
How to Efficiently Produce the Selenium-Enriched Cucumber Fruit with High Yield and Qualities via Hydroponic Cultivation? The Balance between Selenium Supply and CO2 Fertilization

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Sampling and Measurements
2.2.1. Gas-Exchange Rate Measurements
2.2.2. Plant Harvest and Weight Determination
2.2.3. Leaf Pigment Concentration Determination
2.2.4. Fruit Organic Nutrient Concentration Determination
2.2.5. Fruit Inorganic Mineral Concentration Determination
2.3. Statistical Analysis
3. Results
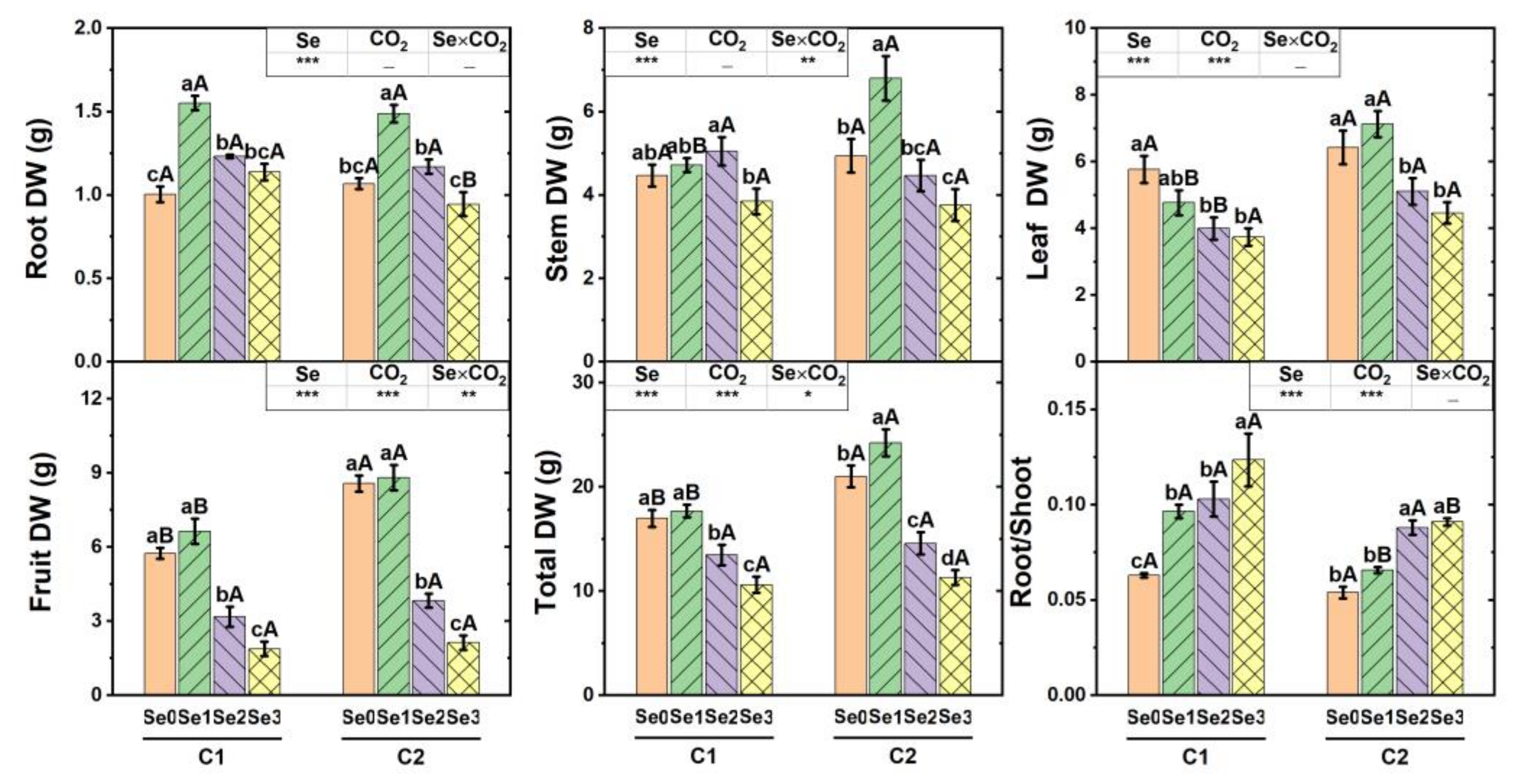
3.1. Dry Weight of Cucumber Plant
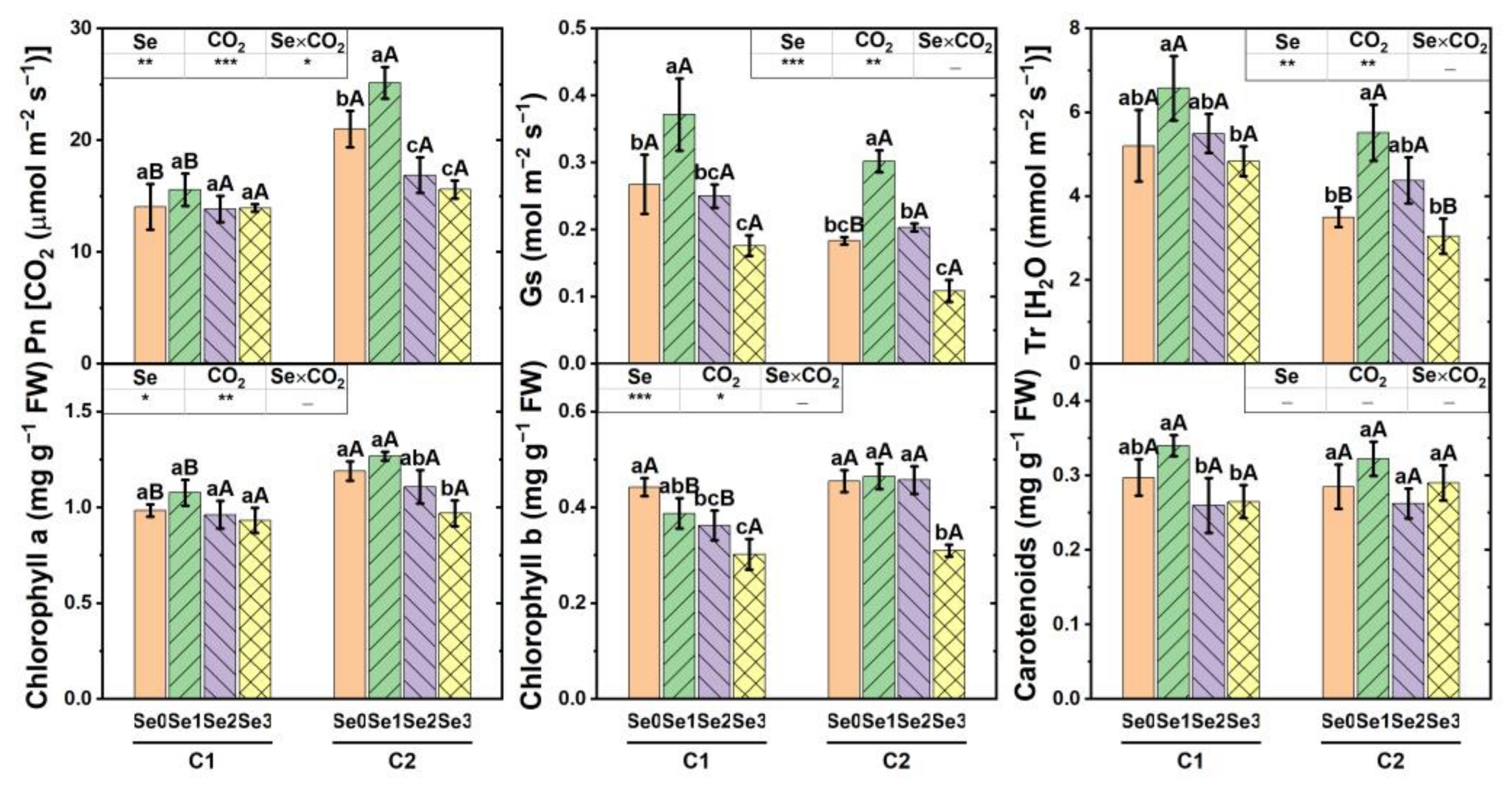
3.2. Gas Exchange Property and Pigment Concentration in Cucumber Leaf
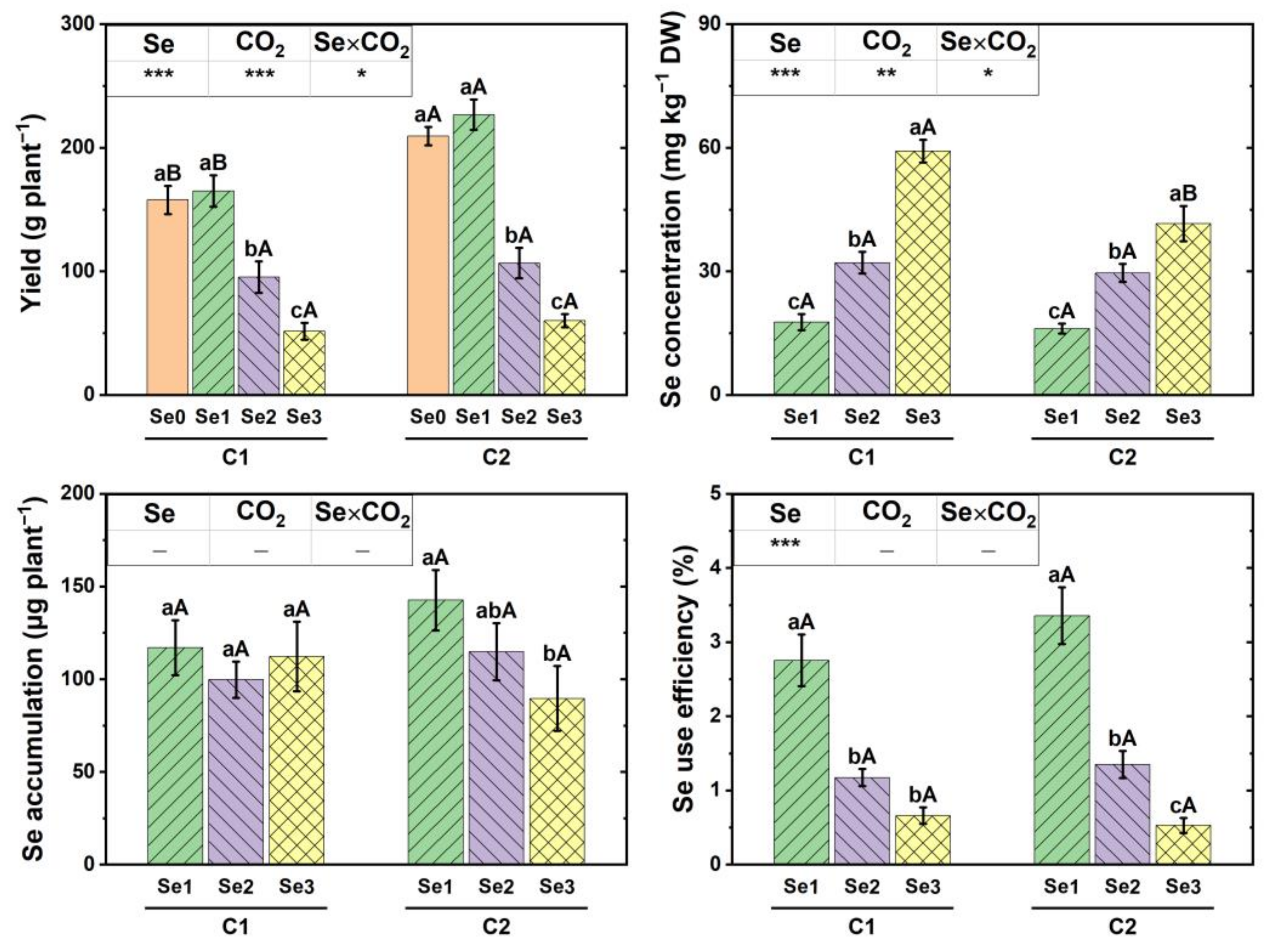
3.3. Yield and Se Biofortification of Cucumber Fruit
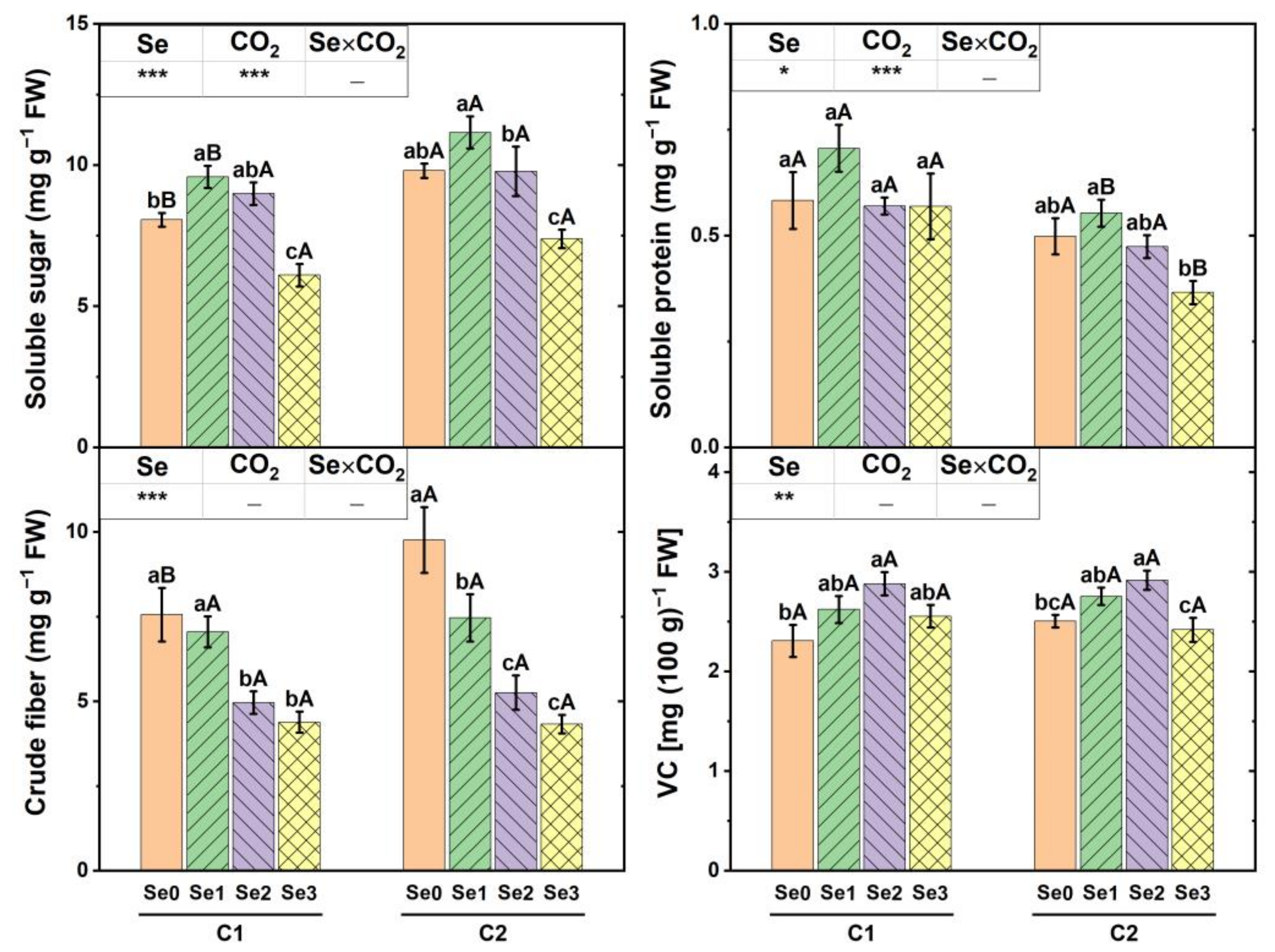
3.4. Organic Nutrients in Cucumber Fruit
3.5. Inorganic Minerals in Cucumber Fruit
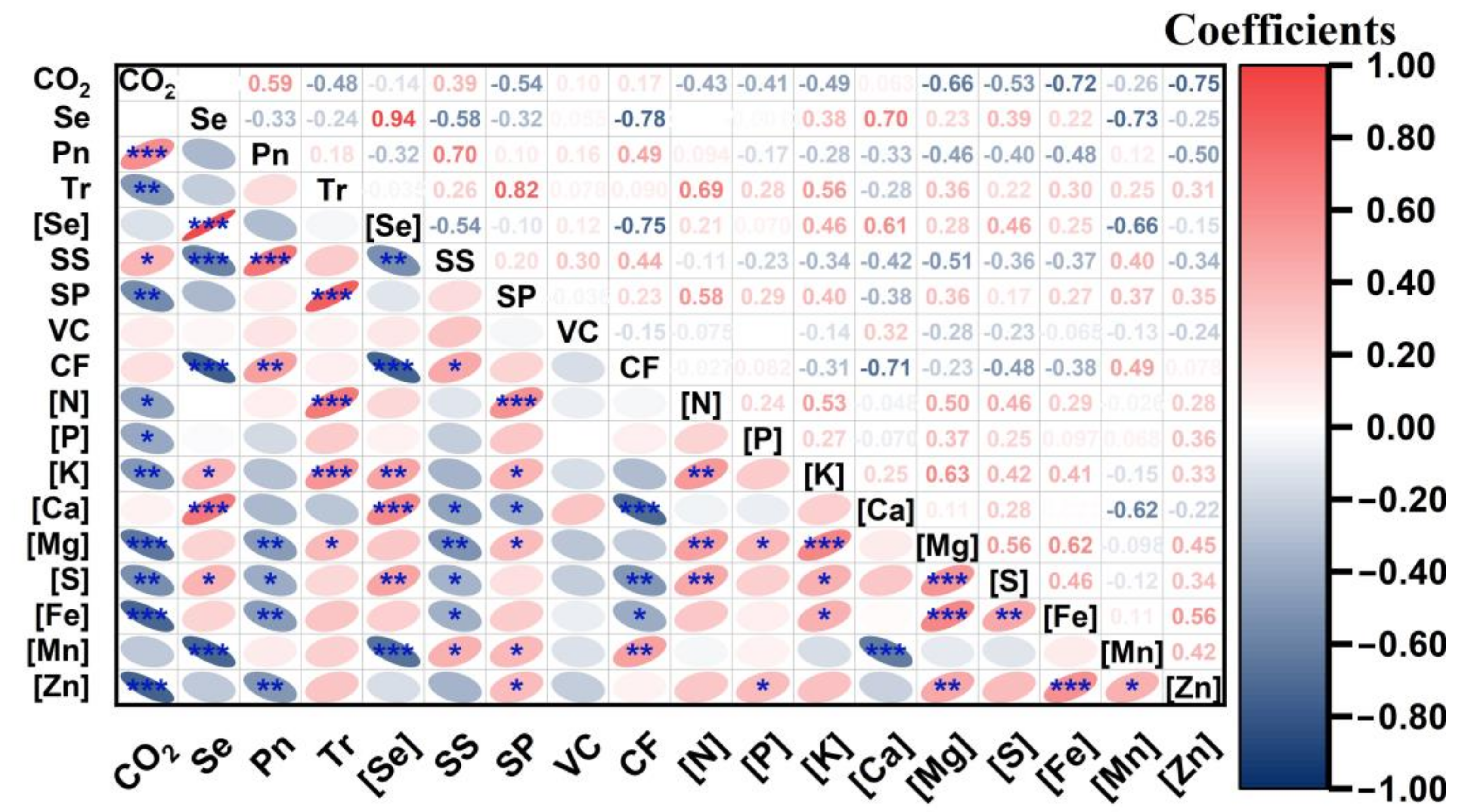
3.6. Correlations between [CO2], Se Supply and Nutritional Qualities of Cucumber Fruits
4. Discussion
4.1. Effects of [CO2] and Se Supply Levels on the Growth and Gas Exchange Parameters of Cucumber Plants
4.2. Effects of [CO2] and Se Supply Levels on the Yield and Se Biofortification of Cucumber Fruits
4.3. Effects of [CO2] and Se Supply Levels on the Nutritional Qualities of Cucumber Fruits
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, X.; He, H.; Xiang, J.; Yin, H.; Hou, T. Selenium-Containing Proteins/Peptides from Plants: A Review on the Structures and Functions. J. Agric. Food Chem. 2020, 68, 15061–15073. [Google Scholar] [CrossRef] [PubMed]
- Wen, D. Selenium in horticultural crops. Sci. Hortic. 2021, 289, 110441. [Google Scholar] [CrossRef]
- Zhao, M.; Luo, T.; Zhao, Z.; Rong, H.; Zhao, G.; Lei, L. Food Chemistry of Selenium and Controversial Roles of Selenium in Affecting Blood Cholesterol Concentrations. J. Agric. Food Chem. 2021, 69, 4935–4945. [Google Scholar] [CrossRef] [PubMed]
- Tapiero, H.; Townsend, D.M.; Tew, K.D. The antioxidant role of selenium and seleno-compounds. Biomed. Pharmacother. 2003, 57, 134–144. [Google Scholar] [CrossRef]
- Khatiwada, S.; Subedi, A. A Mechanistic Link Between Selenium and Coronavirus Disease 2019 (COVID-19). Curr. Nutr. Rep. 2021, 10, 125–136. [Google Scholar] [CrossRef]
