
Parasitism Potential and Laboratory Rearing of Psyllaephagus sp., a Newly Discovered Parasitoid of Cacopsylla chinensis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Insects
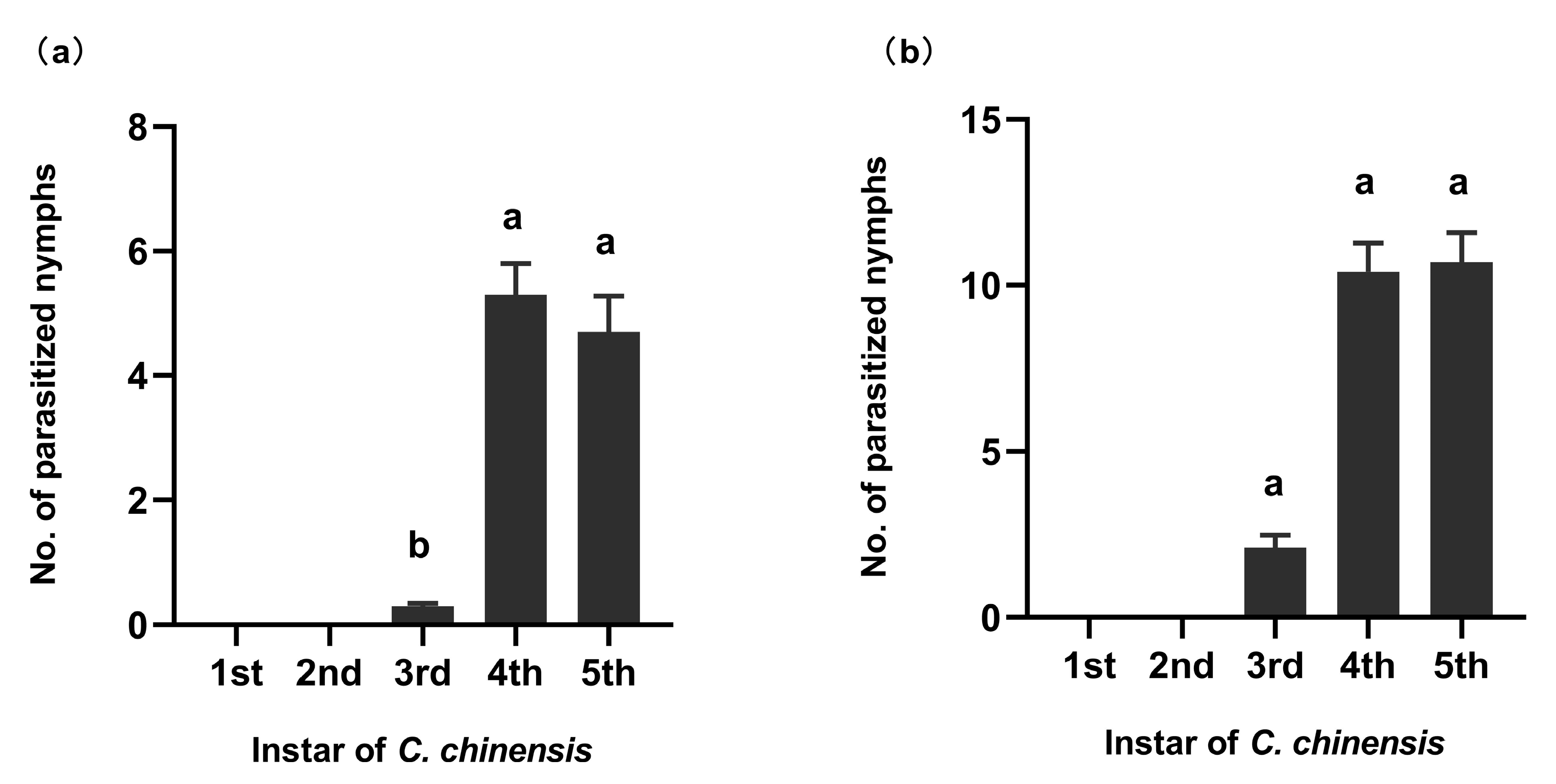
2.2. Wasps’ Host Stage Preference
2.3. Developmental Duration and Fecundity of Psyllaephagus sp.
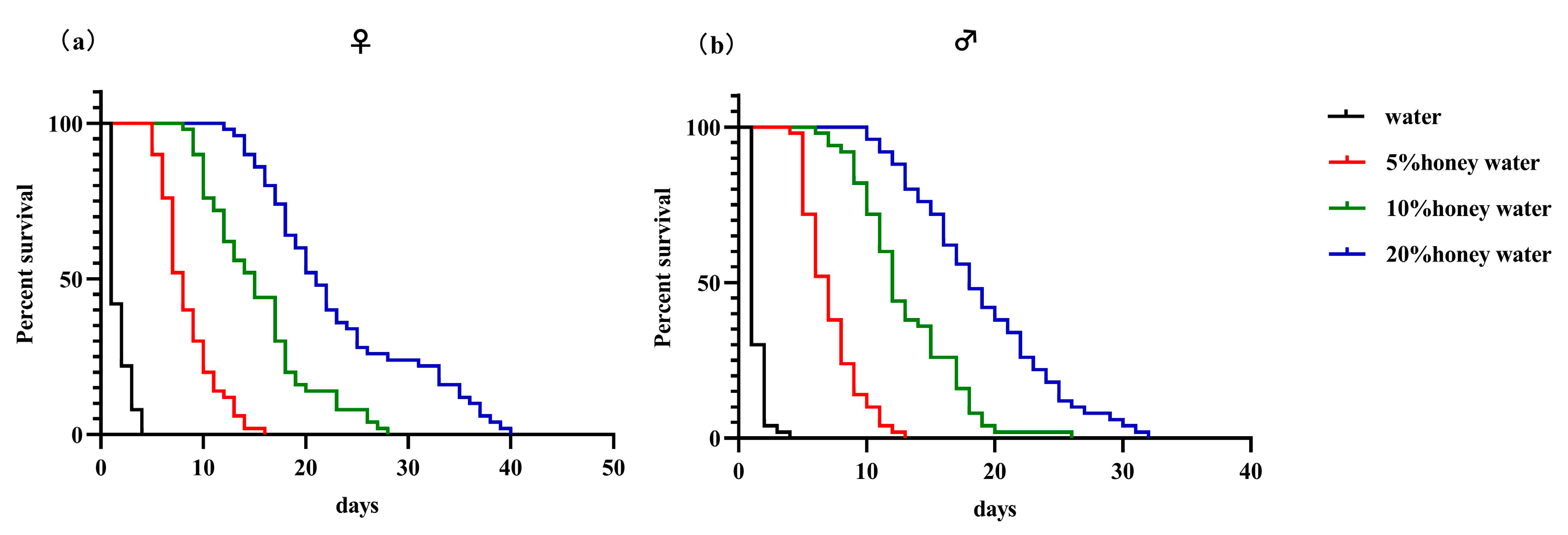
2.4. Influence of Supplementary Nutrition on Psyllaephagus sp. Adult Longevity
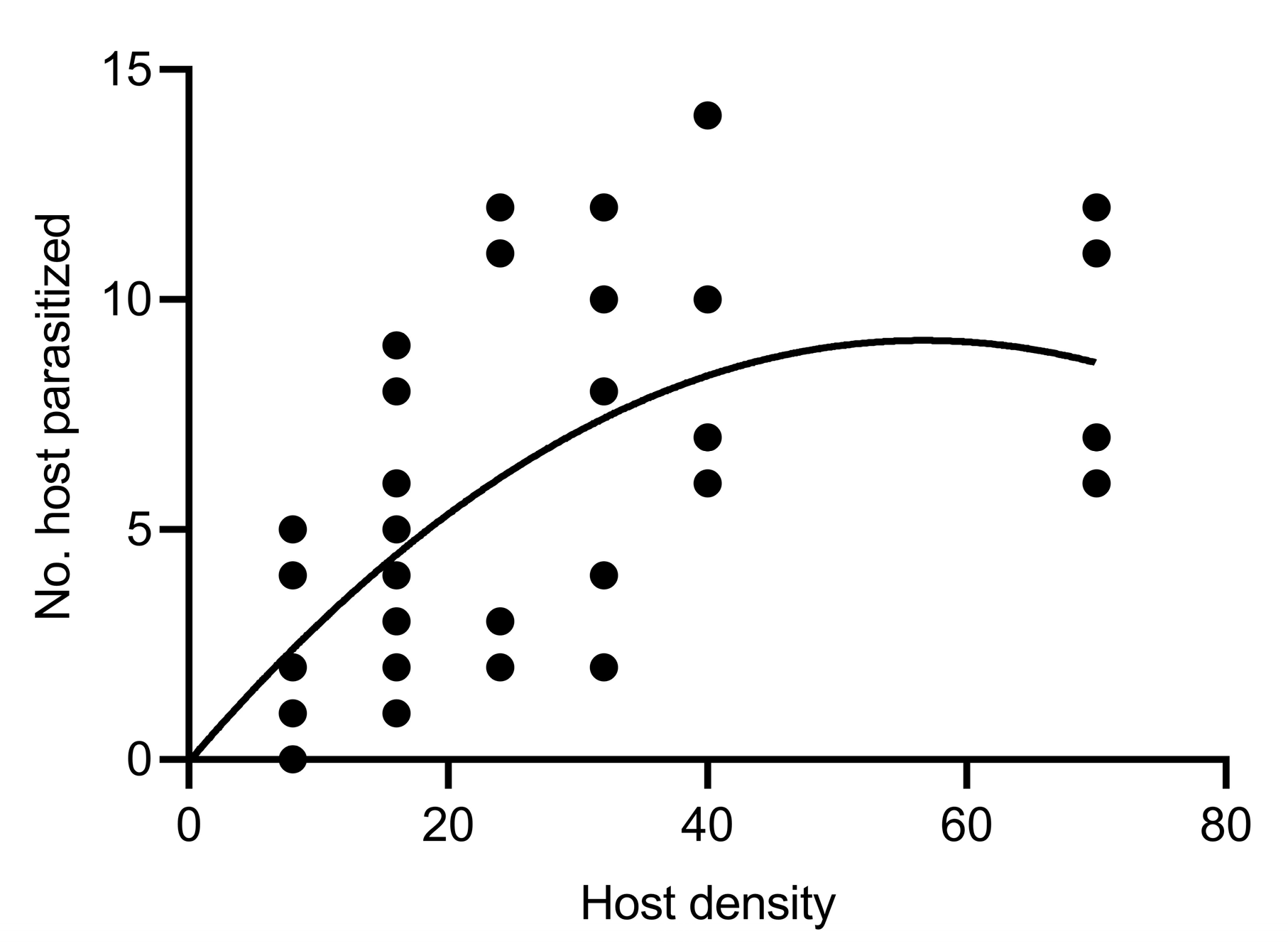
2.5. Functional Response of Psyllaephagus sp. Females to C. chinensis Nymphs
