
Genome-Wide Characterization of the SAMS Gene Family in Cotton Unveils the Putative Role of GhSAMS2 in Enhancing Abiotic Stress Tolerance
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Identification and Physiochemical Analysis of SAMS Genes in Gossypium hirsutum
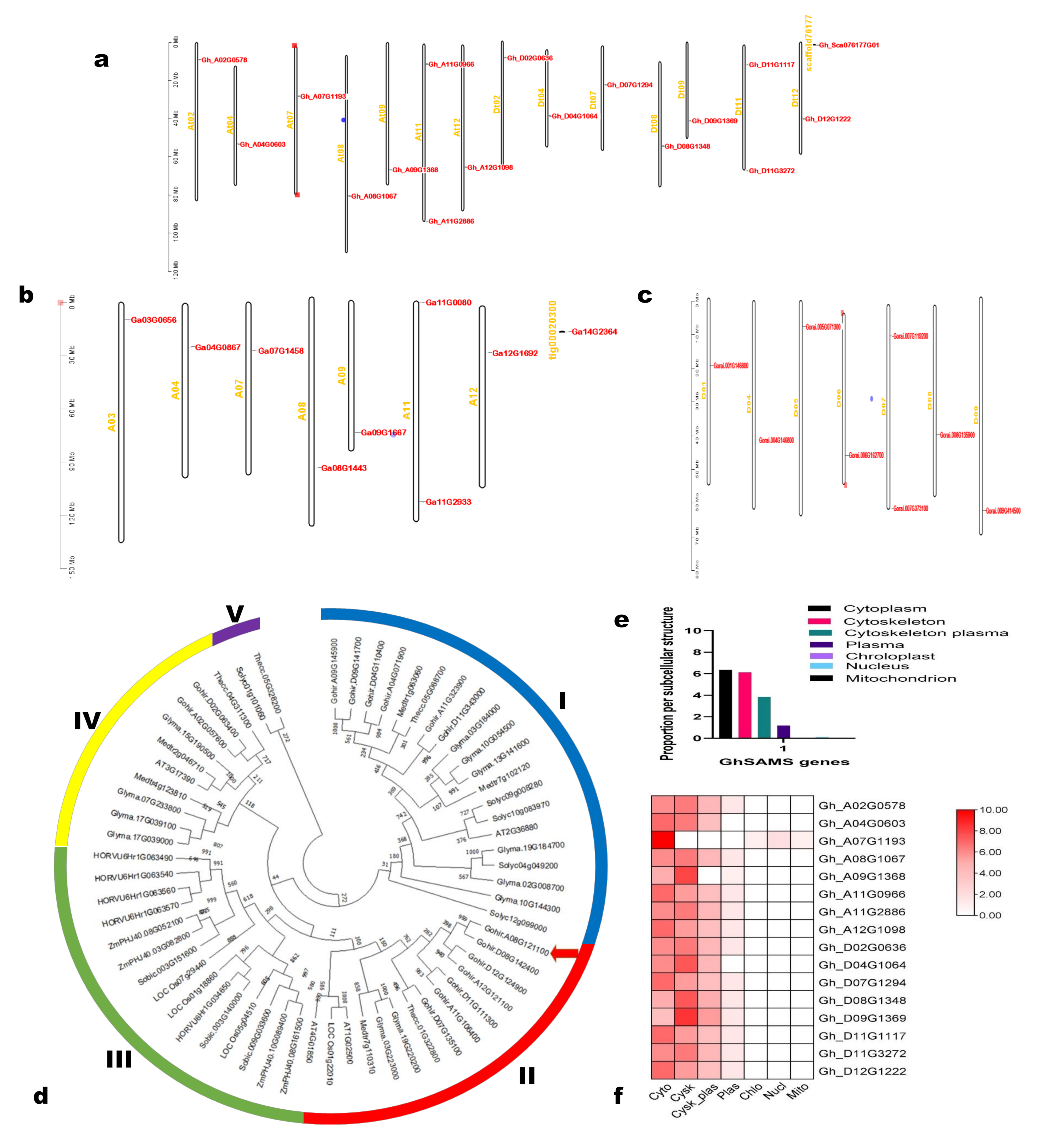
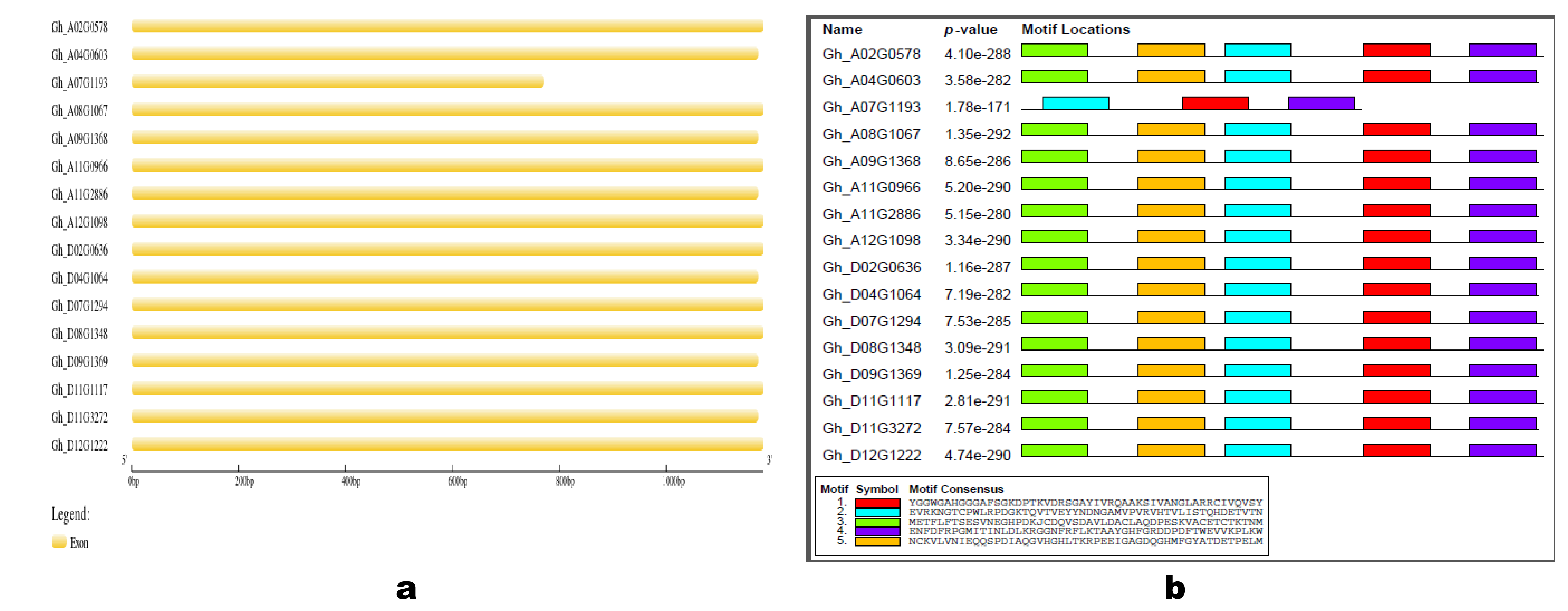
2.2. Chromosomal Location, Phylogenetic Analysis, Prediction of Subcellular Localization, Gene Structure, Cis-Acting Elements, and Conserved Motifs Analyses
2.3. Plant Materials and Treatments
2.4. RNA Extraction and RT-qPCR Assays
2.5. Identification of Pray Proteins
2.6. Construction of Yeast Two-Hybrid Library, Bait Cloning, and Auto Activation Analysis
2.7. cDNA Libraries Screening and Yeast Two-Hybrid Interaction Assay
2.8. Virus-Induced Gene Silencing of GhSAMS2 in G. hirsutum and Stress Treatments
2.9. Measurement of the Physiological and Morphological Parameters
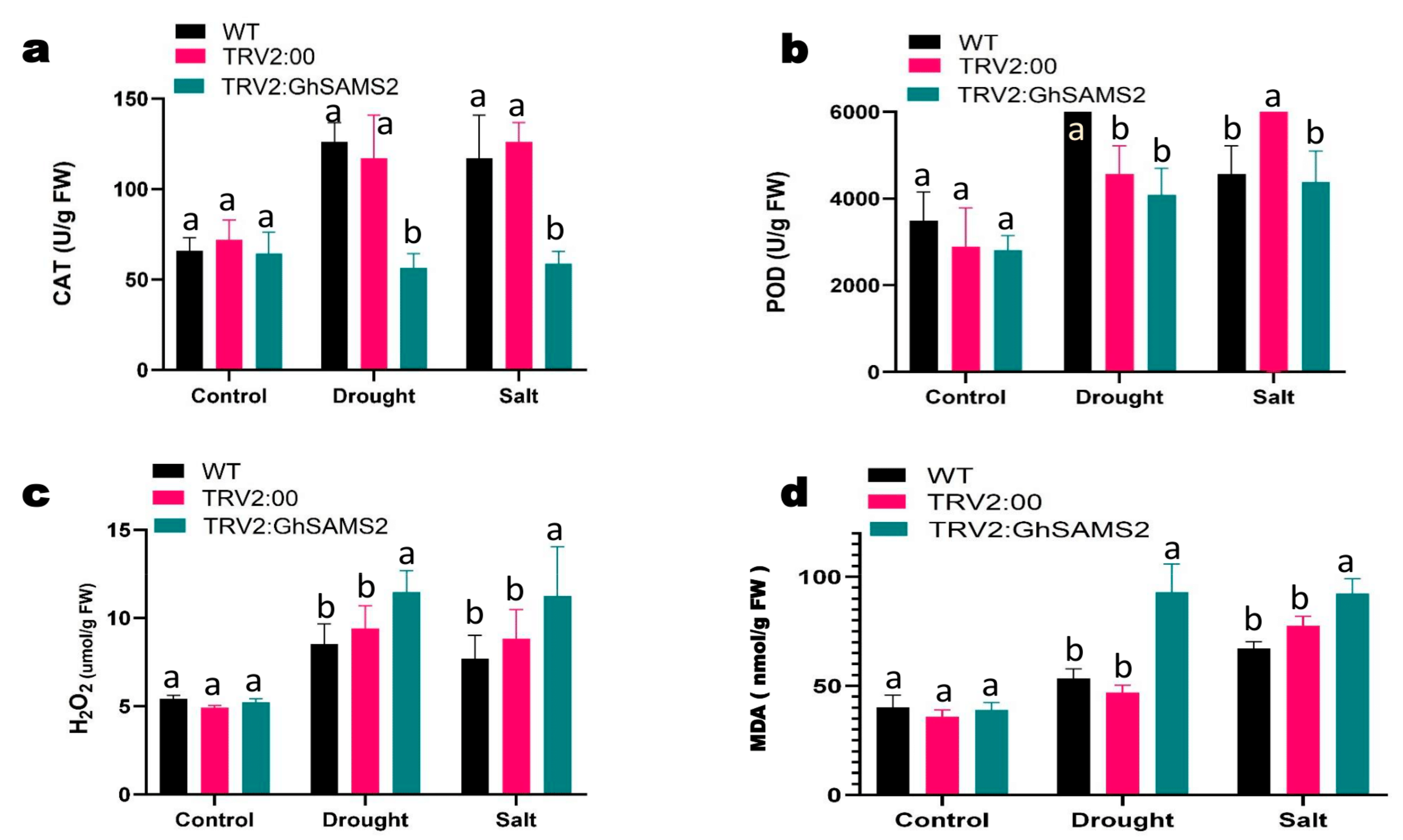
2.10. Estimation of Oxidant and Antioxidant Enzyme Activities
2.11. Statistical Analysis
3. Results
3.1. Identification, Physiochemical Properties, Chromosomal Distribution, Phylogenetic Analysis, and Subcellular Localization
3.2. Gene Structure, Conserved Motifs, and Cis-Acting Elements Analyses
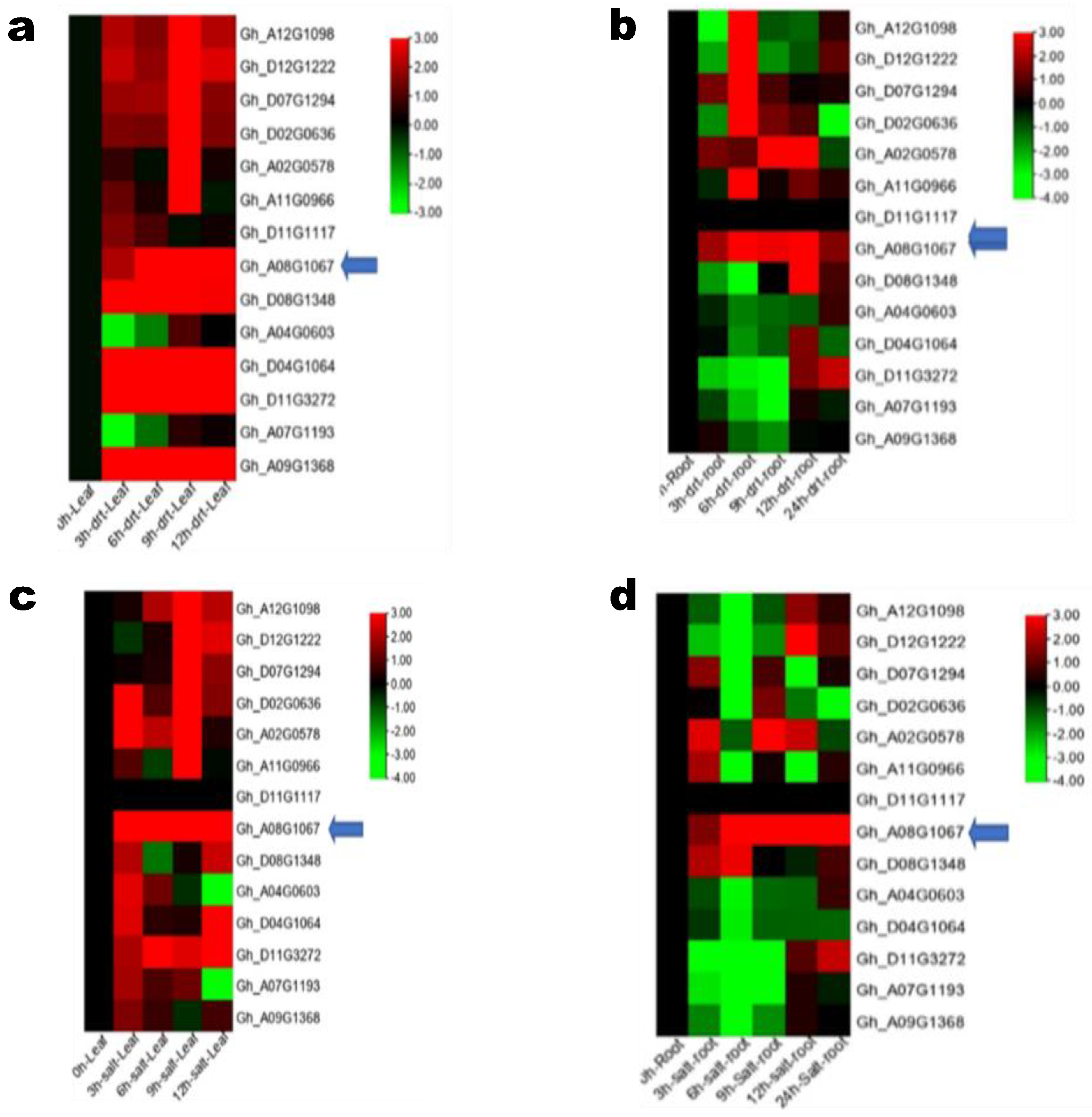
3.3. GhSAMS Genes Expression under Drought and Salt Stress
3.4. Identification of CBL10 Interacting Proteins from the Cotton AD Library under Drought and Salt Stress Using the Y2H System
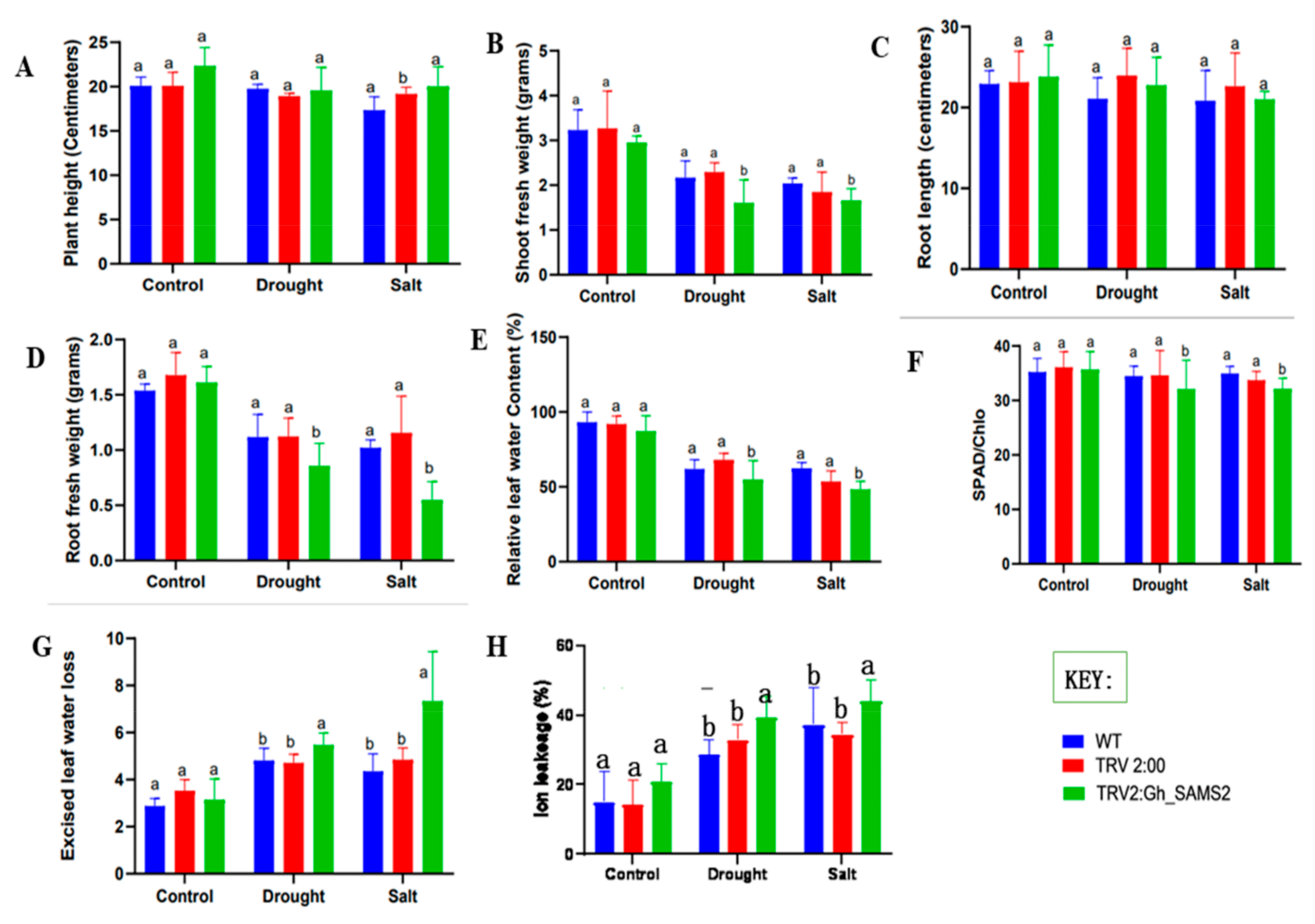
