
Fungicidal Activity of Caproate Produced by Clostridium sp. strain E801, a Bacterium Isolated from Cocopeat Medium Subjected to Anaerobic Soil Disinfestation

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Conidial Suspension
2.2. Laboratory-Scale ASD
2.3. High-Throughput Sequencing of 16S rRNA Gene Amplicons
2.4. Isolation of the Predominant Bacterium from ASD-Treated Soil
2.5. Phylogenetic Analysis of Strain E801
2.6. Effect of Culture Supernatant of Strain E801 on the Growth of Fol
2.7. Effect of Volatile Fatty Acids on the Viability of Fol Conidia and R. solanacearum Cells
2.8. Effect of Volatile Fatty Acids on the Growth of Fol at a Low Temperature
2.9. Evaluation of Fungicidal Activity of Caproate in Cocopeat Medium
2.10. Analytical Techniques
2.11. Nucleotide Sequence Accession Numbers
3. Results
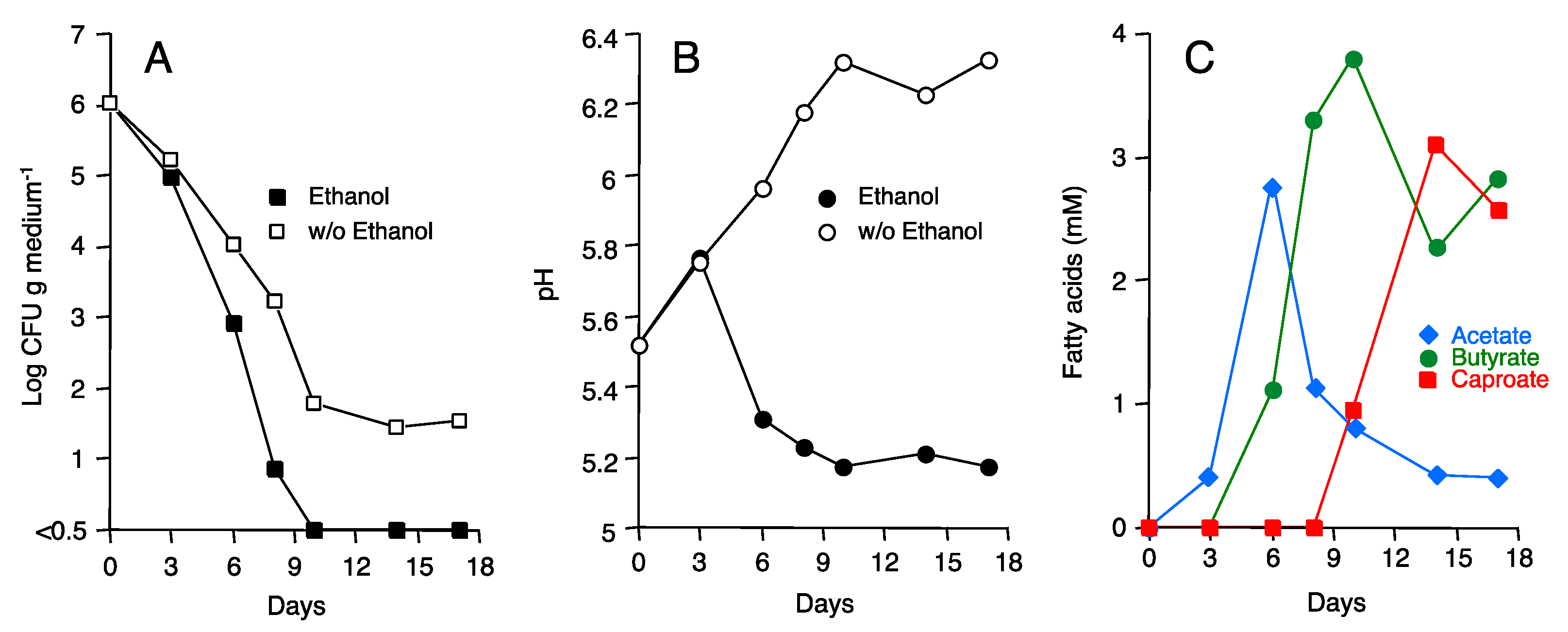
3.1. Laboratory-Scale ASD with Ethanol as Carbon Source
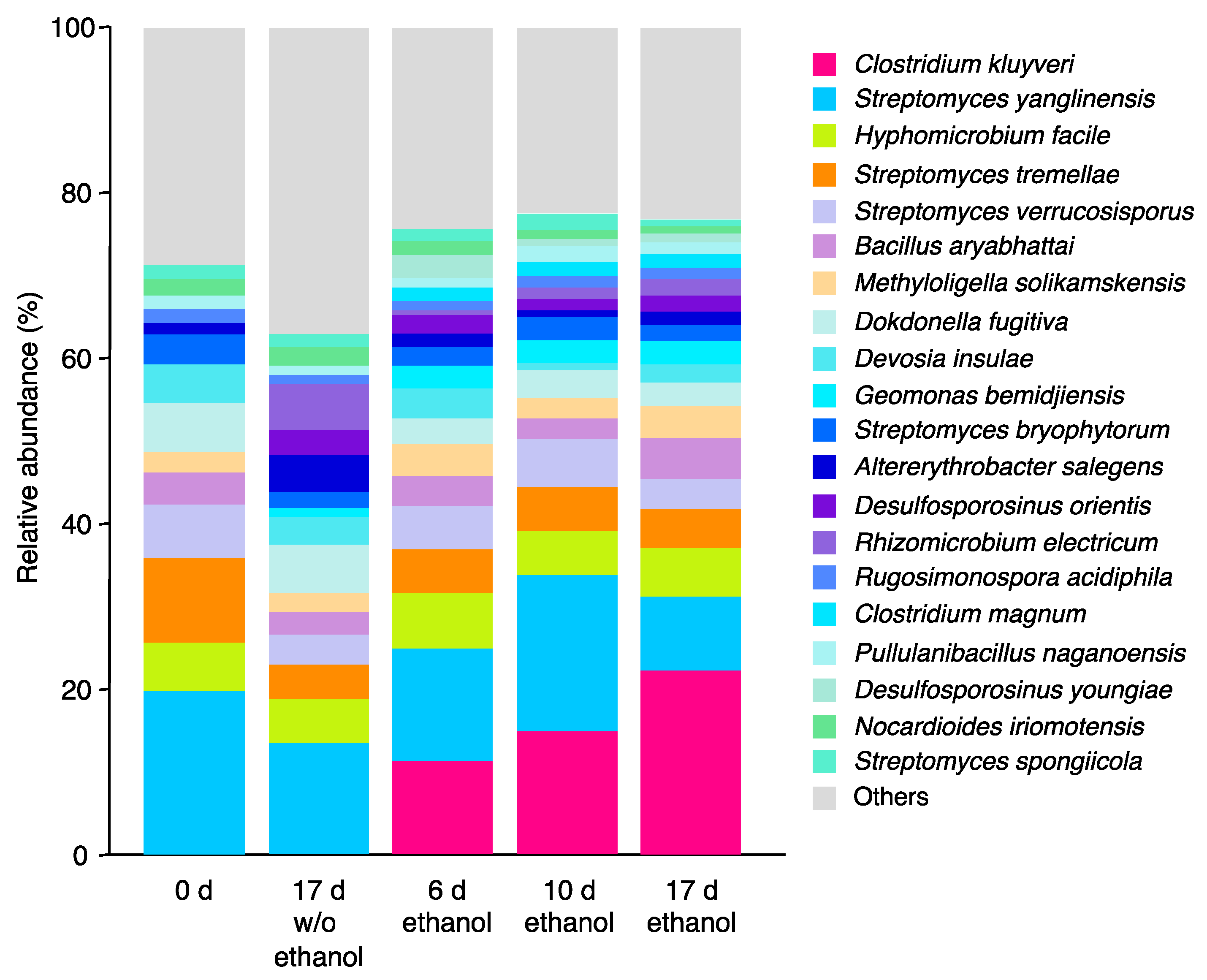
3.2. Microbial Community Shift during ASD
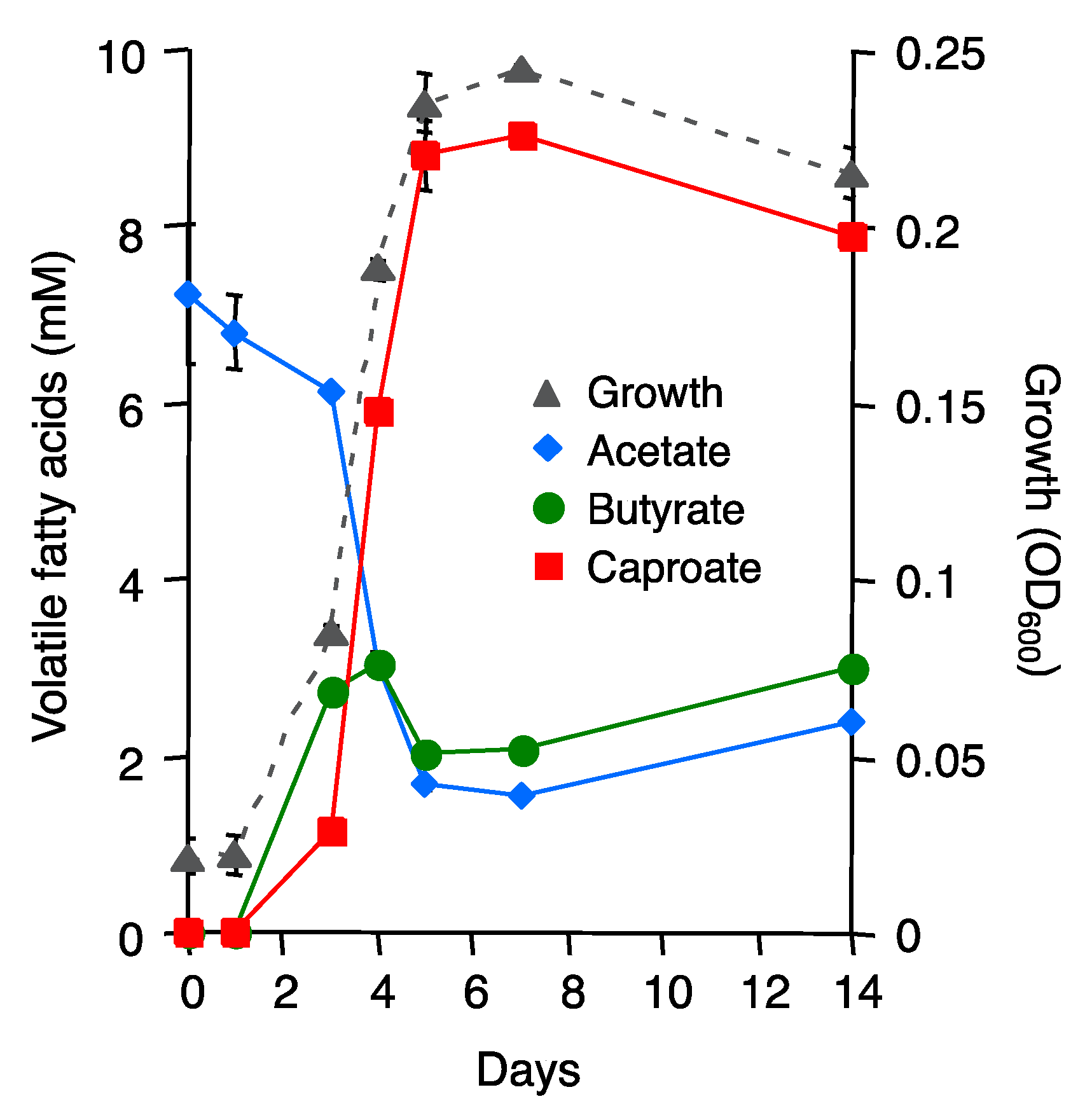
3.3. Isolation of Clostridium sp. E801 from ASD-Treated Cocopeat Medium
3.4. Effect of Culture Supernatant of Strain E801 on the Growth of Fol
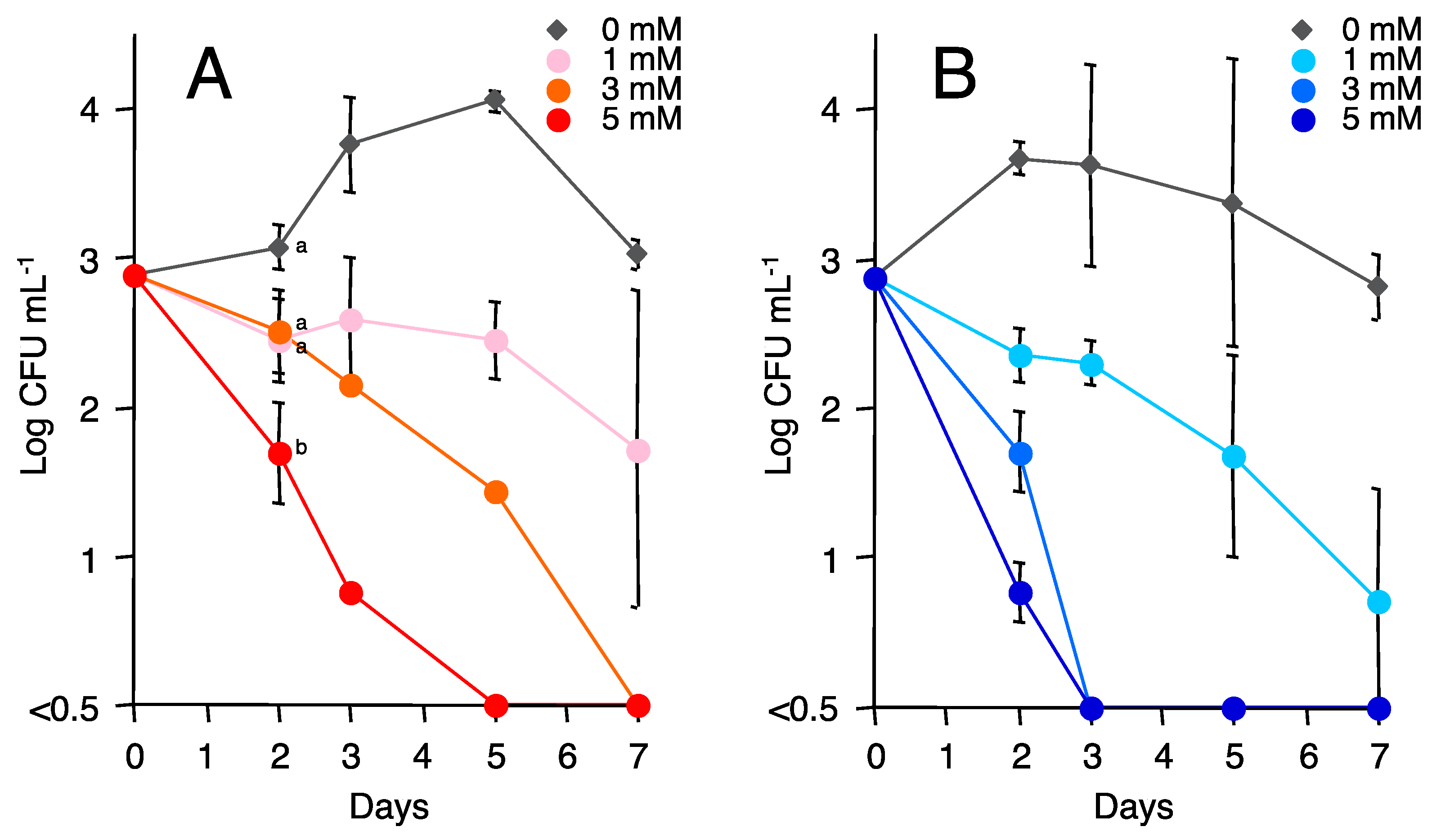
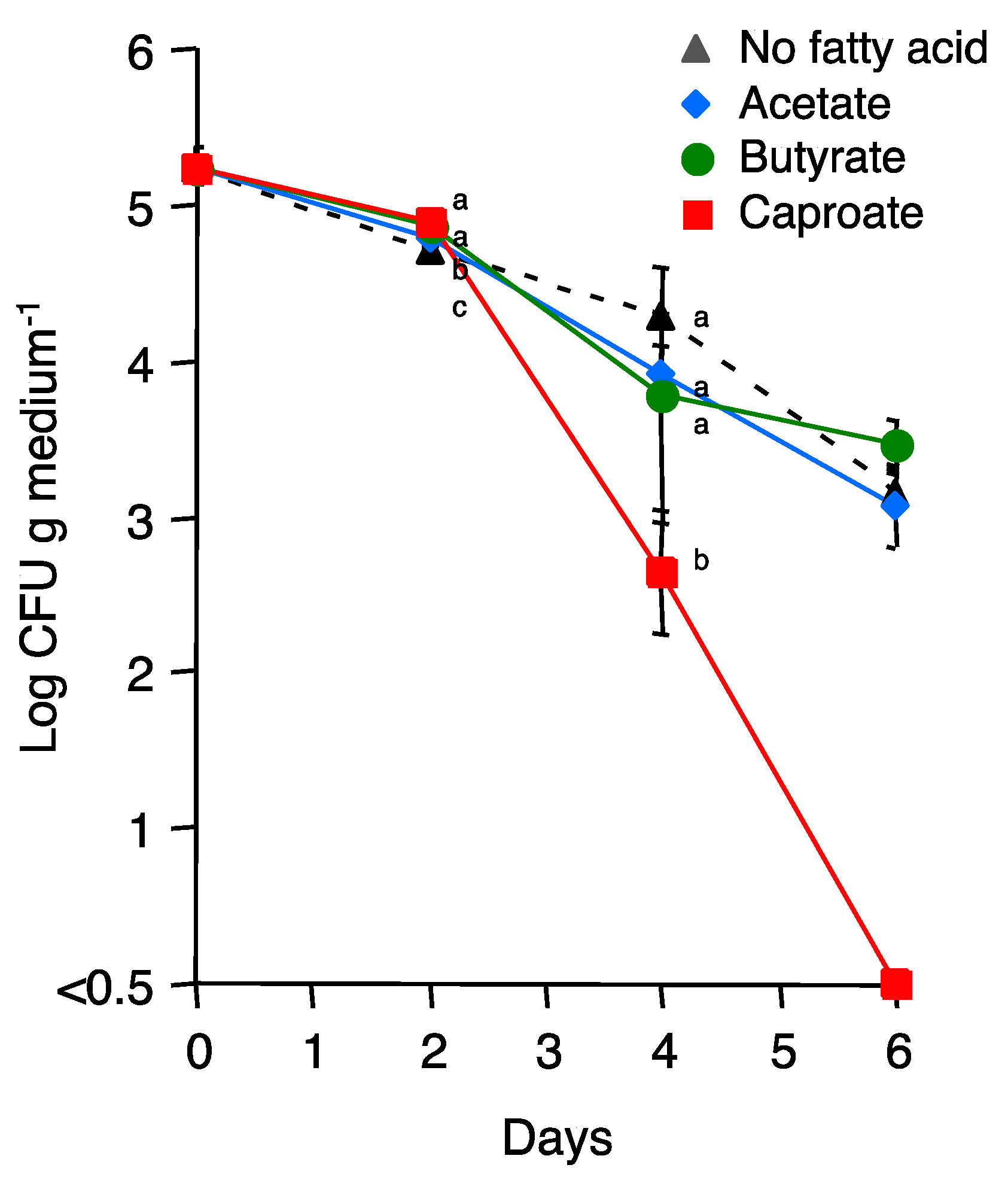
3.5. Effect of Caproate on the Viability of Fol Conidia and R. solanacearum Cells
3.6. Effect of Caproate on the Growth of Fol at Low Temperature
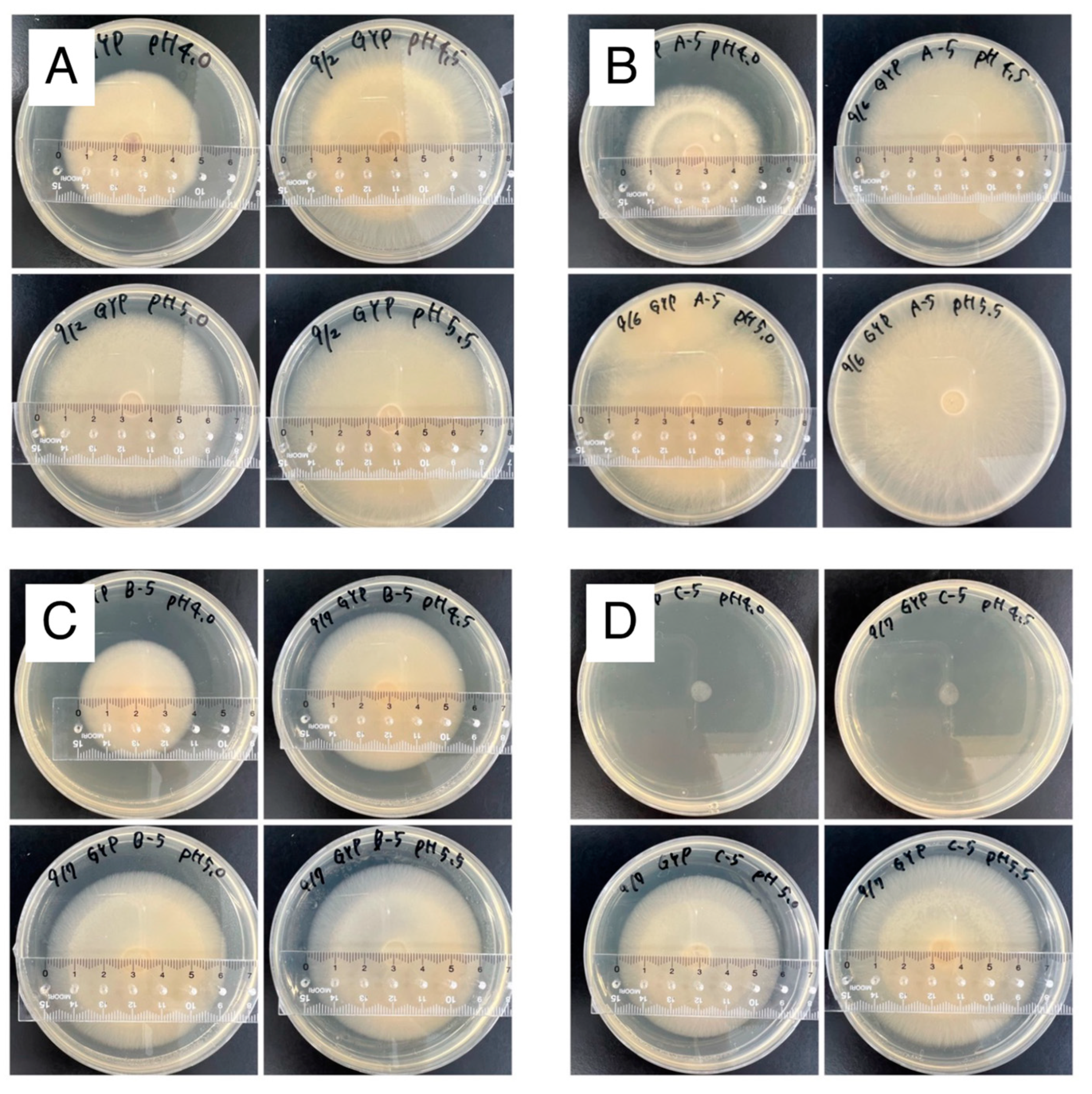
3.7. Evaluation of Fungicidal Activity of Caproate in Cocopeat Medium
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mihajlovic, M.; Rekanovic, E.; Hrustic, J.; Grahovac, M.; Tanovic, B. Methods for management of soil-borne plant pathogens. Pestic. Fitomed. 2017, 32, 9–24. [Google Scholar] [CrossRef] [Green Version]
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for management of soil-borne diseases in crop production. Agriculture 2020, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Thomas, W.B. Methyl bromide: Effective pest management tool and environmental threat. J. Nematol. 1996, 28, 586–589. [Google Scholar] [PubMed]
