

Anaerobic Soil Disinfestation Promotes Soil Microbial Stability and Antagonistic Bacteria Abundance in Greenhouse Vegetable Production Systems
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Experimental Site
2.2. Experimental Design and Field Management
2.3. Soil Sample Collection, DNA Extraction and High-Throughput Sequencing
2.4. Bioinformatics Analysis
2.5. Statistical Analysis
3. Results
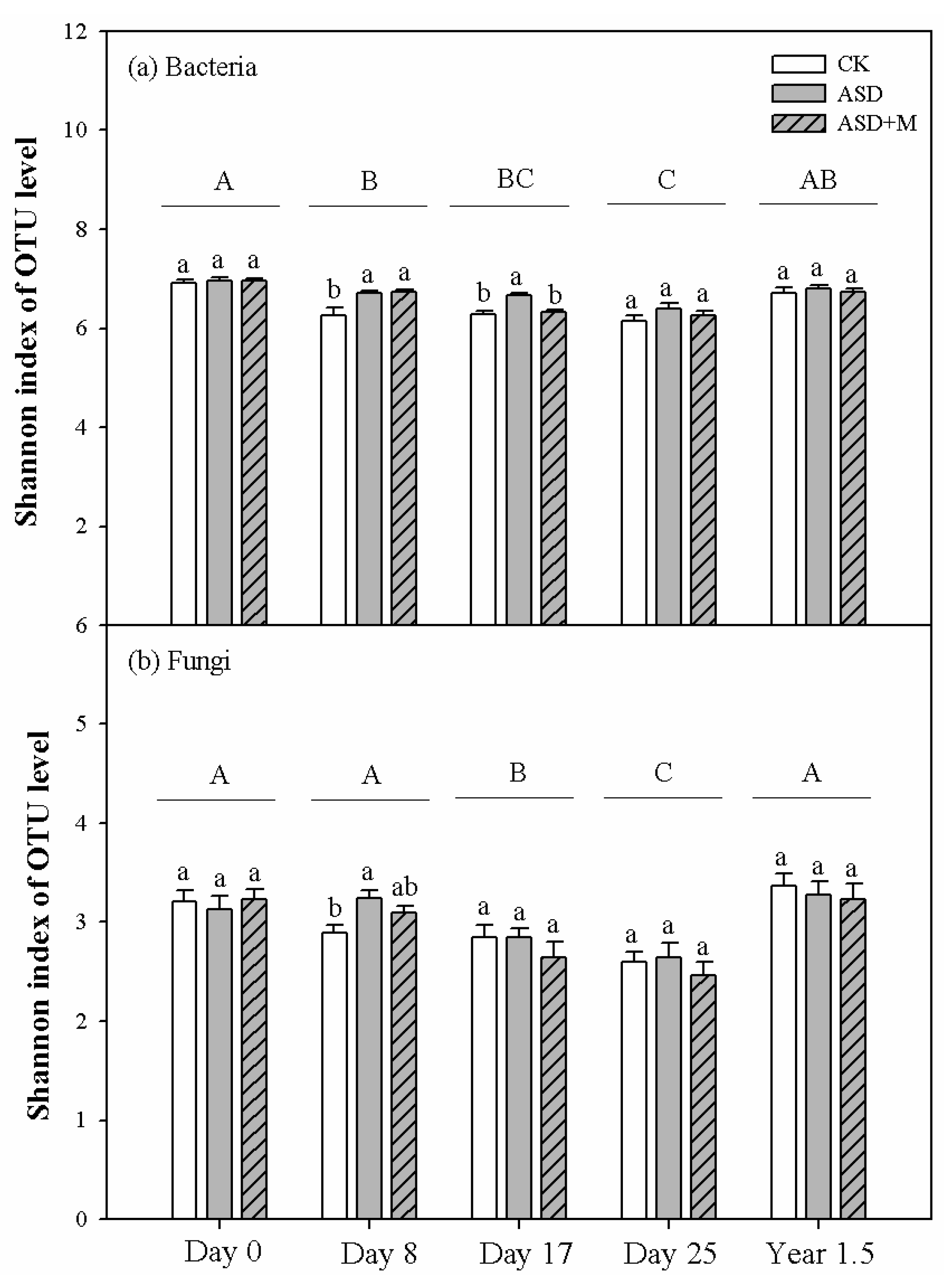
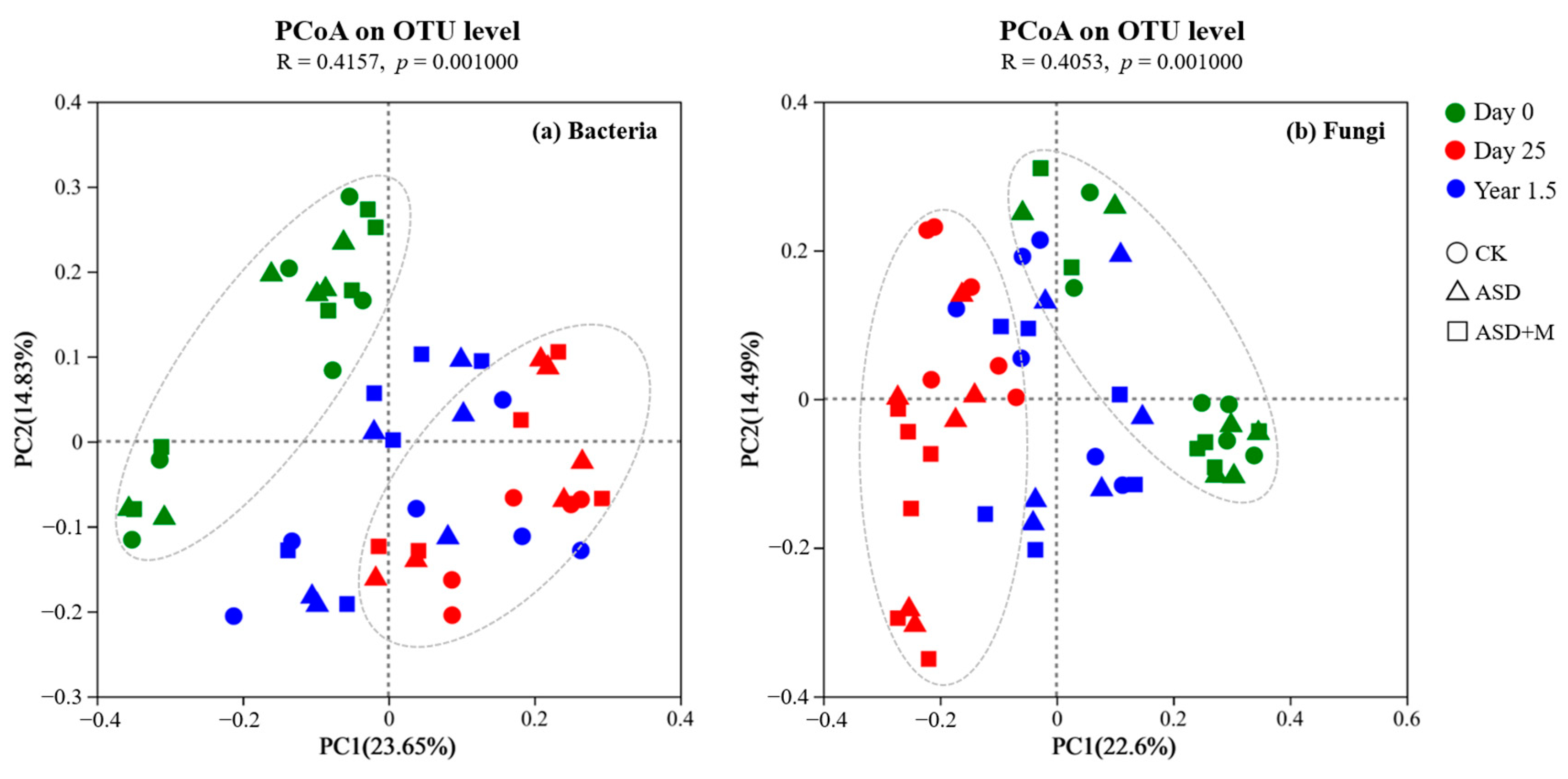
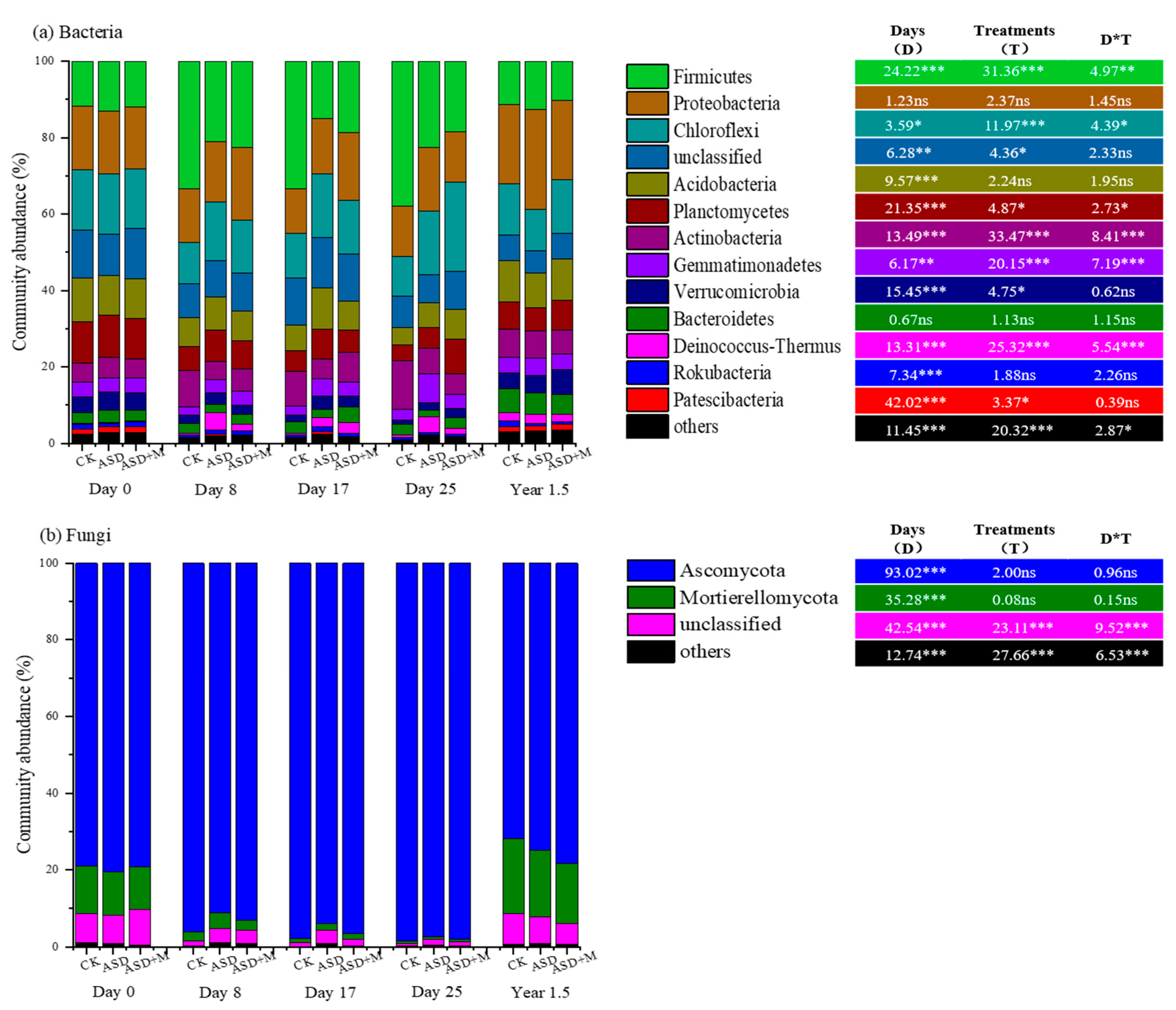
3.1. Soil Microbial Diversity and Composition
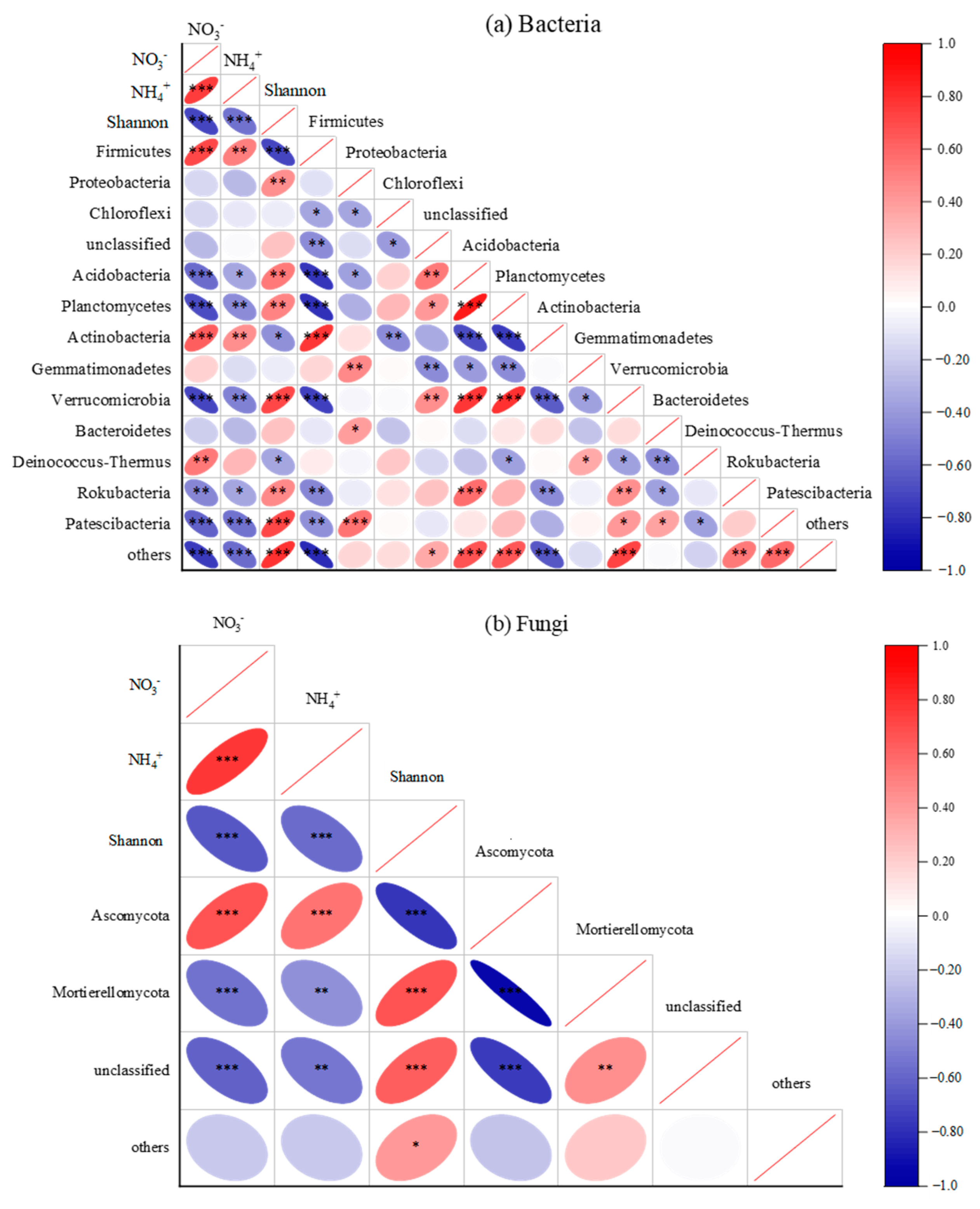
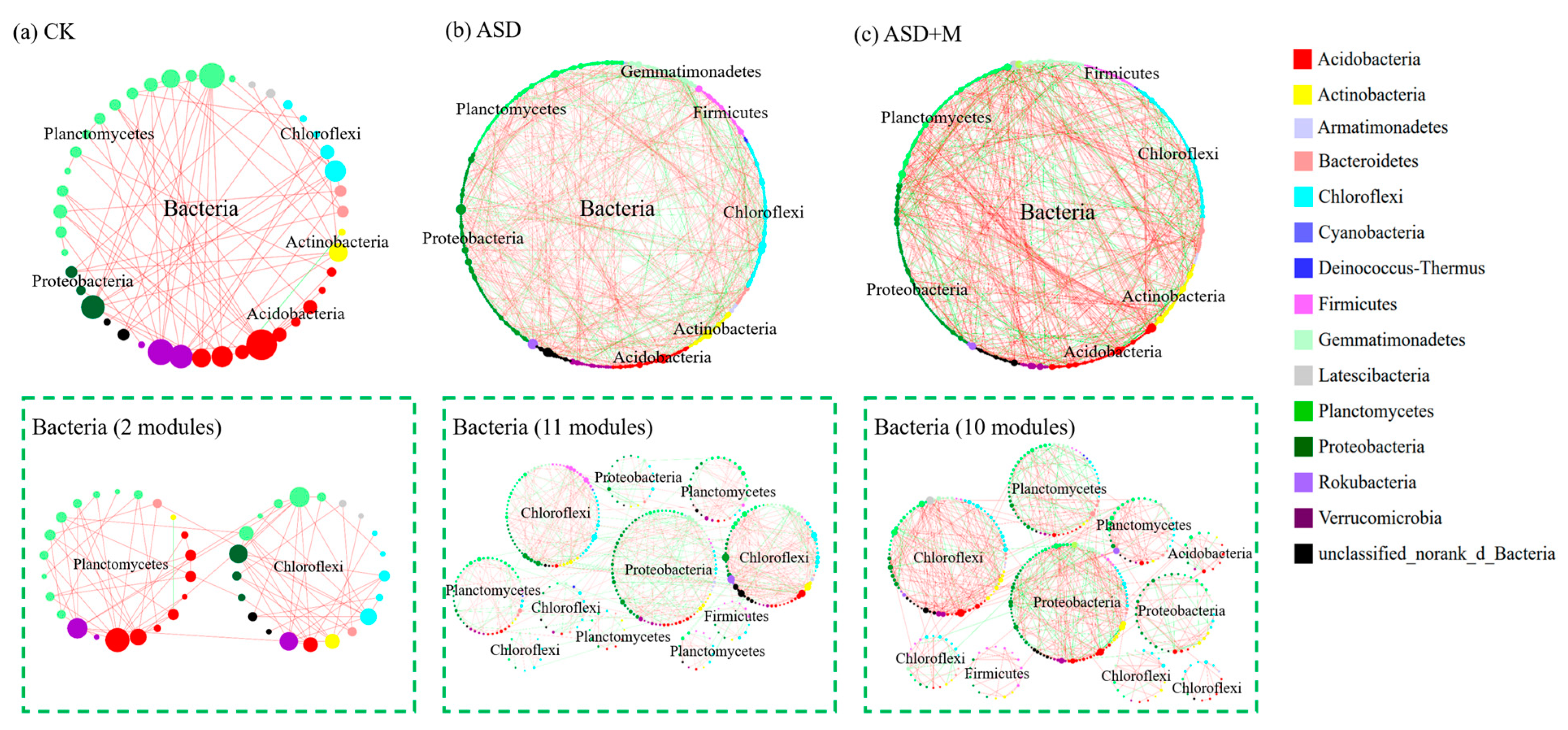
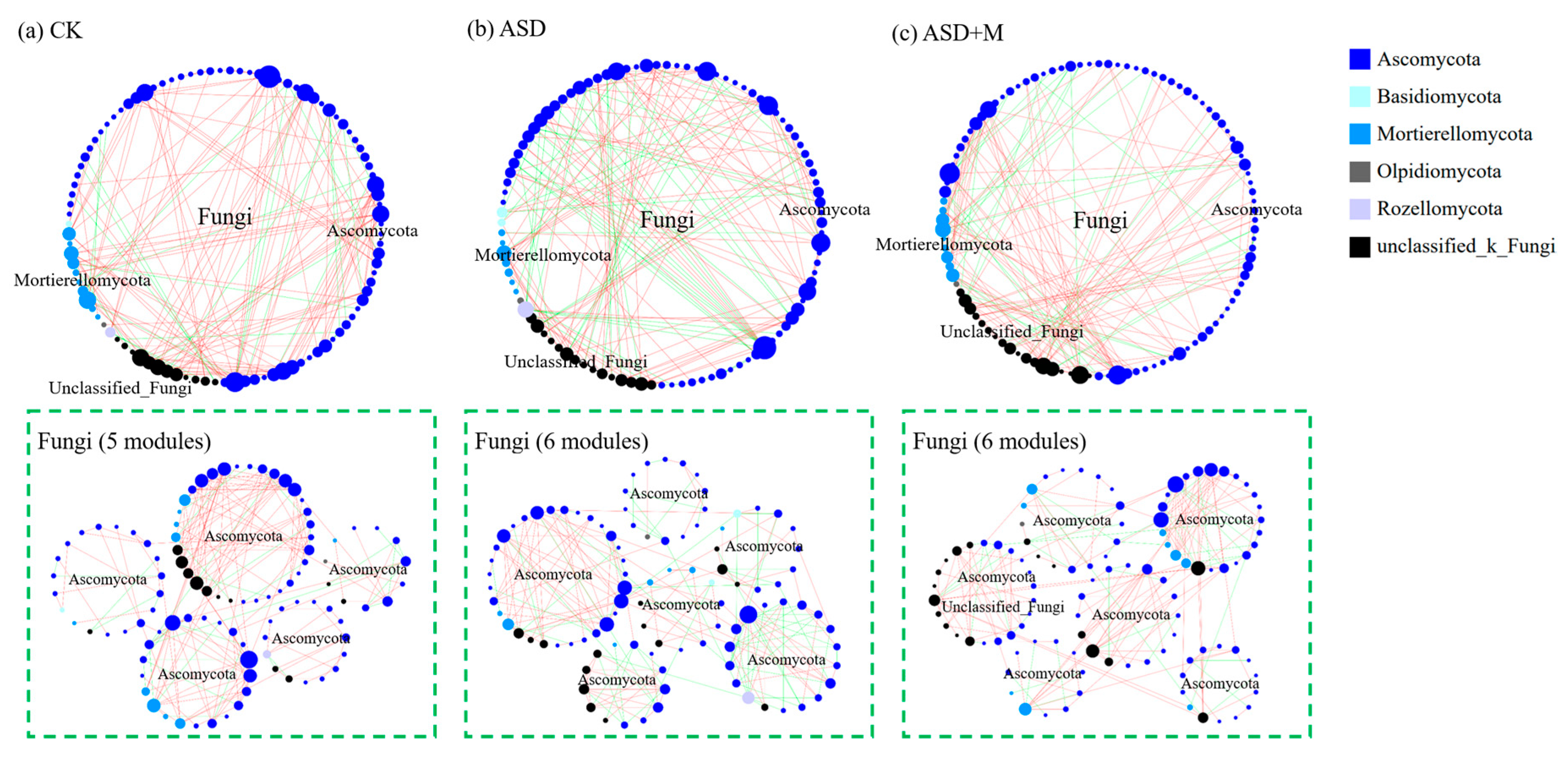
3.2. Soil Microbial Network Complexity
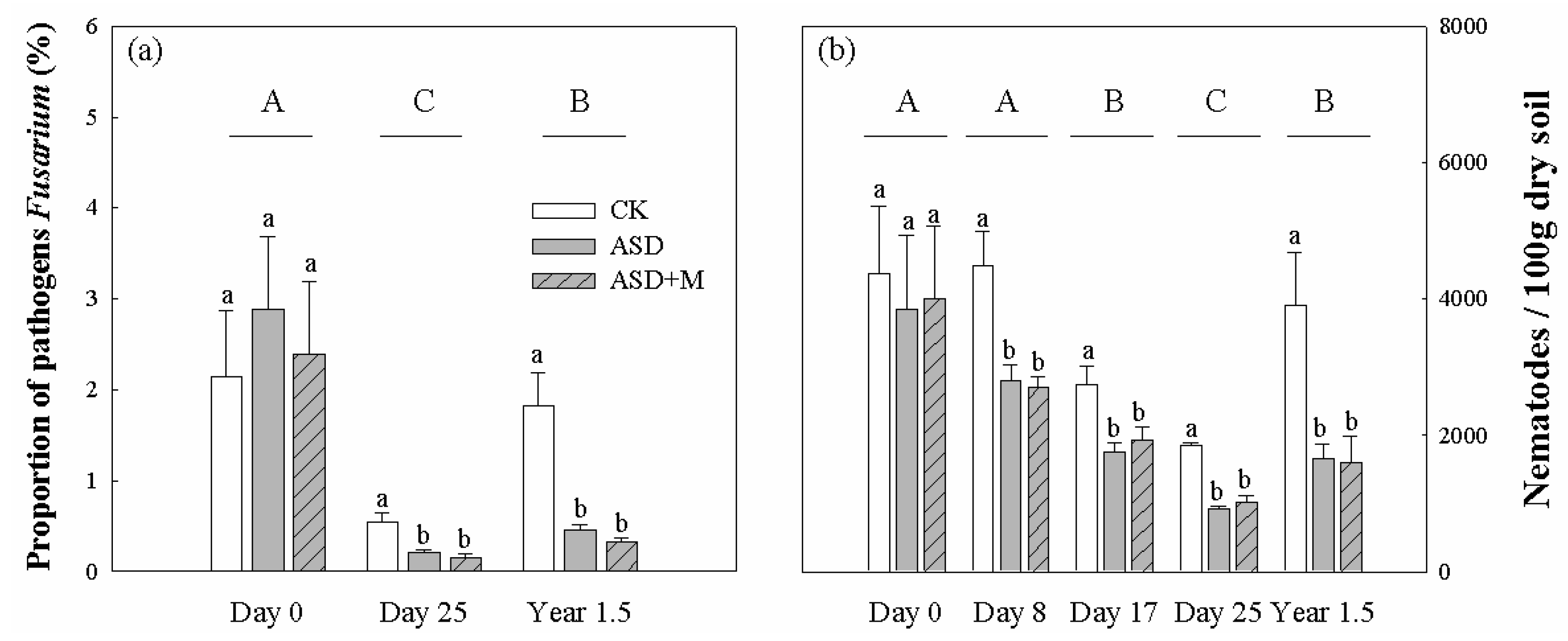
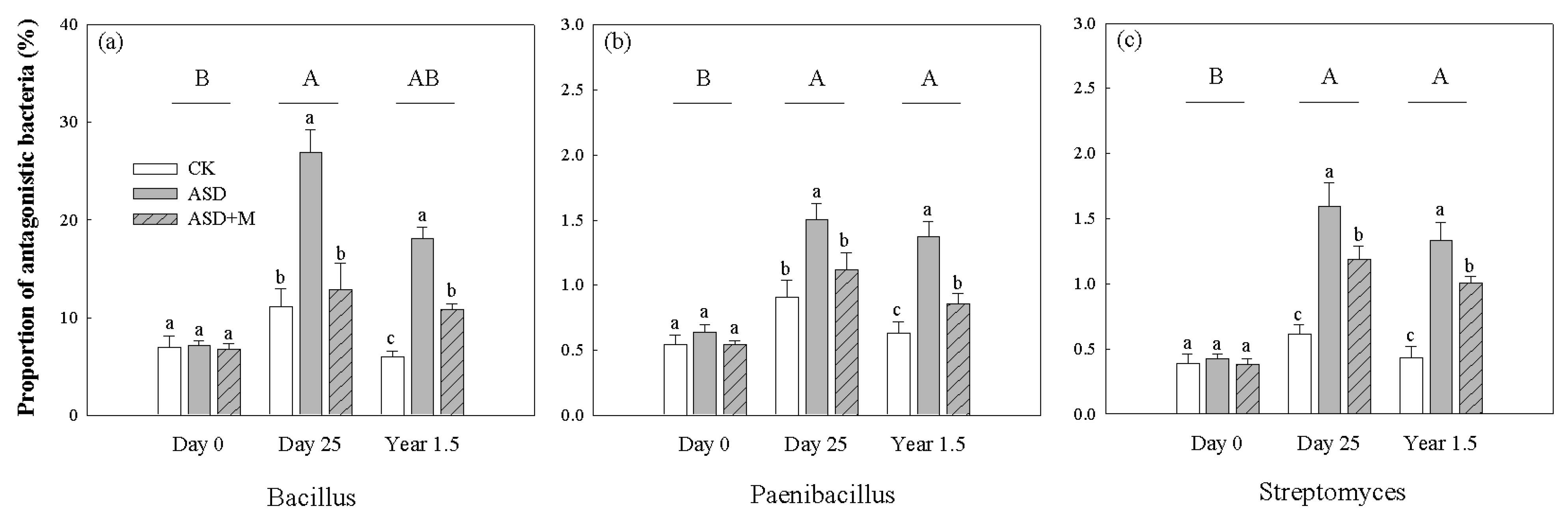
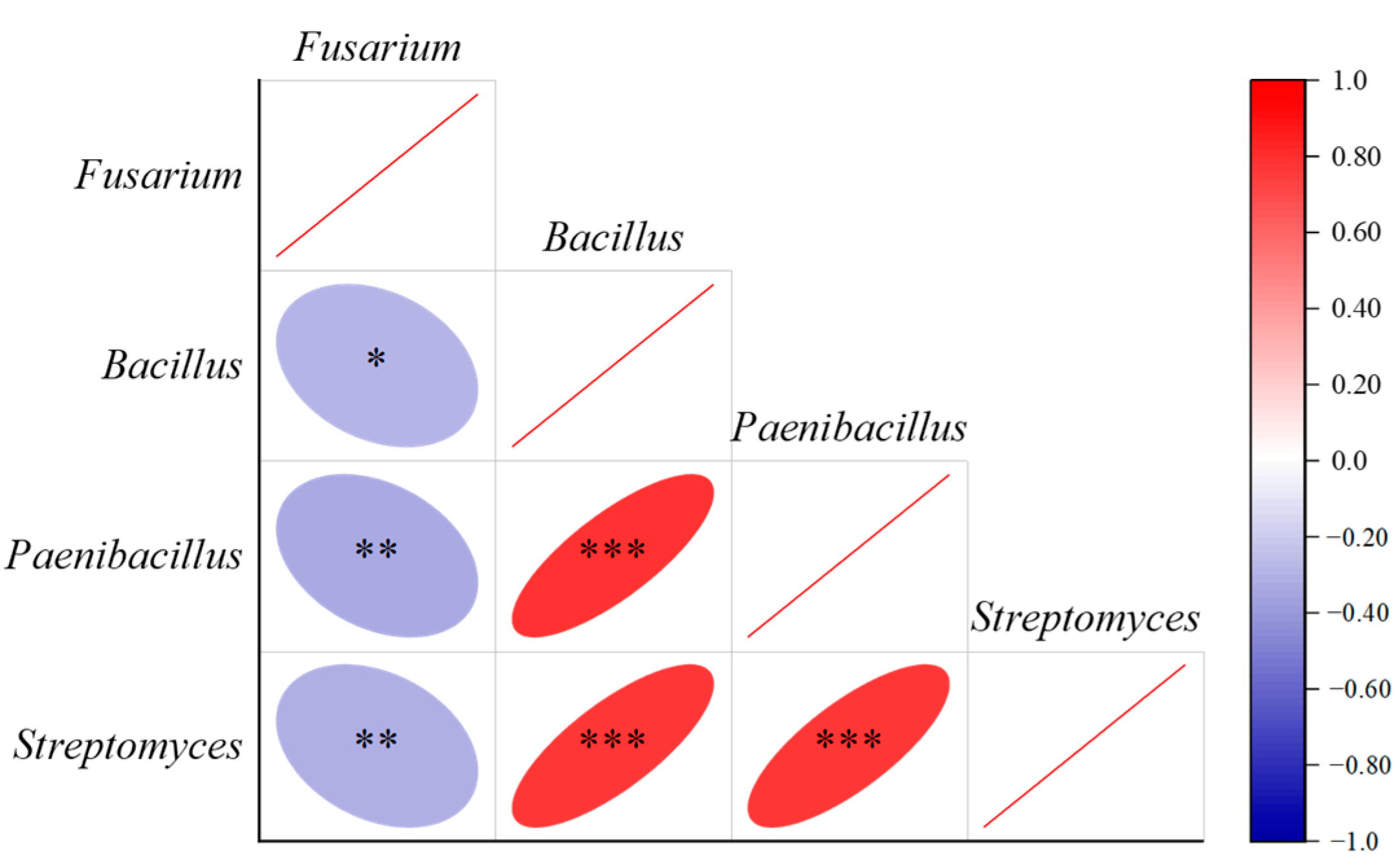
3.3. Plant Pathogens, Soil Nematodes and Antagonistic Bacteria
4. Discussion
4.1. ASD Changed Soil Microbial Community Composition and Increased Bacterial Network Complexity
4.2. Effectively Alleviating Fungal and Nematode Diseases
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, X.X.; Zhao, F.Y.; Zhang, G.X.; Zhang, Y.Y.; Yang, L.J. Vermicompost improves tomato yield and quality and the biochemical properties of soils with different tomato planting history in a greenhouse study. Front. Plant Sci. 2017, 8, 1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qasim, W.; Wan, L.; Lv, H.F.; Zhao, Y.M.; Hu, J.; Meng, F.Q.; Lin, S.; Butterbach-Bahl, K. Impact of anaerobic soil disinfestation on seasonal N2O emissions and N leaching in greenhouse vegetable production system depends on amount and quality of organic matter additions. Sci. Total Environ. 2022, 830, 154673. [Google Scholar] [CrossRef] [PubMed]