3.5. GhSAMS2 Gene Silencing Significantly Increased Sensitivity to Drought and Salt Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gu, Z.; Huang, C.; Li, F.; Zhou, X. A versatile system for functional analysis of genes and microRNAs in cotton. Plant Biotechnol. J. 2014, 12, 638–649. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Kim, S.H.; Palaniyandi, S.A.; Yang, S.H.; Suh, J.W. Expression of potato S-adenosyl-l-methionine synthase (SbSAMS) gene altered developmental characteristics and stress responses in transgenic Arabidopsis plants. Plant Physiol. Biochem. 2015, 87, 84–91. [Google Scholar] [CrossRef]
- Mehari, T.G.; Xu, Y.; Umer, M.J.; Shiraku, M.L.; Hou, Y.; Wang, Y.; Yu, S.; Zhang, X.; Wang, K.; Cai, X.; et al. Multi-Omics-Based Identification and Functional Characterization of Gh_A06G1257 Proves Its Potential Role in Drought Stress Tolerance in Gossypium hirsutum. Front. Plant Sci. 2021, 12, 2092. [Google Scholar] [CrossRef]
- Turan, M.A.; Elkarim, A.H.A.; Taban, N.; Taban, S. Effect of salt stress on growth, stomatal resistance, proline and chlorophyll concentrations on maize plant. Afr. J. Agric. Res. 2009, 4, 893–897. [Google Scholar]
- Shi, D.; Sheng, Y. Effect of various salt-alkaline mixed stress conditions on sunflower seedlings and analysis of their stress factors. Environ. Exp. Bot. 2005, 54, 8–21. [Google Scholar] [CrossRef]
- Yang, C.W.; Xu, H.H.; Wang, L.L.; Liu, J.; Shi, D.C.; Wang, D.L. Comparative effects of salt-stress and alkali-stress on the growth, photosynthesis, solute accumulation, and ion balance of barley plants. Photosynthetica 2009, 47, 79–86. [Google Scholar] [CrossRef]