- Liu, Q.; Zhao, X.; Ma, J.; Mu, Y.; Wang, Y.; Yang, S.; Wu, Y.; Wu, F.; Zhou, Y. Selenium (Se) plays a key role in the biological effects of some viruses: Implications for COVID-19. Environ. Res. 2021, 196, 110984. [Google Scholar] [CrossRef]
- Zhang, J.; Saad, R.; Taylor, E.W.; Rayman, M.P. Selenium and selenoproteins in viral infection with potential relevance to COVID-19. Redox Biol. 2020, 37, 101715. [Google Scholar] [CrossRef]
- Dinh, Q.T.; Wang, M.; Tran, T.A.T.; Zhou, F.; Wang, D.; Zhai, H.; Peng, Q.; Xue, M.; Du, Z.; Bañuelos, G.S.; et al. Bioavailability of selenium in soil-plant system and a regulatory approach. Crit. Rev. Environ. Sci. Technol. 2018, 49, 443–517. [Google Scholar] [CrossRef]
- White, P.J. Selenium accumulation by plants. Ann. Bot. 2016, 117, 217–235. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, I.; Smoleń, S.; Czernicka, M.; Halka, M.; Kęska, K.; Pitala, J. Effect of Selenium Form and Salicylic Acid on the Accumulation of Selenium Speciation Forms in Hydroponically Grown Lettuce. Agriculture 2020, 10, 584. [Google Scholar] [CrossRef]
- Malagoli, M.; Schiavon, M.; dall’Acqua, S.; Pilon-Smits, E.A. Effects of selenium biofortification on crop nutritional quality. Front. Plant Sci. 2015, 6, 280. [Google Scholar] [CrossRef] [Green Version]
- Raina, M.; Sharma, A.; Nazir, M.; Kumari, P.; Rustagi, A.; Hami, A.; Bhau, B.S.; Zargar, S.M.; Kumar, D. Exploring the new dimensions of selenium research to understand the underlying mechanism of its uptake, translocation, and accumulation. Physiol. Plant 2021, 171, 882–895. [Google Scholar] [CrossRef]
- Gruda, N. Impact of Environmental Factors on Product Quality of Greenhouse Vegetables for Fresh Consumption. Crit. Rev. Plant Sci. 2005, 24, 227–247. [Google Scholar] [CrossRef]
- Gruda, N.; Savvas, D.; Colla, G.; Rouphael, Y. Impacts of genetic material and current technologies on product quality of selected greenhouse vegetables—A review. Eur. J. Hortic. Sci. 2018, 83, 319–328. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Herforth, A.; Arimond, M.; Alvarez-Sanchez, C.; Coates, J.; Christianson, K.; Muehlhoff, E. A Global Review of Food-Based Dietary Guidelines. Adv. Nutr. 2019, 10, 590–605. [Google Scholar] [CrossRef] [Green Version]