- Velders, G.J.M.; Andersen, S.O.; Daniel, J.S.; Fahey, D.W.; McFarland, M. The importance of the Montreal Protocol in protecting climate. Proc. Natl. Acad. Sci. USA 2007, 104, 4814–4819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desaeger, J.; Dickson, D.W.; Locascio, S.J. Methyl bromide alternatives for control of root-knot nematode (Meloidogyne spp.) in tomato production in Florida. J. Nematol. 2017, 49, 140–149. [Google Scholar]
- Shinmura, A. Causal agent and control of root rot of welsh onion. PSJ Soilborne Dis. Workshop Rep. 2000, 20, 133–143. [Google Scholar]
- Blok, W.J.; Lamers, J.G.; Termorshuizen, A.J.; Bollen, G.J. Control of soil-borne plant pathogens by incorporating fresh organic amendments followed by tarping. Phytopathology 2000, 90, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Momma, N.; Kobara, Y.; Uematsu, S.; Kita, N.; Shinmura, A. Development of biological soil disinfestations in Japan. Appl. Microbiol. Biotechnol. 2013, 97, 3801–3809. [Google Scholar] [CrossRef]
- Strauss, S.L.; Kluepfel, D.A. Anaerobic soil disinfestation: A chemical-independent approach to pre-plant control of plant pathogens. J. Integr. Agric. 2015, 14, 2309–2318. [Google Scholar] [CrossRef]
- Priyashantha, A.K.H.; Attanayake, R.N. Can anaerobic soil disinfestation (ASD) be a game changer in tropical agriculture? Pathogens 2021, 10, 133. [Google Scholar] [CrossRef]
- Ueki, A.; Kaku, N.; Ueki, K. Role of anaerobic bacteria in biological soil disinfestation for elimination of soil-borne plant pathogens in agriculture. Appl. Microbiol. Biotechnol. 2018, 102, 6309–6318. [Google Scholar] [CrossRef]
- Shennan, C.; Muramoto, J.; Lamers, J.; Mazzola, M.; Rosskopf, E.N.; Kokalis-Burelle, N.; Momma, N.; Butler, D.M.; Kobara, Y. Anaerobic soil disinfestation for soil borne disease control in strawberry and vegetable systems: Current knowledge and future directions. Acta Hortic. 2014, 1044, 165–175. [Google Scholar] [CrossRef]