- Ahmad, S.T.; Sima, N.A.K.K.; Mirzaei, H.H. Effects of sodium chloride on physiological aspects of Salicornia persica growth. J. Plant Nutr. 2013, 36, 401–414. [Google Scholar] [CrossRef]
- Ahuja, I.; de Vos, R.C.H.; Bones, A.M.; Hall, R.D. Plant molecular stress responses face climate change. Trends Plant Sci. 2010, 15, 664–674. [Google Scholar] [CrossRef]
- Yang, L.; Zhang, Y.; Zhu, N.; Koh, J.; Ma, C.; Pan, Y.; Yu, B.; Chen, S.; Li, H. Proteomic analysis of salt tolerance in sugar beet monosomic addition line M14. J. Proteome Res. 2013, 12, 4931–4950. [Google Scholar] [CrossRef]
- He, M.W.; Wang, Y.; Wu, J.Q.; Shu, S.; Sun, J.; Guo, S.R. Isolation and characterization of S-Adenosylmethionine synthase gene from cucumber and responsive to abiotic stress. Plant Physiol. Biochem. 2019, 141, 431–445. [Google Scholar] [CrossRef]
- Heidari, P.; Mazloomi, F.; Nussbaumer, T.; Barcaccia, G. Insights into the SAM Synthetase Gene Family and Its Roles in Tomato Seedlings under Abiotic Stresses and Hormone Treatments. Plants 2020, 9, 586. [Google Scholar] [CrossRef]
- Roje, S. S-Adenosyl-l-methionine: Beyond the universal methyl group donor. Phytochemistry 2006, 67, 1686–1698. [Google Scholar] [CrossRef]
- Bürstenbinder, K.; Rzewuski, G.; Wirtz, M.; Hell, R.; Sauter, M. The role of methionine recycling for ethylene synthesis in Arabidopsis. Plant J. 2007, 49, 238–249. [Google Scholar] [CrossRef]
