
Rhizosphere Microbiomes of Amaranthus spp. Grown in Soils with Anthropogenic Polyelemental Anomalies
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil Sampling
2.2. Soil Characterization
2.3. Experimental Design
2.4. A 16S rRNA Gene-Based Metagenomic Analysis of Rhizosphere Soil
2.5. Bioinformatics and Statistical Analysis
3. Results
3.1. Soil Characteristics
3.2. Metagenomic Analysis of Rhizosphere Microbial Communities of Amaranthus spp.
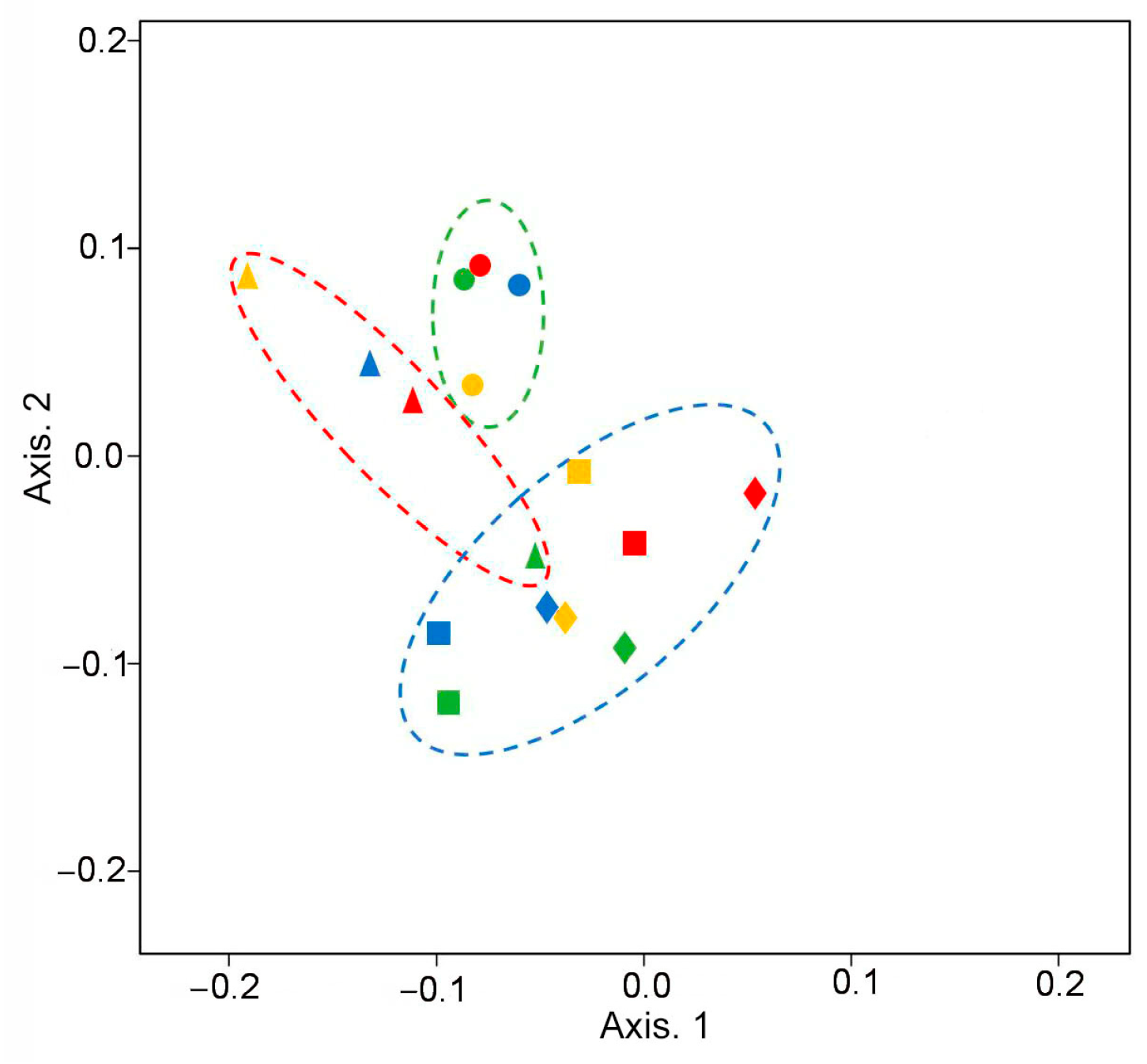
3.2.1. Diversity of Rhizosphere Communities
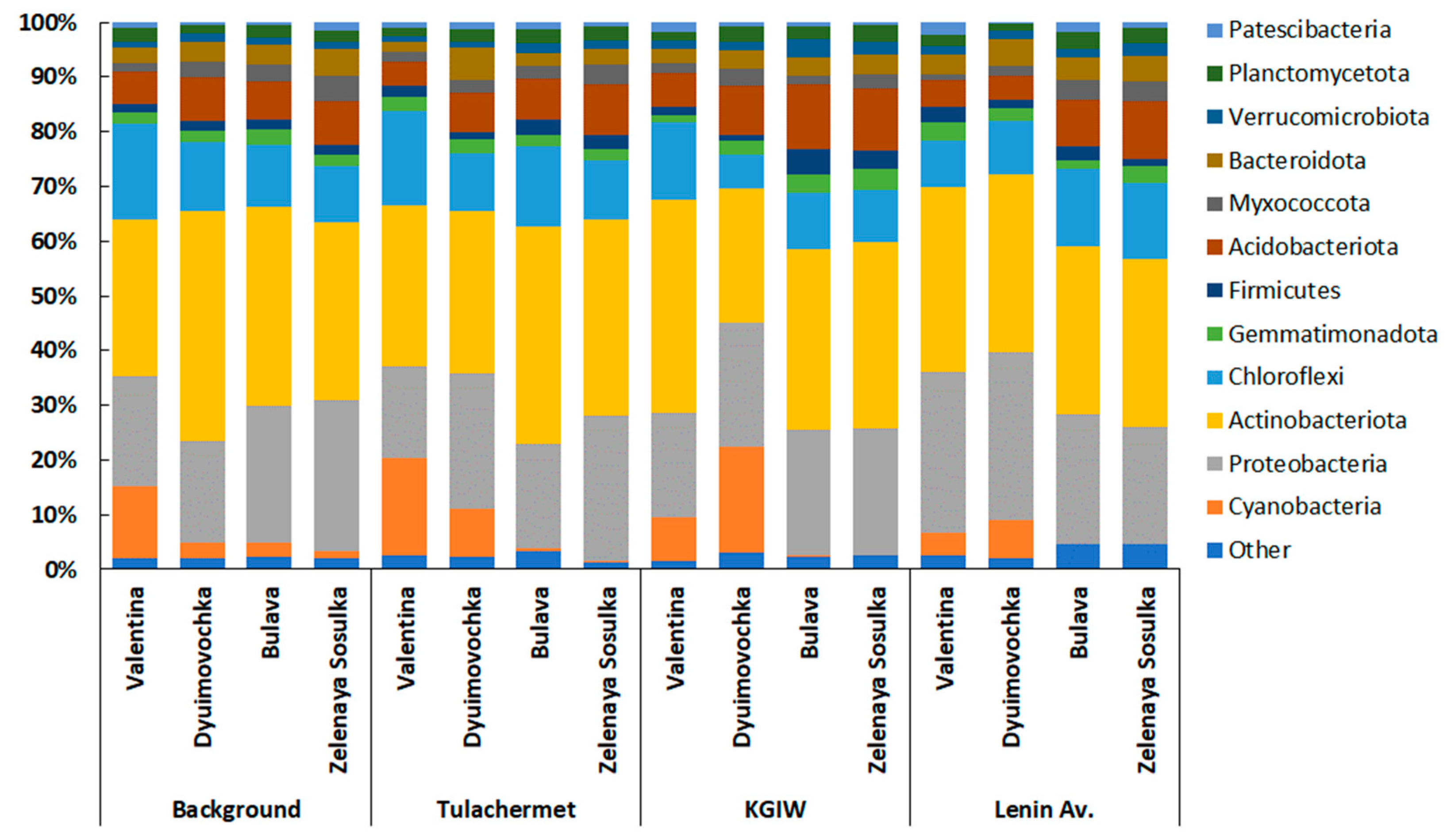
3.2.2. Taxonomic Structure of Rhizosphere Communities
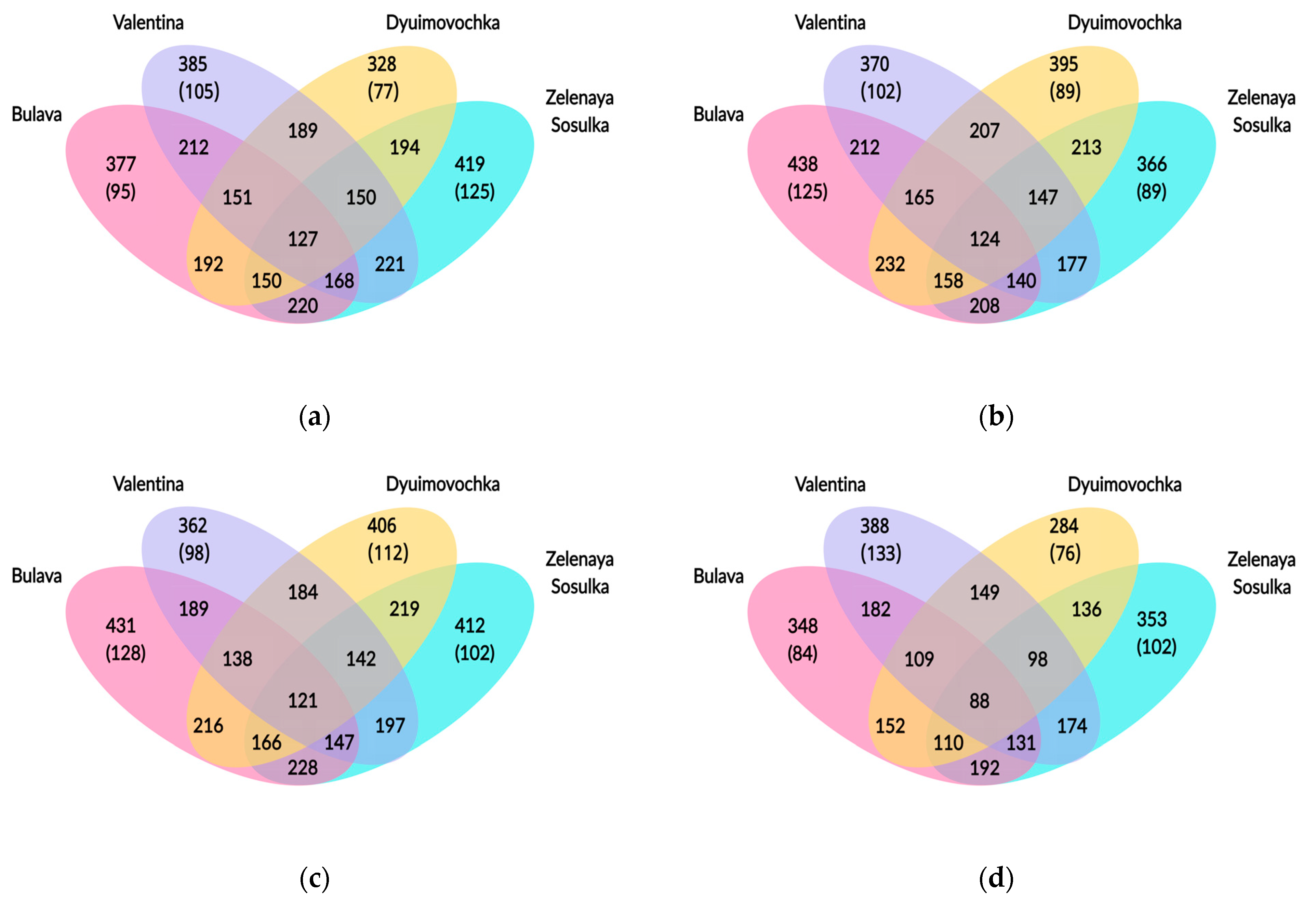
3.2.3. Shared and Unique Taxa among Rhizosphere Microbial Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bais, H.P.; Broeckling, C.D.; Vivanco, J.M. Root exudates modulate plant–microbe interactions in the rhizosphere. In Secondary Metabolites in Soil Ecology. Soil Biology; Karlovsky, P., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; Volume 14, pp. 241–252. [Google Scholar]
- Fan, D.; Schwinghamer, T.; Smith, D.L. Isolation and diversity of culturable rhizobacteria associated with economically important crops and uncultivated plants in Québec, Canada. Syst. Appl. Microbiol. 2018, 41, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Mohanram, S.; Kumar, P. Rhizosphere microbiome: Revisiting the synergy of plant-microbe interactions. Ann. Microbiol. 2019, 69, 307–320. [Google Scholar] [CrossRef]
- Srivastava, R.; Kanda, T.; Yadav, S.; Mishra, R.; Atri, N. Cyanobacteria in rhizosphere: Dynamics, diversity, and symbiosis. In Plant, Soil and Microbes in Tropical Ecosystems. Rhizosphere Biology; Dubey, S.K., Verma, S.K., Eds.; Springer: Singapore, 2021; pp. 51–69. [Google Scholar]
- Moreno-Espíndola, I.P.; Ferrara-Guerrero, M.J.; de León-González, F.; Rivera-Becerril, F.; Mayorga-Reyes, L.; Pérez, N.O. Enzymatic activity and culturable bacteria diversity in rhizosphere of amaranth, as indicators of crop phenological changes. Bot. Sci. 2018, 96, 640–649. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Dubey, A. Rhizosphere microbiome: Engineering bacterial competitiveness for enhancing crop production. J. Adv. Res. 2020, 24, 337–352. [Google Scholar] [CrossRef] [PubMed]