- Momma, N.; Yamamoto, K.; Simandi, P.; Shishido, M. Role of organic acids in the mechanism of biological soil disinfestation (BSD). J. Gen. Plant Pathol. 2006, 72, 247–252. [Google Scholar] [CrossRef]
- Hewavitharana, S.S.; Ruddell, D.; Mazzola, M. Carbon source-dependent antifungal and nematicidal volatiles derived during anaerobic soil disinfestation. Eur. J. Plant Pathol. 2014, 140, 39–52. [Google Scholar] [CrossRef]
- van Agtmaal, M.; van Os, G.J.; Gera Hol, W.H.; Hundscheid, M.P.J.; Runia, W.T.; Hordijk, C.A.; de Boer, W. Legacy effects of anaerobic soil disinfestation on soil bacterial community composition and production of pathogen-suppressing volatiles. Front. Microbiol. 2015, 6, 701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Momma, N.; Kobara, Y.; Momma, M. Fe2+ and Mn2+, potential agents to induce suppression of Fusarium oxysporum for biological soil disinfestation. J. Gen. Plant Pathol. 2011, 77, 331–335. [Google Scholar] [CrossRef]
- Ueki, A.; Takeshita, T.; Ishioka, G.; Kaku, N.; Ueki, K. Degradation of the fungal cell wall by clostridial strains isolated from soil subjected to biological soil disinfestation and biocontrol of Fusarium wilt disease of spinach. Appl. Microbiol. Biotechnol. 2017, 101, 8267–8277. [Google Scholar] [CrossRef]
- Ueki, A.; Takeshita, T.; Ishioka, G.; Kaku, N.; Ueki, K. Production of β-1,3-glucanase and chitosanase from clostridial strains isolated from the soil subjected to biological soil disinfestation. AMB Express 2019, 9, 114. [Google Scholar] [CrossRef] [Green Version]
- Momma, N.; Momma, M.; Kobara, Y. Biological soil disinfestation using ethanol: Effect on Fusarium oxysporum f. sp. lycopersici and soil microorganisms. J. Gen. Plant Pathol. 2010, 76, 336–344. [Google Scholar] [CrossRef]