- Ma, C.; Wang, Y.; Gu, D.; Nan, J.; Chen, S.; Li, H. Overexpression of S-adenosyl-L-methionine synthetase 2 from sugar beet M14 increased arabidopsis tolerance to salt and oxidative stress. Int. J. Mol. Sci. 2017, 18, 847. [Google Scholar] [CrossRef] [Green Version]
- Jang, S.J.; Wi, S.J.; Choi, Y.J.; An, G.; Park, K.Y. Increased polyamine biosynthesis enhances stress tolerance by preventing the accumulation of reactive oxygen species: T-DNA mutational analysis of Oryza sativa lysine decarboxylase-like protein 1. Mol. Cells 2012, 34, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Gupta, K.; Dey, A.; Gupta, B. Plant polyamines in abiotic stress responses. Acta Physiol. Plant. 2013, 35, 2015–2036. [Google Scholar] [CrossRef]
- Wang, K.L.C.; Li, H.; Ecker, J.R. Ethylene biosynthesis and signaling networks. Plant Cell 2002, 14, 131–152. [Google Scholar] [CrossRef] [Green Version]
- Vandenbussche, F.; Vaseva, I.; Vissenberg, K.; Van Der Straeten, D. Ethylene in vegetative development: A tale with a riddle. New Phytol. 2012, 194, 895–909. [Google Scholar] [CrossRef]
- Li, W.; Han, Y.; Tao, F.; Chong, K. Knockdown of SAMS genes encoding S-adenosyl-l-methionine synthetases causes methylation alterations of DNAs and histones and leads to late flowering in rice. J. Plant Physiol. 2011, 168, 1837–1843. [Google Scholar] [CrossRef]
- Chen, M.; Chen, J.; Fang, J.; Guo, Z.; Lu, S. Down-regulation of S-adenosylmethionine decarboxylase genes results in reduced plant length, pollen viability, and abiotic stress tolerance. Plant Cell Tissue Organ Cult. 2014, 116, 311–322. [Google Scholar] [CrossRef]