- Castellano-Hinojosa, A.; Strauss, S.L. Insights into the taxonomic and functional characterization of agricultural crop core rhizobiomes and their potential microbial drivers. Sci. Rep. 2021, 11, 10068. [Google Scholar] [CrossRef]
- Moorthy, A.; Balasundaram, U. Rhizosphere metagenomics: Methods and challenges. In Omics Science for Rhizosphere Biology, Rhizosphere Biology; Pudake, R.N., Sahu, B.B., Kumari, M., Sharma, A.K., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2021; pp. 1–20. [Google Scholar]
- Ke, J.; Wang, B.; Yoshikuni, Y. Microbiome engineering: Synthetic biology of plant-associated microbiomes in sustainable agriculture. Trends Biotechnol. 2021, 39, 244–261. [Google Scholar] [CrossRef]
- Ofek-Lalzar, M.; Sela, N.; Goldman-Voronov, M.; Green, S.J.; Hadar, Y.; Minz, D. Niche and host-associated functional signatures of the root surface microbiome. Nat. Commun. 2014, 5, 4950. [Google Scholar] [CrossRef] [Green Version]
- Schreiter, S.; Ding, G.-C.; Heuer, H.; Neumann, G.; Sandmann, M.; Grosch, R.; Kropf, S.; Smalla, K. Effect of the soil type on the microbiome in the rhizosphere of field-grown lettuce. Front. Microbiol. 2014, 5, 144. [Google Scholar] [CrossRef]
- Sun, X.; Song, B.; Xu, R.; Zhang, M.; Gao, P.; Lin, H.; Sun, W. Root-associated (rhizosphere and endosphere) microbiomes of the Miscanthus sinensis and their response to the heavy metal contamination. J. Environ. Sci. 2021, 104, 387–398. [Google Scholar] [CrossRef]
- Neu, A.T.; Allen, E.E.; Roy, K. Defining and quantifying the core microbiome: Challenges and prospects. Proc. Natl. Acad. Sci. USA 2021, 118, e2104429118. [Google Scholar] [CrossRef]
