

Plant Architecture Influences the Population Transpiration and Canopy Temperature in Winter Wheat Genotypes
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Design
2.3. Measurements
2.3.1. Morphological Traits
2.3.2. Leaf Transpiration Traits
2.3.3. Canopy Traits
2.3.4. Water Use Traits
2.4. Statistical Analysis
3. Results
3.1. Meteorological Conditions
3.2. Genotypic Comparison between Jing 411 and Jinmai 47 in the Morphological Traits and Gas Exchange Parameters
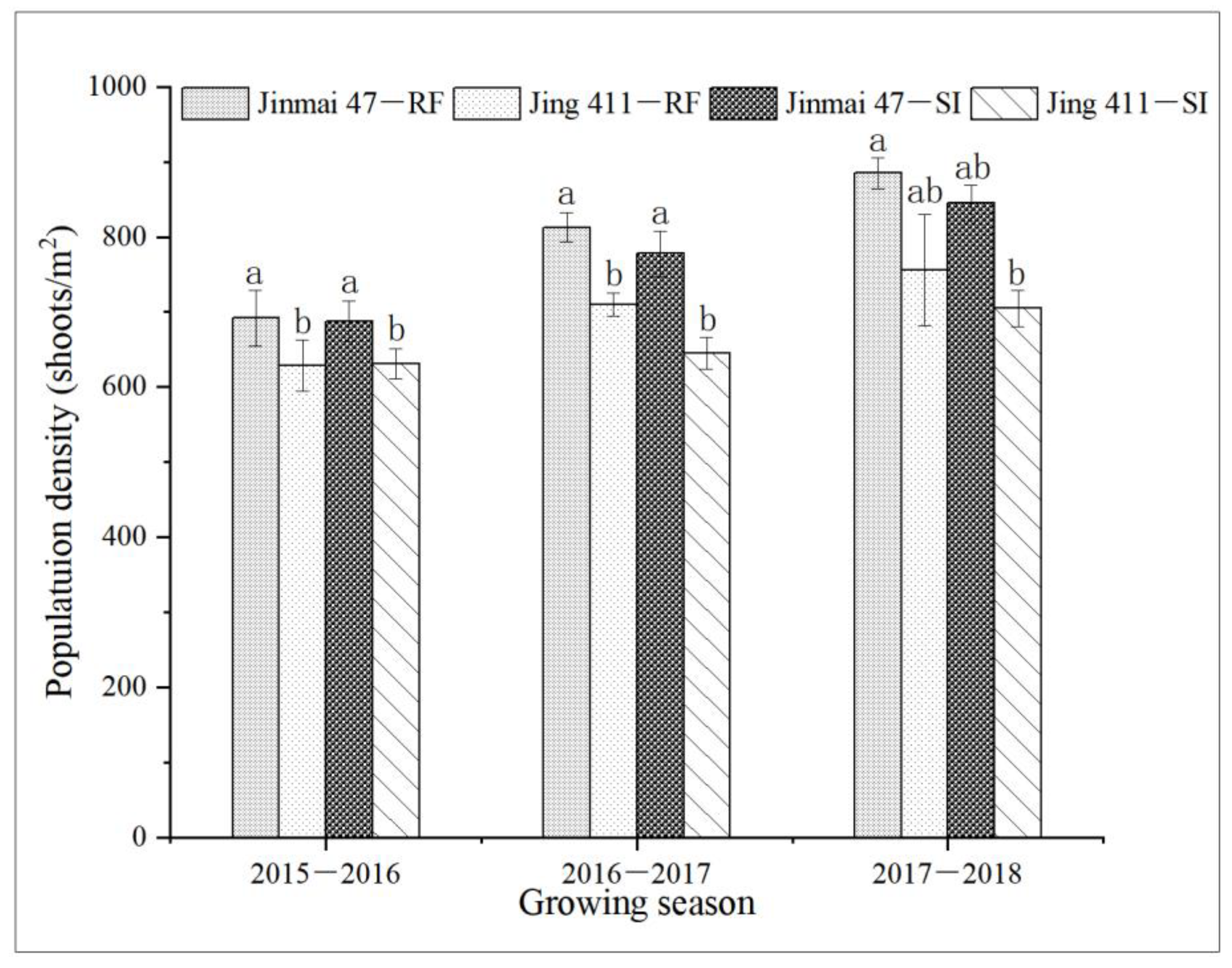
3.3. Genotypic Comparison between Jing 411 and Jinmai 47 in the Canopy Traits