- Momma, N. Biological soil disinfestation (BSD) of soilborne pathogens and its possible mechanisms. Jpn. Agric. Res. Q. 2008, 42, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, N. Selective media for Fusarium oxysporum. J. Gen. Plant Pathol. 2007, 73, 342–348. [Google Scholar] [CrossRef]
- Takahashi, S.; Tomita, J.; Nishioka, K.; Hisada, T.; Nishijima, M. Development of a prokaryotic universal primer for simultaneous analysis of Bacteria and Archaea using next-generation sequencing. PLoS ONE 2014, 9, e105592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aronesty, E. Comparison of sequencing utility programs. Open Bioinform. J. 2013, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335336. [Google Scholar] [CrossRef] [Green Version]
- Seedorf, H.; Fricke, W.F.; Veith, B.; Brüggemann, H.; Liesegang, H.; Strittmatter, A.; Miethke, M.; Buckel, W.; Hinderberger, J.; Li, F.; et al. The genome of Clostridium kluyveri, a strict anaerobe with unique metabolic features. Proc. Natl. Acad. Sci. USA 2008, 105, 2128–2133. [Google Scholar] [CrossRef] [Green Version]
- Yamamura, S.; Kurasawa, H.; Kashiwabara, Y.; Hori, T.; Aoyagi, T.; Nakajima, N.; Amachi, S. Soil microbial communities involved in reductive dissolution of arsenic from arsenate-laden minerals with different carbon sources. Environ. Sci. Technol. 2019, 53, 12398–12406. [Google Scholar] [CrossRef] [PubMed]
- Hiraishi, A. Direct automated sequencing of 16S rDNA amplified by polymerase chain reaction from bacterial cultures without DNA purification. Lett. Appl. Microbiol. 1992, 15, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Saito, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Kelman, A. The relationship of pathogenicity in Pseudomonas solanacearum to colony appearance on a tetrazolium medium. Phytopathology 1954, 44, 693–695. [Google Scholar]