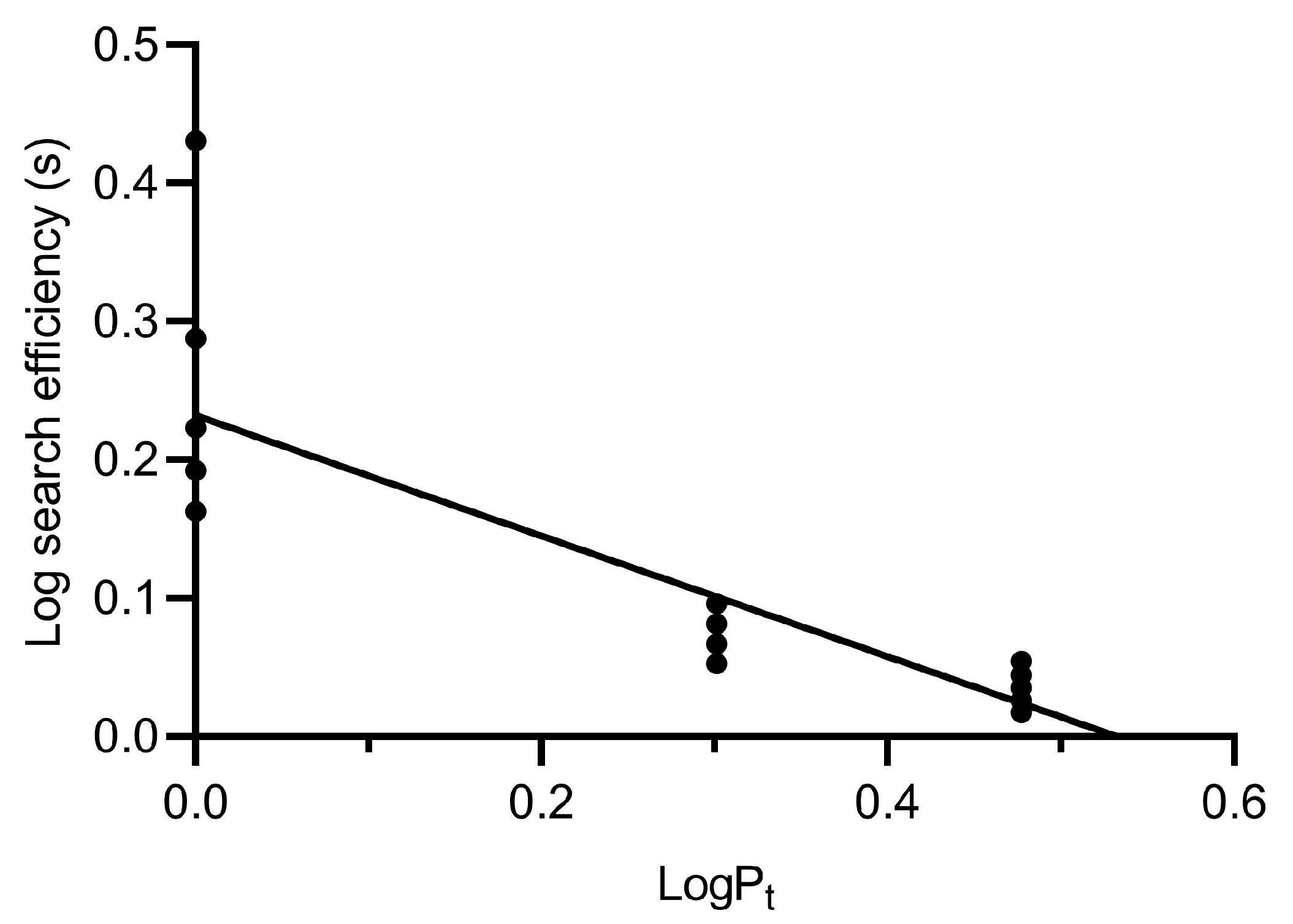
2.6. Optimal Host/Parasitoid Ratios
2.7. Statistical Analysis
3. Results
3.1. Wasps’ Host Stage Preference
3.2. Developmental Duration and Fecundity of Psyllaephagus sp.
3.3. Influence of Supplementary Nutrition on Psyllaephagus sp. Adult Longevity
3.4. Functional Response of Psyllaephagus sp. Females to C. chinensis Nymphs
3.5. Optimal Host/Parasitoid Ratios
4. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dong, Y.H.; Qian, J.R.; Zhou, L.Q.; Wang, H.S. Study on occurrence law and control strategy of Psylla chinensis in southern China. Acta Agric. Jiangxi 2009, 21, 118–120. [Google Scholar]
- Qiao, L.X. Occurrence and comprehensive control of Psylla chinensis in China. Mod. Rural Sci. Technol. 2017, 6, 32. [Google Scholar]
- DuPont, S.T.; Strohm, C.; Nottingham, L.; Rendon, D. Evaluation of an integrated pest management program for central Washington pear orchards. Biol. Control 2021, 152, 104390. [Google Scholar] [CrossRef]
- Gullan, P.J.; Martin, J.H. Sternorrhyncha (jumping plant-lice, whiteflies, aphids and scale insects). In Encyclopedia of Insects, 2nd ed.; Resh, V.H., Cardé, R.T., Eds.; Academic Press: Cambridge, MA, USA, 2009; pp. 957–967. [Google Scholar] [CrossRef]
- Li, D.L.; Wang, P.; Zhang, C.T. Review of research status and control of Cacopsylla chinensis in China. Shanxi Fruits 2003, 4, 30–31. [Google Scholar]
- Noyes, J.S. Universal Chalcidoidea Database. Available online: https://www.nhm.ac.uk/chalcidoids (accessed on 22 November 2022).