- Guo, Z.; Tan, J.; Zhuo, C.; Wang, C.; Xiang, B.; Wang, Z. Abscisic acid, H2O2 and nitric oxide interactions mediated cold-induced S-adenosylmethionine synthetase in Medicago sativa subsp. Falcata that confers cold tolerance through up-regulating polyamine oxidation. Plant Biotechnol. J. 2014, 12, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; He, M.; Jahan, M.S.; Wu, J.; Gu, Q.; Shu, S.; Sun, J.; Guo, S. Cscdpk6, a cssams1-interacting protein, affects polyamine/ethylene biosynthesis in cucumber and enhances salt tolerance by overexpression in tobacco. Int. J. Mol. Sci. 2021, 22, 11133. [Google Scholar] [CrossRef]
- Ding, C.; Chen, T.; Yang, Y.; Liu, S.; Yan, K.; Yue, X.; Zhang, H.; Xiang, Y.; An, L.; Chen, S. Molecular cloning and characterization of an S-adenosylmethionine synthetase gene from Chorispora bungeana. Gene 2015, 572, 205–213. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Magwanga, R.O.; Yang, X.; Jin, D.; Cai, X.; Hou, Y.; Wei, Y.; Zhou, Z.; Wang, K.; Liu, F. Genetic regulatory networks for salt-alkali stress in Gossypium hirsutum with differing morphological characteristics. BMC Genom. 2020, 21, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. The Proteomics Protocols Handbook; Humana Press: Totowa, NJ, USA, 2005; pp. 571–608. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Gibson, T.J.; Higgins, D.G. Multiple Sequence Alignment Using ClustalW and ClustalX. Curr. Protoc. Bioinform. 2003, 1, 2–3. [Google Scholar] [CrossRef]