- Mowlick, S.; Takehara, T.; Kaku, N.; Ueki, K.; Ueki, A. Proliferation of diversified clostridial species during biological soil disinfestation incorporated with plant biomass under various conditions. Appl. Microbiol. Biotechnol. 2013, 97, 8365–8379. [Google Scholar] [CrossRef] [Green Version]
- Mowlick, S.; Inoue, T.; Takehara, T.; Kaku, N.; Ueki, K.; Ueki, A. Changes and recovery of soil bacterial communities influenced by biological soil disinfestation as compared with chloropicrin-treatment. AMB Express 2013, 3, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Liu, L.; Wen, T.; Zhu, R.; Zhang, J.; Cai, Z. Illumina MiSeq investigations on the changes of microbial community in the Fusarium oxysporum f.sp. cubense infected soil during and after reductive soil disinfestation. Microbiol. Res. 2015, 181, 33–42. [Google Scholar] [PubMed]
- Tan, X.; Liao, H.; Shu, L.; Yao, H. Effect of different substrates on soil microbial community structure and the mechanism of reductive soil disinfestation. Front. Microbiol. 2019, 10, 2851. [Google Scholar] [CrossRef] [PubMed]
- Poret-Peterson, A.T.; Sayed, N.; Glyzewski, N.; Forbes, H.; González-Orta, E.T.; Kluepfel, D.A. Temporal responses of microbial communities to anaerobic soil disinfestation. Microb. Ecol. 2020, 80, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Momma, N.; Usami, T.; Shishido, M. Detection of Clostridium sp. inducing biological soil disinfestation (BSD) and suppression of pathogens causing fusarium wilt and bacterial wilt of tomato by gases evolved during BSD. Soil Microorg. 2007, 61, 3–9, (In Japanese with English Abstract). [Google Scholar]
- Huang, X.Q.; Wen, T.; Zhang, J.B.; Meng, L.; Zhu, T.B.; Liu, L.L.; Cai, Z.C. Control of soil-borne pathogen Fusarium oxysporum by biological soil disinfestation with incorporation of various organic matters. Eur. J. Plant Pathol. 2015, 143, 223–235. [Google Scholar] [CrossRef]
- Hewavitharana, S.S.; Klarer, E.; Reed, A.J.; Leisso, R.; Poirier, B.; Honaas, L.; Rudell, D.R.; Mazzola, M. Temporal dynamics of the soil metabolome and microbiome during simulated anaerobic soil disinfestation. Front. Microbiol. 2019, 10, 2365. [Google Scholar] [CrossRef] [Green Version]
