
Differences in Soil Microbiota of Continuous Cultivation of Ganoderma leucocontextum
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cultivation
2.2. Collection of Soil Samples
2.3. Soil Physicochemical Properties
2.4. The Number of Cultivatable Microorganisms
2.5. Sequencing for Microbial Diversity Analysis
2.6. Bioinformatics Analysis
2.7. Yield Measurement
2.8. Statistical Analysis
3. Results
3.1. Comparison of Soil Physicochemical Properties
3.2. The Number of Culturable Microorganisms
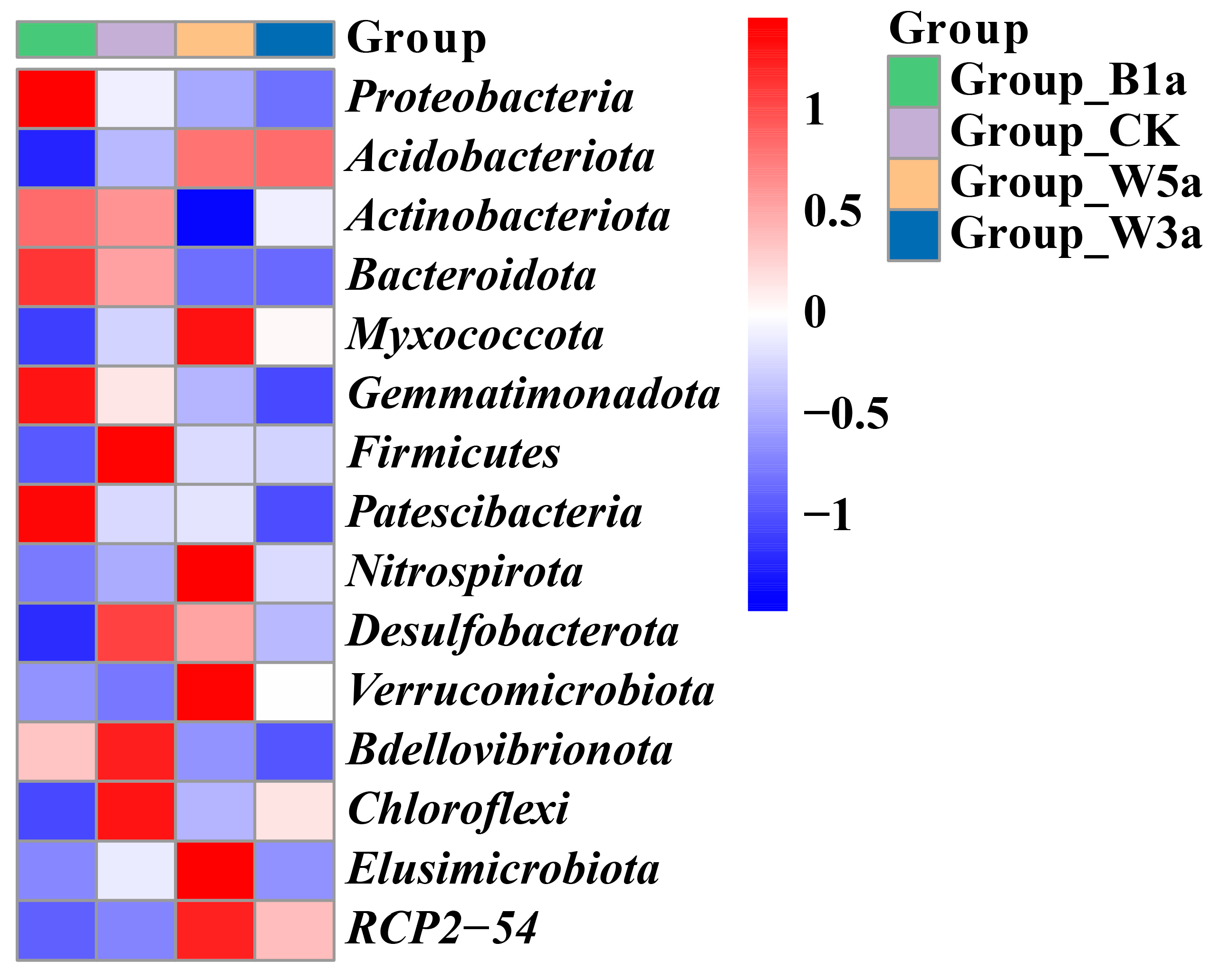
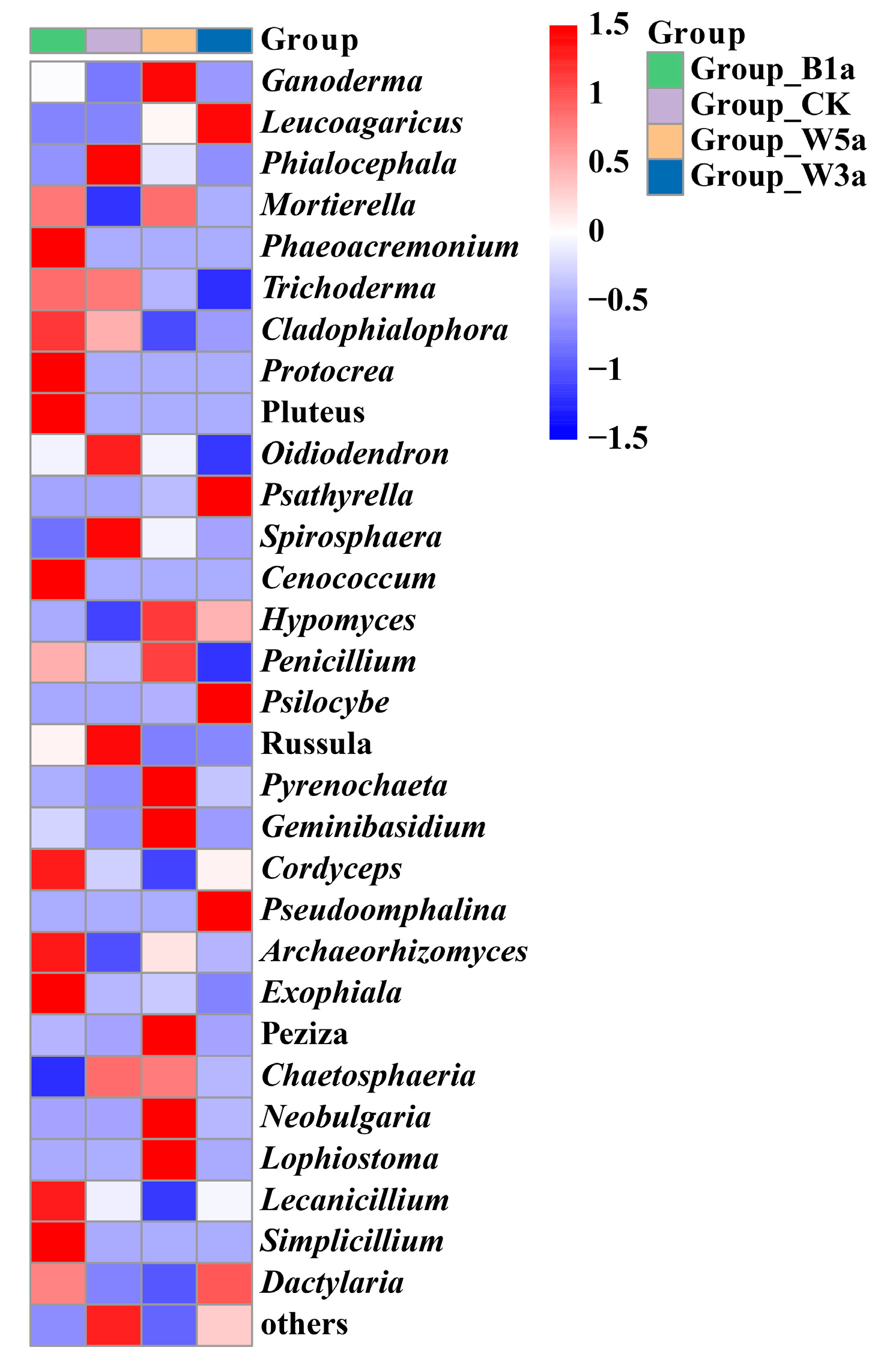
3.3. Microbial Community Structure
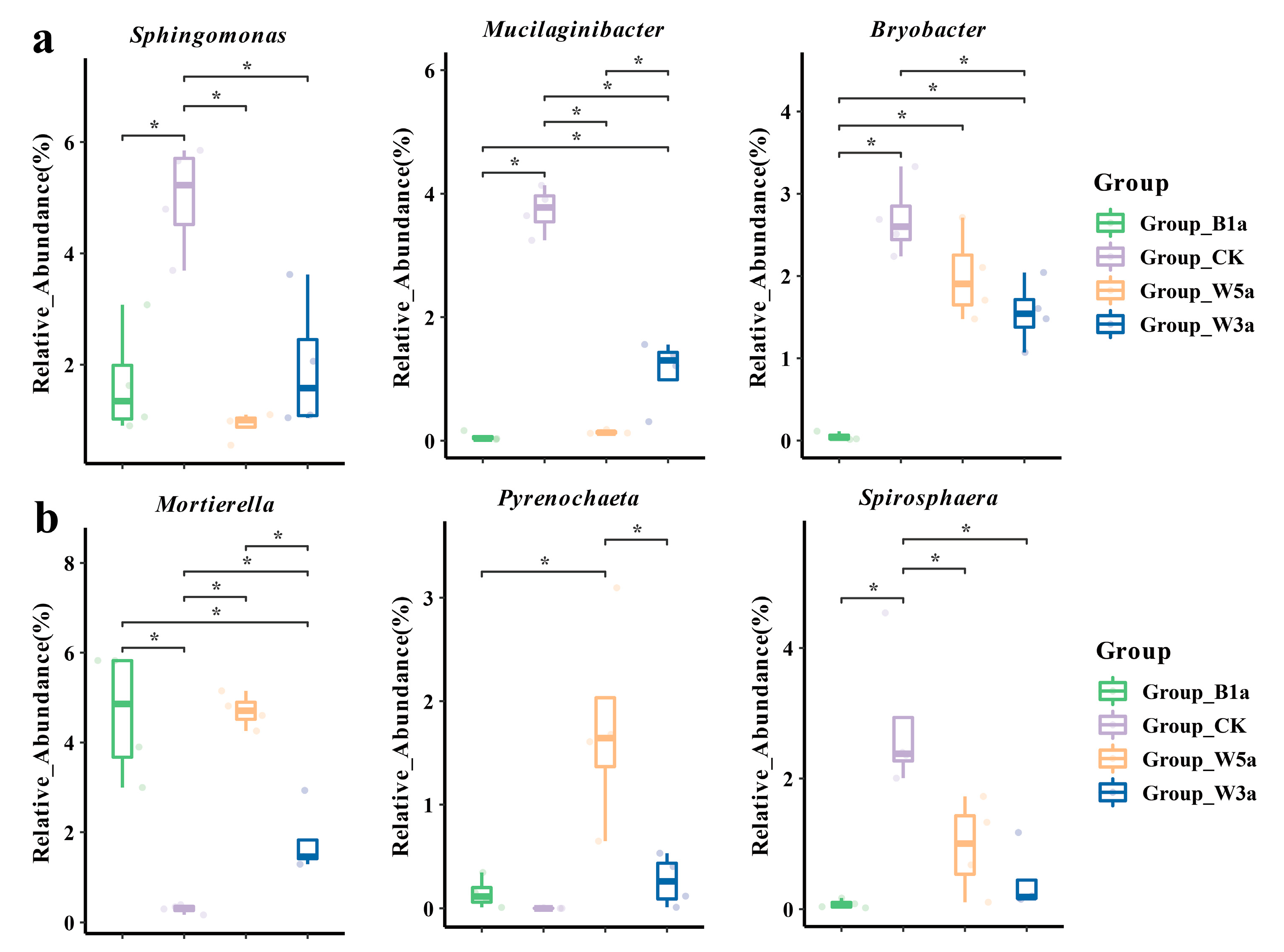
3.3.1. Bacterial Community Composition
3.3.2. Fungal Community Composition
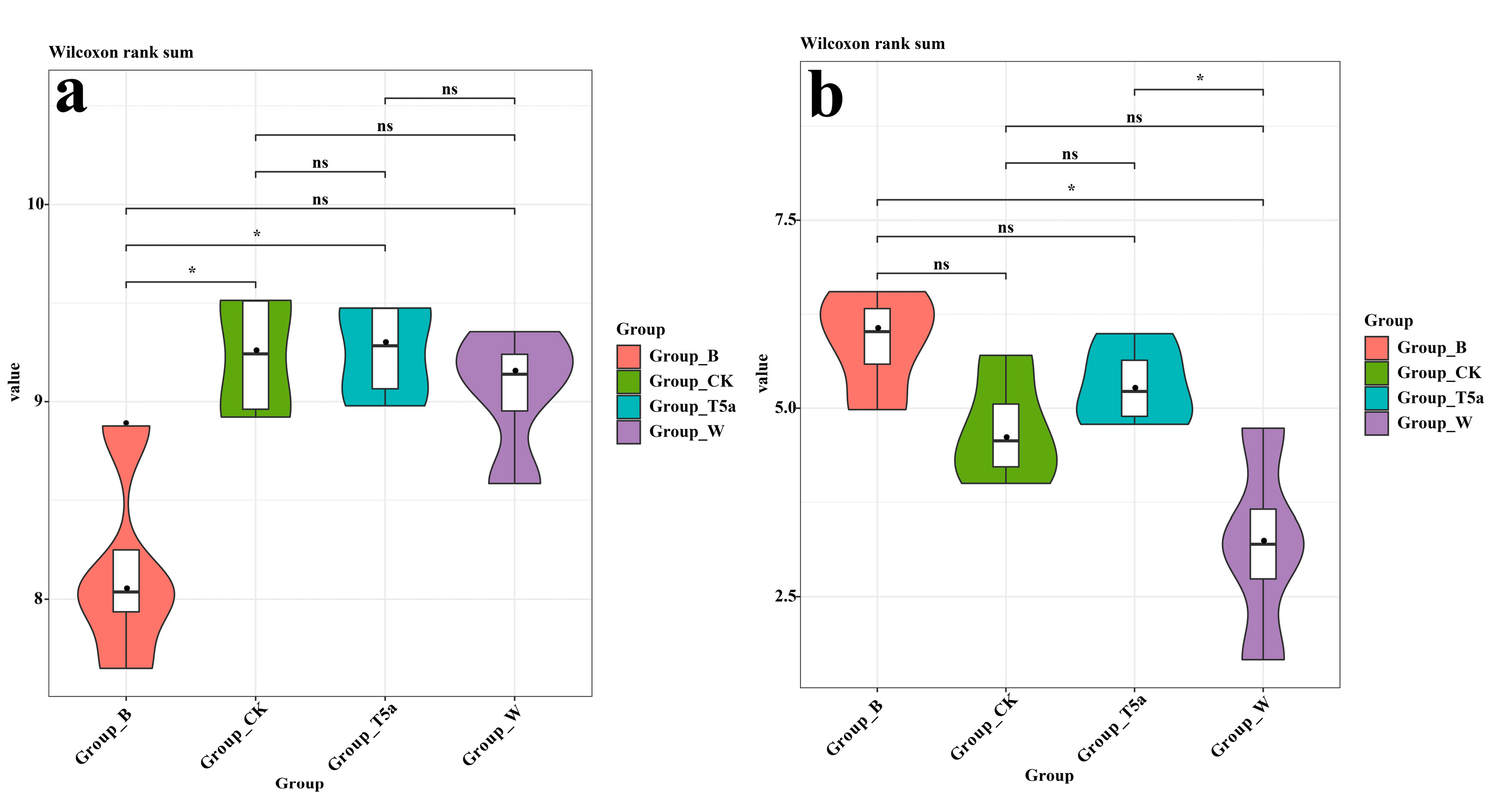
3.4. Microbial Alpha Diversity Analysis
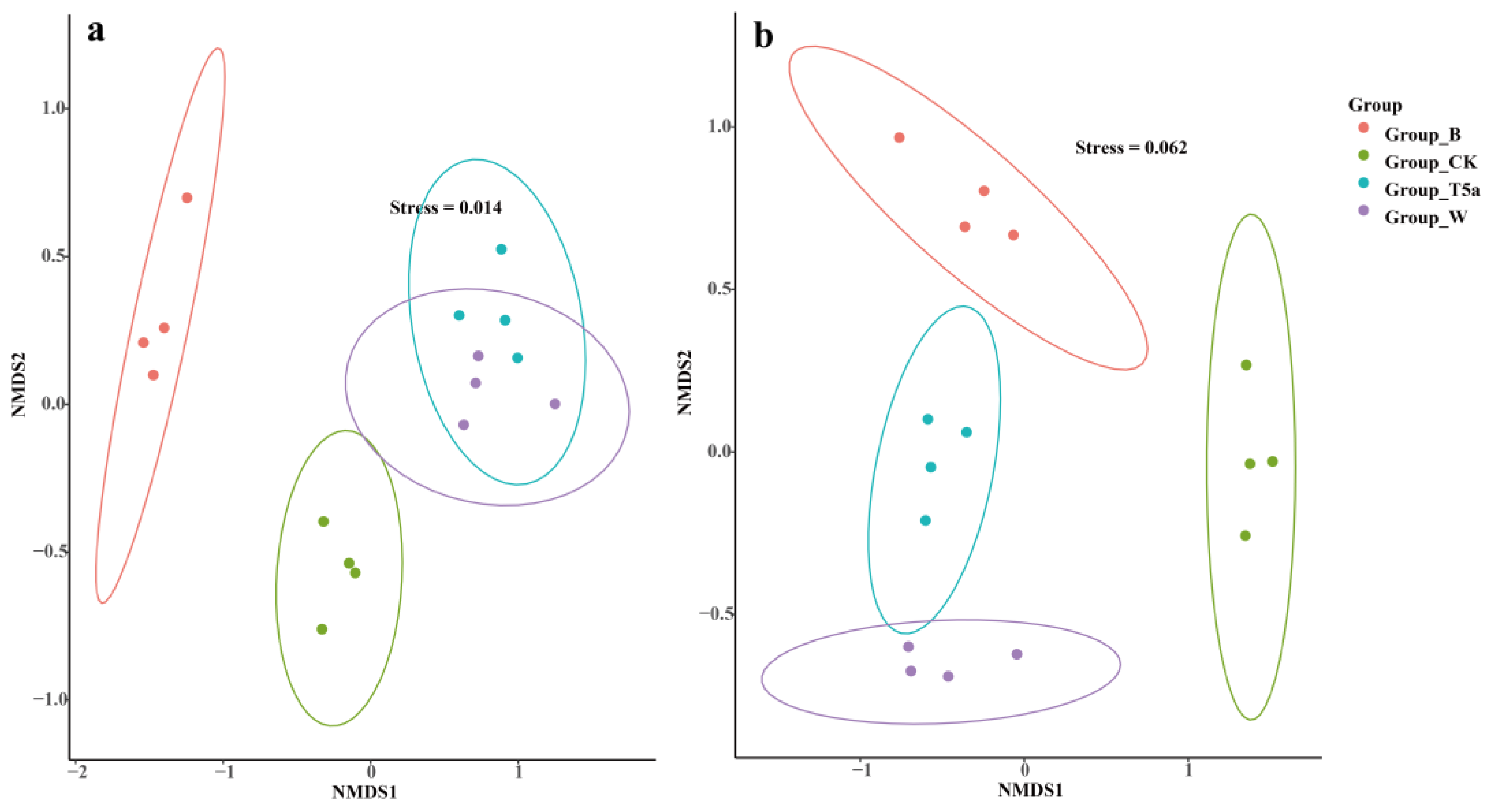
3.5. Microbial Beta Diversity Analysis
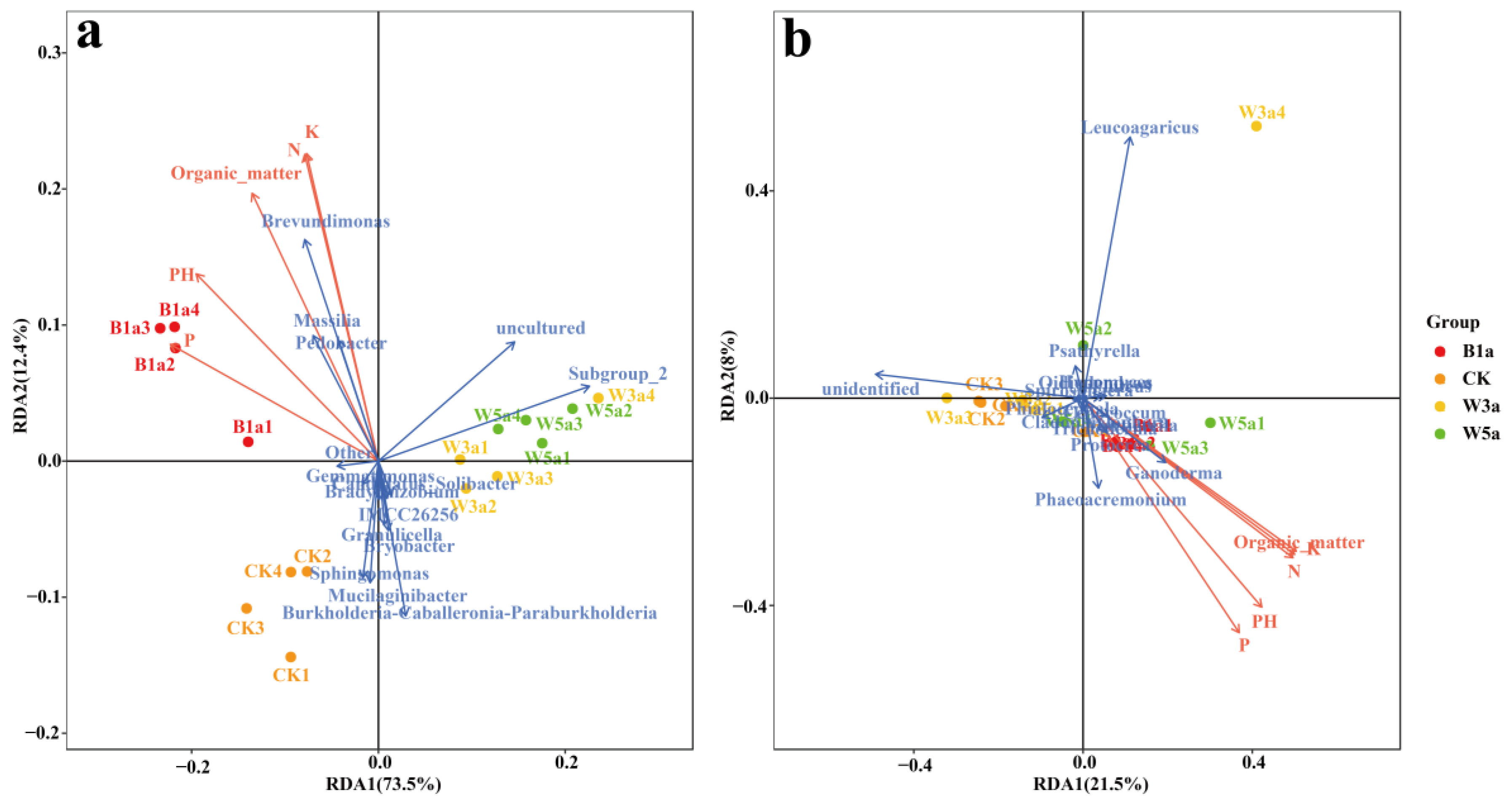
3.6. Multivariate Analysis of Microorganisms
3.7. Correlation between Microbial Flora and Soil Physicochemistry
3.8. Effects on Yield with Continuous Cultivated Soil
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Li, T.H.; Hu, H.P.; Deng, W.Q.; Wu, S.H.; Wang, D.M.; Tsering, T. Ganoderma leucocontextum, a new member of the G. lucidum complex from Southwestern China. Mycoscience 2015, 56, 81–85. [Google Scholar] [CrossRef]
- Wu, F.; Zhou, L.W.; Yang, Z.L.; Yang, Z.L.; Bau, T.; Li, T.H.; Dai, Y.C. Resource diversity of Chinese macrofungi: Edible, medicinal and poisonous species. Fungal Divers 2019, 98, 1–76. [Google Scholar] [CrossRef]
- Yao, C.X.; Tian, G.T.; Wang, H. Three new cultivars of Ganoderma leucocontextum. Mycosystema 2022, 41, 2095–2097. [Google Scholar] [CrossRef]
- Chen, H.Y.; Zhang, J.J.; Ren, J.W.; Wang, W.Z.; Xiong, W.P.; Zhang, Y.D.; Bao, L.; Liu, H.M. Triterpenes and meroterpenes with neuroprotective effects from Ganoderma leucocontextum. Chem. Biodivers. 2018, 15, 177. [Google Scholar] [CrossRef]