- Miller, V.; Yusuf, S.; Chow, C.K.; Dehghan, M.; Corsi, D.J.; Lock, K.; Popkin, B.; Rangarajan, S.; Khatib, R.; Lear, S.A.; et al. Availability, affordability, and consumption of fruits and vegetables in 18 countries across income levels: Findings from the Prospective Urban Rural Epidemiology (PURE) study. Lancet Glob. Health 2016, 4, e695–e703. [Google Scholar] [CrossRef] [Green Version]
- Boeing, H.; Bechthold, A.; Bub, A.; Ellinger, S.; Haller, D.; Kroke, A.; Leschik-Bonnet, E.; Muller, M.J.; Oberritter, H.; Schulze, M.; et al. Critical review: Vegetables and fruit in the prevention of chronic diseases. Eur. J. Nutr. 2012, 51, 637–663. [Google Scholar] [CrossRef] [Green Version]
- Gruda, N. Assessing the impact of environmental factors on the quality of greenhouse produce. In Achieving Sustainable Greenhouse Cultivation; Marcelis, L., Heuvelink, E., Eds.; Burleigh Dodds Science Publishing Limited: Cambridge, UK, 2019. [Google Scholar] [CrossRef]
- Puccinelli, M.; Malorgio, F.; Terry, L.A.; Tosetti, R.; Rosellini, I.; Pezzarossa, B. Effect of selenium enrichment on metabolism of tomato (Solanum lycopersicum) fruit during postharvest ripening. J. Sci. Food Agric. 2019, 99, 2463–2472. [Google Scholar] [CrossRef] [Green Version]
- Funes-Collado, V.; Morell-Garcia, A.; Rubio, R.; López-Sánchez, J.F. Selenium uptake by edible plants from enriched peat. Sci. Hortic. 2013, 164, 428–433. [Google Scholar] [CrossRef] [Green Version]
- Rahim, F.P.; Rocio, C.G.; Adalberto, B.M.; Lidia Rosaura, S.C.; Maginot, N.H. Agronomic Biofortification with Selenium in Tomato Crops (Solanum lycopersicon L. Mill). Agriculture 2020, 10, 486. [Google Scholar] [CrossRef]
- Hu, W.; Su, Y.; Zhou, J.; Zhu, H.; Guo, J.; Huo, H.; Gong, H. Foliar application of silicon and selenium improves the growth, yield and quality characteristics of cucumber in field conditions. Sci. Hortic. 2022, 294, 110776. [Google Scholar] [CrossRef]
- Puccinelli, M.; Malorgio, F.; Pezzarossa, B. Selenium Enrichment of Horticultural Crops. Molecules 2017, 22, 933. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.J.; Meacham, S.L.; Kruskall, L.J. The antioxidants-vitamin C, vitamin E, selenium, and carotenoids. J. Agromedicine 2003, 9, 65–82. [Google Scholar] [CrossRef] [PubMed]
- Monsen, E.R. Dietary reference intakes for the antioxidant nutrients: Vitamin C, vitamin E, selenium, and carotenoids. J. Am. Diet. Assoc. 2000, 100, 637–640. [Google Scholar] [CrossRef]