- Barker, H.A. The production of caproic and butyric acids by the methane fermentation of ethyl alcohol. Arch. Mikrobiol. 1937, 8, 415–421. [Google Scholar] [CrossRef]
- Bertsch, J.; Siemund, A.L.; Kremp, F.; Müller, V. A novel route for ethanol oxidation in the acetogenic bacterium, Acetobacterium woodii: The acetoaldehyde/ethanol dehydrogenase pathway. Environ. Microbiol. 2016, 18, 2913–2922. [Google Scholar] [CrossRef]
- Igarashi, K.; Kato, S. Reductive transformation of Fe(III) (oxyhydr)oxides by mesophilic homoacetogens in the genus Sporomusa. Front. Microbiol. 2021, 12, 600808. [Google Scholar] [CrossRef]
- Katase, M.; Kubo, C.; Ushio, S.; Ootsuka, E.; Takeuchi, T.; Mizukubo, T. Nematicidal activity of volatile fatty acids generated from wheat bran in reductive soil disinfection. Nematol. Res. 2009, 39, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Wen, T.; Zhang, J.; Meng, L.; Zhu, T.; Cai, Z. Toxic organic acids produced in biological soil disinfestation mainly caused the suppression of Fusarium oxysporum f. sp. cubense. BioControl 2015, 60, 113–124. [Google Scholar] [CrossRef]
- Jarboe, L.R.; Royce, L.A.; Liu, P. Understanding biocatalyst inhibition by carboxylic acids. Front. Microbiol. 2013, 4, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Ruan, W.; Li, J.; Xu, H.; Wang, J.; Gao, Y.; Wang, J. Biological control of phytopathogenic fungi by fatty acids. Mycopathologia 2008, 166, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Leyva, M.O.; Vicedo, B.; Finiti, I.; Flors, V.; Del Amo, G.; Real, M.D.; García-Agustín, P.; González-Bosch, C. Preventive and post-infection control of Botrytis cinerea in tomato plants by hexanoic acid. Plant Pathol. 2008, 57, 1038–1046. [Google Scholar] [CrossRef]
