
Phytolith Content Negatively Affects Forage Quality of Eragrostis curvula (Schrad.) Nees
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Phytolith Extraction and Experimental Design
2.3. Statistical Analysis
2.4. Genome-Wide Analysis to Look for Lsi Genes and Phylogenetic Analysis
2.5. Microscopic Observations
3. Results
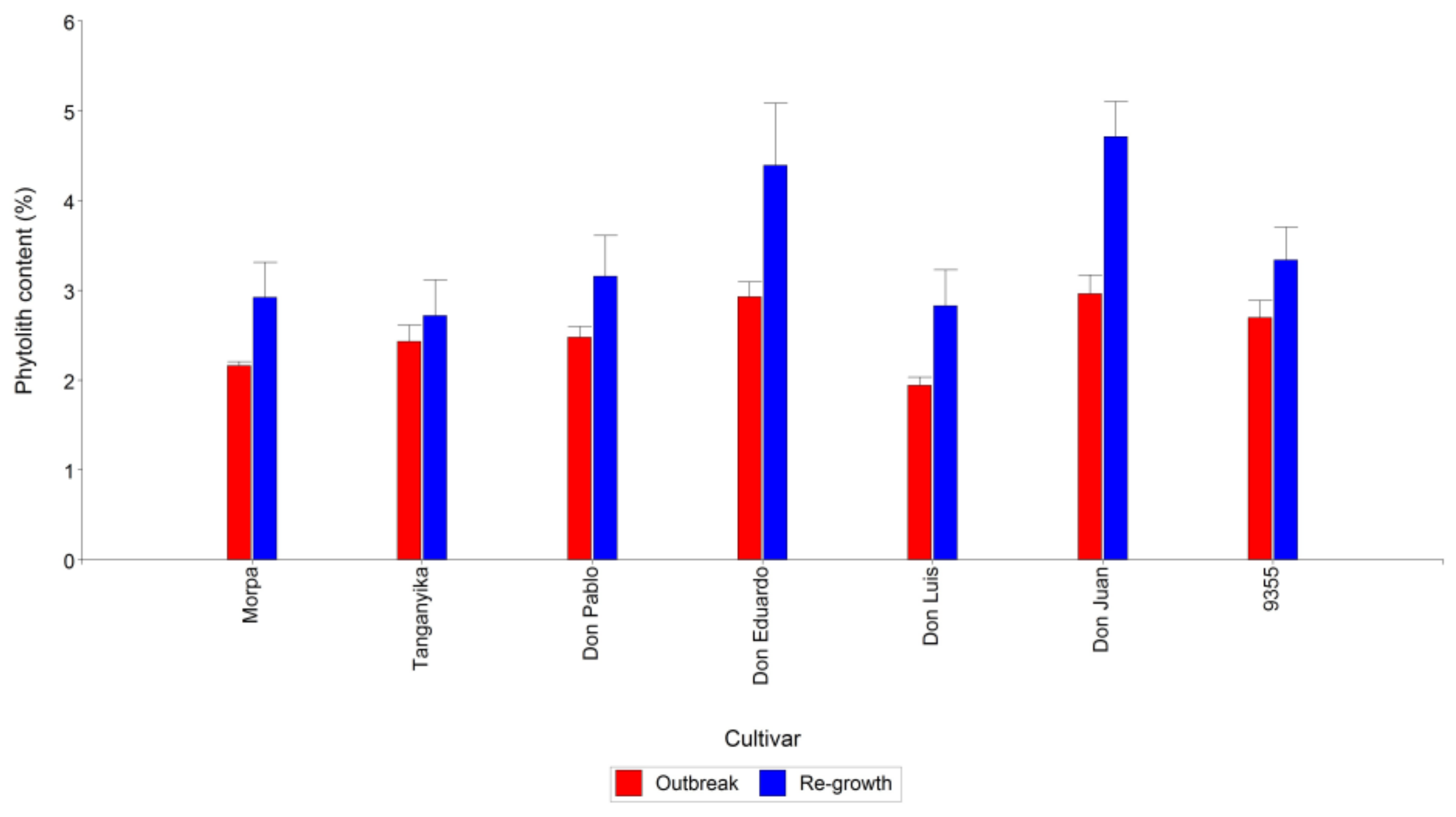
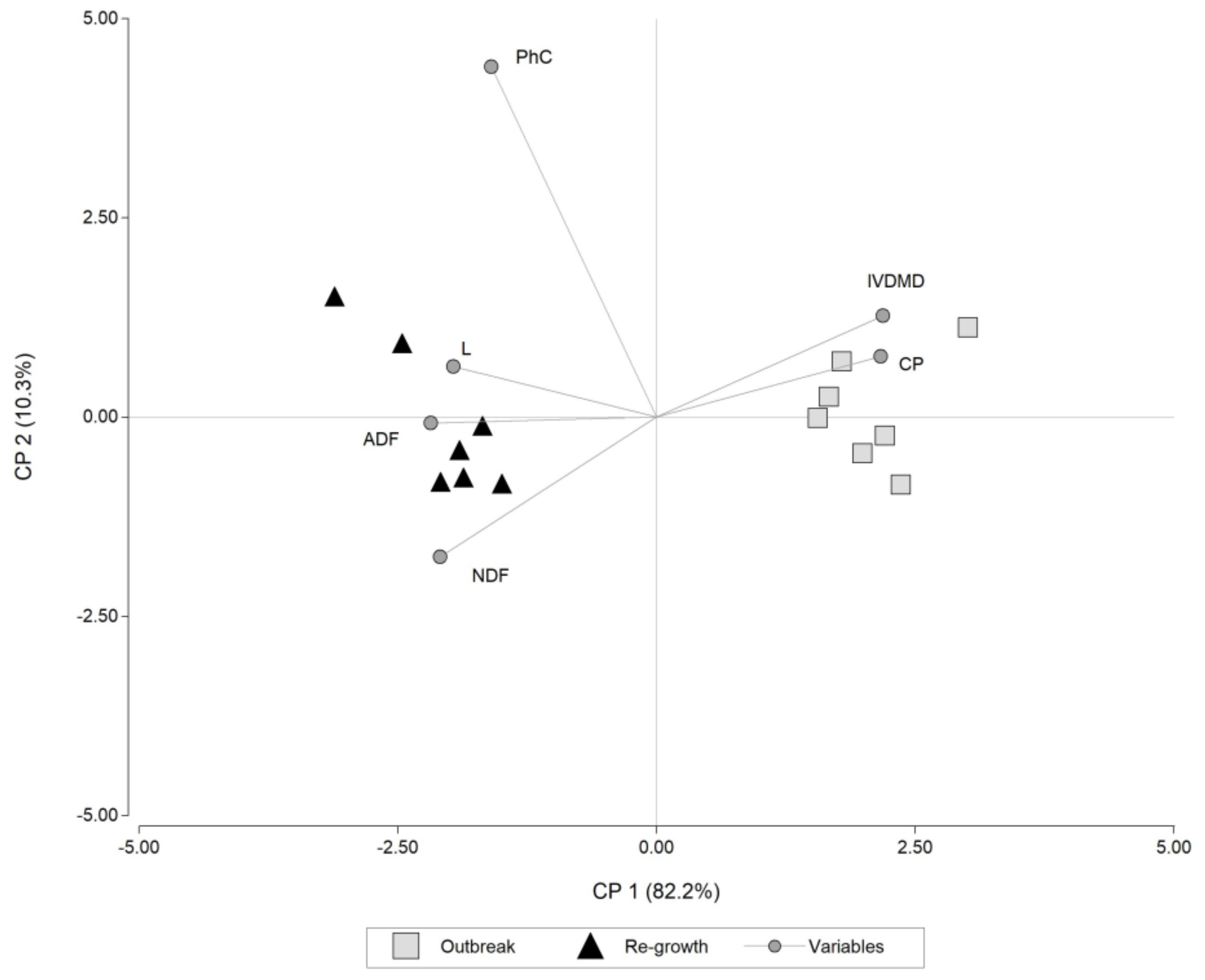