- Zhang, J.J.; Ma, K.; Han, J.J.; Wang, K.; Chen, H.Y.; Bao, L.; Liu, L.; Xiong, W.P.; Zhang, Y.D.; Huang, Y.; et al. Eight new triterpenoids with inhibitory activity against HMG -CoA reductase from the medical mushroom Ganoderma leucocontextum collected in Tibetan plateau. Fitoterapia 2018, 130, 79–88. [Google Scholar] [CrossRef]
- Wang, K.; Bao, L.; Xiong, W.P.; Ma, K.; Han, J.J.; Wang, W.Z.; Yin, W.B.; Liu, H.M. Lanostane triterpenes from the Tibetan medicinal mushroom Ganoderma leucocontextum and their inhibitory effects on HMG-CoA reductase and α-glucosidase. J. Nat. Prod. 2015, 78, 1977–1989. [Google Scholar] [CrossRef]
- Van Griendsven, L.J.L.D. Science and Cultivation of Edible Fungi 2000; Taylor and Francis: London, UK, 2012; ISBN 9781466539075/9058091457/978-9058091451. [Google Scholar]
- Buckley, D.H.; Schmidt, T.M. The structure of microbial communities in soil and the lasting impact of cultivation. Microb. Ecol. 2001, 42, 11–21. [Google Scholar] [CrossRef]
- Yao, C.X.; Liang, M.T.; Ma, Y.H.; Liu, J.X.; Zhang, S.S.; Chen, X.; Tian, G.T. Isolation and Identification of Soil-borne Pathogenic Microorganisms in Ganoderma leucocontextum. S.W. Chin. J. Agric. Sci. 2022, 35, 2811–2818. [Google Scholar] [CrossRef]
- Longley, R.; Benucci, G.M.N.; Mills, G.; Bonito, G. Fungal and bacterial community dynamics in substrates during the cultivation of morels (Morchella rufobrunnea) indoors. FEMS Microbiol. Lett. 2019, 366, fnz215. [Google Scholar] [CrossRef]
- Carrasco, J.; Preston, G.M. Growing edible mushrooms: A conversation between bacteria and fungi. Environ. Microbiol. 2020, 22, 858–872. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Wang, Y.L.; Zhang, P.; Yu, F.Q.; Jesús, P. Macrofungi cultivation inshady forest areas signifificantly increases microbiome diversity, abundance and functional capacity in soil furrows. J. Fungi 2021, 7, 775. [Google Scholar] [CrossRef]
