wang, L.; Cao, T.; He, W.; Lu, S.; Li, F.; Zhang, Z.; Chang, T.; Tian, X. Legacy effect of plant chemical defence substances on litter decomposition. Plant Soil 2023, 287. [Google Scholar] [CrossRef]
- Kumara, S.N.; Parkinson, T.J.; Laven, R.; Donaghy, D.J. The Influence of Rotational Length, along with Pre-and Post-Grazing Measures on Nutritional Composition of Pasture during Winter and Spring on New Zealand Dairy Farms. Animals 2022, 12, 1934. [Google Scholar] [CrossRef]
- Bergvall, U.A.; Co, M.; Bergström, R.; Sjöberg, P.J.; Waldebäck, M.; Turner, C. Anti-browsing effects of birch bark extract on fallow deer. Eur. J. For. Res. 2013, 132, 717–725. [Google Scholar] [CrossRef]
- Borkowski, J.; Obidziński, A. The composition of the autumn and winter diets in two Polish populations of fallow deer. Acta Theriol. 2003, 48, 539–546. [Google Scholar] [CrossRef]
- Landete-Castillejos, T.; Currey, J.D.; Estevez, J.A.; Fierro, Y.; Calatayud, A.; Ceacero, F.; Garcia, A.J.; Gallego. Do drastic Feather effects on diet influence chan ges In chemical composition, mechanical properties and structure in deer antlers? Bone 2010, 47, 815–825. [Google Scholar] [CrossRef]
- Strzetelski, J.A.; Brzóska, F.; Kowalski, Z.M.; Osięgłowski, S. Feeding Recommendation for Ruminants and Feed Tables; Publisher MA-NRI INRA: Krakow, Poland, 2014. [Google Scholar]
- Månsson, J.; Nilsson, L.; Felton, A.M.; Jarnemo, A. Habitat and crop selection by red deer in two different landscape types. Agric. Ecosyst. Environ. 2021, 318, 107483. [Google Scholar] [CrossRef]
- Dryden, G.M. Quantitative nutrition of deer: Energy, protein and water. Anim. Prod. Sci. 2011, 51, 292–302. [Google Scholar] [CrossRef]
- Marshal, J.P.; Krausman, P.R.; Bleich, V.C. Rainfall, temperature, and forage dynamics affect nutritional quality of desert mule deer forage. Rangel. Ecol. Manag. 2005, 58, 360–365. [Google Scholar] [CrossRef]
- Marell, A.; Hofgaard, A.; Danell, K. Nutrient dynamics of reindeer forage species along snow-melt gradients at different ecological scales. Basic Appl. Ecol. 2006, 7, 13–30. [Google Scholar] [CrossRef]
- Wam, H.K.; Histol, T.; Nybakken, L.; Solberg, E.J.; Hjeljord, O. Transient nutritional peak in browse foliage after forest clearing advocates cohort management of ungulates. Basic Appl. Ecol. 2016, 17, 252–261. [Google Scholar] [CrossRef]
- Perkins, J.R. Supplemental feeding. Texas Parks and Wildlife Department Fisheries and Wildlife Division. Contribution of Federal Aid Project W-129-M. Reproduced from PWD BK W7000-033 (11/91). 1991. Available online: https://tpwd.texas.gov/publications/pwdpubs/media/pwd_bk_w7000_0033.pdf (accessed on 1 April 2023).
- Obidziński, A.; Kiełtyk, P.; Borkowski, J.; Bolibok, L.; Remuszko, K. Autumn-winter diet overlap of fallow, red, and roe deer in forest ecosystems, Southern Poland. Open Life Sci. 2013, 8, 8–17. [Google Scholar] [CrossRef]
- USDA-NRCS. Wildlife Food Plot. Fish and Wildlife Habitat Management Guidesheet; USDANRCS: Minneapolis, MN, USA, 1999.