3.1. Phytolith Content and the Interaction between Cultivars and Cutting Time
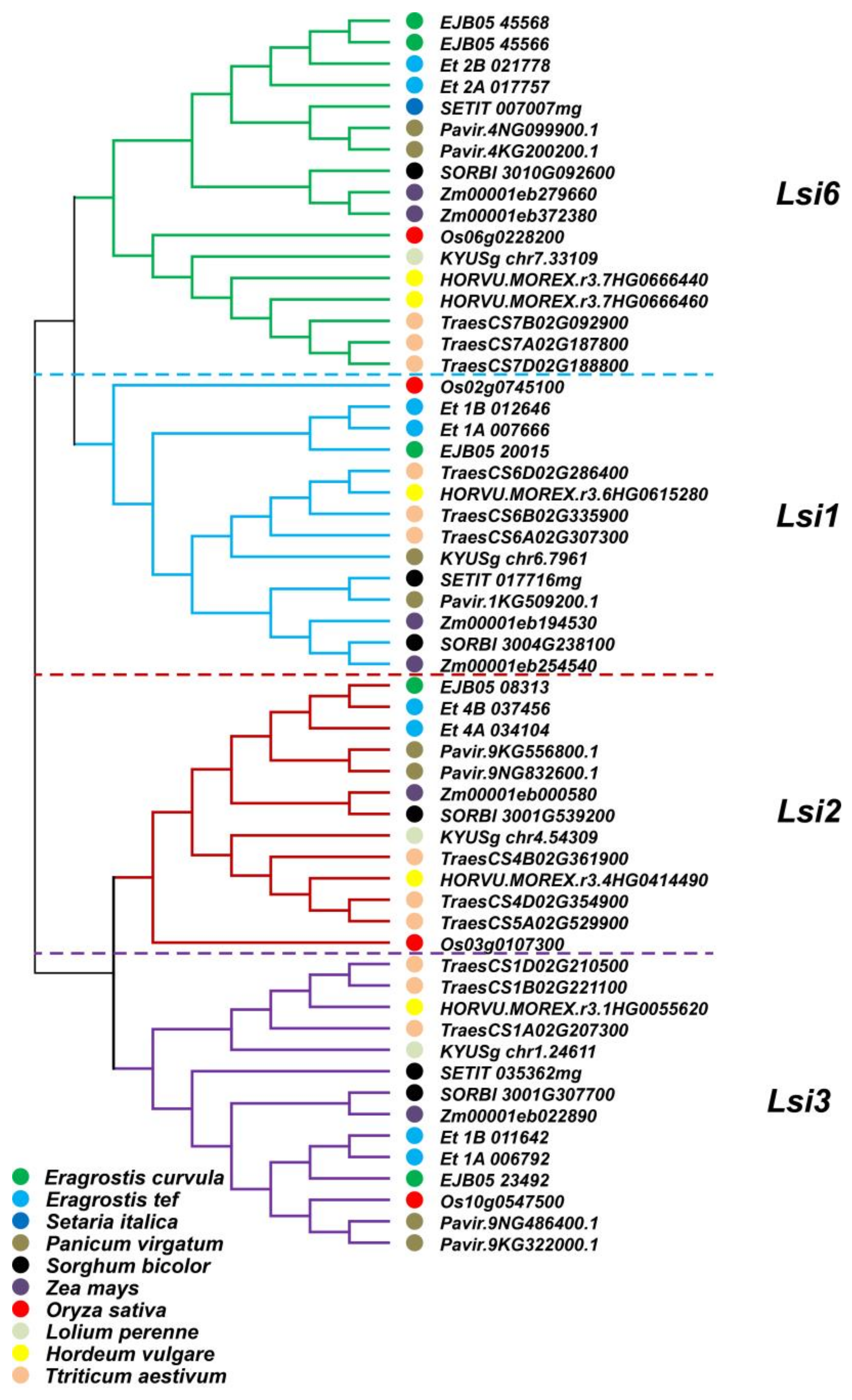
3.2. Genome-Wide Analysis to Look for Lsi Genes and Phylogenetic Analysis
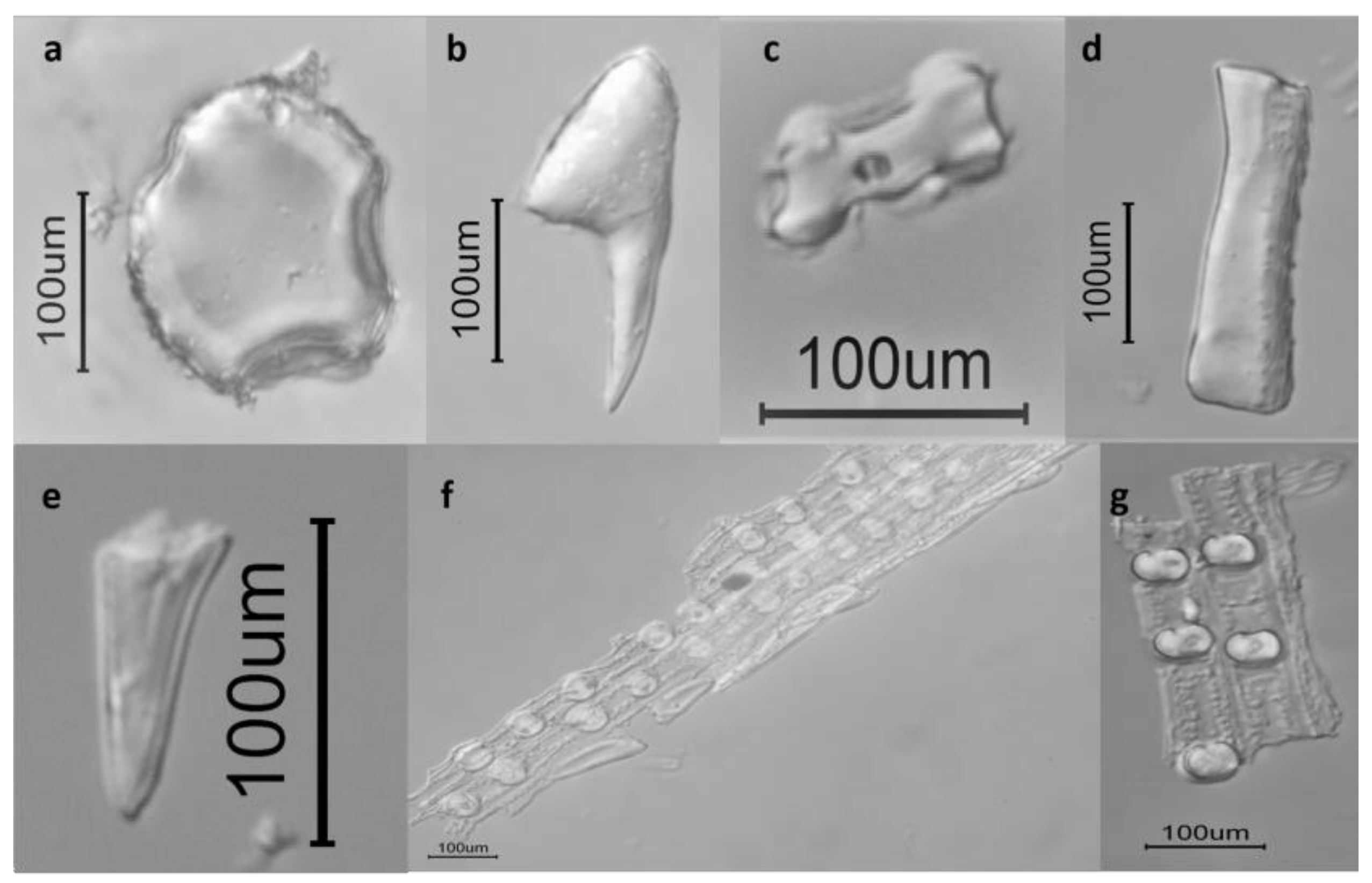
3.3. Microscopic Observations
4. Discussion
5. Conclusions
- (1)
- This is the first study on the phytolith content and the relationship with forage quality parameters in different genotypes and phenological stages in Eragrostis curvula.
- (2)
- The content of phytoliths in E. curvula, expressed as a percentage of dry matter, is similar to the one usually shown by other forage grasses from semi-arid regions.
- (3)
- Based on the criterion that lower phytolith content implies better palatability and digestibility, the best cultivars analyzed here were Don Luis, Morpa, and Tanganyika. Additionally, those with the lowest forage value were Don Juan and Don Eduardo.
- (4)
- The low forage quality of weeping lovegrass could be attributed, in part, to the content of phytoliths, particularly at the re-growth stage.
- (5)
- The variation in the degree of silicification observed in the analyzed cultivars was independent of the plant phenological stage.
- (6)
- The sequence of Lsi genes involved in Si uptake and transport was conserved across the grass family.