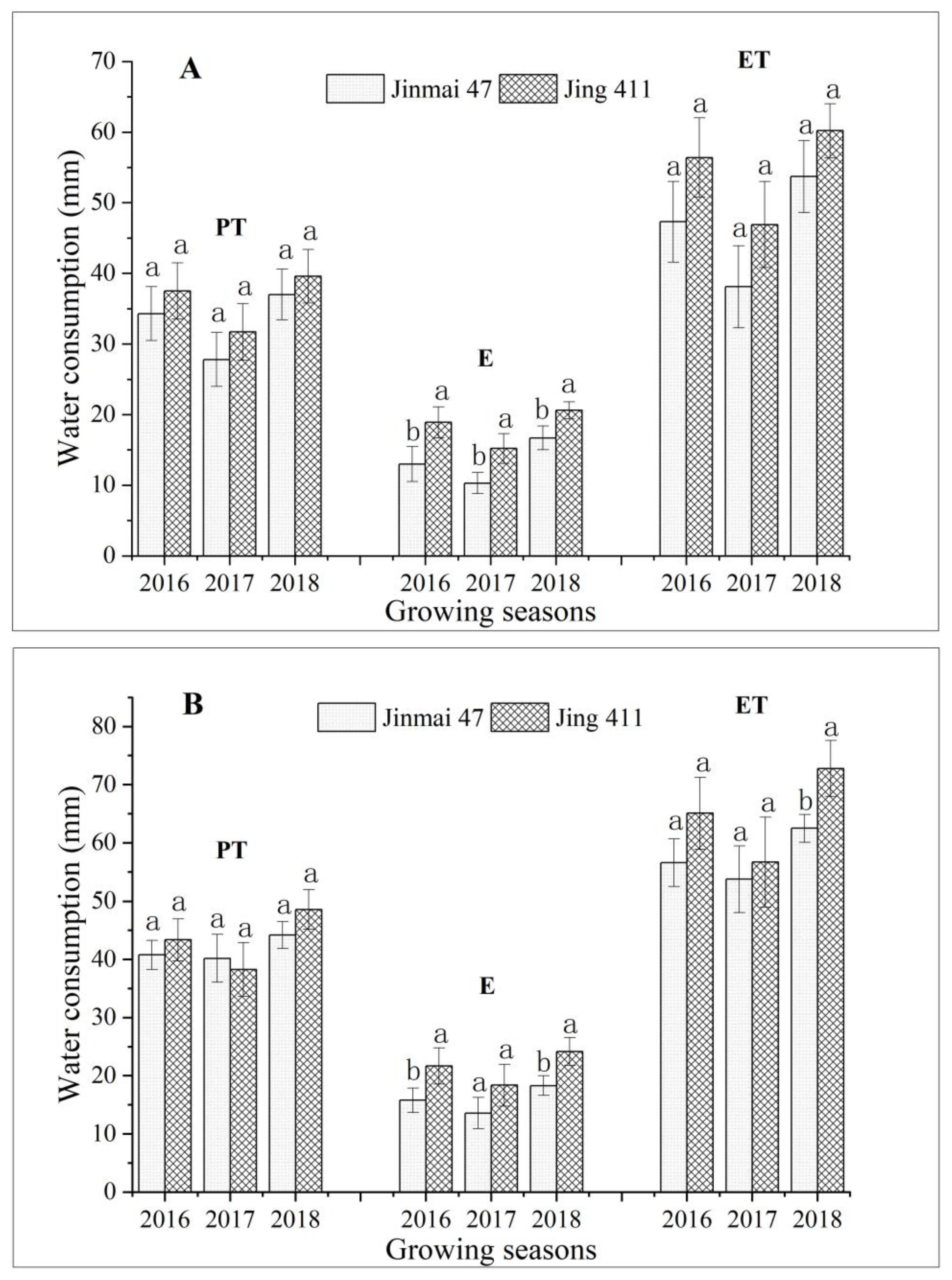
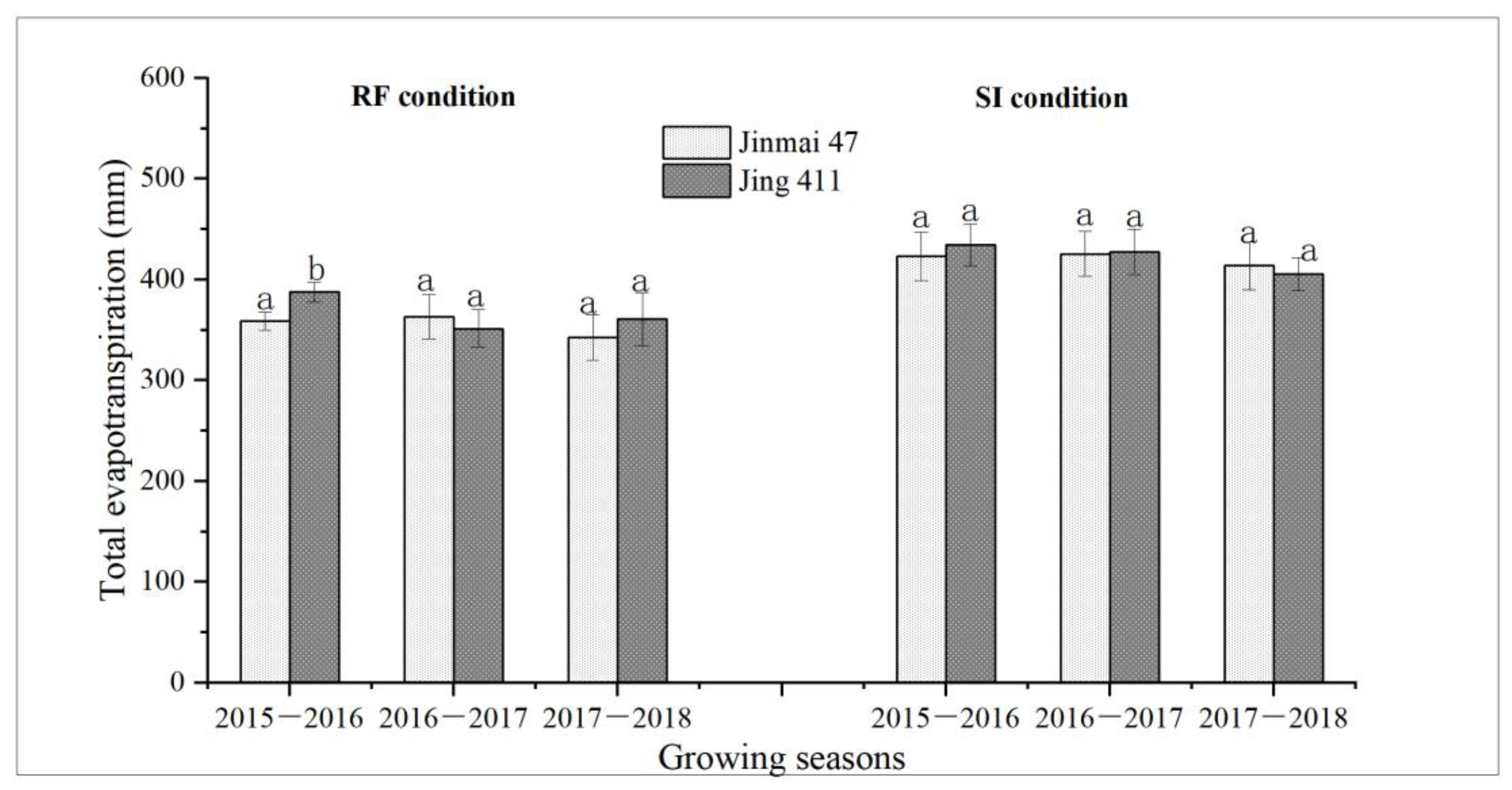
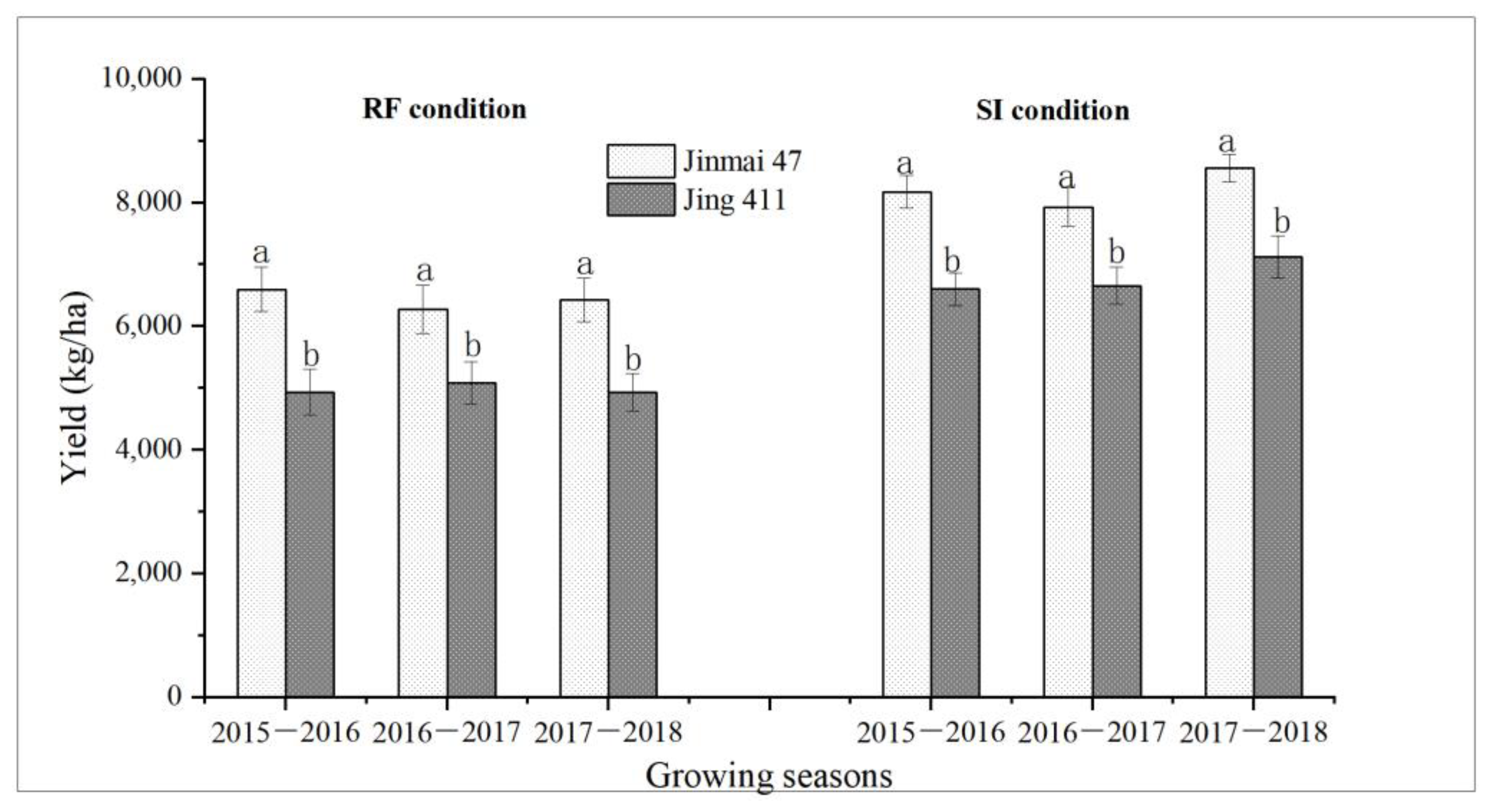
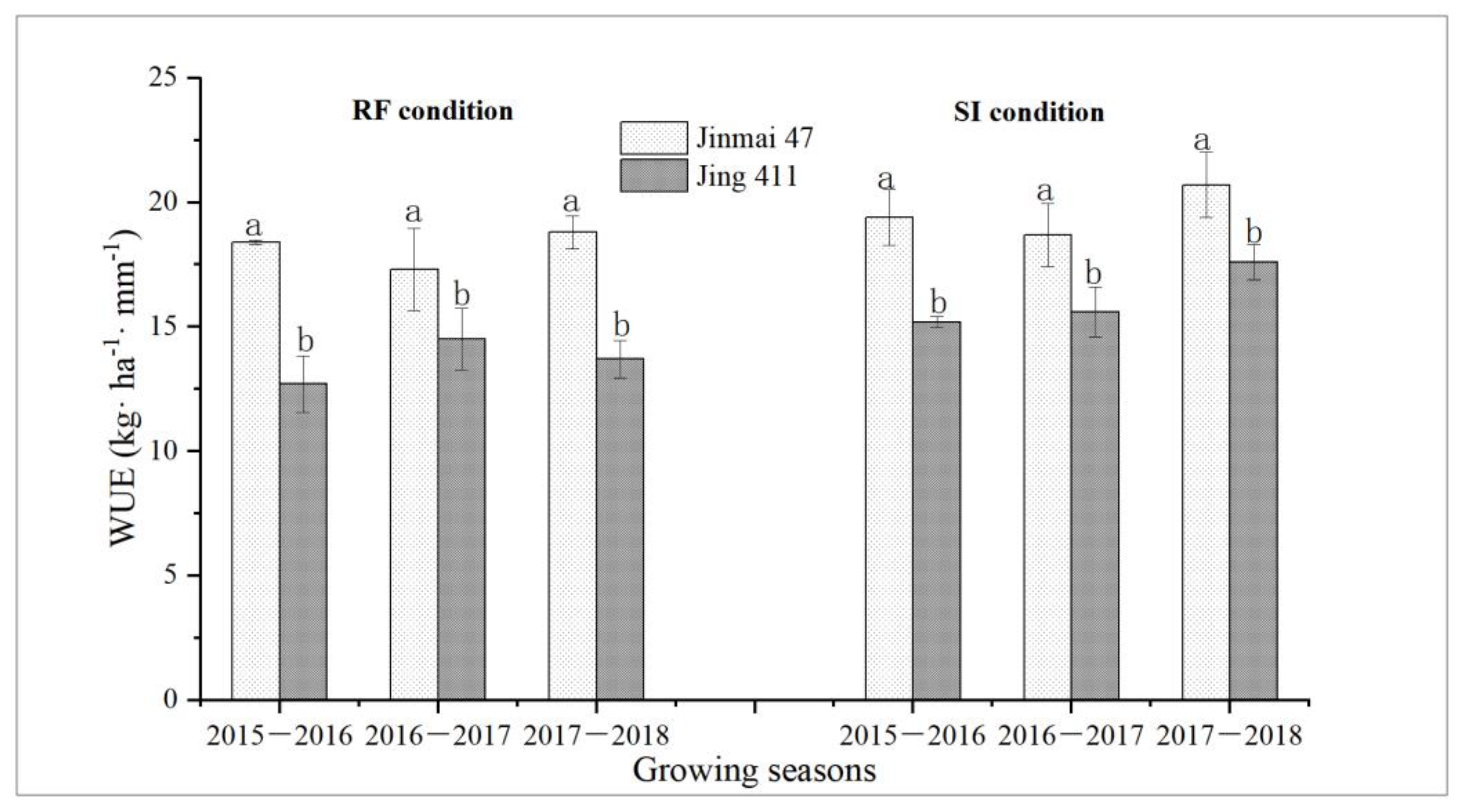
3.4. Water Consumption and Grain Yield of Jinmai 47 and Jing 411
4. Discussion
4.1. Flat-leafed Jinmai 47 Maintained a Cooler Canopy and Conserved Water Relative to the Upright-Leafed Jing 411
4.2. Flat-Leafed Type Jinmai 47 Yielded Higher Than Upright-Leafed Type Jing 411
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- National Bureau of Statistics of China. China Statistical Year-Book; China Statistics Press: Beijing, China, 2021.