- Shin, H.T.; Hudson, R.J.; Gao, X.H.; Suttie, J.M. Nutritional requirements and management strategies for farmed deer-review. Asian-Australas. J. Anim. Sci. 2000, 13, 561–573. [Google Scholar] [CrossRef]


{kind=link}
| Feature | Summer Sown Pen (SS) | Summer Pen (S) | Winter Pen (W) | Control Area (C) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| M | SE | t | M | SE | t | M | SE | t | |||
| Area (ha) | 9.15 | - | - | 4.5 | - | - | 9.5 | - | - | 9.0 | |
| Animal density (pcs ha−1) | 12 | - | - | 2 | - | - | 12 | - | - | - | |
| Body mass of fallow deer (kg) | before grazing | 64.5 | 4.68 | −4.01, p = 0.001 * | 29.5 | 4.85 | −3.42, p = 0.004 * | 31 | 5.23 | 2.13, p = 0.049 * | - |
| after grazing | 93.9 | 3.11 | 48.5 | 3.47 | 42 | 5.63 | - | ||||
| Weight gain (kg) | 29.5 | 3.18 | - | 19 | 3.40 | 11 | 3.04 | - | - | ||
| Number of grazing months | 8 (April–November) | - | - | 8 (April–November) | - | - | 4 (December–March) | - | - | - | |
| Area with trees (ha) | 3.3 | - | - | 1.3 | - | - | 0.2 | - | - | 0.5 | |
| Tree species | Mainly Betula pendula, Malus sylvestris, Malus domestica, Prunus domestica, Pinus sylvestris | ||||||||||
| Groups of Plant Species | Pens | Control Area (C) | |||
|---|---|---|---|---|---|
| Summer (S) | Winter (W) | Summer Sown (SS) | |||
| Valuable grasses FV = 7–8 | Mean | 65.7 | 47.4 | 66.1 | 46.1 |
| Species * | Poa pratensis, Dactylis glomerata, Alopecurus pratensis, Lolium perenne | Poa pratensis, Dactylis glomerata, Lolium perenne | Poa pratensis, Dactylis glomerata, Alopecurus pratensis, Lolium perenne, Festuca pratensis, Phleum pratense | Dactylis glomerata, Arrhenatherum elatius | |
| Other grasses FV ≤ 6 | Mean | 7.1 | 7.3 | 16.3 | 14.4 |
| Species * | Festuca rubra | Festuca rubra | Avenula pubescens, Festuca rubra | Elymus repens, Festuca rubra | |
| Legumes FV = 7–8 | Mean | 8.1 | 11.2 | 8.3 | 0.9 |
| Species * | Trifolium repens | Trifolium repens | Trifolium repens | - | |
| Sedges FV = 2 | Mean | 0.1 | 0 | 0.3 | 0 |
| Species * | - | - | - | - | |
| Herbs FV = 4–6 | Mean | 9.6 | 7 | 4.7 | 16.1 |
| Species * | Taraxacum officinale | Taraxacum officinale | - | Achillea millefolium | |
| Weeds FV ≤ 3 | Mean | 9.4 | 27.1 | 4.3 | 22.5 |
| Species * | Urtica dioica | Geranium pusillum, Polygonum aviculare, Urtica dioica | - | Artemisia vulgaris, Chaerophyllum aromaticum | |
| Study Objects | Season | Dry Matter % | Crude Ash g kg−1 DM | Crude Protein g kg−1 DM | Crude Fat g kg−1 DM | Crude Fibre g kg−1 DM | NFE g kg−1 DM |
|---|---|---|---|---|---|---|---|
| Summer pen (S) | Spring | 16.52 ± 2.83 a | 94.37 a ± 9.52 | 223.36 b ± 38.93 | 28.53 a ± 2.78 | 249.26 a ± 33.36 | 404.48 a ± 54.30 |
| Autumn | 30.07 b ± 5.66 | 87.54 a ± 7.64 | 116.94 a ± 31.91 | 27.79 a ± 3.10 | 281.66 a ± 23.17 | 486.08 b ± 19.15 | |
| Total | 23.30 ± 8.43 | 90.95 ± 8.58 | 170.15 ± 66.41 | 28.16 ± 2.66 | 265.46 ± 31.22 | 445.28 ± 57.65 | |
| Winter pen (W) | Spring | 28.91 a ± 0.01 | 111.78 a ± 1.69 | 274.08 b ± 0.49 | 24.76 a ± 0.18 | 197.64 a ± 0.04 | 391.75 a ± 1.05 |