- Higgs, D.J.; Morris, V.C.; Levander, O.A. Effect of cooking on selenium content of foods. J. Agric. Food Chem. 1972, 20, 678–680. [Google Scholar] [CrossRef]
- Dong, Z.; Liu, Y.; Dong, G.; Wu, H. Effect of boiling and frying on the selenium content, speciation, and in vitro bioaccessibility of selenium-biofortified potato (Solanum tuberosum L.). Food Chem. 2021, 348, 129150. [Google Scholar] [CrossRef]
- Gupta, M.; Gupta, S. An Overview of Selenium Uptake, Metabolism, and Toxicity in Plants. Front. Plant Sci. 2016, 7, 2074. [Google Scholar] [CrossRef] [Green Version]
- Bisbis, M.B.; Gruda, N.; Blanke, M. Potential impacts of climate change on vegetable production and product quality—A review. J. Clean. Prod. 2018, 170, 1602–1620. [Google Scholar] [CrossRef]
- Dong, J.; Gruda, N.; Li, X.; Tang, Y.; Zhang, P.; Duan, Z. Sustainable vegetable production under changing climate: The impact of elevated CO2 on yield of vegetables and the interactions with environments-A review. J. Clean. Prod. 2020, 253, 119920. [Google Scholar] [CrossRef]
- Dong, J.; Gruda, N.; Lam, S.K.; Li, X.; Duan, Z. Effects of elevated CO2 on nutritional quality of vegetables: A review. Front. Plant Sci. 2018, 9, 924. [Google Scholar] [CrossRef]
- Li, X.; Dong, J.; Gruda, N.; Chu, W.; Duan, Z. Does the short-term fluctuation of mineral element concentrations in the closed hydroponic experimental facilities affect the mineral concentrations in cucumber plants exposed to elevated CO2? Plant Soil 2021, 465, 125–141. [Google Scholar] [CrossRef]
- Sardans, J.; Grau, O.; Chen, H.Y.H.; Janssens, I.A.; Ciais, P.; Piao, S.; Penuelas, J. Changes in nutrient concentrations of leaves and roots in response to global change factors. Glob. Chang Biol. 2017, 23, 3849–3856. [Google Scholar] [CrossRef] [Green Version]
- Bai, B.; Wang, Z.; Gao, L.; Chen, W.; Shen, Y. Effects of selenite on the growth of alfalfa (Medicago sativa L. cv. Sadie 7) and related physiological mechanisms. Acta Physiol. Plant. 2019, 41, 78. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B.; Matraszek, R.; Pogorzelec, M. The dual effects of two inorganic selenium forms on the growth, selected physiological parameters and macronutrients accumulation in cucumber plants. Acta Physiol. Plant. 2015, 37, 41. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Dong, J.; Gruda, N.S.; Chu, W.; Duan, Z. Interactive effects of the CO2 enrichment and nitrogen supply on the biomass accumulation, gas exchange properties, and mineral elements concentrations in cucumber plants at different growth stages. Agronomy 2020, 10, 139. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, K. Nutrient Solution Culture; Pak-Kyo, Co.: Tokyo, Japan, 1982; p. 251. [Google Scholar]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Van Soest, P.J. Use of detergents in the analysis of fibrous feeds. 2. A rapid method for the determination of fiber and lignin. J. Assoc. Off. Agric. Chem. 1963, 46, 829–835. [Google Scholar] [CrossRef]
- Nelson, W.L.; Somers, G.F. Determination of Ascorbic Acid—Application of the Indophenol-Xylene Extraction Method to Determination in Large Numbers of Tomato and Tomato Juice Samples. Ind. Eng. Chem. 1945, 17, 754–756. [Google Scholar] [CrossRef]
- Zhao, F.; McGrath, S.P.; Crosland, A.R. Comparison of three wet digestion methods for the determination of plant sulphur by inductively coupled plasma atomic emission spectroscopy (ICP-AES). Commun. Soil Sci. Plant Anal. 1994, 25, 407–418. [Google Scholar] [CrossRef]
- Zhang, M.; Tang, S.; Huang, X.; Zhang, F.; Pang, Y.; Huang, Q.; Yi, Q. Selenium uptake, dynamic changes in selenium content and its influence on photosynthesis and chlorophyll fluorescence in rice (Oryza sativa L.). Environ. Exp. Bot. 2014, 107, 39–45. [Google Scholar] [CrossRef]
- Van der Kooi, C.J.; Reich, M.; Löw, M.; De Kok, L.J.; Tausz, M. Growth and yield stimulation under elevated CO2 and drought: A meta-analysis on crops. Environ. Exp. Bot. 2016, 122, 150–157. [Google Scholar] [CrossRef]
- Mortensen, L.M. Review: CO2 enrichment in greenhouses. Crop responses. Sci. Hortic. 1987, 33, 1–25. [Google Scholar] [CrossRef]
- Haghighi, M.; Sheibanirad, A.; Pessarakli, M. Effects of selenium as a beneficial element on growth and photosynthetic attributes of greenhouse cucumber. J. Plant Nutr. 2015, 39, 1493–1498. [Google Scholar] [CrossRef] [Green Version]
- Jozwiak, W.; Mleczek, M.; Politycka, B. The effect of exogenous selenium on the growth and photosynthetic pigments content of cucumber seedlings. Fresenius Environ. Bull. 2016, 25, 142–152. [Google Scholar]
- Feng, R.; Wei, C.; Tu, S. The roles of selenium in protecting plants against abiotic stresses. Environ. Exp. Bot. 2013, 87, 58–68. [Google Scholar] [CrossRef]