- Wu, F.; Zhen, W.; Yang, Z.; Zu, G. A new species of Psyllaephagus (Hymenoptera: Encyrtidae) from China.; parasitoid of Macrohomotoma sinica (Hemiptera: Homotomidae) on Ficus concinna. Biodiv. Data J. 2021, 9, e63253. [Google Scholar] [CrossRef]
- Ma, F.L. Systematic Studies on Encyrtidae (Hymenoptera: Chalcidoidea) from Northeastern China. Ph.D. Thesis, Northeast Forestry University, Liaoning, China, 2004. [Google Scholar]
- Li, H.L. The Taxonomic and Biodiversity of Encyrtidae from Shanghai City. Ph.D. Thesis, Shanghai Normal University, Shanghai, China, 2010. [Google Scholar]
- Tan, Y.G.; Zhao, J.Z. One new species and one newly recorded species of the genus Psyllaephagus Ashmead from China (Hymenoptera: Encyrtidae). J. Hubei Univ. 1999, 21, 174–176. [Google Scholar]
- Xu, Z.H.; Chen, W.; Yu, H.; Li, B.J. Notes on Psyllaephagus.; a genus new to China with descriptions of two new species (Hymenoptera: Encyrtidae). Sci. Silvae Sin. 2000, 36, 39–41. [Google Scholar]
- Xu, Z.H.; Chou, L.Y.; Hong, S.C. Notes on three encyrtid parasitoids of Triozasyzygii in Taiwan with description of one new species (Hymenoptera: Encyrtidae). Chin. J. Entomol. 2000, 20, 9–12. [Google Scholar]
- Zhang, Y.Z. Generic Revision of Chinese Encyrtidae (Hymenoptera: Chalcidoidea). Ph.D. Thesis, Chinese Academy of Sciences, Beijing, China, 2001. [Google Scholar]
- Tang, X.L.; Zhang, Y.Z.; Hu, H.Y. Seven new record species of Psyllaephagus (Hymenoptera: Encyrtidae) from China. Entomotaxonomia 2016, 38, 63–78. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, P.C.; Ma, B.X.; Zhang, Y.Z. Psyllaephagus arenarius (Hymenoptera: Encyrtidae), a newly recorded parasitoid of Bactericera gobica (Hemiptera: Psyllidae) in China. Acta Entomol. Sin. 2017, 60, 842–846. [Google Scholar] [CrossRef]
- Noyes, J.S.; Fallahzadeh, M. Psyllaephagus zdeneki sp. nov. (Hymenoptera: Encyrtidae) from Iran.; A parasitoid of Euphyllura pakistanica (Hemiptera: Psyllidae). Acta. Soc. Zool.Bohem. 2005, 69, 203–208. [Google Scholar]
- Dahlsten, D.L.; Rowney, D.L.; Copper, W.A.; Tassan, R.L.; Chaney, W.E.; Robb, K.L.; Tjosvold, S.; Bianchi, M.; Lane, P. Parasitoid wasp controls blue gum psyllid. Calif. Agr. 1998, 52, 31–34. [Google Scholar] [CrossRef] [Green Version]
- Noyes, J.S.; Hanson, P. Encyrtidae (Hymenoptera: Chalcidoidea) of Costa Rica: The genera and species associated with jumping plant-lice (Homoptera: Psylloidea). Bull. Nat. Hist. Museum. Entomol. Ser. 1996, 65, 105–164. [Google Scholar]
- Daane, K.M.; Sime, K.R.; Dahlsten, D.L.; Andrews, J.W.; Zuparko, R.L. The biology of Psyllaephagus bliteus Riek (Hymenoptera: Encyrtidae).; a parasitoid of the red gum lerp psyllid (Hemiptera: Psylloidea). Biol. Control 2005, 32, 228–235. [Google Scholar] [CrossRef]
- Jones, M.E.; Daane, K.M.; Paine, T.D. Establishment of Psyllaephagus parvus and P. perplexans as serendipitous biological control agents of Eucalyptus psyllids in southern California. BioControl 2011, 56, 735–744. [Google Scholar] [CrossRef] [Green Version]
- Ge, Y.; Liu, P.P.; Zhang, L.; Snyder, W.E.; Smith, O.M.; Shi, W.P. A sticky situation: Honeydew of the pear psylla disrupts feeding by its predator Orius sauteri. Pest Manag. Sci. 2020, 76, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Gai, Y.P.; Ji, X.L.; Sun, X.G.; Liu, Y.S. Studies on the external morphological characters of Psylla chineses Yang et Li nymphs. J. Shandong Agric. Univ. 2000, 31, 253–256. [Google Scholar]