- Kang, H.; Chang, W.; Yun, S.; Lee, Y. Roles of ascospores and arthroconidia of Xylogone ganodermophthora in development of yellow rot in cultivated mushroom, Ganoderma lucidum. Plant Pathol. J. 2011, 27, 138–147. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.M.; Chen, D.X.; Chen, Y.G. Alelopathic effect of dominant microflora on its mycelium of continuous cropping obstacles Ganoderma lucidum in field cultivation. Chin. J. Trop. Crops 2016, 37, 372–375. [Google Scholar] [CrossRef]
- Yuan, Y.; Li, L.; Huang, H.C.; Liu, G.H.; Xie, F.Q.; Fu, J.S.; Wu, X.P. Analysis of bacterial community in Ganoderma lingzhi continuous cropping soil based on 16S rDNA amplicon sequencing. Chin. Agric. Sci. Bull. 2021, 37, 116–123. [Google Scholar] [CrossRef]
- Yuan, Y.; Huang, H.C.; Ye, L.Y.; Fu, J.S.; Wu, X.P. Analysis of fungal community in continuous cropping soil of Ganoderma lingzhi. Mycosystema 2019, 38, 2112–2121. [Google Scholar] [CrossRef]
- Ren, F.; Zhang, Y.G.; Yu, H.; Zhang, Y.A. Ganoderma lucidum cultivation affect microbial community structure of soil, wood segments and tree roots. Sci. Rep. 2020, 10, 3435. [Google Scholar] [CrossRef] [Green Version]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Mulvaney, R.L.; Khan, S.A. Diffusion methods to determine different forms of nitrogen in soil hydrolysates. Soil Sci. Soc. Am. J. 2001, 65, 1284–1292. [Google Scholar] [CrossRef]
- Truog, E. The determination of the readily availably phosphorus soils. J. Am. Soc. Agron. 1930, 22, 874. [Google Scholar] [CrossRef] [Green Version]
- Ieggli, C.V.S.; Bohrer, D.; Do Nascimento, P.C.; De Carvalho, L.M. Determination of sodium, potassium, calcium, magnesium, zinc and iron in emulsified chocolate samples by flame atomic absorption spectrometry. Food Chem. 2011, 124, 1189–1193. [Google Scholar] [CrossRef]