- Sun, H.Y.; Liu, C.M.; Zhang, X.Y.; Shen, Y.J.; Zhang, Y.Q. Effects of irrigation on water balance, yield and WUE of winter wheat in the North China Plain. Agric. Water Manag. 2006, 85, 211–218. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Qin, W.L.; Xie, J.N. Improving water use efficiency in grain production of winter wheat and summer maize in the North China Plain: A review. Front. Agr. Sci. Eng. 2016, 3, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Shen, Y. Estimation of agricultural water consumption from meteorological and yield data: A case study of Hebei, North China. PLoS ONE 2013, 8, e58685. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.L.; Chen, Y.Q.; Pacenka, S.; Gao, W.S.; Ma, L.; Wang, G.Y.; Yan, P.; Sui, P.; Steenhuis, T.S. Effect of diversified crop rotations on groundwater levels and crop water productivity in the North China Plain. J. Hydrol. 2015, 522, 428–438. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Qin, W.L.; Chen, S.Y.; Shao, L.W.; Sun, H.Y. Response of yield and WUE of winter wheat to water stress during the past three decades-A case study in the North China Plain. Agric. Water Manag. 2017, 179, 47–54. [Google Scholar] [CrossRef]
- Blum, A. Drought resistance, water-use efficiency, and yield potential-are they compatible, dissonant, or mutually exclusive? Aust. J. Agric. Res. 2005, 56, 1159–1168. [Google Scholar] [CrossRef] [Green Version]
- Fischer, R.A.; Rees, D.; Sayre, K.D.; Lu, Z.M.; Condon, A.G. Wheat yield progress associated with higher stomatal conductance and photosynthetic rate, and cooler canopies. Crop Sci. 1998, 38, 1467–1475. [Google Scholar] [CrossRef]
- Bota, J.; Flexas, J.; Medrano, H. Genetic variability of photosynthesis and water use in Balearic grapevine cultivars. Ann. Appl. Biol. 2001, 138, 353–361. [Google Scholar] [CrossRef]
- Ratnayaka, H.H.; Kincaid, D. Gas exchange and leaf ultrastructure of tinnevelly senna, Cassia angustifolia, under drought and nitrogen stress. Crop Sci. 2005, 45, 840–847. [Google Scholar] [CrossRef]
- Saeidi, M.; Abdoli, M. Effect of Drought Stress during Grain Filling on Yield and Its Components, Gas Exchange Variables, and Some Physiological Traits of Wheat Cultivars. J. Agr. Sci. Tech. Iran 2015, 17, 885–898. [Google Scholar]
- Zhang, X.Y.; Wang, Y.J.; Huang, G.R.; Feng, F.; Liu, X.Y.; Guo, R.; Gu, F.X.; Hu, X.; Yang, Z.G.; Zhong, X.L.; et al. Atmospheric humidity and genotype are key determinants of the diurnal stomatal conductance pattern. J. Agron. Crop Sci. 2020, 206, 161–168. [Google Scholar] [CrossRef]
- Condon, A.G.; Richards, R.A.; Rebetzke, G.J.; Farquhar, G.D. Improving intrinsic water-use efficiency and crop yield. Crop Sci. 2002, 42, 122–131. [Google Scholar] [PubMed]
- Tafesse, E.G.; Warkentin, T.D.; Bueckert, R.A. Canopy architecture and leaf type as traits of heat resistance in pea. Field Crops Res. 2019, 241, 107561. [Google Scholar] [CrossRef]
- Tharakan, P.J.; Volk, T.A.; Nowak, C.A.; Ofezu, G.J. Assessment of canopy structure, light interception, and light-use efficiency of first year regrowth of shrub willow (Salix sp.). BioEnerg. Res. 2008, 1, 229–238. [Google Scholar] [CrossRef]
- Li, W.; Niu, Z.; Chen, H.Y.; Li, D. Characterizing canopy structural complexity for the estimation of maize LAI based on ALS data and UAV stereo images. Int. J. Remote Sens. 2017, 38, 2106–2116. [Google Scholar] [CrossRef]
- Reihardt, D.; Kuhlemerier, C. Plant architecture. EMBO Rep. 2002, 3, 846–851. [Google Scholar] [CrossRef]
- Falster, D.S.; Westoby, M. Leaf size and angle vary widely across species: What consequences for light interception? New Phytol. 2003, 158, 509–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wullschleger, S.D.; Oosterhuis, D.M. Photosynthesis of individual field-grown cotton leaves during ontogeny. Photosynth. Res. 1990, 23, 163–170. [Google Scholar] [CrossRef]