| Autumn | 24.84 a ± 6.30 | 93.36 a ± 14.31 | 167.65 a ± 41.31 | 26.53 a ± 3.91 | 288.12 a ± 43.96 | 424.35 a ± 15.08 | |
| Total | 26.46 ± 4.98 | 100.73 ± 14.31 | 210.22 ± 65.20 | 25.82 ± 2.93 | 251.92 ± 58.50 | 411.31 ± 20.81 | |
| Summer sown pen (SS) | Spring | 18.79 a ± 1.42 | 91.67 a ± 4.38 | 208.57 b ± 17.55 | 28.59 a ± 8.68 | 233.73 a ± 13.60 | 437.45 a ± 2.45 |
| Autumn | 30.08 b ± 2.32 | 85.63 a ± 3.82 | 126.21 a ± 10.24 | 25.98 a ± 6.29 | 296.65 ± 15.87 | 465.53 a ± 4.69 | |
| Total | 24.43 ± 6.42 | 88.65 ± 4.94 | 167.39 ± 46.91 | 27.28 ± 6.93 | 265.18 ± 36.91 | 451.49 ± 15.74 | |
| Control area (C) | Spring | 21.43 a ± 0.56 | 99.92 a ± 1.20 | 176.69 a ± 11.71 | 27.64 a ± 11.08 | 240.50 a ± 7.21 | 455.24 a ± 1.27 |
| Autumn | 20.39 a ± 3.53 | 99.77 a ± 11.98 | 118.23 a ± 19.87 | 25.11 a ± 2.32 | 314.24 a ± 33.79 | 442.64 a ± 18.66 | |
| Total | 20.91 ± 2.33 | 99.85 ± 7.61 | 147.46 ± 35.19 | 26.38 ± 7.29 | 277.37 ± 45.92 | 448.94 ± 13.69 |
| Study Objects | Season | ME (MJ) | UFL | dCP (g) | PDI (g) |
|---|---|---|---|---|---|
| Summer pen (S) | Spring | 11.05 b ± 0.40 | 0.83 b ± 0.04 | 125.08 b ± 21.80 | 77.82 b ± 4.25 |
| Autumn | 9.81 a ± 0.35 | 0.72 a ± 0.03 | 65.49 a ± 17.87 | 63.77 a ± 5.00 | |
| Total | 10.43 ± 0.76 | 0.77 ± 0.07 | 95.28 ± 37.19 | 70.80 ± 8.74 | |
| Winter pen (W) | Spring | 11.53 b ± 0.02 | 0.88 b ± 0.00 | 153.48 b ± 0.27 | 83.65 b ± 0.14 |
| Autumn | 10.34 a ± 0.43 | 0.77 a ± 0.04 | 93.88 a ± 23.13 | 71.07 a ± 4.18 | |
| Total | 10.82 ± 0.72 | 0.81 ± 0.07 | 117.72 ± 36.51 | 76.10 ± 7.50 | |
| Summer sown pen (SS) | Spring | 10.92 b ± 0.17 | 0.82 b ± 0.02 | 116.80 b ± 9.83 | 76.18 b ± 3.01 |
| Autumn | 9.91 a ± 0.14 | 0.73 a ± 0.01 | 70.68 a ± 5.73 | 66.41 a ± 0.40 | |
| Total | 10.41 ± 0.57 | 0.77 ± 0.05 | 93.74 ± 26.27 | 71.30 ± 5.68 | |
| Control area (C) | Spring | 10.45 a ± 0.10 | 0.78 a ± 0.01 | 98.95 a ± 6.56 | 71.81 a ± 2.57 |
| Autumn | 9.66 a ± 0.22 | 0.71 a ± 0.02 | 66.21 a ± 11.12 | 64.14 a ± 2.65 | |
| Total | 10.05 ± 0.46 | 0.74 ± 0.04 | 82.58 ± 19.70 | 67.98 ± 4.80 |
| Study Objects | Season | Ca (g) | P (g) | Mg (g) | Na (g) | K (g) | Cu (mg) | Zn (mg) | Fe (mg) |
|---|---|---|---|---|---|---|---|---|---|
| Summer pen (S) | Spring | 5.35 ± 1.31 | 4.83 ± 0.61 | 2.18 ± 0.38 | 0.36 ± 0.10 | 36.12 ± 4.51 | 7.6 ± 1.7 | 25.5 ± 5.0 | 143.1 ± 72.2 |
| Autumn | 5.16 ± 0.43 | 3.20 ± 0.44 | 2.03 ± 0.14 | 0.25 ± 0.07 | 24.14 ± 5.04 | 6.9 ± 1.4 | 47.9 ± 4.3 | 83.1 ± 15.6 | |
| Total | 5.26 ± 0.88 | 4.01 ± 1.01 | 2.11 ± 0.26 | 0.31 ± 0.10 | 30.14 ± 7.83 | 7.2 ± 1.5 | 36.7 ± 13.0 | 113.1 ± 57.1 | |
| Winter pen (W) | Spring | 8.13 ± 0.05 | 4.81 ± 0.20 | 2.45 ± 0.01 | 0.59 ± 0.24 | 32.47 ± 0.72 | 5.8 ± 0.1 | 34.7 ± 0.2 | 357.8 ± 22.7 |
| Autumn | 6.95 ± 1.52 | 3.61 ± 0.34 | 2.34 ± 0.17 | 0.64 ± 0.37 | 31.15 ± 6.10 | 6.1 ± 0.4 | 30.9 ± 4.4 | 106.2 ± 2.2 | |
| Total | 7.42 ± 1.26 | 4.10 ± 0.71 | 2.38 ± 0.14 | 0.62 ± 0.29 | 31.69 ± 4.39 | 6.0 ± 0.4 | 32.4 ± 3.7 | 206.8 ± 138.3 | |
