
Specific Gibberellin 2-Oxidase 3 (SbGA2ox3) Mutants Promote Yield and Stress Tolerance in Sorghum bicolor
Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of the EMS Sorghum Mutant Population
2.2. Investigation of the Agronomic Character of Plants in the Field
2.3. EMS Library Preparation and Sequencing
2.4. Mutation Density Evaluation
2.5. SbGA2ox3 Sequences and Phylogenetic Analysis
2.6. Determination of Soluble Sugar and Malondialdehyde Content
2.7. Gene Expression Analysis of SbGA2ox3 in Sorghum
2.8. Statistical Analysis
3. Results
3.1. Development of the Sorghum bicolor Mutant Population
3.2. Prediction and Structural Analysis of the Sorghum SbGA2ox3 Gene
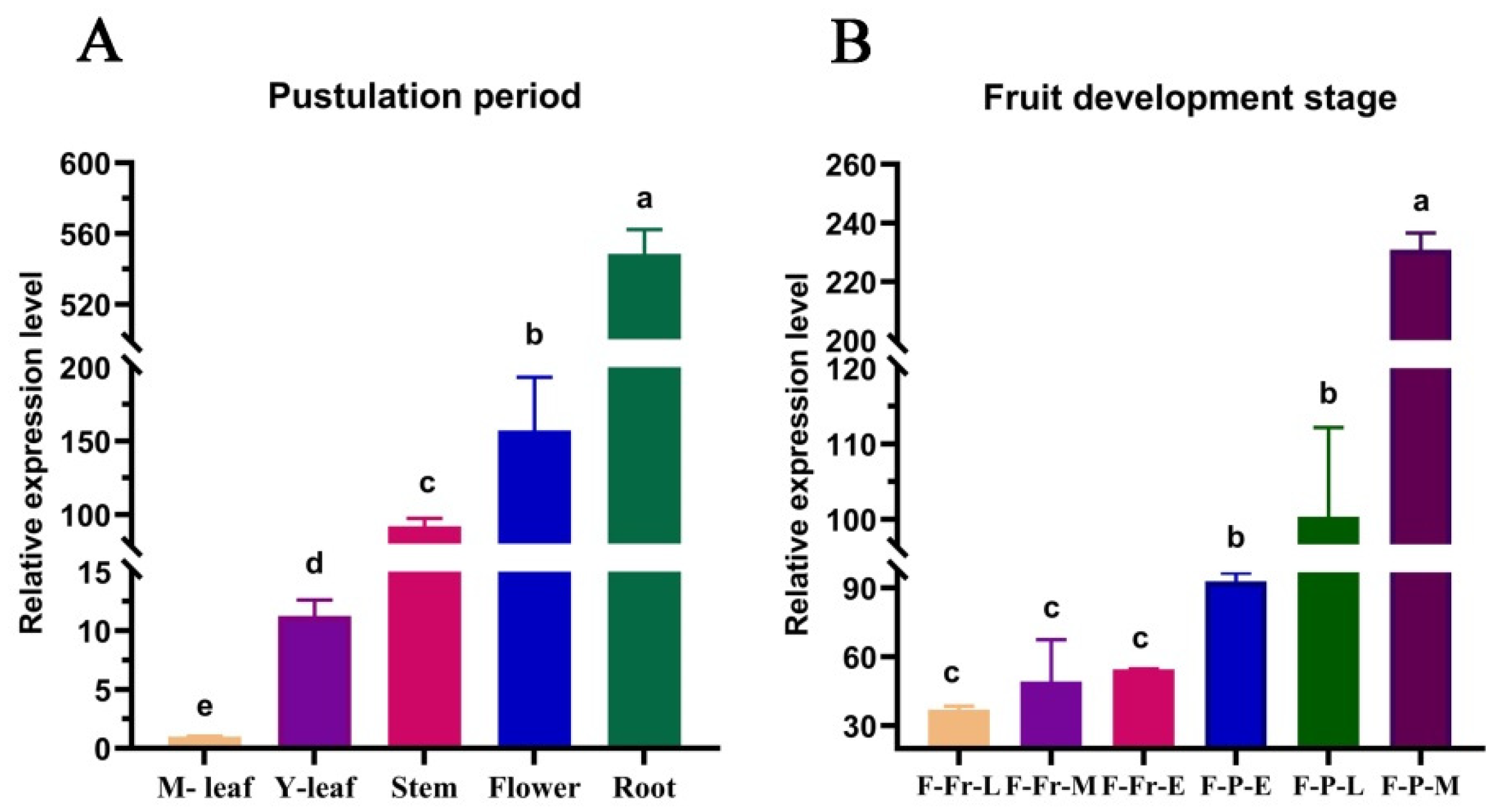
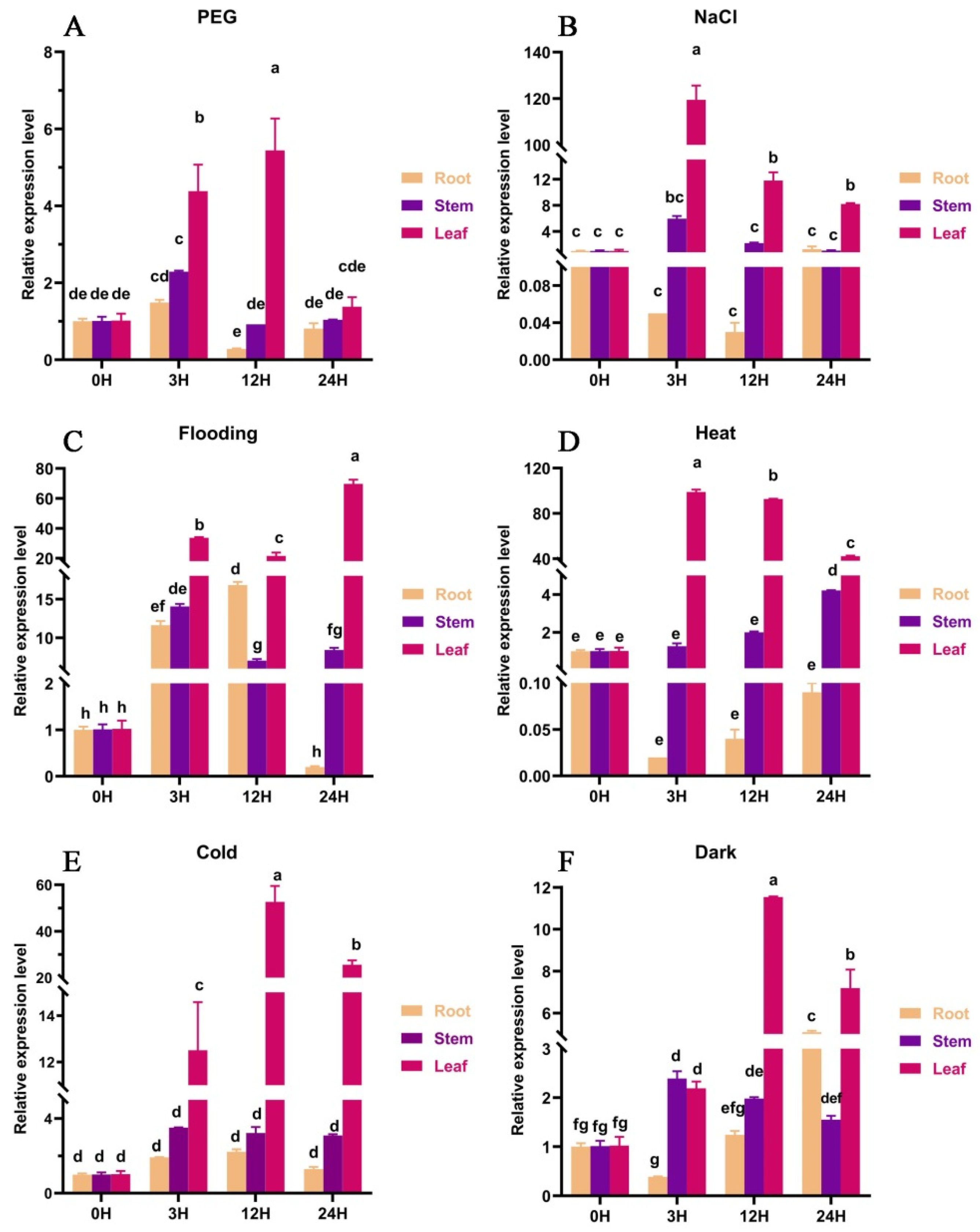
3.3. Expression Analysis of the Sorghum SbGA2ox3 Gene
3.4. Mutant Screening of SbGA2ox3
3.5. Drought Resistance and Phenotypic Analysis of Mutants
4. Discussion
5. Conclusions
Conclusions and Prospects
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xie, P.; Shi, J.Y.; Tang, S.Y.; Chen, C.X.; Khan, A.; Zhang, F.X.; Xiong, Y.; Li, C.; He, W.; Wang, G.D.; et al. Control of Bird Feeding Behavior by Tannin1 through Modulating the Biosynthesis of Polyphenols and Fatty Acid-Derived Volatiles in Sorghum. Mol. Plant 2019, 12, 1315–1324. [Google Scholar] [CrossRef] [PubMed]
- Ouyang, H.; Jiang, J.M.; Du, Q.L.; Fang, Y.P.; Li, X.Y.; Xie, X. Identification and Bioinformatics Analysis of Shaker Protein Family of Potassium Channels in Sorghum. J. Mt. Agric. Biol. 2021, 40, 1–9, (In Chinese with English abstract). [Google Scholar]
- Mace, E.S.; Tai, S.S.; Gilding, E.K.; Li, Y.H.; Prentis, P.J.; Bian, L.L.; Campbell, B.C.; Hu, W.S.; Innes, D.J.; Han, X.; et al. Whole-genome sequencing reveals untapped genetic potential in Africa’s indigenous cereal crop sorghum. Nat. Commun. 2013, 4, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, H.Q.; Li, Z.G.; Leng, C.Y.; Lu, C.; Luo, H.; Liu, Y.M.; Wu, X.Y.; Liu, Z.Q.; Shang, L.; Jing, H.C. Sorghum breeding in the genomic era: Opportunities and challenges. Theor. Appl. Genet. 2021, 134, 1899–1924. [Google Scholar] [CrossRef] [PubMed]
- Brenton, Z.W.; Cooper, E.A.; Myers, M.T.; Boyles, R.E.; Shakoor, N.; Zielinski, K.J.; Rauh, B.L.; Bridges, W.C.; Morris, G.P.; Kresovich, S. A Genomic Resource for the Development, Improvement, and Exploitation of Sorghum for Bioenergy. Genetics 2016, 204, 21. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Huang, C.; Zhao, W.; Dai, B.; Shen, C.; Zhang, B.; Li, D.; Lin, Z. Identification of QTL for Fiber Quality and Yield Traits Using Two Immortalized Backcross Populations in Upland Cotton. PLoS ONE 2016, 11, e0166970. [Google Scholar] [CrossRef] [Green Version]