- Balal, R.M.; Shahid, M.A.; Javaid, M.M.; Iqbal, Z.; Anjum, M.A.; Garcia-Sanchez, F.; Mattson, N.S. The role of selenium in amelioration of heat-induced oxidative damage in cucumber under high temperature stress. Acta Physiol. Plant. 2016, 38, 158. [Google Scholar] [CrossRef]
- Hajiboland, R.; Sadeghzade, N. Effect of selenium on CO2 and NO3− assimilation under low and adequate nitrogen supply in wheat (Triticum aestivum L.). Photosynthetica 2014, 52, 501–510. [Google Scholar] [CrossRef]
- Lehotai, N.; Kolbert, Z.; Peto, A.; Feigl, G.; Ordog, A.; Kumar, D.; Tari, I.; Erdei, L. Selenite-induced hormonal and signalling mechanisms during root growth of Arabidopsis thaliana L. J. Exp. Bot. 2012, 63, 5677–5687. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Dong, J.; Gruda, N.S.; Li, X.; Duan, Z. Elevated root-zone temperature promotes the growth and alleviates the photosynthetic acclimation of cucumber plants exposed to elevated [CO2]. Environ. Exp. Bot. 2022, 194, 104694. [Google Scholar] [CrossRef]
- De Almeida, H.J.; Carmona, V.V.; Dutra, A.F.; Cecílio Filho, A.B. Growth and physiological responses of cabbage cultivars biofortified with inorganic selenium fertilizers. Sci. Hortic. 2022, 302, 111154. [Google Scholar] [CrossRef]
- Businelli, D.; D’Amato, R.; Onofri, A.; Tedeschini, E.; Tei, F. Se-enrichment of cucumber (Cucumis sativus L.), lettuce (Lactuca sativa L.) and tomato (Solanum lycopersicum L. Karst) through fortification in pre-transplanting. Sci. Hortic. 2015, 197, 697–704. [Google Scholar] [CrossRef]
- Zhou, X.; Yang, J.; Kronzucker, H.J.; Shi, W. Selenium Biofortification and Interaction with other Elements in Plants: A Review. Front. Plant Sci. 2020, 11, 586421. [Google Scholar] [CrossRef]
- Li, H.F.; McGrath, S.P.; Zhao, F.J. Selenium uptake, translocation and speciation in wheat supplied with selenate or selenite. New Phytol. 2008, 178, 92–102. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, B.; Li, W.; Che, R.; Deng, K.; Li, H.; Yu, F.; Ling, H.; Li, Y.; Chu, C. OsPT2, a phosphate transporter, is involved in the active uptake of selenite in rice. New Phytol. 2014, 201, 1183–1191. [Google Scholar] [CrossRef] [Green Version]
- Sabatino, L.; La Bella, S.; Ntatsi, G.; Iapichino, G.; D’Anna, F.; De Pasquale, C.; Consentino, B.B.; Rouphael, Y. Selenium biofortification and grafting modulate plant performance and functional features of cherry tomato grown in a soilless system. Sci. Hortic. 2021, 285, 110095. [Google Scholar] [CrossRef]