- Caccalano, M.N.; Dilarri, G.; Domingues, D.S.; Ferreira, H. Hexanoic acid: A new potential substitute for copper-based agrochemicals against citrus canker. J. Appl. Microbiol. 2021, 131, 2488–2499. [Google Scholar] [CrossRef]
- Horita, M.; Kitamoto, H.K. Biological soil disinfestation using bioethanol fermentation products: Role of residual organic substances. J. Gen. Plant Pathol. 2015, 81, 304–314. [Google Scholar] [CrossRef]
- Shrestha, U.; Augé, R.M.; Butler, D.M. A meta-analysis of the impact of anaerobic soil disinfestation on pest suppression and yield of horticultural crops. Front. Plant Sci. 2016, 7, 1254. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, U.; Ownley, B.H.; Bruce, A.; Rosskopf, E.N.; Butler, D.M. Anaerobic soil disinfestation efficacy against Fosarium oxysporum is affected by soil temperature, amendment type, rate, and C:N ratio. Phytopathology 2021, 111, 1380–1392. [Google Scholar] [CrossRef]
- Hewavitharana, S.S.; Klarer, E.; Muramoto, J.; Shennan, C.; Mazzola, M. Analysis of environmental variables and carbon input on soil microbiome, metabolome and disease control efficacy in strawberry attributable to anaerobic soil disinfestation. Microorganisms 2021, 9, 1638. [Google Scholar] [CrossRef]
- Rosskopf, E.N.; Burelle, N.; Hong, J.; Butler, D.M.; Noling, J.W.; He, Z.; Booker, B.; Sances, F. Comparison of anaerobic soil disinfestation and drip-applied organic acids for raised-bed specialty crop production in Florida. Acta Hortic. 2014, 1044, 221–228. [Google Scholar] [CrossRef]
- Yin, Y.; Zhang, Y.; Karakashev, D.B.; Wang, J.; Angelidaki, I. Biological caproate production by Clostridium kluyveri from ethanol and acetate as carbon sources. Bioresour. Technol. 2017, 241, 638–644. [Google Scholar] [CrossRef] [Green Version]
- Otten, J.K.; Zou, Y.; Papoutsakis, E.T. The potential of caproate (hexanoate) production using Clostridium kluyveri syntrophic cocultures with Clostridium acetobutylicum or Clostridium saccharolyticum. Front. Bioeng. Biotechnol. 2022, 10, 965614. [Google Scholar] [CrossRef] [PubMed]