- Zar, J.H. Biostatistical Analysis, 5th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2010. [Google Scholar]
- Visser, M.E.; Driessen, G. Indirect mutual interference in parasitoids. Neth. J. Zool. 1991, 41, 214–227. [Google Scholar] [CrossRef]
- Juliano, S.A. Nonlinear Curve Fitting: Predation and Functional Response Curves. Ph.D. Thesis, Oxford University, Oxford, UK, 2001. [Google Scholar]
- Campos, V.; Araya, J.E. Functional response of Nabis punctipennis Blanchard to Acyrthosiphon pisum Harris in the laboratory. Chile J. Agric. Anim. Sci. 2017, 33, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.F.; Matos, C.H.; de Oliveira, C.R.; Da Silva, T.G.; Lima Neto, I.F. Functional and numerical responses of Stethorus tridens Gordon (Coleoptera: Coccinellidae) preying on Tetranychus bastosi Tuttle, Baker & Sales (Acari: Tetranychidae) on physic nut (Jatropha curcas). Biol. Control 2017, 111, 1–5. [Google Scholar] [CrossRef]
- Ge, Y.; Zhang, L.; Qin, Z.F.; Wang, Y.; Liu, P.P.; Tan, S.Q.; Fu, Z.; Olivia, M.S.; Shi, W.P. Different predation capacities and mechanisms of Harmonia axyridis (Coleoptera: Coccinellidae) on two morphotypes of pear psylla Cacopsylla chinensis (Hemiptera: Psyllidae). PLoS ONE 2019, 14, e0215834. [Google Scholar] [CrossRef] [Green Version]
- Jalali, M.A.; Tirry, L.; De Clercq, P. Effect of temperature on the functional response of Adalia bipunctata to Myzus persicae. BioControl 2010, 55, 261–269. [Google Scholar] [CrossRef] [Green Version]
- Mahdian, K.; Vantornhout, I.; Tirry, L.; De Clercq, P. Effects of temperature on predation by the stinkbugs Picromerus bidens and Podisus maculiventris (Heteroptera: Pentatomidae) on noctuid caterpillars. Bull. Entomol. Res. 2006, 96, 489–496. [Google Scholar] [CrossRef]
- R Core Team, R. R Core Team R: A Language and Environment for Statistical Computing; R Core Team R: Vienna, Austria, 2022. [Google Scholar]
- Stehr, F.W. Parasitoides and predators in pest management. In Introduction to Insect Pest Management; Mecalf, R.L., Luckmann, W.H., Eds.; Wiley & Sons, A Wiley-Interscience Publication: New York, NY, USA, 1982. [Google Scholar]
- Hemerik, L.; Harvey, J.A. Flexible larval development and the timing of destructive feeding by a solitary endoparasitoid: An optimal foraging problem in evolutionary perspective. Ecol. Entomol. 1999, 24, 308–315. [Google Scholar] [CrossRef]
- Pacheco da Silva, V.C.; Garciaa, M.S.; Botton, M. Biology of Blepyrus clavicornis (Compere) (Hymenoptera: Encyrtidae), a parasitoid of Pseudococcus viburni (Signoret) (Hemiptera: Pseudococcidae). Rev. Brasil. Entomol. 2017, 61, 257–261. [Google Scholar] [CrossRef]
- Sandanayaka, W.R.M.; Charles, J.G.; Allan, D.J. Aspects of the reproductive biology of Pseudaphycus maculipennis (Hym: Encyrtidae), a parasitoid of obscure mealybug, Pseudococcus viburni (Hem: Pseudococcidae). Biol. Control 2009, 48, 30–35. [Google Scholar] [CrossRef]
- Boulton, R.A.; Collins, L.A.; Shuker, D.M. Beyond sex allocation: The role of mating systems in sexual selection in parasitoid wasps. Biol. Rev. 2015, 90, 599–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broadley, H.J.; Gould, J.R.; Sullivan, L.T.; Wang, X.Y.; Hoelmer, K.A.; Hickin, M.L.; Elkinton, J.S. Life history and rearing of Anastatus orientalis (Hymenoptera: Eupelmidae), an egg parasitoid of the spotted lanternfly (Hemiptera: Fulgoridae). Environ. Entomol. 2021, 50, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Mazzon, L.; Visentini, A.; Girolami, V. Sex ratio of European populations of Neodryinus typhlocybae (Ashmead) (Hymenoptera: Dryinidae) parasitoid of Metcalfa pruinosa (Say) (Homoptera: Flatidae). Frustula Entomol. 2000, 23, 30–36. [Google Scholar]