- He, J.Z.; Zheng, Y.; Chen, C.R.; He, Y.Q.; Zhang, L.M. Microbial composition and diversity of an upland red soil under long-term fertilization treatments as revealed by culture-dependent and culture-independent approaches. J. Soils Sediments 2008, 8, 349–358. [Google Scholar] [CrossRef]
- Magoc, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Caporaso, J.G. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Meth. 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Li, Q.; Xiang, P.; Zhang, T.; Wu, Q.; Bao, Z.; Tu, W.; Li, L.; Zhao, C. The effect of phosphate mining activities on rhizosphere bacterial communities of surrounding vegetables and crops. Sci. Total Environ. 2022, 821, 153479. [Google Scholar] [CrossRef]
- Li, Q.; Wu, Q.; Zhang, T.; Xiang, P.; Bao, Z.; Tu, W.; Li, L.; Wang, Q. Phosphate mining activities affect crop rhizosphere fungal communities. Sci. Total Environment. 2022, 838 Pt 2, 156196. [Google Scholar] [CrossRef]
- Sudheer, S.; Alzorqi, I.; Ali, A.; Cheng, P.G.; Siddiqui, Y.; Manickam, S. Determination of the biological efficiency and antioxidant potential of Lingzhi or Reishi medicinal mushroom, Ganoderma lucidum (Agaricomycetes), cultivated using different agro-wastes in Malaysia. Int. J. Med. Mushrooms 2018, 20, 89–100. [Google Scholar] [CrossRef]
- Gea, F.J.; Navarro, M.J.; Santos, M.; Diánez, F.; Carrasco, J. Control of Fungal Diseases in Mushroom Crops while Dealing with Fungicide Resistance: A Review. Microorganisms 2021, 9, 585. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Fan, T.; Jin, L.; Lei, P.; Shao, C.; Wu, S.; Yang, Y.; He, Y.; Ren, R.; Xu, J. Soil microbial diversity and functional capacity associated with the production of edible mushroom Stropharia rugosoannulata in croplands. PeerJ 2022, 10, e14130. [Google Scholar] [CrossRef]
- Lauber, C.L.; Ramirez, K.S.; Aanderud, Z.; Lennon, J.; Fierer, N. Temporal variability in soil microbial communities across landuse types. ISME J. 2013, 7, 1641–1650. [Google Scholar] [CrossRef]