- Aoyagi, T.; Hanada, S.; Itoh, H.; Sato, Y.; Ogata, A.; Friedrich, M.W.; Kikuchi, Y.; Hori, T. Ultra-high-sensitivity stable-isotope probing of rRNA by high-throughput sequencing of isopycnic centrifugation gradients. Environ. Microbiol. Rep. 2015, 7, 282–287. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amendment | Medium pH | |||
|---|---|---|---|---|
| 5.5 | 5.0 | 4.5 | 4.0 | |
| No addition | 5.66 ± 0.09 A | 5.36 ± 0.73 A | 5.10 ± 0.30 | 5.58 ± 0.29 |
| Culture supernatant | 6.04 ± 0.20 B | 4.49 ± 0.23 AB | <0.50 * | <0.50 * |
| Heat-treated sup. | 5.64 ± 0.07 A | 5.73 ± 0.26 B | <0.50 * | <0.50 * |
| Amendment | Medium pH | |||
|---|---|---|---|---|
| 5.5 | 5.0 | 4.5 | 4.0 | |
| Acetate | 6.18 ± 0.17A | 5.84 ± 0.06 A | 5.60 ± 0.12 | 5.53 ± 0.20 |
| Butyrate | 5.87 ± 0.19A | 5.61 ± 0.10 B | 5.40 ± 0.04 | 5.00 ± 0.13 |
| Caproate | 3.39 ± 0.01B | 2.39 ± 0.11 C | <0.50 * | <0.50 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shirane, S.; Momma, N.; Usami, T.; Suzuki, C.; Hori, T.; Aoyagi, T.; Amachi, S. Fungicidal Activity of Caproate Produced by Clostridium sp. strain E801, a Bacterium Isolated from Cocopeat Medium Subjected to Anaerobic Soil Disinfestation. Agronomy 2023, 13, 747. https://doi.org/10.3390/agronomy13030747
Shirane S, Momma N, Usami T, Suzuki C, Hori T, Aoyagi T, Amachi S. Fungicidal Activity of Caproate Produced by Clostridium sp. strain E801, a Bacterium Isolated from Cocopeat Medium Subjected to Anaerobic Soil Disinfestation. Agronomy. 2023; 13(3):747. https://doi.org/10.3390/agronomy13030747
Chicago/Turabian StyleShirane, Shota, Noriaki Momma, Toshiyuki Usami, Chiharu Suzuki, Tomoyuki Hori, Tomo Aoyagi, and Seigo Amachi. 2023. "Fungicidal Activity of Caproate Produced by Clostridium sp. strain E801, a Bacterium Isolated from Cocopeat Medium Subjected to Anaerobic Soil Disinfestation" Agronomy 13, no. 3: 747. https://doi.org/10.3390/agronomy13030747