- Marois, J.J.; Wright, D.L.; Wiatrak, P.J.; Vargas, M.A. Effect of row width and nitrogen on cotton morphology and canopy micro-climate. Crop Sci. 2004, 44, 870–877. [Google Scholar] [CrossRef]
- Balota, M.; Payne, W.A.; Evett, S.R.; Peters, T.R. Morphological and physiological traits associated with canopy temperature depression in three closely related wheat lines. Crop Sci. 2008, 48, 1897–1910. [Google Scholar] [CrossRef] [Green Version]
- Thapa, S.; Jessup, K.E.; Pradhan, G.P.; Rudd, J.C.; Liu, S.Y.; Mahan, J.R.; Devkota, R.N.; Baker, J.A.; Xue, Q.W. Canopy temperature depression at grain filling correlates to winter wheat yield in the U.S. Southern High Plains. Field Crop Res. 2018, 217, 11–19. [Google Scholar] [CrossRef]
- Jones, H.G.; Serraj, R.; Loveys, B.R.; Xiong, L.Z.; Wheaton, A.; Price, A.H. Thermal infrared imaging of crop canopies for the remote diagnosis and quantification of plant responses to water stress in the field. Funct. Plant Biol. 2009, 36, 978–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balota, M.; Payne, W.A.; Evett, S.R.; Lazar, M.D. Canopy temperature depression sampling to assess grain yield and genotypic differentiation in winter wheat. Crop Sci. 2007, 47, 1518–1529. [Google Scholar] [CrossRef] [Green Version]
- Khush, G.S. What it will take to feed 5.0 billion rice consumers in 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Q.; Zhou, X.B.; Chen, Y.H.; Yu, S.L. Water consumption characteristics of winter wheat grown using different planting patterns and deficit irrigation regime. Agric. Water Manag. 2012, 105, 8–12. [Google Scholar]
- Chen, S.Y.; Zhang, X.Y.; Sun, H.Y.; Ren, T.S.; Wang, Y.M. Effects of winter wheat row spacing on evapotranpsiration, grain yield and water use efficiency. Agric. Water Manag. 2010, 97, 1126–1132. [Google Scholar] [CrossRef]
- Webber, H.; White, J.W.; Kimball, B.A.; Ewert, F.; Asseng, S.; Rezaei, E.E.; Pinter, J.P.J.; Hatfield, J.L.; Reynolds, M.P.; Behnam, A.B.M.; et al. Physical robustness of canopy temperature models for crop heat stress simulation across environments and production conditions. Field Crops Res. 2018, 216, 75–88. [Google Scholar] [CrossRef] [Green Version]
- Kumari, M.; Pudake, R.N.; Singh, V.P.; Joshi, A.K. Association of stay green trait with canopy temperature depression and yield under terminal heat stress in wheat (Triticum aestivum L.). Euphytica 2013, 190, 87–97. [Google Scholar] [CrossRef]
- Mason, R.E.; Hays, D.B.; Mondal, S.; Ibrahim, M.H.; Basnet, B.R. QTL for yield, yield components and canopy temperature depression in wheat under late sown field conditions. Euphytica 2013, 194, 243–259. [Google Scholar] [CrossRef]
- Mondal, S.; Mason, R.E.; Huggins, T.; Hays, D.B. QTL on wheat (Triticum aestivum L.) chromosomes 1B, 3D and 5A are associated with constitutive production of leaf cuticular wax and may contribute to lower leaf temperatures under heat stress. Euphytica 2015, 201, 123–130. [Google Scholar] [CrossRef]
- Pradhan, G.P.; Xue, Q.; Jessup, K.E.; Rudd, J.C.; Liu, S.; Devkota, R.N.; Mahan, J.R. Cooler canopy contributes to higher yield and drought tolerance in new wheat cultivars. Crop Sci. 2014, 54, 2275–2284. [Google Scholar] [CrossRef]
- Cossani, C.M.; Slafer, G.A.; Savin, R. Do barley and wheat (bread and durum) differ in grain weight stability through seasons and water-nitrogen treatments in a Mediterranean location? Field Crops Res. 2011, 121, 240–247. [Google Scholar] [CrossRef]