- Bulgarelli, D.; Rott, M.; Schlaeppi, K.; Themaat, E.V.L.V.; Ahmadinejad, N.; Assenza, F.; Schulze-Lefert, P. Revealing structure and assembly cues for Arabidopsis root-inhabiting bacterial microbiota. Nature 2012, 488, 91–95. [Google Scholar] [CrossRef]
- Muller, D.B.; Vogel, C.; Bai, Y.; Vorholt, J.A. The plant microbiota: Systems-level insights and perspectives. Annu. Rev. Genet. 2016, 50, 211–234. [Google Scholar] [CrossRef] [Green Version]
- Frindte, K.; Pape, R.; Werner, K.; Löfer, J.; Knief, C. Temperature and soil moisture control microbial community composition in an arctic-alpine ecosystem along elevational and micro-topographic gradients. ISME J. 2019, 13, 2031–2043. [Google Scholar] [CrossRef]
- Plassart, P.; Prévost-Bouré, N.C.; Uroz, S.; Dequiedt, S.; Stone, D.; Creamer, R.; Lemanceau, P. Soil parameters, land use, and geographical distance drive soil bacterial communities along a European transect. Sci. Rep. 2019, 9, e605. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Baquerizo, M.; Oliverio, A.M.; Brewer, T.E.; Benavent-González, A.; Eldridge, D.J.; Bardgett, R.D.; Fierer, N. A global atlas of the dominant bacteria found in soil. Science 2018, 359, 320–325. [Google Scholar] [CrossRef] [Green Version]
- Wei, Z.; Gu, Y.; Friman, V.P.; Kowalchuk, G.A.; Xu, Y.C.; Shen, Q.R.; Jousset, A. Initial soil microbiome composition and functioning predetermine future plant health. Sci. Adv. 2019, 5, eaaw0759. [Google Scholar] [CrossRef] [Green Version]
- Goh, Y.K.; Zoqratt, M.Z.H.M.; Goh, Y.K.; Ayub, Q.; Ting, A.S.Y. Determining soil microbial communities and their influence on Ganoderma disease incidences in oil palm (Elaeis guineensis) via high-throughput sequencing. Biology 2020, 9, 424. [Google Scholar] [CrossRef]
- Wang, T.T.; Hao, Y.W.; Zhu, M.Z.; Yu, S.T.; Ran, W.; Xue, C.; Ling, N.; Shen, Q.R. Characterizing diferences in microbial community composition and function between Fusarium wilt diseased and healthy soils under watermelon cultivation. Plant Soil 2019, 438, 421–433. [Google Scholar] [CrossRef]