- FAO. Food and Agriculture Data. 2020. Available online: http://www.fao.org/faostat/zh/#data (accessed on 22 May 2022).
- Gao, L.H.; Qu, M.; Ren, H.Z.; Sui, X.L.; Chen, Q.Y. Structure, function, application and ecological benefit of single-slope, energy-efficient solar greenhouse in China. Hortic. Technol. 2010, 20, 626–631. [Google Scholar] [CrossRef] [Green Version]
- Fan, Z.B.; Lin, S.; Zhang, X.M.; Jiang, Z.M.; Yang, K.C.; Jian, D.D.; Chen, Y.Z.; Li, J.L.; Chen, Q.; Wang, J.G. Conventional flooding irrigation causes an overuse of nitrogen fertilizer and low nitrogen use efficiency in intensively used solar greenhouse vegetable production. Agric. Water Manag. 2014, 144, 11–19. [Google Scholar] [CrossRef]
- Lv, H.F.; Lin, S.; Wang, Y.F.; Lian, X.J.; Zhao, Y.M.; Li, Y.J.; Du, J.Y.; Wang, Z.X.; Wang, J.G.; Butterbach-Bahl, K. Drip fertigation significantly reduces nitrogen leaching in solar greenhouse vegetable production system. Environ. Pollut. 2019, 245, 694–701. [Google Scholar] [CrossRef]
- Qasim, W.; Xia, L.L.; Lin, S.; Wan, L.; Zhao, Y.M.; Butterbach-Bahl, K. Global greenhouse vegetable production systems are hotspots of soil N2O emissions and nitrogen leaching: A meta-analysis. Environ. Pollut. 2021, 272, 116372. [Google Scholar] [CrossRef]
- Shen, W.; Hu, M.; Qian, D.; Xue, H.; Gao, N.; Lin, X. Microbial deterioration and restoration in greenhouse-based intensive vegetable production systems. Plant Soil 2021, 463, 1–18. [Google Scholar] [CrossRef]