- Henry, I.M.; Nagalakshmi, U.; Lieberman, M.C.; Ngo, K.J.; Krasileva, K.V.; Vasquez-Gross, H.; Akhunova, A.; Akhunov, E.; Dubcovsky, J.; Tai, T.H.; et al. Efficient Genome-Wide Detection and Cataloging of EMS-Induced Mutations Using Exome Capture and Next-Generation Sequencing. Plant Cell 2014, 26, 1382–1397. [Google Scholar] [CrossRef] [Green Version]
- Tsuda, M.; Kaga, A.; Anai, T.; Shimizu, T.; Sayama, T.; Takagi, K.; Machita, K.; Watanabe, S.; Nishimura, M.; Yamada, N.; et al. Construction of a high-density mutant library in soybean and development of a mutant retrieval method using amplicon sequencing. BMC Genom. 2015, 16, 1014. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.Z.; Duan, L.; Sun, M.H.; Yang, Z.; Li, H.Y.; Hu, K.M.; Yang, H.; Liu, L. Current trends and insights on EMS mutagenesis application to studies on plant abiotic stress tolerance and development. Front. Plant Sci. 2023, 13, 1052569. [Google Scholar] [CrossRef]
- Lethin, J.; Shakil, S.S.M.; Hassan, S.; Sirijovski, N.; Töpel, M.; Olsson, O.; Aronsson, H. Development and characterization of an EMS-mutagenized wheat population and identification of salt-tolerant wheat lines. BMC Plant Biol. 2020, 20, 18. [Google Scholar] [CrossRef] [Green Version]
- Ferrie, A.M.R.; Bhowmik, P.; Rajagopalan, N.; Kagale, S. CRISPR/Cas9-Mediated Targeted Mutagenesis in Wheat Doubled Haploids. Methods Mol. Biol. 2020, 2072, 183–198. [Google Scholar] [PubMed]
- Martin, B.; Ramiro, M.; Martinez-Zapater, J.M.; Alonso-Blanco, C. A high-density collection of EMS-induced mutations for TILLING in Landsberg erecta genetic background of Arabidopsis. BMC Plant Biol. 2009, 9, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chhun, T.; Aya, K.; Asano, K.; Yamamoto, E.; Morinaka, Y.; Watanabe, M.; Kitano, H.; Ashikari, M.; Matsuoka, M.; Ueguchi-Tanaka, M. Gibberellin regulates pollen viability and pollen tube growth in rice. Plant Cell 2007, 19, 3876–3888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aya, K.; Ueguchi-Tanaka, M.; Kondo, M.; Hamada, K.; Yano, K.; Nishimura, M.; Matsuoka, M. Gibberellin modulates another development in rice via the transcriptional regulation of GAMYB. Plant Cell 2009, 21, 1453–1472. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.; Chu, C. Gibberellin Metabolism and Signaling: Targets for Improving Agronomic Performance of Crops. Plant Cell Physiol. 2020, 61, 1902–1911. [Google Scholar] [CrossRef]