- Tan, Y.; Cui, Y.S.; Li, H.Y.; Kuang, A.X.; Li, X.R.; Wei, Y.L.; Ji, X.L. Rhizospheric soil and root endogenous fungal diversity and composition in response to continuous Panaxnoto ginseng cropping practices. Microbiol. Res. 2016, 194, 10–19. [Google Scholar] [CrossRef]
- Bao, Z.; Feng, H.; Tu, W.; Li, L.; Li, Q. Method and mechanism of chromium removal from soil: A systematic review. Environ. Sci. Pollut. Res. 2022, 29, 35501–35517. [Google Scholar] [CrossRef]
- Feng, Y.; Yuan, Q.; Yang, Z.; Zhang, T. Effect of Sphingomonas sp. strain on degradation of polyphenols in redried tobacco leaves. Acta Tab. Sin. 2019, 25, 19–24. [Google Scholar] [CrossRef]
- Liu, J.; Lin, H.; Dong, Y.; Li, B. Elucidating the biodegradation mechanism of tributyl phosphate (TBP) by Sphingomonas sp. isolated from TBP-contaminated mine-tailings. Environ. Pollut. 2019, 250, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Huq, M.A.; Akter, S.; Lee, S.Y. Mucilaginibacter formosus sp. nov., a bacterium isolated from road-side soil. Antonie Van Leeuwenhoek 2019, 112, 513–521. [Google Scholar] [CrossRef]
- Gadkari, P.; Haggblom, M.; Barkay, T.; Kerkhof, L.; Mannisto, M. Ecology and Physiological Potential of Tundra Soil Bacteria; Rutgers University, School of Graduate Studies: New Brunswick, NJ, USA, 2019; pp. 1–31. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Komaitis, M.; Aggelis, G. Single cell oil (SCO) production by Mortierella isabellina grown on high-sugar content media. Bioresour. Technol. 2004, 95, 287–291. [Google Scholar] [CrossRef]
- Rousk, J.; Brookes, P.C.; Bååth, E. Contrasting soil pH effects on fungal and bacterial growth suggest functional redundancy in carbon mineralization. Appl. Environ. Microbiol. 2009, 75, 1589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, Z.; Wang, X.; Wang, Q.; Zou, L.; Peng, L.; Li, L.; Tu, W.; Li, Q. A novel method of domestication combined with ARTP to improve the reduction ability of Bacillus velezensis to Cr(VI). J. Environ. Chem. Eng. 2023, 11, 109091. [Google Scholar] [CrossRef]