- Zhao, Y.M.; Lv, H.F.; Qasim, W.; Wan, L.; Wang, Y.F.; Lian, X.J.; Liu, Y.N.; Hu, J.; Wang, Z.X.; Li, G.Y.; et al. Drip fertigation with straw incorporation significantly reduces N2O emission and N leaching while maintaining higher vegetable yields in solar greenhouse production. Environ. Pollut. 2021, 273, 116521. [Google Scholar] [CrossRef]
- Lv, H.F.; Zhao, Y.M.; Wang, Y.F.; Wan, L.; Wang, J.G.; Butterbach-Bahl, K.; Lin, S. Conventional flooding irrigation and over fertilization drives soil pH decrease not only in the top- but also in subsoil layers in solar greenhouse vegetable production systems. Geoderma 2020, 363, 114156. [Google Scholar] [CrossRef]
- Zhou, X.G.; Gao, D.M.; Liu, J.; Qiao, P.L.; Zhou, X.L.; Lu, H.B.; Wu, X.; Liu, D.; Jin, X.; Wu, F.Z. Changes in rhizosphere soil microbial communities in a continuously monocropped cucumber (Cucumis sativus L.) system. Eur. J. Soil Biol. 2014, 60, 1–8. [Google Scholar] [CrossRef]
- Lamers, J.G.; Runia, W.T.; Molendijk, L.P.G.; Bleeker, P.O. Perspectives of anaerobic soil disinfestation. Acta Hortic. 2010, 883, 277–284. [Google Scholar] [CrossRef] [Green Version]
- Meng, T.; Zhu, T.; Zhang, J.; Xie, Y.; Sun, W.; Yuan, L.; Cai, Z. Liming accelerates the NO3− removal and reduces N2O emission in degraded vegetable soil treated by reductive soil disinfestation (RSD). J. Soils Sediments 2015, 15, 1968–1976. [Google Scholar] [CrossRef]
- Butler, D.M.; Kokalis-Burelle, N.; Muramoto, J.; Shennan, C.; McCollum, T.G.; Rosskopf, E.N. Impact of anaerobic soil disinfestation combined with soil solarization on plant-parasitic nematodes and introduced inoculum of soil-borne plant pathogens in raised-bed vegetable production. Crop Prot. 2012, 39, 33–40. [Google Scholar] [CrossRef]