- Balota, M.; Green, A.J.; Griffery, C.A.; Pitman, R.; Thomason, W. Genetic gains for physiological traits associated with yield in soft red winter wheat in the Eastern United States from 1919 to 2009. Eur. J. Agron. 2017, 84, 7–83. [Google Scholar] [CrossRef]
- Rebetzke, G.J.; Rattey, A.R.; Farquhar, G.D.; Richards, R.A.; Condon, A.G. Genomic regions for canopy temperature and their genetic association with stomatal conductance and grain yield in wheat. Funct. Plant Biol. 2013, 40, 14–33. [Google Scholar] [CrossRef]
- Johnson, R.C.; Witters, R.E.; Ciha, A.J. Apparent photosynthesis, evapotranspiration, and light penetration in two contrasting hard red winter wheat canopies. Agron. J. 1981, 73, 419–422. [Google Scholar] [CrossRef]
- Holmes, M.G.; Keiller, D.R. Effects of pubescence and waxes on the reflectance of leaves in the ultraviolet and photosynthetic wavebands: A comparison of a range of species. Plant Cell Environ. 2002, 25, 85–93. [Google Scholar] [CrossRef]
- Thapa, S.; Stewart, B.A.; Xue, Q.; Pokhrel, P.; Barkley, T.; Bhandari, M. Growing corn in clumps reduces canopy temperature and improves microclimate. J. Crop Improv. 2016, 30, 614–631. [Google Scholar] [CrossRef]
- Umair, M.; Shen, Y.J.; Qi, Y.Q.; Zhang, Y.C.; Ahmad, A.; Pei, H.W.; Liu, M.Y. Evaluation of the CropSyst model during wheat-maize rotations on the North China Plain for identifying soil evaporation losses. Front. Plant Sci. 2017, 8, 1667. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, H.; Eslick, R.F.; Aase, J.K. Canopy temperature of barley as influenced by morphological characteristics. Agron. J. 1973, 65, 425–428. [Google Scholar] [CrossRef]
- Herbert, T.J. Variation in interception of the direct solar beam by top canopy layers. Ecology 1991, 72, 17–22. [Google Scholar] [CrossRef]
- Herbert, T.J. Random wind-induced leaf orientation-effect upon maximization of whole plant photosynthesis. Photosynthetica 1992, 26, 601–607. [Google Scholar]
- Prasad, P.V.V.; Pisipati, S.R.; Ristic, Z.; Bukovnik, U.; Fritz, A.K. Impact of Nighttime Temperature on Physiology and Growth of Spring Wheat. Crop Sci. 2008, 48, 2372–2380. [Google Scholar] [CrossRef]
- Murchie, E.H.; Chen, Y.Z.; Hubbart, S.; Peng, S.B.; Horton, P. Interactions between senescence and leaf orientation determine in situ patterns of photosynthesis and photoinhibition in field-grown rice. Plant Physiol. 1999, 119, 553–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, M.S.; Reynolds, M.P.; Jalal-Kamali, M.R.; Moussa, M.; Feltaous, Y.; Tahir, I.S.A.; Barma, N.; Vargas, M.; Mannes, Y.; Baum, M. The yield correlations of selectable physiological traits in a population of advanced spring wheat lines grown in warm and drought environments. Field Crops Res. 2012, 128, 129–136. [Google Scholar] [CrossRef]
- Purushothaman, R.; Thudi, M.; Krishnamurthy, L.; Upadhyaya, H.D.; Kashiwagi, J.; Gowda, C.L.L.; Varshney, R.K. Association of mid-reproductive stage canopy temperature depression with the molecular markers and grain yields of chickpea (Cicerarietinum L.) germplasm under terminal drought. Field Crops Res. 2015, 174, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Zhang, Z.; Tao, F.L.; Palosuo, T.; Rötter, R.P. Impacts of heat stress on leaf area index and growth duration of winter wheat in the North China Plain. Field Crops Res. 2018, 222, 230–237. [Google Scholar] [CrossRef] [Green Version]
- de Wit, C.T. Transpiration and Crop Yields; Institute of Biological and Chemical Research on Field Crops and Herbage: Wageningen, The Netherlands, 1958; Volume 64.6, p. 24. [Google Scholar]