- Cipriano, P.E.; Siueia Júnior, M.; de Souza, R.R.; da Silva, D.F.; da Silva, R.F.; Faquin, V.; de Souza Silva, M.L.; Guilherme, L.R.G. Macronutrients content of radishes and the influence of biofortification with selenium. Sci. Hortic. 2022, 296, 110908. [Google Scholar] [CrossRef]
- Peng, Q.; Zhang, Z.; Su, R.; Zhang, X.; Lambers, H.; He, H. Phosphorus and selenium uptake, root morphology, and carboxylates in the rhizosheath of alfalfa (Medicago sativa) as affected by localised phosphate and selenite supply in a split-root system. Funct. Plant Biol. 2021, 48, 1161–1174. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Randle, W.M.; Mills, H.A. Nutrient accumulation in leaf tissue of rapid-cyclingbrassuca oleracearesponds to increasing sodium selenate concentrations. J. Plant Nutr. 2000, 23, 927–935. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B. Effect of selenium on selected macronutrients in maize plants. J. Elementol. 2008, 13, 513–519. [Google Scholar]
- Guerrero, B.; Llugany, M.; Palacios, O.; Valiente, M. Dual effects of different selenium species on wheat. Plant Physiol. Biochem. 2014, 83, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.A.; Wick, J.E.; Famuyide, I.M.; McGaw, L.J.; Mühling, K.H. Selenium Enrichment of Green and Red Lettuce and the Induction of Radical Scavenging Potential. Horticulturae 2021, 7, 488. [Google Scholar] [CrossRef]
- Teixeira, L.S.; Pimenta, T.M.; Brito, F.A.L.; Malheiros, R.S.P.; Arruda, R.S.; Araujo, W.L.; Ribeiro, D.M. Selenium uptake and grain nutritional quality are affected by nitrogen fertilization in rice (Oryza sativa L.). Plant Cell Rep. 2021, 40, 871–880. [Google Scholar] [CrossRef]
- Huang, S.; Yu, K.; Xiao, Q.; Song, B.; Yuan, W.; Long, X.; Cai, D.; Xiong, X.; Zheng, W. Effect of bio-nano-selenium on yield, nutritional quality and selenium content of radish. J. Food Compos. Anal. 2023, 115, 104927. [Google Scholar] [CrossRef]
- Gonzalez-Lemus, U.; Medina-Perez, G.; Espino-Garcia, J.J.; Fernandez-Luqueno, F.; Campos-Montiel, R.; Almaraz-Buendia, I.; Reyes-Munguia, A.; Urrutia-Hernandez, T. Nutritional Parameters, Biomass Production, and Antioxidant Activity of Festuca arundinacea Schreb. Conditioned with Selenium Nanoparticles. Plants 2022, 11, 2326. [Google Scholar] [CrossRef]