- King, B.H. Offspring sex ratios in parasitoid wasps. Q. Rev. Biol. 1987, 62, 367–369. [Google Scholar] [CrossRef]
- Bugila, A.A.A.; Franco, J.C.; Silva, E.B.; Branco, M. Suitability of five mealybug species (Hemiptera, Pseudococcidae) as hosts for the solitary parasitoid Anagyrus sp. nr. pseudococci (Girault) (Hymenoptera: Encyrtidae), Biocontrol. Sci. Techn. 2015, 25, 108–120. [Google Scholar] [CrossRef]
- King, B.H. Offspring sex ratio and number in response to proportion of host sizes and ages in the parasitoid wasp Spalangia cameroni (Hymenoptera: Pteromalidae). Environ. Entomol. 2002, 31, 505–508. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Gong, X.M.; Chen, Y.Z.; Pan, S.Y.; Dai, Y.N.; Hu, H.Y.; Liu, P.C. Effect of maternal age on primary and secondary sex ratios in the ectoparasitoid wasp Pachycrepoideus vindemmiae. Entomol. Exp. Appl. 2022, 170, 468–476. [Google Scholar] [CrossRef]
- Liu, P.C.; Men, J.; Zhao, B.; Wei, J.R. Fitness-related offspring sex allocation of Anastatus disparis, a gypsy moth egg parasitoid, on different-sized host species. Entomol. Exp. Appl. 2017, 163, 281–286. [Google Scholar] [CrossRef]
- Liu, P.C.; Wei, H.X.; Cao, D.D.; Wei, J.R. Relationships amongst sex ratio of progeny in Anastatus disparis (Hymenoptera: Eupelmidae), sperm depletion and decreased fecundity. Appl. Entomol. Zool. 2020, 55, 25–30. [Google Scholar] [CrossRef]
- Cochard, P.; Galstian, T.; Cloutier, C. The proportion of blue light affects parasitoid wasp behavior in LED-extended photoperiod in greenhouses: Increased parasitism and offspring sex ratio bias. Biol. Control 2019, 133, 9–17. [Google Scholar] [CrossRef]
- Murray, T.J.; Withers, T.M.; Mansfield, S. Choice versus no-choice test interpretation and the role of biology and behavior in parasitoid host specificity tests. Biol. Control 2010, 52, 153–159. [Google Scholar] [CrossRef]
- Yang, X.B.; Campos-Figueroa, M.; Silva, A.; Henne, D.C. Functional response.; prey stage preference, and mutual interference of the Tamarixia triozae (Hymenoptera: Eulophidae) on tomato and bell pepper. J. Econ. Entomol. 2015, 108, 414–424. [Google Scholar] [CrossRef]
- Islam, K.S.; Copland, M.J.W. Host preference and progeny sex ratio in a solitary koinobiont mealybug endoparasitoid, Anagyrus pseudococci (Girault), in response to its host stage. Biocontrol Sci. Techn. 1997, 7, 449–456. [Google Scholar] [CrossRef]
- Tillman, P.G. Functional response of Microplitis croceipes and Cardiochiles nigriceps (Hymenoptera: Braconidae) to variation in density of tobacco budworm (Lepidoptera: Noctuidae). Environ. Entomol. 1996, 25, 524–528. [Google Scholar] [CrossRef]
- Savino, V.; Coviella, C.E.; Luna, M.G. Reproductive biology and functional response of Dineulophus phtorimaeae, a natural enemy of the tomato moth Tuta Absoluta. J. Insect Sci. 2012, 12, 153. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.P.; Li, H.M.; Lu, Y.H.; Zhang, F.; Tim, H.; Ulrich, K.; Wu, K.M. Functional response and mutual interference of Peristenus spretus (Hymenoptera: Braconidae), a parasitoid of Apolygus lucorum (Heteroptera: Miridae). Biocontr. Sci. Technol. 2014, 24, 247–256. [Google Scholar] [CrossRef]
- Ebrahimifar, J.; Jamshidnia, A.; Allahyari, H. Functional Response of Eretmocerus delhiensis on Trialeurodes vaporariorum by Parasitism and Host Feeding. J. Insect Sci. 2017, 17, 56. [Google Scholar] [CrossRef] [Green Version]