- Rosskopf, E.N.; Serrano-Pérez, P.; Hong, J.; Shrestha, U.; del Carmen Rodríguez-Molina, M.; Martin, K.; Butler, D. Anaerobic soil disinfestation and soilborne pest management. In Organic Amendments and Soil Suppressiveness in Plant Disease Management; Springer: Cham, Switzerland, 2015; pp. 277–305. [Google Scholar] [CrossRef]
- Yong, T.; Cui, Y.; Li, H. Rhizospheric soil and root endogenous fungal diversity and composition in response to continuous Panaxnotoginseng cropping practices. Microbiol. Res. 2017, 194, 10–19. [Google Scholar]
- Huang, X.Q.; Zhao, J.; Zhou, X.; Zhang, J.B.; Cai, Z.C. Differential responses of soil bacterial community and functional diversity to reductive soil disinfestation and chemical soil disinfestation. Geoderma 2019, 348, 124–134. [Google Scholar] [CrossRef]
- Liang, B.; Kang, L.; Ren, T.; Junliang, L.; Chen, Q.; Wang, J. The impact of exogenous N supply on soluble organic nitrogen dynamics and nitrogen balance in a greenhouse vegetable system. J. Environ. Manag. 2015, 154, 351–357. [Google Scholar] [CrossRef]
- Bergmann, G.T.; Bates, S.T.; Eilers, K.G.; Lauber, C.L.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. The under-recognized dominance of Verrucomicrobia in soil bacterial communities. Soil Biol. Biochem. 2011, 43, 1450–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, L.B.; Li, Y.; Wang, Z.S.; Yu, Y.; Zhang, N.; Yang, C.; Zeng, Q.C.; Wang, Q. Synthetic community with six Pseudomonas strains screened from garlic rhizosphere microbiome promotes plant growth. Microb. Biotechnol. 2020, 14, 488–502. [Google Scholar] [CrossRef]
- Deng, Y.; Jiang, Y.H.; Yang, Y.F.; He, Z.L.; Luo, F.; Zhou, J.Z. Molecular ecological network analyses. BMC Bioinform. 2012, 13, 113–132. [Google Scholar] [CrossRef] [Green Version]