- Dreher, K.; Callis, J. Ubiquitin, hormones, and biotic stress in plants. Ann Bot. 2007, 99, 787–822. [Google Scholar] [CrossRef]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The role of gibberellin signaling in plant responses to abiotic stress. J. Exp. Biol. 2014, 217, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Fujioka, S.; Yamane, H.; Spray, C.R.; Katsumi, M.; Phinney, B.O.; Gaskin, P.; Macmillan, J.; Takahashi, N. The dominant non-gibberellin-responding dwarf mutant (D8) of maize accumulates native gibberellins. Proc. Nat. Acad. Sci. USA 1988, 85, 9031–9035. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.H.; Zhang, F.M.; Ge, S. Evolutionary rate patterns of the Gibberellin pathway. BMC Evol. Biol. 2009, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Wang, X.; Ge, S.; Rao, G.Y. Divergence and adaptive evolution of the gibberellin oxidase genes in plants. BMC Evol. Biol. 2015, 15, 15. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.C.; Underhill, S.J.R. Breadfruit (Artocarpus altilis) gibberellin 2-oxidase genes in stem elongation and abiotic stress response. Plant Physiol. Biochem. 2016, 98, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Wuddineh, W.A.; Mazarei, M.; Zhang, J.Y.; Poovaiah, C.R.; Mann, D.G.J.; Ziebell, A.; Sykes, R.W.; Davis, M.F.; Udvardi, M.K.; Stewart, C.N. Identification and overexpression of gibberellin 2-oxidase (GA2ox) in switchgrass (Panicum virgatum L.) for improved plant architecture and reduced biomass recalcitrance. Plant Biotechnol. J. 2015, 13, 636–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, T.; Miura, K.; Itoh, H.; Tatsumi, T.; Ueguchi-Tanaka, M.; Ishiyama, K.; Kobayashi, M.; Agrawal, G.K.; Takeda, S.; Abe, K.; et al. An overview of gibberellin metabolism enzyme genes and their related mutants in rice. Plant Physiol. 2004, 134, 1642–1653. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zheng, L.L.; Wang, X.N.; Hu, Z.B.; Zheng, Y.; Chen, Q.H.; Hao, X.C.; Xiao, X.; Wang, X.B.; Wang, G.D.; et al. Comprehensive expression analysis of Arabidopsis GA2-oxidase genes and their functional insights. Plant Sci. 2019, 285, 1–13. [Google Scholar] [CrossRef]
- Busov, V.B.; Meilan, R.; Pearce, D.W.; Ma, C.; Rood, S.B.; Strauss, S.H. Activation tagging of a dominant gibberellin catabolism gene (GA 2-oxidase) from poplar that regulates tree stature. Plant Physiol. 2003, 132, 1283–1291. [Google Scholar] [CrossRef] [Green Version]