- Tazerouni, Z.; Talebi, A.A.; Fathipour, Y.; Soufbaf, M. Agespecific functional response of Aphidius matricariae and Praon volucre (Hym.: Braconidae) on Aphis gossypii (Hem.: Aphididae). J. Entomol. Soc. Iran 2017, 36, 239–248. [Google Scholar]
- Bodino, N.; Ferracini, C.; Tavella, L. Functional response and age specific foraging behaviour of Necremnus tutae and N. Cosmopterix, native natural enemies of the invasive pest Tuta absoluta in Mediterranean area. J. Pest Sci. 2019, 92, 1467–1478. [Google Scholar] [CrossRef]
- Hassell, M.P.; Lawton, J.H.; Beddington, J.R. Sigmoid functional responses by invertebrate predators and parasitoids. J. Anim. Ecol. 1977, 46, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Van Lenteren, J.; Bakker, K. Functional responses in invertebrates. Neth. J. Zool. 1975, 26, 567–572. [Google Scholar] [CrossRef] [Green Version]
- Sagarra, L.; Vincent, C.; Peters, N.; Stewart, R.J. Effect of host density, temperature, and photoperiod on the fitness of Anagyrus kamali, a parasitoid of the hibiscus mealybug Maconellicoccus hirsutus. Entomol. Exp. Appl. 2000, 96, 141–147. [Google Scholar] [CrossRef]
- Holling, C.S. Some characteristics of simple types of predation and parasitism. Can. Entomol. 1959, 91, 385–398. [Google Scholar] [CrossRef]
- Tazerouni, Z.; Talebi, A.A.; Rezaei, M. Functional response of parasitoids: Its impact on biological control. In Parasitoids: Biology, Behavior and Ecology; Donnelly, E., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2019; pp. 35–58. [Google Scholar]
- Streams, F.A. Effect of prey size on attack components of the functional response by Notonecta undulata. Oecologia 1994, 98, 57–63. [Google Scholar] [CrossRef]
- Ziaei Madbouni, M.A.; Samih, M.; Namvar, P.; Biondi, A. Temperature-dependent functional response of Nesidiocoris tenuis (Hemiptera: Miridae) to different densities of pupae of cotton whitefly, Bemisia tabaci (Hemiptera: Aleyrodidae). Eur. J. Entomol. 2017, 114, 325–331. [Google Scholar] [CrossRef]
- Hemerik, L.; Yano, E. Scaling up from individual behaviour of Orius sauteri foraging on Thrips palmi to its daily functional response. Popul. Ecol. 2011, 53, 563–572. [Google Scholar] [CrossRef] [Green Version]
- Okuyama, T. On selection of functional response models: Holling’s models and more. BioControl 2013, 58, 293–298. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Distilled Water | 5% Honey Water | 10% Honey Water | 20% Honey Water | |
|---|---|---|---|---|
| Females | 1.72 ± 0.14 Da | 8.44 ± 0.38 Ca | 15.32 ± 0.74 Ba | 23.20 ± 1.12 Aa |
| Males | 1.36 ± 0.09 Db | 7.14 ± 0.30 Cb | 13.08 ± 0.57 Bb | 19.00 ± 0.80 Ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, Z.; Feng, M.; Zhang, L.; Ge, Y.; Huang, X.; Shi, W. Parasitism Potential and Laboratory Rearing of Psyllaephagus sp., a Newly Discovered Parasitoid of Cacopsylla chinensis. Agronomy 2023, 13, 943. https://doi.org/10.3390/agronomy13030943
Qin Z, Feng M, Zhang L, Ge Y, Huang X, Shi W. Parasitism Potential and Laboratory Rearing of Psyllaephagus sp., a Newly Discovered Parasitoid of Cacopsylla chinensis. Agronomy. 2023; 13(3):943. https://doi.org/10.3390/agronomy13030943
Chicago/Turabian StyleQin, Zifang, Mingyue Feng, Liu Zhang, Yang Ge, Xinzheng Huang, and Wangpeng Shi. 2023. "Parasitism Potential and Laboratory Rearing of Psyllaephagus sp., a Newly Discovered Parasitoid of Cacopsylla chinensis" Agronomy 13, no. 3: 943. https://doi.org/10.3390/agronomy13030943