- Rahman, K.; Othman, R. Influence of pH levels on disease development in oil palm seedling roots infected with Ganoderma boninense. Rhizosphere 2020, 13, 100181. [Google Scholar] [CrossRef]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Li, Q.; Xiang, P.; Li, L.; Zhang, T.; Wu, Q.; Bao, Z.; Tu, W. Phosphorus mining activities alter endophytic bacterial communities and metabolic functions of surrounding vegetables and crops. Plant Soil 2023, 485, 1–20. [Google Scholar] [CrossRef]
- Ke, L.Q.; Li, P.D.; Xu, J.P.; Wang, Q.S.; Wang, L.L.; Wen, H.P. Microbial communities and soil chemical features associated with commercial production of the medicinal mushroom Ganoderma lingzhi in soil. Sci. Rep. 2019, 9, 15839. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Information | Sample ID | Sample Group |
|---|---|---|
| Medium bag cultivation ① soil of Group Ⅰ (after 1 year) | B1a.1 | B1a |
| Medium bag cultivation ② soil of Group Ⅰ (after 1 year) | B1a.2 | |
| Medium bag cultivation ③ soil of Group Ⅰ (after 1 year) | B1a.3 | |
| Medium bag cultivation ④ soil of Group Ⅰ (after 1 year) | B1a.4 | |
| CK ① uncultivated soil | CK.1 | CK |
| CK ② uncultivated soil | CK.2 | |
| CK ③ uncultivated soil | CK.3 | |
| CK ④ uncultivated soil | CK.4 | |
| Log cultivation ① Group Ⅱ (after 3 year) | W3a.1 | W3a |
| Log cultivation ② Group Ⅱ (after 3 year) | W3a.2 | |
| Log cultivation ③ Group Ⅱ (after 3 year) | W3a.3 | |
| Log cultivation ④ Group Ⅱ (after 3 year) | W3a.4 | |
| Log cultivation ① Group Ⅲ (after 5 year) | W5a.1 | W5a |
| Log cultivation ② Group Ⅲ (after 5 year) | W5a.2 | |
| Log cultivation ③ Group Ⅲ (after 5 year) | W5a.3 | |
| Log cultivation ④ Group Ⅲ (after 5 year) | W5a.4 |