- Meng, T.Z.; Ren, G.D.; Wang, G.F.; Ma, Y. Impacts on soil microbial characteristics and their restorability with different soil disinfestation approaches in intensively cropped greenhouse soils. Appl. Microbiol. Biotechnol. 2019, 103, 6369–6383. [Google Scholar] [CrossRef]
- Ji, C.Y.; Ye, R.Z.; Yin, Y.F.; Sun, X.F.; Ma, H.L.; Gao, R. Reductive soil disinfestation with biochar amendment modified microbial community composition in soils under plastic greenhouse vegetable production. Soil Tillage Res. 2022, 218, 105323. [Google Scholar] [CrossRef]
- Liu, L.; Chen, S.; Zhao, J.; Zhou, X.; Wang, B.; Li, Y.; Zheng, G.; Zhang, J.; Cai, Z.; Huang, X. Watermelon planting is capable to restructure the soil microbiome that regulated by reductive soil disinfestation. Appl. Soil Ecol. 2018, 129, 52–60. [Google Scholar] [CrossRef]
- Bouskill, N.J.; Lim, H.C.; Borglin, S.; Salve, R.; Wood, T.E.; Silver, W.L.; Brodie, E.L. Pre-exposure to drought increases the resistance of tropical forest soil bacterial communities to extended drought. ISME J. 2013, 7, 384–394. [Google Scholar] [CrossRef]
- Naylor, D.; DeGraaf, S.; Purdom, E.; Coleman-Derr, D. Drought and host selection influence bacterial community dynamics in the grass root microbiome. ISME J. 2017, 11, 2691–2704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeBruyn, L.M.; Nixon, L.T.; Fawaz, M.N.; Johnson, A.M.; Radosevich, M. Global biogeography and quantitative seasonal dynamics of gemmatimonadetes in soil. Appl. Environ. Microbiol. 2011, 77, 6295–6300. [Google Scholar] [CrossRef] [Green Version]
- Wolińska, A.; Kuzniar, A.; Zielenkiewicz, U.; Izak, D.; Szafranek-Nakonieczna, A.; Banach, A.; Błaszczyk, M. Bacteroidetes as a sensitive biological indicator of agricultural soil usage revealed by a culture independent approach. Appl. Soil Ecol. 2017, 119, 128–137. [Google Scholar] [CrossRef]