- Lu, N.; Wu, L.; Zhang, X.; Zhang, Y.; Shan, C. Selenium improves the content of vitamin C in the fruit of strawberry by regulating the enzymes responsible for vitamin C metabolism. Plant Soil Environ. 2022, 68, 205–211. [Google Scholar] [CrossRef]
- Handa, N.; Kohli, S.K.; Sharma, A.; Thukral, A.K.; Bhardwaj, R.; Abd_Allah, E.F.; Alqarawi, A.A.; Ahmad, P. Selenium modulates dynamics of antioxidative defence expression, photosynthetic attributes and secondary metabolites to mitigate chromium toxicity in Brassica juncea L. plants. Environ. Exp. Bot. 2019, 161, 180–192. [Google Scholar] [CrossRef]
- Luo, L.; Zhang, J.; Zhang, K.; Wen, Q.; Ming, K.; Xiong, H.; Ning, F. Peanut selenium distribution, concentration, speciation, and effects on proteins after exogenous selenium biofortification. Food Chem. 2021, 354, 129515. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments 1 | [Se] (mg kg−1 FW) | RDA (g day−1) | UL (g day−1) |
|---|---|---|---|
| C1Se1 | 0.714 | 77.03 | 560.2 |
| C1Se2 | 1.067 | 51.55 | 374.9 |
| C1Se3 | 2.163 | 25.43 | 184.9 |
| C2Se1 | 0.630 | 87.30 | 634.9 |
| C2Se2 | 1.073 | 51.26 | 372.8 |
| C2Se3 | 1.480 | 37.16 | 270.3 |
| Factor 1 | [N] | [P] | [K] | [Ca] | [Mg] | [S] | [Fe] | [Mn] | [Zn] |
|---|---|---|---|---|---|---|---|---|---|
| CO2 | * 2 | * | ** | NS | *** | ** | *** | * | *** |
| Se | NS 3 | NS | NS | *** | * | NS | NS | *** | * |
| CO2 × Se | NS | NS | NS | NS | NS | NS | NS | NS | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Li, D.; Gruda, N.S.; Zhu, C.; Duan, Z.; Li, X. How to Efficiently Produce the Selenium-Enriched Cucumber Fruit with High Yield and Qualities via Hydroponic Cultivation? The Balance between Selenium Supply and CO2 Fertilization. Agronomy 2023, 13, 922. https://doi.org/10.3390/agronomy13030922
Wang Z, Li D, Gruda NS, Zhu C, Duan Z, Li X. How to Efficiently Produce the Selenium-Enriched Cucumber Fruit with High Yield and Qualities via Hydroponic Cultivation? The Balance between Selenium Supply and CO2 Fertilization. Agronomy. 2023; 13(3):922. https://doi.org/10.3390/agronomy13030922
Chicago/Turabian StyleWang, Ziying, Di Li, Nazim S. Gruda, Chunwu Zhu, Zengqiang Duan, and Xun Li. 2023. "How to Efficiently Produce the Selenium-Enriched Cucumber Fruit with High Yield and Qualities via Hydroponic Cultivation? The Balance between Selenium Supply and CO2 Fertilization" Agronomy 13, no. 3: 922. https://doi.org/10.3390/agronomy13030922