- Hortona, P.; Park, K.J.; Obayashi, T.; Nakai, K. Protein subcellular localization prediction with WoLF PSORT. Ser. Adv. Bioinform. Comput. Biol. 2006, 3, 39–48. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.; Guo, A.Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Mehari, T.G.; Xu, Y.; Magwanga, R.O.; Umer, M.J.; Kirungu, J.N.; Cai, X.; Hou, Y.; Wang, Y.; Yu, S.; Wang, K.; et al. Genome wide identification and characterization of light-harvesting Chloro a/b binding (LHC) genes reveals their potential role in enhancing drought tolerance in Gossypium hirsutum. J. Cott. Res. 2021, 4, 15. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. Preparing the nutrient solution. In The Water-Culture Method for Growing Plants Without Soil; Circular California Agricultural Experiment Station: Berkeley, CA, USA, 1950; Volume 347, pp. 29–31. [Google Scholar]
- Mehari, T.G.; Xu, Y.; Magwanga, R.O.; Umer, M.J.; Shiraku, M.L.; Hou, Y.; Wang, Y.; Wang, K.; Cai, X.; Zhou, Z.; et al. Identification and functional characterization of Gh_D01G0514 (GhNAC072) transcription factor in response to drought stress tolerance in cotton. Plant Physiol. Biochem. 2021, 166, 361–375. [Google Scholar] [CrossRef]
- Magwanga, R.O.; Lu, P.; Kirungu, J.N.; Lu, H.; Wang, X.; Cai, X.; Zhou, Z.; Zhang, Z.; Salih, H.; Wang, K.; et al. Characterization of the late embryogenesis abundant (LEA) proteins family and their role in drought stress tolerance in upland cotton. BMC Genet. 