- Galperin, M.Y. Genome diversity of spore-forming firmicutes. Microbiol. Spectr. 2013, 1, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannula, S.E.; Boschker, H.T.S.; Boer, W.D.; Veen, J.A.V. 13C pulse-labeling assessment of the community structure of active fungi in the rhizosphere of a genetically starch-modified potato (Solanum tuberosum) cultivar and its parental isoline. New Phytol. 2012, 194, 784–799. [Google Scholar] [CrossRef]
- Bastida, F.; Hernández, T.; Albaladejo, J.; García, C. Phylogenetic and functional changes in the microbial community of long-term restored soils under semiarid climate. Soil Biol. Biochem. 2013, 65, 12–21. [Google Scholar] [CrossRef]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; Bodegom, P.M. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef] [Green Version]
- Araki, H.; Hamada, A.; Hossain, M.A.; Takahashi, T. Waterlogging at jointing and/or after anthesis in wheat induces early leaf senescence and impairs grain filling. Field Crops Res. 2012, 137, 27–36. [Google Scholar] [CrossRef]
- Peralta, A.L.; Ludmer, S.; Matthews, J.W. Bacterial community response to changes in soil redox potential along a moisture gradient in restored wetlands. Ecol. Eng. 2014, 73, 246–253. [Google Scholar] [CrossRef]
- Li, L.J.; Zhu-Barker, X.; Ye, R.Z.; Doane, T.A.; Horwath, W.R. Soil microbial biomass size and soil carbon influence the priming effect from carbon inputs depending on nitrogen availability. Soil Biol. Biochem. 2018, 119, 41–49. [Google Scholar] [CrossRef]