| Summer sown pen (SS) | Spring | 7.03 ± 1.20 | 4.87 ± 0.32 | 2.28 ± 0.38 | 0.41 ± 0.18 | 33.68 ± 1.58 | 9.0 ± 0.7 | 28.8 ± 2.6 | 127.8 ± 23.2 |
| Autumn | 6.14 ± 0.75 | 3.14 ± 0.49 | 2.17 0.06 | 0.32 ± 0.11 | 19.53 ± 2.34 | 7.3 ± 2.0 | 87.3 ± 69.5 | 149.5 ± 44.2 | |
| Total | 6.58 ± 1.02 | 4.01 ± 1.02 | 2.22 ± 0.24 | 0.36 ± 0.14 | 26.61 ± 7.95 | 8.2 ± 1.6 | 58.0 ± 54.4 | 138.6 ± 33.7 | |
| Control area (C) | Spring | 7.64 ± 0.40 | 4.40 ± 0.46 | 2.40 ± 0.10 | 0.33 ± 0.11 | 34.24 ± 2.76 | 3.1 ± 2.6 | 37.4 ± 4.3 | 159.0 ± 52.1 |
| Autumn | 11.81 ± 0.86 | 3.72 ± 0.64 | 2.91 ± 0.12 | 0.58 ± 0.35 | 22.12 ± 4.37 | 5.4 ± 0.3 | 47.2 ± 14.7 | 128.3 ± 57.0 | |
| Total | 9.72 ± 2.37 | 4.06 ± 0.62 | 2.65 ± 0.29 | 0.45 ± 0.27 | 28.18 ± 7.40 | 4.4 ± 1.8 | 42.3 ± 11.1 | 143.6 ± 51.2 |
| Analysed Variable | Spring | Autumn | ||||
|---|---|---|---|---|---|---|
| F | p | Differences between Pens | F | p | Differences between Pens | |
| Crude ash (g kg−1 DM) | 8.475 | 0.007 * | W > S (p = 0.016); W > SS (p = 0.007) | 1.159 | 0.383 | - |
| Crude protein (g kg−1 DM) | 10.077 | 0.004 * | W > S (p = 0.027); W > C (p = 0.003) | 2.134 | 0.174 | - |
| Crude fat (g kg−1 DM) | 0.188 | 0.901 | - | 0.215 | 0.883 | - |
| Crude fibre (g kg−1 DM) | 4.570 | 0.038 * | S > W (p = 0.035) | 0.619 | 0.622 | - |
| NFE (g kg−1 DM) | 3.471 | 0.070 | - | 8.993 | 0.006 * | S > W (p = 0.005); S > C (p = 0.036); SS > W (p = 0.047) |
| Ca (g) | 5.312 | 0.026 * | W > S (p = 0.040) | 24.957 | <0.001 * | C > S (p < 0.001); C > W (p = 0.002); C > SS (p < 0.001) |
| P (g) | 1.405 | 0.310 | - | 1.089 | 0.407 | - |
| Mg (g) | 0.819 | 0.518 | - | 22.771 | <0.001 * | C > S (p < 0.001); C > W (p = 0.005); C > SS (p = 0.001) |
| Na (g) | 3.987 | 0.052 | - | 1.647 | 0.254 | - |
| K (g) | 0.946 | 0.462 | - | 3.759 | 0.059 | - |
| Cu (mg) | 2.447 | 0.138 | - | 1.395 | 0.313 | - |
| Zn (mg) | 6.641 | 0.014 * | C > S (p = 0.015) | 1.334 | 0.329 | - |
| Fe (mg) | 15.533 | 0.001 * | W > S (p = 0.002); W > SS (p = 0.001); W > C (p = 0.004) | 1.807 | 0.223 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulik, M.; Tajchman, K.; Lipiec, A.; Bąkowski, M.; Ukalska-Jaruga, A.; Ceacero, F.; Pecio, M.; Steiner-Bogdaszewska, Ż. The Impact of Rotational Pasture Management for Farm-Bred Fallow Deer (Dama dama) on Fodder Quality in the Context of Animal Welfare. Agronomy 2023, 13, 1155. https://doi.org/10.3390/agronomy13041155
Kulik M, Tajchman K, Lipiec A, Bąkowski M, Ukalska-Jaruga A, Ceacero F, Pecio M, Steiner-Bogdaszewska Ż. The Impact of Rotational Pasture Management for Farm-Bred Fallow Deer (Dama dama) on Fodder Quality in the Context of Animal Welfare. Agronomy. 2023; 13(4):1155. https://doi.org/10.3390/agronomy13041155
Chicago/Turabian StyleKulik, Mariusz, Katarzyna Tajchman, Antoni Lipiec, Maciej Bąkowski, Aleksandra Ukalska-Jaruga, Francisco Ceacero, Monika Pecio, and Żaneta Steiner-Bogdaszewska. 2023. "The Impact of Rotational Pasture Management for Farm-Bred Fallow Deer (Dama dama) on Fodder Quality in the Context of Animal Welfare" Agronomy 13, no. 4: 1155. https://doi.org/10.3390/agronomy13041155