2018, 19, 1–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mustafa, R.; Shafiq, M.; Mansoor, S.; Briddon, R.W.; Scheffler, B.E.; Scheffler, J.; Amin, I. Virus-Induced Gene Silencing in Cultivated Cotton (Gossypium spp.) Using Tobacco Rattle Virus. Mol. Biotechnol. 2016, 58, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Kirungu, J.N.; Magwanga, R.O.; Pu, L.; Cai, X.; Xu, Y.; Hou, Y.; Zhou, Y.; Cai, Y.; Hao, F.; Zhou, Z.; et al. Knockdown of Gh_A05G1554 (GhDHN_03) and Gh_D05G1729 (GhDHN_04) Dehydrin genes, Reveals their potential role in enhancing osmotic and salt tolerance in cotton. Genomics 2020, 112, 1902–1915. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Shiraku, M.L.; Magwanga, R.O.; Cai, X.; Kirungu, J.N.; Xu, Y.; Mehari, T.G.; Hou, Y.; Wang, Y.; Wang, K.; Peng, R.; et al. Knockdown of 60S ribosomal protein L14-2 reveals their potential regulatory roles to enhance drought and salt tolerance in cotton. J. Cott. Res. 2021, 4, 27. [Google Scholar] [CrossRef]
- Chen, Y.; Li, C.; Zhang, B.; Yi, J.; Yang, Y.; Kong, C.; Lei, C.; Gong, M. The Role of the Late Embryogenesis-Abundant (LEA) Protein Family in Development and the Abiotic Stress Response: A Comprehensive Expression Analysis of. Genes 2019, 10, 148. [Google Scholar] [CrossRef] [Green Version]
- Lei, K.; Liu, A.; Fan, S.; Peng, H.; Zou, X.; Zhen, Z. Identification of TPX2 Gene Family in Upland Cotton and its Functional Analysis in Cotton. Genes 2019, 10, 508. [Google Scholar] [CrossRef] [Green Version]
- Fu, D.Q.; Zhu, B.Z.; Zhu, H.L.; Jiang, W.B.; Luo, Y.B. Virus-induced gene silencing in tomato fruit. Plant J. 2005, 43, 299–308. [Google Scholar] [CrossRef]