- Chen, Y.; Ding, Q.; Chao, Y.; Wei, X.; Wang, S.; Qiu, R. Structural development and assembly patterns of the root-associated microbiomes during phytoremediation. Sci. Tot. Environ. 2018, 644, 1591–1601. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, T.H.; Cloutier-Hurteau, B.; Al-Otaibi, F.; Turmel, M.-C.; Yergeau, E.; Courchesne, F.; St-Arnaud, M. Early rhizosphere microbiome composition is related to the growth and Zn uptake of willows introduced to a former landfill. Environ. Microbiol. 2015, 17, 3025–3038. [Google Scholar] [CrossRef] [PubMed]
- Kour, D.; Rana, K.L.; Yadav, N.; Yadav, A.N.; Kumar, A.; Meena, V.S.; Singh, B.; Chauhan, V.S.; Dhaliwal, H.S.; Saxena, A.K. Rhizospheric microbiomes: Biodiversity, mechanisms of plant growth promotion, and biotechnological applications for sustainable agriculture. In Plant Growth Promoting Rhizobacteria for Agricultural Sustainability (From Theory to Practices); Kumar, A., Meena, V.S., Eds.; Springer Nature Singapore Pte Ltd.: Singapore, 2019; pp. 19–65. [Google Scholar]
- Barra Caracciolo, A.; Terenzi, V. Rhizosphere microbial communities and heavy metals. Microorganisms 2021, 9, 1462. [Google Scholar] [CrossRef]
- Lundberg, D.S.; Lebeis, S.L.; Paredes, S.H.; Yourstone, S.; Gehring, J.; Malfatti, S.; Tremblay, J.; Engelbrektson, A.; Kunin, V.; del Rio, T.G.; et al. Defining the core Arabidopsis thaliana root microbiome. Nature 2012, 488, 86–90. [Google Scholar] [CrossRef] [Green Version]
- Muratova, A.Y.; Gorelova, S.V.; Sungurtseva, I.Y.; Zelenova, N.A. Rhizospheric microbiomes of Sorghum bicolor grown on soils with anthropogenic polyelement anomalies. In BIO Web of Conferences PLAMIC2020; EDP Sciences: Les Ulis, France, 2020; Volume 23, p. 03008. [Google Scholar]
- Gins, M.S. Biologicheski Aktivnyye Veshchestva Amaranta Amarantin: Svoystva, Mekhanizmy Deystviya i Prakticheskoye Ispol’zovaniye (Biologically Active Substances of Amaranth Amaranthine: Properties, Mechanisms of Action and Practical Use); RUDN: Moscow, Russia, 2002; 183p. (In Russian) [Google Scholar]
- Shevyakova, N.I.; Cheremisina, A.I.; Kuznetsov, V.V. Phytoremediation potential of Amaranthus hybrids: Antagonism between nickel and iron and chelating role of polyamines. Russ. J. Plant Physiol. 2011, 58, 634–642. [Google Scholar] [CrossRef]