- Banerjee, S.; Walder, F.; Büchi, L.; Meyer, M.; Held, A.Y.; Gattinger, A.; Keller, T.; Charles, R.; Van Der Heijden, M.G.A. Agricultural intensification reduces microbial network complexity and the abundance of keystone taxa in roots. ISME J. 2019, 13, 1722–1736. [Google Scholar] [CrossRef] [Green Version]
- Ochoa-Hueso, R.; Collins, S.L.; Delgado-Baquerizo, M.; Hamonts, K.; Pockman, W.T.; Sinsabaugh, R.L.; Smith, M.D.; Knapp, A.K.; Power, S.A. Drought consistently alters the composition of soil fungal and bacterial communities in grasslands from two continents. Glob. Chang. Biol. 2018, 24, 2818–2827. [Google Scholar] [CrossRef] [PubMed]
- Momma, N.; Kobara, Y.; Uematsu, S.; Kita, N.; Shinmura, A. Development of biological soil disinfestations in Japan. Appl. Microbiol. Biotechnol. 2013, 97, 3801–3809. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Zhao, H.H.; Li, M.L.; Zhang, J.B.; Zhang, L. Effects of strong reductive approach on root-knot nematodes, the growth of tomato and soil physicochemical properties. Chin. J. Ecol. 2018, 37, 6. (In Chinese) [Google Scholar]
- Momma, N. Biological soil disinfestation (BSD) of soilborne pathogens and its possible mechanisms. Jpn. Agric. Res. Q. 2008, 42, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Ebihara, Y.; Uematsu, S. Survival of strawberry-pathogenic fungi Fusarium oxysporum f. sp. fragariae, Phytophthora cactorum and Verticillium dahliae under anaerobic conditions. J. Gen. Plant Pathol. 2014, 80, 50–58. [Google Scholar] [CrossRef]
- Bonanomi, G.; Antignani, V.; Capodilupo, M.; Scala, F. Identifying the characteristics of organic soil amendments that suppress soil-borne plant diseases. Soil Biol. Biochem. 2010, 42, 136–144. [Google Scholar] [CrossRef]