- Lo, S.F.; Ho, T.H.D.; Liu, Y.L.; Jiang, M.J.; Hsieh, K.T.; Chen, K.T.; Yu, L.C.; Lee, M.H.; Chen, C.Y.; Huang, T.P.; et al. Ectopic expression of specific GA2 oxidase mutants promotes yield and stress tolerance in rice. Plant Biotechnol. J. 2017, 15, 850–864. [Google Scholar] [CrossRef] [Green Version]
- Shi, J.B.; Wang, N.; Zhou, H.; Xu, Q.H.; Yan, G.T. The role of gibberellin synthase gene GhGA2ox1 in upland cotton (Gossypium hirsutum L.) responses to drought and salt stress. Biotechnol. Appl. Biochem. 2019, 66, 298–308. [Google Scholar] [CrossRef]
- Agharkar, M.; Lomba, P.; Altpeter, F.; Zhang, H.; Kenworthy, K.; Lange, T. Stable expression of AtGA2ox1 in a low-input turfgrass (Paspalum notatum Flugge) reduces bioactive gibberellin levels and improves turf quality under field conditions. Plant Biotechnol. J. 2007, 5, 791–801. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.Y.; Ping, J.A.; Zhao, W.J. Genetic Quality Improvement of Brewing Sorghum in China: Research Progress. J. Agric. 2019, 9, 21–25, (In Chinese with English abstract). [Google Scholar]
- Paterson, A.H.; Bowers, J.E.; Bruggmann, R.; Dubchak, I.; Grimwood, J.; Gundlach, H.; Haberer, G.; Hellsten, U.; Mitros, T.; Poliakov, A.; et al. The Sorghum bicolor genome and the diversification of grasses. Nature 2009, 457, 551–556. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.L. EMS mutagenesis of sorghum and construction of mutant germplasm bank. Chin. Grassl. Soc. 2018, 15–20, (In Chinese with English abstract). [Google Scholar]
- Zhang, W.; Zhao, W.J.; Bai, W.B.; Zhang, Y.; Wang, G.; Shao, R.F.; Xue, D.D. Screening and Identification of EMS Mutants from Sweet Sorghum. Chin. Agric. Sci. Bull. 2020, 36, 22–25, (In Chinese with English abstract). [Google Scholar]
- Kulkarni, K.P.; Vishwakarma, C.; Sahoo, S.P.; Lima, J.M.; Nath, M.; Dokku, P.; Gacche, R.N.; Mohapatra, T.; Robin, S.; Sarla, N.; et al. A substitution mutation in OsCCD7 cosegregates with dwarf and increased tillering phenotype in rice. J. Genet. 2014, 93, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Hua, W.; Tan, C.; Xie, J.Z.; Zhu, J.H.; Shang, Y.; Yang, J.M.; Zhang, X.Q.; Wu, X.J.; Wang, J.M.; Li, C.D. Alternative splicing of a barley gene results in an excess-tillering and semi-dwarf mutant. Theor. Appl. Genet. 2020, 133, 163–177. [Google Scholar] [CrossRef]
- Luo, Z.; Janssen, B.J.; Snowden, K.C. The molecular and genetic regulation of shoot branching. Plant Physiol. 2021, 187, 1033–1044. [Google Scholar] [CrossRef]