- (7)
- The results obtained in this work constitute a valuable source for breeding programs to select genotypes based on the phytolith content to improve the E. curvula forage quality.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nawaz, M.A.; Zakharenko, A.M.; Zemchenko, I.V.; Haider, M.S.; Ali, M.A.; Imtiaz, M.; Chung, G.; Tsatsakis, A.; Sun, S.; Golokhvast, K.S. Phytolith Formation in Plants: From Soil to Cell. Plants 2019, 8, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, O.; Puppe, D.; Kaczorek, D.; Prakash, N.B.; Schaller, J. Silicon in the Soil–Plant Continuum: Intricate Feedback Mechanisms within Ecosystems. Plants 2021, 10, 652. [Google Scholar] [CrossRef] [PubMed]
- Bowdery, D. Phytolith Analysis Applied to Pleistocene-Holocene Archaeological Sites in the Australian Arid Zone; British Archaeological Reports Ltd.: Oxford, UK, 1998; p. 216. [Google Scholar]
- Feng, J.; Yamaji, N.; Mitani-Ueno, N. Transport of silicon from roots to panicles in plants. Proc. Jpn. Acad. Ser. B 2011, 87, 377–385. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, N.; Mitatni, N.; Ma, J.F. A transporter regulating silicon distribution in rice shoots. Plant Cell 2008, 20, 1381–1389. [Google Scholar] [CrossRef] [Green Version]
- Yamaji, N.; Sakurai, G.; Mitani-Ueno, N.; Ma, J.F. Orchestration of three transporters and distinct vascular structures in node for intervascular transfer of silicon in rice. Proc. Natl. Acad. Sci. USA 2015, 112, 11401–11406. [Google Scholar] [CrossRef] [Green Version]
- Sendulsky, T.; Labouriau, L.G. Corpos silicosos de gramíneas dos Cerrados. I. AABC 1966, 38, 159–185. [Google Scholar]
- Zucol, A.F. Fitolitos. III. Asociaciones fitolíticas de Piptochaetium montevidense (Stipeae: Arundinoideae: Poaceae). Una nueva metodología descriptiva. Bol. Soc. Argent. Bot. 2001, 36, 69–85. [Google Scholar]
- Zurro, D. El análisis de fitolitos y su papel en el estudio del consumo de recursos vegetales en la prehistoria: Bases para una propuesta metodológica materialista. Trab. Prehist. 2006, 63, 35–54. [Google Scholar] [CrossRef] [Green Version]
- Carnelli, A.L.; Madella, M.; Theurillat, J.P.; Ammann, B. Aluminum in the opal silica reticule of phytoliths: A new tool in palaeoecological studies. Am. J. Bot. 2002, 89, 346–351. [Google Scholar] [CrossRef]
- Epstein, E. The anomaly of silicon in plant biology. Proc. Natl. Acad. Sci. USA 1994, 91, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Piperno, D.R. Phytolyth Analysis: An Archaeological and Geological Perspective; Elsevier: Amsterdam, The Netherlands, 2014; Volume 22, pp. 119–120. [Google Scholar]
- Salvant, N.; Snyder, G.; Danoff, L. Silicon management and sustentable rice production. Adv. Agron. 1997, 58, 151–199. [Google Scholar]
- Salem, E.M.M.; Kenawey, M.K.M.; Saudy, H.S.; Mubarak, M. Influence of Silicon Forms on Nutrients Accumulation and Grain Yield of Wheat Under Water Deficit. Gesunde Pflanz. 2022, 74, 539–548. [Google Scholar] [CrossRef]
- Saudy, H.S.; Salem, E.M.M.; El-Momen, W.R. A Effect of Potassium Silicate and Irrigation on Grain Nutrient Uptake and Water Use Efficiency of Wheat Under Calcareous Soils. Gesunde Pflanz. 2022. [Google Scholar] [CrossRef]
- Borda, O.A.; Barón, F.H.; Gómez, M.I. Silicon as a beneficial element in forage oat (Avena sativa L.): Physiological responses of growth and management. Agron. Colomb. 2007, 25, 273–279. [Google Scholar]
- Sánchez, P. Properties and Management of Soils in the Tropics, 1st ed.; Cruz, M., Escoto, J.B., Eds.; IICA Library Venezuela: San José, Costa Rica, 1981; p. 235. [Google Scholar]
- Guntzer, F.; Keller, C.; Meunier, J.D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef] [Green Version]
- Massey, F.P.; Ennos, A.R.; Hartley, S.E. Grasses and the resource availability hypothesis: The importance of silica-based defences. J. Ecol. 2007, 95, 414–424. [Google Scholar] [CrossRef] [Green Version]
- Bon, M.P.; Inga, K.G.; Utsi, T.A.; Jónsdóttir, I.S.; Bråthen, K.A. Forage quality in tundra grasslands under herbivory: Silicon based defences, nutrients and their ratios in grasses. J. Ecol. 2021, 110, 129–143. [Google Scholar] [CrossRef]