- Chinmayee, M.D.; Mahesh, B.; Pradesh, S.; Mini, I.; Swapna, T.S. The assessment of phytoremediation potential of invasive weed Amaranthus spinosus L. Appl. Biochem. Biotechnol. 2012, 167, 1550–1559. [Google Scholar] [CrossRef]
- Ziarati, P.; Alaedini, S. The phytoremediation technique for cleaning up contaminated soil by Amaranthus sp. J. Environ. Anal. Toxicol. 2014, 4, 1000208. [Google Scholar]
- Odiyi, B.; Ologundudu, F.A.; Adegbite, T. Phytoremediation potential of Amaranthus hybridus L. (Caryophyllales: Amaranthaceae) on soil amended with brewery effluent. Braz. J. Biol. Sci. 2019, 6, 401–411. [Google Scholar] [CrossRef] [Green Version]
- Gorelova, S.V.; Gins, M.S.; Frontasyeva, M.V. Phytoextraction of toxic elements by Amaranthus tricolor grown on technogenically polluted soils in open ground conditions. Chim. Techno Acta 2022, 9, 202292S8. [Google Scholar] [CrossRef]
- Song, N.; Zhang, X.; Wang, F.; Zhang, C.; Tang, S. Elevated CO2 increases Cs uptake and alters microbial communities and biomass in the rhizosphere of Phytolacca americana Linn (pokeweed) and Amaranthus cruentus L. (purple amaranth) grown on soils spiked with various levels of Cs. J. Environ. Radioact. 2012, 112, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Mishra, J.; Singh, R.; Arora, N.K. Alleviation of heavy metal stress in plants and remediation of soil by rhizosphere microorganisms. Front. Microbiol. 2017, 8, 1706. [Google Scholar] [CrossRef] [Green Version]
- Nambisan, S.; Sunil, M.; Choudhary, B.; Srinivasan, S. Cyanobacteria is uniquely enriched in the roots of grain amaranths. bioRxiv 2019, 540484. [Google Scholar] [CrossRef]
- Gorelova, S.V.; Gorbunov, A.V.; Frontasyeva, M.V.; Sylina, A.K. Toxic elements in the soils of urban ecosystems and technogenic sources of pollution. WSEAS Trans. Environ. Dev. 2020, 16, 608–618. [Google Scholar] [CrossRef]
- GOST 53123-2008; Soil Quality. Sampling. Part 5. Guidance on the Procedure for the Investigation of Urban and Industrial Sites with Regard to Soil Contamination. Russian Gost: Moscow, Russian, 2008. Available online: https://docs.cntd.ru/document/1200074384 (accessed on 1 October 2022).