- Till; Bradley, J.; Cooper, J.; Tai, T.H.; Colowit, P.; Greene, E.A.; Henikoff, S.; Comai, L. Discovery of chemically induced mutations in rice by TILLING. BMC Plant Biol. 2007, 7, 19. [Google Scholar]
- Gottwald, S.; Bauer, P.; Komatsuda, T.; Lundqvist, U.; Stein, N. TILLING in the two-rowed barley cultivar ‘Barke’ reveals preferred sites of functional diversity in the gene HvHox1. BMC Res. Notes 2009, 2, 258. [Google Scholar] [CrossRef] [Green Version]
- Till, B.J.; Reynolds, S.H.; Weil, C.; Springer, N.; Burtner, C.; Young, K.; Bowers, E.; Codomo, C.A.; Enns, L.C.; Odden, A.R.; et al. Discovery of induced point mutations in maize genes by TILLING. BMC Plant Biol. 2004, 4, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okabe, Y.; Asamizu, E.; Saito, T.; Matsukura, C.; Ariizumi, T.; Bres, C.; Rothan, C.; Mizoguchi, T.; Ezura, H. Tomato TILLING Technology: Development of a Reverse Genetics Tool for the Efficient Isolation of Mutants from Micro-Tom Mutant Libraries. Plant Cell Physiol. 2011, 52, 1994–2005. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.M.; Mace, E.; Tao, Y.F.; Cruickshank, A.; Hunt, C.; Hammer, G.; Jordan, D. Large-scale genome-wide association study reveals that drought-induced lodging in grain sorghum is associated with plant height and traits linked to carbon remobilization. Theor. Appl. Genet. 2020, 133, 3201–3215. [Google Scholar] [CrossRef]
- Schomburg, F.M.; Bizzell, C.M.; Lee, D.J.; Zeevaart, J.A.D.; Amasino, R.M. Overexpression of a novel class of gibberellin 2-oxidases decreases gibberellin levels and creates dwarf plants. Plant Cell 2003, 15, 151–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.G.; Wang, B.; Yuan, L.; Zhang, X.X.; Shi, X.; Zhao, X.F.; Tang, J.H. QTL mapping for kernel-related traits based on the single-segment substitution lines in maize. J. Maize Sci. 2013, 21, 35–40, (In Chinese with English abstract). [Google Scholar]
- Lo, S.F.; Yang, S.Y.; Chen, K.T.; Hsing, Y.I.; Zeevaart, J.A.; Chen, L.J.; Yu, S.M. A novel class of gibberellin 2-oxidases control semi dwarfism, tillering, and root development in rice. Plant Cell 2008, 20, 2603–2618. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Agronomic Traits | Type | Number of Mutant (Line) | Variation Frequency | Total Number of Mutant Plants (Line) | Total Variation Frequency |
|---|---|---|---|---|---|
| Tillering | Weak tillering | 4 | 0.96% | 9 | 2.16% |
| Strong tillering | 5 | 1.20% | |||
| Branchiness | Weak branching | 36 | 8.67% | 45 | 10.84% |