- Massey, F.P.; Ennos, A.R.; Hartley, S.E. Silica in grasses as a defence against insect herbivores: Contrasting effects on folivores and phloem feeders. J. Ecol. 2006, 75, 595–603. [Google Scholar] [CrossRef]
- Massey, F.P.; Hartley, S.E. Experimental demonstration of the antiherbivore effects of silica in grasses: Impacts on foliage digestibility and vole growth rates. Proc. Biol. Sci. 2006, 273, 2299–2304. [Google Scholar] [CrossRef] [Green Version]
- Hunt, J.W.; Dean, A.P.; Webster, R.E.; Jonhson, G.N.; Ennos, A.R. A Novel Mechanism by which Silica Defends Grasses Against Herbivory. Ann. Bot. 2008, 102, 653–656. [Google Scholar] [CrossRef] [Green Version]
- Van der Vorm, P.D.J. Dry ashing of plant material and dissolution of the ash in HF for the colorimetric determination of silicon. Commun. Soil Sci. Plant Anal. 1987, 18, 1181–1189. [Google Scholar] [CrossRef]
- Taber, H.G.; Shogren, D.; Gang, L. Extraction of silicon from plant tissue with dilute HCl and HF and measurement by modified inductive coupled argon plasma procedures. Commun. Soil Sci. Plant Anal. 2002, 33, 1661–1670. [Google Scholar] [CrossRef]
- Jones, R.L.; Dreher, G.B. Silicon. In Methods of Soil Analysis. Part 3. Chemical Methods; Sparks, D.L., Ed.; SSSA Book Series 5; Soil Science Society of America and American Society of Agronomy: Madison, WI, USA, 1996; pp. 627–637. [Google Scholar]
- Rovner, I. Potential of opal phytoliths for use in palaeoecological reconstruction. Quat. Res. 1971, 3, 345–359. [Google Scholar]
- Rovner, I.; Russ, J.C. Darwin and design in phytolith systematics: Morphometric methods for mitigating redundancy. In Phytolith Systematics; Rapp, G., Mulholland, S.C., Eds.; Plenum Press: New York, NY, USA, 1992; pp. 253–276. [Google Scholar]
- Zucol, A.F.; Brea, M. Phytolith systematics, guideline for a classificatory system. A study case in the Alvear formation (lower Pleistocene), Entre Ríos, Argentine. Ameghiniana 2005, 42, 685–704. [Google Scholar]
- Mandela, M.; Alexandre, A.; Ball, T. International Code for Phytolith Nomenclature 1.0 (ICPN). Ann. Bot. 2005, 96, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Luciani, G.F.; Sobanski, M.G.; Meier, M.S.; Polci, P.A.; Miranda, R.; Echenique, C.V. Weeping lovegrass yield and nutritional quality provides an alternative to beef cattle feeding in semiarid environments of Argentina. Crop Sci. 2012, 52, 1955–1965. [Google Scholar] [CrossRef]
- Leigh, J.H. The relative paptability of various varieties of weeping lovegrass (Eragrostis curvula (Schrad) Nees). Grass Forage Sci. 1961, 16, 135–140. [Google Scholar] [CrossRef]
- Wilson, J.; Minson, D. Comparative digestibility of tropical and temperate forage—A contrast between grasses and legumes. J. Aust. Inst. Agric. Sci. 1980, 46, 247–249. [Google Scholar]
- Wilson, J. Cell wall characteristics in relation to forage digestion by ruminants. J. Agric. Sci. 1994, 122, 173–182. [Google Scholar] [CrossRef]
- Laborde, H. Quality and nutritional value. In Weeping Grass, Its Biology and Management; Fernández, O., Brevedad, R., Gargajo, A., Eds.; CERZOS and Department of Agronomy University Nacional del Sur: Bahía Blanca, Argentina, 1991; pp. 325–347. [Google Scholar]
- Soil Survey Staff. Soil Taxonomy: A Basic System of Soil Classification for Making and Interpreting Soil Surveys, 2nd ed.; Natural Resources Conservation Service, US Government Print Office: Washington, DC, USA, 1999; p. 786. [Google Scholar]
- De Campos, A.C.; Labouriau, L.G. Corpos silicosos de gramíneas dos Cerrados. II. Pesqui. Agropecu. Bras. 1969, 4, 143–151. [Google Scholar]
- Da Silva, S.T.; Labouriau, L.G. Corpos silicosos de gramíneas dos Cerrados. III. Pesqui. Agropecu. Bras. 1970, 5, 167–182. [Google Scholar]
- Labouriau, L.G. Phytolith work in Brazil: A mini-review. Phytolith. Newsl. 1983, 2, 6–11. [Google Scholar]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat, Versión 2010; Grupo InfoStat. 2010. Available online: http://www.infostat.com.ar.