| Treatment | pH | OM (g/kg) | N (mg/kg) | P (mg/kg) | K (mg/kg) |
|---|---|---|---|---|---|
| B1a | 8.0 a | 19.8 a | 44 a | 19.2 a | 973 a |
| CK | 5.5 b | 5.3 c | 12 c | 13.2 b | 189 c |
| W3a | 5.1 b | 10.6 b | 29 b | 10.5 b | 437 b |
| W5a | 5.4 b | 8.3 b | 26 b | 10.2 b | 638 b |
| Treatment | Total Count | Bacterium Count | Actinomycete Count | Fungus Count | ||||
|---|---|---|---|---|---|---|---|---|
| (104 CFU/g) | % | (104 CFU/g) | % | (104 CFU/g) | % | (104 CFU/g) | % | |
| B1a | 1553.23 a | 212.05 | 842.81 a | 571.56 | 697.15 a | 89.03 | 13.276 a | 284.59 |
| CK | 497.75 b | / | 125.50 b | / | 368.80 b | / | 3.452 b | / |
| W3a | 174.33 c | −64.98 | 87.38 c | −30.37 | 84.43 c | −77.11 | 2.513 b | −27.20 |
| W5a | 63.35 c | −87.27 | 60.37 c | −51.90 | * | * | 2.985 b | −13.53 |
| Yield | Medium Bag | Log | |||
|---|---|---|---|---|---|
| B-CK | B1a | W-CK | W3a | W5a | |
| fresh mushroom (g/bag·log) | 148.8 ± 12.37 | 91.2 ± 7.91 | 294.1 ± 25.32 | 108.3 ± 8.14 | 25.5 ± 2.13 |
| dry mushroom (g/bag·log) | 41.2 ± 5.67 | 25.3 ± 3.79 | 108.9 ± 9.11 | 40.0 ± 6.29 | 9.4 ± 1.71 |
| biological efficiency (%) | 24.8% | 15.2% | 9.8% | 3.6% | 0.85% |
| conversion rate (%) | 6.87% | 4.22% | 3.63% | 1.33% | 0.31% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, C.; Tao, N.; Liu, J.; Liang, M.; Wang, H.; Tian, G. Differences in Soil Microbiota of Continuous Cultivation of Ganoderma leucocontextum. Agronomy 2023, 13, 888. https://doi.org/10.3390/agronomy13030888
Yao C, Tao N, Liu J, Liang M, Wang H, Tian G. Differences in Soil Microbiota of Continuous Cultivation of Ganoderma leucocontextum. Agronomy. 2023; 13(3):888. https://doi.org/10.3390/agronomy13030888
Chicago/Turabian StyleYao, Chunxin, Nan Tao, Jiaxun Liu, Mingtai Liang, Hui Wang, and Guoting Tian. 2023. "Differences in Soil Microbiota of Continuous Cultivation of Ganoderma leucocontextum" Agronomy 13, no. 3: 888. https://doi.org/10.3390/agronomy13030888