| Medium branching | 6 | 1.45% | |||
| Strong branching | 3 | 0.72% | |||
| Leaf color | Albino seedling | 10 | 2.41% | 19 | 4.58% |
| Yellow seedling | 6 | 1.45% | |||
| Purple leaf | 3 | 0.72% | |||
| Spike type | Around dispersion | 101 | 24.34% | 227 | 54.70% |
| Medium dispersion | 26 | 6.27% | |||
| Medium tight | 61 | 14.70% | |||
| Tight | 39 | 9.40% | |||
| Spike shape | Umbrella-type | 111 | 26.75% | 196 | 47.23% |
| Spindle-shaped | 34 | 8.19% | |||
| Bar-shaped | 51 | 12.29% | |||
| Chaff coating degree | Coating 1/2 | 4 | 0.96% | 116 | 28% |
| All coated | 112 | 26.99% | |||
| Glume color | Brown | 114 | 27.47% | 313 | 75.42% |
| Black | 187 | 45.06% | |||
| Grey | 12 | 2.89% | |||
| Plant growth period | Mature early | 85 | 20.48% | 99 | 23.86% |
| Mature late | 14 | 3.37% | |||
| Abortion degree | Unbeaten | 1 | 0.24% | 67 | 16.14% |
| Minority abortion | 59 | 14.22% | |||
| Majority abortion | 7 | 1.69% | |||
| Plant height | Height | 41 | 9.88% | 98 | 23.61% |
| Low | 57 | 13.73% |
| Plant Number | Normal Base | Mutant Base | Normal Amino Acid | Mutant Amino Acid | Mutation Type |
|---|---|---|---|---|---|
| 3614 | G/G/T/C/G/T/C/G | A/A/G/G/A/G/G/T | Ser/Ala/Val/Leu/Gln/Ser/Pro/Ala | Asn/Thr/Gly/Val/Gln/Ala/Ala/Ser | Missense mutation |
| 3408 | G | A | Ser | Ser | Silent mutation |
| 3266 | C | T | Pro | Pro | Silent mutation |
| 3034 | C | G | Silent mutation | ||
| 3013 | G | A | Val | Met | Missense mutation |
| 3425 | - | T | Insertion mutation (intron) | ||
| 3540 | - | T | insertion mutation (intron) | ||
| 3510 | - | G | insertion mutation (intron) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weng, W.; Tang, Y.; Xiong, R.; Bai, Q.; Gao, A.; Yao, X.; Wu, W.; Ma, C.; Cheng, J.; Ruan, J. Specific Gibberellin 2-Oxidase 3 (SbGA2ox3) Mutants Promote Yield and Stress Tolerance in Sorghum bicolor. Agronomy 2023, 13, 908. https://doi.org/10.3390/agronomy13030908
Weng W, Tang Y, Xiong R, Bai Q, Gao A, Yao X, Wu W, Ma C, Cheng J, Ruan J. Specific Gibberellin 2-Oxidase 3 (SbGA2ox3) Mutants Promote Yield and Stress Tolerance in Sorghum bicolor. Agronomy. 2023; 13(3):908. https://doi.org/10.3390/agronomy13030908
Chicago/Turabian StyleWeng, Wenfeng, Yong Tang, Ruiqi Xiong, Qing Bai, Anjing Gao, Xin Yao, Weijiao Wu, Chao Ma, Jianping Cheng, and Jingjun Ruan. 2023. "Specific Gibberellin 2-Oxidase 3 (SbGA2ox3) Mutants Promote Yield and Stress Tolerance in Sorghum bicolor" Agronomy 13, no. 3: 908. https://doi.org/10.3390/agronomy13030908