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Carballo, J.; Santos, B.A.; Zappacosta, D.; Garbus, I.; Selva, J.P.; Gallo, C.A.; Díaz, A.; Albertini, E.; Cáccamo, M.; Echenique, V. A high-quality genome of Eragrostis curvula grass provides insights into Poáceae evolution and supports new strategies to enhance forage quality. Sci. Rep. 2019, 9, 10250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanBuren, R.; Man Wai, C.; Wang, X.; Pardo, J.; Yocca, A.E.; Wang, H.; Chaluvadi, S.R.; Han, G.; Bryant, D.; Edger, P.P.; et al. Exceptional subgenome stability and functional divergence in the allotetraploid Ethiopian cereal teff. Nat. Commun. 2020, 11, 884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.J.; Ning, Y.Q.; Zhang, S.W.; Chen, Q.; Shao, C.R.; Guo, Y.W.; Zhou, J.X.; Li, L.; Chen, S.; He, X.J. IDN2 and its paralogs form a complex required for RNA-directed DNA methylation. PLoS Genet. 2012, 8, e1002693. [Google Scholar] [CrossRef]
- Jiao, Y.; Peluso, P.; Shi, J.; Liang, T.; Stitzer, M.C.; Wang, B.; Campbell, M.S.; Stein, J.C.; Wei, X.; Chin, C.S.; et al. Improved maize reference genome with single-molecule technologies. Nature 2017, 546, 524–527. [Google Scholar] [CrossRef]
- McCormick, R.F.; Truong, S.K.; Sreedasyam, A.; Jenkins, J.; Shu, S.; Sims, D.; Kennedy, M.; Amirebrahimi, M.; Weers, B.D.; McKinley, B.; et al. The Sorghum bicolor reference genome: Improved assembly, gene annotations, a transcriptome atlas, and signatures of genome organization. Plant. J. 2018, 93, 338–354. [Google Scholar] [CrossRef] [Green Version]
- International Wheat Genome Sequencing Consortium (IWGSC); Kanyuka, K.; King, R. Shifting the Limits in Wheat Research and Breeding Using a Fully Annotated Reference Genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [Green Version]
- Mascher, M.; Wicker, T.; Jenkins, J.; Plott, C.; Lux, T.; Koh, C.S.; Ens, J.; Gundlach, H.; Boston, L.B.; Tulpová, Z.; et al. Long-read sequence assembly: A technical evaluation in barley. Plant Cell 2021, 33, 1888–1906. [Google Scholar] [CrossRef]
- Frei, D.; Veekman, E.; Grogg, D.; Stoffel-Studer, I.; Morishima, A.; Shimizu-Inatsugi, R.; Yates, S.; Shimizu, K.K.; Frey, J.E.; Studer, B.; et al. Ultralong Oxford Nanopore reads enable the development of a reference-grade perennial ryegrass genome assembly. Genome Biol. Evol. 2021, 13, evab159. [Google Scholar] [CrossRef] [PubMed]
- Lovell, J.T.; MacQueen, A.H.; Mamidi, S.; Bonnette, J.; Jenkins, J.; Napier, J.D.; Sreedasyam, A.; Healey, A.; Session, A.; Shu, S.; et al. Genomic mechanisms of climate adaptation in polyploid bioenergy switchgrass. Nature 2021, 590, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Gargano, A.O.; Aduriz, M.A.; Arelovich, H.M.; Amela, M.I. Forage yield and nutritive value of Eragrostis curvula and Digitaria eriantha in central-south semi-arid Argentina. Tropl. Grassl. 2001, 35, 161–167. [Google Scholar]
- Rovner, I. Macro and Micro ecological Reconstruction Using Plant Opal Phytolith Analysis. Geoarchaeology 1988, 3, 155–163. [Google Scholar] [CrossRef]
- Van Soest, P.; Jones, L. Effect of silica in forage upon digestibility. J. Dairy Sci. 1968, 51, 1644–1648. [Google Scholar] [CrossRef]
- Hoover, W. Chemical factors involved in ruminal fiber digestion. J. Dairy Sci 1986, 69, 2755–2766. [Google Scholar] [CrossRef]
- Neethirajan, S.; Gordon, R.; Wang, L. Potential of silica bodies (phytoliths) for nanotechnology. Trends Biotechnol. 2009, 27, 461–467. [Google Scholar] [CrossRef]
- Elger, A.; Lemoine, D.G.; Fenner, M.; Hanley, M.E. Plant ontogeny and chemical defense: Older seedlings are better defended. Oikos 2009, 118, 767–773. [Google Scholar] [CrossRef]
- Jones, L.; Milne, A.; Sanders, J. Tabashir: An opal of plant origin. Science 1966, 151, 464–466. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.S. Relationship between fermentation acid production in the rumen and the requirement for physically effective fiber. J. Dairy Sci. 1997, 80, 1447–1462. [Google Scholar] [CrossRef] [PubMed]