- GOST 26483-85; Soils. Preparation of Salt Extract and Determination of Its pH by CINAO Method. Russian Gost: Moscow, Russian, 1985. Available online: https://docs.cntd.ru/document/1200023490 (accessed on 21 February 2023).
- Malahov, S.G. Interim Guidelines on Soil Pollution Control. 1984. Available online: https://files.stroyinf.ru/Data2/1/4294847/4294847412.htm (accessed on 21 February 2023).
- GOST 26205-91; Soils. Determination of Mobile Compounds of Phosphorus and Potassium by Machigin Method Modified by CINAO. Russian Gost: Moscow, Russian, 1991. Available online: https://docs.cntd.ru/document/1200023449 (accessed on 21 February 2023).
- GOST 26488-85; Soils. Determination of Nitrates by CINAO Method. Russian Gost: Moscow, Russian, 1985. Available online: https://docs.cntd.ru/document/1200023495 (accessed on 21 February 2023).
- GOST 26489-85; Soils. Determination of Exchangeable Ammonium by CINAO Method. Russian Gost: Moscow, Russian, 1986. Available online: https://docs.cntd.ru/document/1200023496 (accessed on 21 February 2023).
- PND F 16.1:2:2.2:2.3:3.64-10; Quantitative Chemical Analysis of Soils. Method for Measuring the Mass Fraction of Oil Products in Samples of Soils, Soils, Bottom Sediments, Silts, Sewage Sludge, Production and Consumption Wastes by the Gravimetric Method. Federal Service for Environmental, Technological and Nuclear Supervision: Moscow, Russia, 2010. Available online: https://files.stroyinf.ru/Data2/1/4293807/4293807051.htm (accessed on 21 February 2023).
- PND F 16.1.42-04; Measurement Method for Measuring the Mass Fraction of Metals and Metal Oxides in Soil Powder Samples by X-Ray Fluorescence Analysis. Federal Service for Environmental, Technological and Nuclear Supervision: Moscow, Russia, 2004. (In Russian)
- GN 2.1.7.2041-06; Predel’no Dopustimye Koncentracii (PDK) Himicheskih Veshchestv v Pochve: Gigienicheskie Normativy.—M.: Federal’nyj Centr Gigieny i Epidemiologii Rospotrebnadzora. Maximum Permissible Concentrations (MPCs) of Chemicals in the Soil: Moscow, Russia, 2006; 15p. Available online: https://files.stroyinf.ru/Data2/1/4293850/4293850511.pdf (accessed on 21 February 2023). (In Russian).
- GN 2.1.7.2511-09; Orientirovochno Dopustimye Koncentracii (ODK) Himicheskih Veshchestv v Pochve: Gigienicheskie Normativy.—M.: Federal’nyj Centr Gigieny i Epidemiologii Rospotrebnadzora. Approximate allowable concentrations (AAC) of Chemical Substances in Soils: Moscow, Russia, 2009; 10p. Available online: https://files.stroyinf.ru/Data2/1/4293828/4293828439.pdf (accessed on 21 February 2023). (In Russian).
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, P.; Parfrey, L.W.; Yarza, P.; Gerken, J.; Pruesse, E.; Quast, C.; Schweer, T.; Peplies, J.; Ludwig, W.; Glockner, F.O. The SILVA and "All-species Living Tree Project (LTP)" taxonomic frameworks. Nucleic Acids Res. 2014, 42, D643–D648. [Google Scholar] [CrossRef] [Green Version]
- Jing, H.; Xia, X.; Liu, H.; Zhou, Z.; Wu, C.; Nagarajan, S. Anthropogenic impact on diazotrophic diversity in the mangrove rhizosphere revealed by nifH pyrosequencing. Front. Microbiol. 2015, 6, 1172. [Google Scholar] [CrossRef] [Green Version]
- Prasanna, R.; Nain, L.; Ancha, R.; Srikrishna, J.; Joshi, M.; Kaushik, B.D. Rhizosphere dynamics of inoculated cyanobacteria and their growth-promoting role in rice crop. Egypt. J. Biol. 2009, 11, 26–36. [Google Scholar]
- Suresh, A.; Soundararajan, S.; Elavarasi, S.; Oscara, F.L.; Thajuddin, N. Evaluation and characterization of the plant growth promoting potentials of two heterocystous cyanobacteria for improving food grains growth. Biocatal. Agric. Biotechnol. 2019, 17, 647–652. [Google Scholar] [CrossRef]
- Toribio, A.J.; Suárez-Estrella, F.; Jurado, M.M.; López, M.J.; López-González, J.A.; Moreno, J. Prospection of cyanobacteria producing bioactive substances and their application as potential phytostimulating agents. Biotechnol. Rep. 2020, 26, e00449. [Google Scholar] [CrossRef] [PubMed]