- Kinsella, K.; Schulthessm, C.P. Rapid quantification of Bacillus subtilis antibiotics in the rhizosphere. Soil Biol. Biochem. 2010, 42, 1009–1192. [Google Scholar] [CrossRef]
- Li, B.H.; Li, Q.; Xiong, L.M.; Kronzucker, H.J.; Krämer, U.; Shi, W.M. Arabidopsis Plastid AMOS1/EGY1 Integrates Abscisic Acid Signaling to Regulate Global Gene Expression Response to Ammonium Stress. Plant Physiol. 2012, 160, 2040–2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Shannon Index | Bacterial Community Structure | Fungal Community Structure | ||
|---|---|---|---|---|
| Bacteria | Fungi | |||
| Days (D) | 33.60 *** | 23.79 *** | 10.79 *** | 13.56 *** |
| Treatments (T) | 10.42 *** | 0.87 ns | 4.61 *** | 5.04 *** |
| D × T | 2.45 * | 1.64 ns | 1.76 ns | 1.53 ns |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Wan, L.; Qasim, W.; Lv, H.; Zhao, Y.; Li, G.; Butterbach-Bahl, K.; Lin, S. Anaerobic Soil Disinfestation Promotes Soil Microbial Stability and Antagonistic Bacteria Abundance in Greenhouse Vegetable Production Systems. Agronomy 2023, 13, 939. https://doi.org/10.3390/agronomy13030939
Hu J, Wan L, Qasim W, Lv H, Zhao Y, Li G, Butterbach-Bahl K, Lin S. Anaerobic Soil Disinfestation Promotes Soil Microbial Stability and Antagonistic Bacteria Abundance in Greenhouse Vegetable Production Systems. Agronomy. 2023; 13(3):939. https://doi.org/10.3390/agronomy13030939
Chicago/Turabian StyleHu, Jing, Li Wan, Waqas Qasim, Haofeng Lv, Yiming Zhao, Guoyuan Li, Klaus Butterbach-Bahl, and Shan Lin. 2023. "Anaerobic Soil Disinfestation Promotes Soil Microbial Stability and Antagonistic Bacteria Abundance in Greenhouse Vegetable Production Systems" Agronomy 13, no. 3: 939. https://doi.org/10.3390/agronomy13030939