- Parra-Terraza, S.; Baca-Castillo, G.A.; Carrillo-González, R.; Kohashi-Shibata, J.; Martínez-Garza, A.; Trejo-López, C. Comparison of three methods of analysis of silicon in cucumber leaf tissue. TERRA Latinoam. 2004, 22, 401–407. [Google Scholar]
- Kilmer, V.J. Silicon. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties; Black, C.A., Ed.; American Society of Agronomy: Madison, WI, USA, 1965; pp. 959–962. [Google Scholar]
- Jones, L.H.; Handreck, K.A. Silica in soils, plant and animals. Adv. Agron. 1967, 19, 107–149. [Google Scholar]
- Coradin, T.; Lopez, P.J.; Gautier, C.; Livage, J. From biogenic to biomimetic silica. Comptes Rendus Palevol 2004, 3, 443–452. [Google Scholar] [CrossRef]
- Vásquez, A.; Meza, C.M.; Rivas, G.M.; Gámez, B.G.; Fierro, A.; Olivares, O. Study on the nutritional properties of Nassella mucronata. In Proceedings of the XVIII Buiatric Congress, Québec, QC, Canada, 11–16 July 2004; Mexican Association of Veterinary Doctors Specialists in Bovines A.C.: Mexico City, Mexico, 2004. [Google Scholar]
- Bokor, B.; Bokorová, S.; Ondoš, S.; Švubová, R.; Lukačová, Z.; Hýblová, M.; Lux, A. Ionome and expression level of Si transporter genes (Lsi1, Lsi2, and Lsi6) affected by Zn and Si interaction in maize. Environ. Sci. Pollut. Res. 2015, 22, 6800–6811. [Google Scholar] [CrossRef]
- Bokor, B.; Ondoš, S.; Vaculík, M.; Bokorová, S.; Weidinger, M.; Lichtscheidl, I.; Turňa, J.; Lux, A. Expression of Genes for Si Uptake, Accumulation, and Correlation of Si with Other Elements in Ionome of Maize Kernel. Front. Plant Sci. 2017, 8, 1063. [Google Scholar] [CrossRef] [Green Version]
- Ge, Y.; Lu, H.; Zhang, J.; Wang, C.; Gao, X. Phytoliths in Inflorescence Bracts: Preliminary Results of an Investigation on Common Panicoideae Plants in China. Front. Plant Sci. 2020, 10, 1736. [Google Scholar] [CrossRef] [Green Version]
- Golokhvast, K.S.; Seryodkin, I.V.; Chaika, V.V.; Zakharenko, A.M.; Pamirsky, I.E. Phytoliths in taxonomy of phylogenetic domains of plants. BioMed Res. Int. 2014, 2014, 648326. [Google Scholar] [CrossRef] [Green Version]
- Erra, G. Systematic assignment and lower paleocommunities from the phytolithic study of Quaternary sediments from Entre Ríos, Argentina. Bol. Soc. Argent. Bot. 2010, 45, 309–319. [Google Scholar]
- Livingstone, D.; Clayton, W. An Altitudinal Cline in Tropical African Grass Floras and Its Paleoecological Significance. Quat. Res. 1980, 13, 392–402. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Sum of Squares | Degrees of Freedom | Mean Squares | F | p-Value |
|---|---|---|---|---|---|
| Block | 0.46 | 2 | 0.23 | ||
| Clipping date | 8.95 | 1 | 8.95 | 25.35 | <0.0001 |
| Cultivar | 11.61 | 6 | 1.94 | 5.48 | 0.0009 |
| Clipping date × Cultivar | 2.31 | 6 | 0.39 | 1.09 | 0.3937 |
| Error | 9.18 | 26 | 0.35 | ||
| Total | 32.51 | 41 |
| Cultivar | Average | N | |||
|---|---|---|---|---|---|
| Don Luis | 2.39 | 6 | A | ||
| Morpa | 2.54 | 6 | A | ||
| Tanganyika | 2.57 | 6 | A | ||
| Don Pablo | 2.82 | 6 | A | ||
| 9355 | 3.02 | 6 | A | B | |
| Don Eduardo | 3.66 | 6 | B | C | |
| Don Juan | 3.84 | 6 | C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gallardo, J.; Díaz, M.; Carballo, J.; Garayalde, A.; Echenique, V. Phytolith Content Negatively Affects Forage Quality of Eragrostis curvula (Schrad.) Nees. Agronomy 2023, 13, 924. https://doi.org/10.3390/agronomy13030924
Gallardo J, Díaz M, Carballo J, Garayalde A, Echenique V. Phytolith Content Negatively Affects Forage Quality of Eragrostis curvula (Schrad.) Nees. Agronomy. 2023; 13(3):924. https://doi.org/10.3390/agronomy13030924
Chicago/Turabian StyleGallardo, Jimena, Marina Díaz, José Carballo, Antonio Garayalde, and Viviana Echenique. 2023. "Phytolith Content Negatively Affects Forage Quality of Eragrostis curvula (Schrad.) Nees" Agronomy 13, no. 3: 924. https://doi.org/10.3390/agronomy13030924