- Lindblad, P. Cyanobacteria in symbiosis with cycads. In Prokaryotic Symbionts in Plants. Microbiology Monographs; Pawlowski, K., Ed.; Springer: Heidelberg, Germany, 2008; Volume 8, pp. 225–233. [Google Scholar]
- Bergman, B.; Johansson, C.; Söderbäck, E. The Nostoc-Gunnera symbiosis. New Phytol. 1992, 122, 379–400. [Google Scholar] [CrossRef]
- Chittora, D.; Meena, M.; Barupal, T.; Swapnil, P.; Sharma, K. Cyanobacteria as a source of biofertilizers for sustainable agriculture. Biochem. Biophys. Rep. 2020, 22, 100737. [Google Scholar] [CrossRef]
- Hermans, S.M.; Buckley, H.L.; Case, B.S.; Curran-Cournane, F.; Taylor, M.; Lear, G. Bacteria as emerging indicators of soil condition. Appl. Environ. Microbiol. 2017, 83, e02826-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dasgupta, D.; Ghosh, R.; Sengupta, T.K. Biofilm-mediated enhanced crude oil degradation by newly isolated Pseudomonas species. Int. Sch. Res. Notices 2013, 2013, 250749. [Google Scholar] [CrossRef] [Green Version]
- Ueshima, M.; Ginn, B.R.; Haack, E.A.; Szymanowski, J.E.S.; Fein, J.B. Cd adsorption onto Pseudomonas putida in the presence and absence of extracellular polymeric substances. Geochim. Cosmochim. Acta 2008, 72, 5885–5895. [Google Scholar] [CrossRef]
- Chettri, B.; Singh, A.K. Kinetics of hydrocarbon degradation by a newly isolated heavy metal tolerant bacterium Novosphingobium panipatense P5:ABC. Bioresour. Technol. 2019, 294, 122190. [Google Scholar] [CrossRef]
- Lee, S.Y.; Lee, Y.Y.; Cho, K.S. Effect of Novosphingobium sp. CuT1 inoculation on the rhizoremediation of heavy metal- and diesel-contaminated soil planted with tall fescue. Environ. Sci. Pollut. Res. 2022, 30, 16612–16625. [Google Scholar] [CrossRef]
- Asaf, S.; Numan, M.; Khan, A.L.; Al-Harrasi, A. Sphingomonas: From diversity and genomics to functional role in environmental remediation and plant growth. Crit. Rev. Biotechnol. 2020, 40, 138–152. [Google Scholar] [CrossRef]
- Nocelli, N.; Bogino, P.C.; Banchio, E.; Giordano, W. Roles of extracellular polysaccharides and biofilm formation in heavy metal resistance of rhizobia. Materials 2011, 9, 418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, P.; Diwan, B. Bacterial exopolysaccharide mediated heavy metal removal: A review on biosynthesis, mechanism and remediation strategies. Biotechnol. Rep. 2017, 13, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.D.; Yang, S.Z.; Li, H.P.; Zhu, H.H. Massilia putida sp. nov., a dimethyl disulfide-producing bacterium isolated from wolfram mine tailing. Int. J. Syst. Evol. Microbiol. 2016, 66, 50–55. [Google Scholar] [CrossRef]
- Chikere, C.B.; Okpokwasili, G.C.; Chikere, B.O. Monitoring of microbial hydrocarbon remediation in the soil. 3 Biotech 2011, 1, 117–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Xiao, E.; Krumins, V.; Häggblom, M.M.; Dong, Y.; Pu, Z.; Li, B.; Wang, Q.; Xiao, T.; Li, F. Rhizosphere microbial response to multiple metal(loid)s in different contaminated arable soils indicates crop-specific metal-microbe interactions. Appl. Environ. Microbiol. 2018, 84, e00701–e00718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diestra, E.; Esteve, I.; Burnat, M.; Maldonado, J.; Solé, A. Isolation and characterization of a heterotrophic bacterium able to grow in different environmental stress conditions, including crude oil and heavy metals. Commun. Curr. Res. Educ. Top. Trends Appl. Microbiol. 2007, 1, 90–99. [Google Scholar]
- Jing, Y.; He, Z.; Yang, X. Role of soil rhizobacteria in phytoremediation of heavy metal contaminated soils. J. Zhejiang Univ. Sci. B (Biomed. Biotechnol.) 2007, 8, 192–207. [Google Scholar] [CrossRef] [Green Version]