- Blum, A. Effective use of water (EUW) and not water-use efficiency (WUE) is the target of crop yield improvement under drought stress. Field Crops Res. 2009, 112, 119–123. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Growing Seasons | Oct. | Nov. | Dec. | Jan. | Feb. | Mar. | Apr. | May. | Jun. | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Rainfall (mm) | 2015–2016 | 8.7 | 42.9 | 2.3 | 1.2 | 9.2 | 0 | 5.6 | 56.9 | 46.6 | 173.4 |
| 2016–2017 | 70.8 | 6.5 | 0 | 0.8 | 3.7 | 11.6 | 0.1 | 23.1 | 3 | 119.6 | |
| 2017–2018 | 58.8 | 0 | 0 | 0 | 0 | 1.7 | 52.7 | 13.5 | 3.4 | 130.1 | |
| Long-term average (1990–2018) | 27.2 | 12.9 | 2 | 2.3 | 4.7 | 9.7 | 20.6 | 35.9 | 28.5 | 143.8 | |
| Mean temperature (°C) | 2015–2016 | 13.7 | 3 | −0.6 | −4.8 | 0.6 | 8.7 | 16.4 | 21.2 | 25.2 | _ |
| 2016–2017 | 13 | 4 | −0.4 | −2 | 1.7 | 8.4 | 17.2 | 23.2 | 25.6 | _ | |
| 2017–2018 | 12.7 | 3.4 | −1.4 | −3.9 | −1.6 | 8.1 | 15.6 | 22.1 | 26.8 | _ | |
| Average of the three seasons | 13.1 | 3.5 | −0.8 | −3.6 | 0.2 | 8.4 | 16.4 | 22.2 | 25.9 | _ | |
| Long-term average (1990–2018) | 13.1 | 4.4 | −1.7 | −3.7 | −0.1 | 6.8 | 14.7 | 20.9 | 24.8 | _ |
| Genotype | Plant Type | Leaf Type | Leaf Color | SPAD Reading | Flag Leaf Inclination Angle (°) | Flag Leaf Area (cm2) | Plant Height (cm) | Basal Internode Diameter (mm) |
|---|---|---|---|---|---|---|---|---|
| Jinmai 47 | Loose type | Flat leaf | Bright green | 48.3 ± 0.8 | 65 ± 4.6 | 19.3 ± 1.5 | 80.5 ± 3.1 | 3.2 ± 0.2 |
| Jing 411 | Compact type | Upright leaf | Dark green | 55.1 ± 1.0 | 35 ± 4.4 | 16.3 ± 1.3 | 69.9 ± 3.0 | 4.8 ± 0.3 |
| Genotype | 2015–2016 | 2016–2017 | 2017–2018 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Pn | Gs | Tr | Pn | Gs | Tr | Pn | Gs | Tr | |
| RF condition | |||||||||
| Jinmai 47 | 17.9 a | 0.312 a | 5.86 a | 16.9 a | 0.279 a | 5.22 a | 20.7 a | 0.422 a | 6.4 a |
| Jing 411 | 15.2 b | 0.249 b | 4.56 b | 13.9 b | 0.264 a | 4.15 a | 16.7 b | 0.327 b | 4.34 b |
| P > F | 0.013 | 0.0297 | 0.0325 | 0.0454 | 0.346 | 0.4394 | 0.0463 | 0.016 | 0.0112 |
| SI condition | |||||||||
| Jinmai 47 | 24.3 a | 0.505 a | 8.24 a | 22.1 a | 0.445 a | 7.33 a | 25.4 a | 0.589 a | 8.76 a |
| Jing 411 | 19.8 b | 0.331 b | 6.22 b | 18.6 b | 0.274 b | 5.73 b | 22.2 a | 0.466 b | 7.21 b |
| P > F | 0.0363 | 0.0009 | 0.0039 | 0.0306 | 0.0007 | 0.0145 | 0.1444 | 0.0061 | 0.0165 |
| Genotype | 2015–2016 | 2016–2017 | 2017–2018 | |||
|---|---|---|---|---|---|---|
| 15 April | 26 April | 20 April | 30 April | 24 April | 9 May | |
| RF condition | ||||||
| Jinmai 47 | 7.2 a | 5.9 a | 6.2 a | 5 a | 7.8 a | 6 a |
| Jing 411 | 5.6 b | 4.4 b | 5.4 b | 4.7 a | 6.6 b | 4.6 b |
| P > F | 0.0023 | 0.0044 | 0.0184 | 0.3658 | 0.0351 | 0.0056 |
| SI condition | ||||||
| Jinmai 47 | 8.7 a | 7 a | 6.8 a | 5.6 a | 8 a | 5.9 a |
| Jing 411 | 7.4 b | 5.6 b | 5.7 b | 5 a | 6.8 b | 5.5 a |
| P > F | 0.0354 | 0.0135 | 0.0309 | 0.0602 | 0.0097 | 0.3005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, G.; Zhang, X.; Wang, Z.; Li, Y.; Liu, X.; Guo, R.; Gu, F.; Liu, E.; Li, S.; Zhong, X.; et al. Plant Architecture Influences the Population Transpiration and Canopy Temperature in Winter Wheat Genotypes. Agronomy 2023, 13, 742. https://doi.org/10.3390/agronomy13030742
Huang G, Zhang X, Wang Z, Li Y, Liu X, Guo R, Gu F, Liu E, Li S, Zhong X, et al. Plant Architecture Influences the Population Transpiration and Canopy Temperature in Winter Wheat Genotypes. Agronomy. 2023; 13(3):742. https://doi.org/10.3390/agronomy13030742
Chicago/Turabian StyleHuang, Guirong, Xinying Zhang, Zhenzhao Wang, Yuzhong Li, Xiaoying Liu, Rui Guo, Fengxue Gu, Enke Liu, Shuying Li, Xiuli Zhong, and et al. 2023. "Plant Architecture Influences the Population Transpiration and Canopy Temperature in Winter Wheat Genotypes" Agronomy 13, no. 3: 742. https://doi.org/10.3390/agronomy13030742