- Santoyo, G.; Moreno-Hagelsieb, G.; Orozco-Mosqueda, M.C.; Glick, B.R. Plant growth-promoting bacterial endophytes. Microbiol. Res. 2016, 183, 92–99. [Google Scholar] [CrossRef]
- Ojuederie, O.; Babalola, O. Microbial and plant-assisted bioremediation of heavy metal polluted environments: A review. Int. J. Environ. Res. Public Health 2017, 14, 1504. [Google Scholar] [CrossRef] [Green Version]
- Hussain, S.S.; Mehnaz, S.; Siddique, K.H.M. Harnessing the plant microbiome for improved abiotic stress tolerance. In Plant Microbiome: Stress Response; Egamberdieva, D., Ahmad, P., Eds.; Springer Nature Singapore Pte Ltd: Singapore, 2018; pp. 21–43. [Google Scholar]
- Babu, A.G.; Kim, J.-D.; Oh, B.-T. Enhancement of heavy metal phytoremediation by Alnus firma with endophytic Bacillus thuringiensis GDB-1. J. Hazard. Mater. 2013, 250–251, 477–483. [Google Scholar] [CrossRef]
- Chen, Z.-J.; Tian, W.; Li, Y.-J.; Sun, L.-N.; Chen, Y.; Zhang, H.; Li, Y.Y.; Han, H. Responses of rhizosphere bacterial communities, their functions and their network interactions to Cd stress under phytostabilization by Miscanthus spp. Environ. Pollut. 2021, 287, 117663. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Zhang, Y.; Rafiq, M.T.; Khan, K.Y.; Pan, F.; Yang, X.; Feng, Y. Improvement of cadmium uptake and accumulation in Sedum alfredii by endophytic bacteria Sphingomonas SaMR12: Effects on plant growth and root exudates. Chemosphere 2014, 17, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Guo, D.; Mahar, A.; Wang, Z.; Muhammad, D.; Li, R.; Wang, P.; Shen, F.; Xue, Q.; Zhang, Z. Role of Streptomyces pactum in phytoremediation of trace elements by Brassica juncea in mine polluted soils. Ecotoxicol. Environ. Saf. 2017, 144, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Preston, G.M. Plant perceptions of plant growth-promoting Pseudomonas. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 907–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshwal, V.K.; Singh, S.B.; Kumar, P.; Chubey, A. Rhizobia unique plant growth promoting rhizobacteria: A review. Int. J. Life Sci. 2013, 2, 74–86. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Background Soil (Yasnaya Polyana) | KGIW | Tulachermet | Lenin Ave. |
|---|---|---|---|---|
| Soil type | Clay loam | Clay loam | Sandy loam | Clay loam |
| pH | 6.20 | 7.26 | 7.35 | 7.29 |
| Total carbon (% of air-dried soil) | 4.87 ± 0.05 | 4.41 ± 0.11 | 4.84 ± 0.33 | 3.83 ± 0.06 |
| Water-soluble carbon (% of total carbon) | 0.14 ± 0.05 | 0.13 ± 0.02 | 0.14 ± 0.06 | 0.12 ± 0.03 |
| Humus (% of air-dried soil) | 8.12 ± 0.16 | 7.14 ± 0.14 | 7.62 ± 0.15 | 6.16 ± 0.12 |
| Humus carbon (%) | 4.72 ± 0.09 | 4.15 ± 0.08 | 4.43 ± 0.09 | 3.58 ± 0.07 |
| N-NO3 (mg/kg dw) | 20.5 ± 1.2 | 28.2 ± 1.6 | 38.6 ± 3.5 | 41.1 ± 2.4 |
| N-NH4 (mg/kg dw) | 12.9 ±1.2 | 6.9 ± 0.7 | 9.5 ± 0.3 | 2.3 ± 0.2 |
| P2O5 (mg/kg dw) | 51.0 ± 4.8 | 159.5 ± 3.6 | 98.4 ± 7.6 | 218.0 ± 6.7 |
| Oil products (g/kg dw) | 1.5 ± 0.6 | 2.6 ± 0.4 | 4.1 ± 1.0 | 2.5 ± 0.7 |
| Metal(loid)s (mg/kg dw): | ||||
| Fe | 15,600 ± 1860 | 78,100 ± 1280 | 120,600 ± 5830 | 37,400 ± 2160 |
| Mn | 1300 ± 67 | 5700 ± 180 | 1100 ± 87 | 1600 ± 58 |
| V | 57 ± 3.0 | 41 ± 4.1 | 136 ± 7.2 | 61 ± 2.9 |
| Ni | 25 ± 3.1 | 31 ± 4.2 | 55 ± 3.3 | 35 ± 2.8 |
| Cu | 29 ± 2.3 | 52 ± 4.1 | 75 ± 0.8 | 378 ± 0.9 |
| Zn | 47 ± 2.0 | 310 ± 9.4 | 161 ± 3.6 | 186 ± 5.3 |
| Pb | 18 ± 2.1 | 72 ± 6.0 | 26 ± 0.7 | 59 ± 1.1 |
| As | 5.1 ± 0.4 | 5.9 ± 0.5 | 6.4 ± 0.2 | 7.3 ± 0.3 |
| Soil | Plant | Observed Features | Chao1 | Shannon Index | Simpson Index | Faith PD |
|---|---|---|---|---|---|---|
| Background | A. tricolor cv. Valentina | 743 | 748.000 | 8.6560 | 0.9964 | 81.03 |
| A. cruentus cv. Dyuimovochka | 716 | 718.111 | 8.6309 | 0.9957 | 73.42 | |
| A. caudatus cv. Bulava | 890 | 898.347 | 8.5701 | 0.9930 | 86.78 | |
| A. caudatus cv. Zelenaya Sosulka | 910 | 916.949 | 8.7466 | 0.9955 | 97.51 | |
| Tulachermet | A. tricolor cv. Valentina | 741 | 743.258 | 8.6019 | 0.9956 | 87.83 |
| A. cruentus cv. Dyuimovochka | 823 | 829.343 | 8.8715 | 0.9970 | 87.65 | |
| A. caudatus cv. Bulava | 1010 | 1014.614 | 9.2749 | 0.9978 | 92.29 | |
| A. caudatus cv. Zelenaya Sosulka | 819 | 831.470 | 8.8402 | 0.9967 | 81.70 | |
| KGIW | A. tricolor cv. Valentina | 807 | 807.786 | 8.9055 | 0.9970 | 82.02 |
| A. cruentus cv. Dyuimovochka | 806 | 806.992 | 8.6081 | 0.9940 | 88.70 | |
| A. caudatus cv. Bulava | 990 | 994.844 | 9.1206 | 0.9970 | 97.06 | |
| A. caudatus cv. Zelenaya Sosulka | 964 | 974.350 | 9.1507 | 0.9974 | 88.53 | |
| Lenin Ave. | A. tricolor cv. Valentina | 832 | 836.833 | 8.9232 | 0.9969 | 85.43 |
| A. cruentus cv. Dyuimovochka | 601 | 602.600 | 8.4401 | 0.9959 | 63.57 | |
| A. caudatus cv. Bulava | 758 | 766.928 | 8.3750 | 0.9926 | 81.42 | |
| A. caudatus cv. Zelenaya Sosulka | 707 | 716.784 | 8.5888 | 0.9957 | 74.67 |
| Plant Cultivars | Bacterial Taxa | |
|---|---|---|
| A. tricolor cv. Valentina | g_Chloronema; s_Scytonema tolypothrichoides | Background; Tulachermet; KGIW |
| g_Leptolyngbya_VRUC_135;s_uncultured bacterium | Background; Tulachermet; Lenin Ave. | |
| g_Alsobacter; s_Alsobacter_metallidurans | Tulachermet; KGIW; Lenin Ave. | |
| g_C0119; s_uncultured_bacterium | Background; Tulachermet; Lenin Ave. | |
| g_Rhodocytophaga; s_uncultured Bacteroidetes | Background; Tulachermet; KGIW | |
| f_Rhodanobacteraceae; g_uncultured; s_uncultured bacterium | Background; Tulachermet; KGIW | |
| g__DS-100; s_unknown | Tulachermet; KGIW; Lenin Ave. | |
| A. cruentus cv. Dyuimovochka | g_Methylophilus; s_uncultured bacterium | Tulachermet; KGIW; Lenin Ave. |
| g_CENA518; s_uncultured_bacterium | Tulachermet; KGIW; Lenin Ave. | |
| A. caudatus cv. Bulava | g_Nakamurella; g_unknown; s_unknown | Background; Tulachermet; KGIW |
| g_Pir4_lineage; s_uncultured Pirellula | Background; Tulachermet; Lenin Ave. | |
| f_Moraxellaceae; g_uncultured; s_uncultured gamma | Background; KGIW; Lenin Ave. | |
| f_0319-7L14; g_0319-7L14; s_uncultured bacterium | Background; Tulachermet; Lenin Ave. | |
| o_Planctomycetales; f_uncultured; g_uncultured | Background; Tulachermet; KGIW | |
| f_Phycisphaeraceae; g_uncultured | Tulachermet; KGIW; Lenin Ave. | |
| f_Kapabacteriales; g_Kapabacteriales | Tulachermet; KGIW; Lenin Ave. | |
| f_Sericytochromatia; g_Sericytochromatia | Background; Tulachermet; KGIW | |
| A. caudatus cv. Zelenaya Sosulka | f_Vicinamibacteraceae; g_Vicinamibacteraceae;s_uncultured Geothrix | Tulachermet; KGIW; Lenin Ave. |
| f_Xanthomonadaceae; g_unknown; s_unknown | Background; Tulachermet; KGIW | |
| f_Rhodanobacteraceae; g_Ahniella; s_uncultured bacterium | Background; Tulachermet; Lenin Ave. | |
| f_Latescibacterota; g_Latescibacterota; s_uncultured soil | Background; Tulachermet; Lenin Ave. | |
| f_Blastocatellaceae; g_Aridibacter; s_uncultured bacterium | Background; KGIW; Lenin Ave. | |
| f_Subgroup 22; g_Subgroup 22 | Tulachermet; KGIW; Lenin Ave. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muratova, A.; Gorelova, S.; Golubev, S.; Kamaldinova, D.; Gins, M. Rhizosphere Microbiomes of Amaranthus spp. Grown in Soils with Anthropogenic Polyelemental Anomalies. Agronomy 2023, 13, 759. https://doi.org/10.3390/agronomy13030759
Muratova A, Gorelova S, Golubev S, Kamaldinova D, Gins M. Rhizosphere Microbiomes of Amaranthus spp. Grown in Soils with Anthropogenic Polyelemental Anomalies. Agronomy. 2023; 13(3):759. https://doi.org/10.3390/agronomy13030759
Chicago/Turabian StyleMuratova, Anna, Svetlana Gorelova, Sergey Golubev, Dilyara Kamaldinova, and Murat Gins. 2023. "Rhizosphere Microbiomes of Amaranthus spp. Grown in Soils with Anthropogenic Polyelemental Anomalies" Agronomy 13, no. 3: 759. https://doi.org/10.3390/agronomy13030759