- Gao, X.; Britt, R.C.; Shan, L.; He, P. Agrobacterium-mediated virus-induced gene silencing assay in cotton. J. Vis. Exp. 2011, 54, e2938. [Google Scholar] [CrossRef] [Green Version]
- Dupadahalli, K. A modified freeze-thaw method for the efficient transformation of Agrobacterium tumefaciens. Curr. Sci. 2007, 93, 770–772. [Google Scholar]
- Tuttle, J.R.; Haigler, C.H.; Robertson, D. Method: Low-cost delivery of the cotton leaf crumple virus-induced gene silencing system. Plant Methods 2012, 8, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirungu, J.N.; Magwanga, R.O.; Lu, P.; Cai, X.; Zhou, Z.; Wang, X.; Peng, R.; Wang, K.; Liu, F. Functional characterization of Gh-A08G1120 (GH3.5) gene reveal their significant role in enhancing drought and salt stress tolerance in cotton. BMC Genet. 2019, 20, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Jawdy, S.; Tschaplinski, T.J.; Tuskan, G.A. Genome-wide identification of lineage-specific genes in Arabidopsis, Oryza and Populus. Genomics 2009, 93, 473–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magwanga, R.O.; Kirungu, J.N.; Lu, P.; Yang, X.; Dong, Q.; Cai, X.; Xu, Y.; Wang, X.; Zhou, Z.; Hou, Y.; et al. Genome wide identification of the trihelix transcription factors and overexpression of Gh_A05G2067 (GT-2), a novel gene contributing to increased drought and salt stresses tolerance in cotton. Physiol. Plant. 2019, 167, 447–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarie, S.; Hanaoka, N.; Kubota, F.; Agata, W.; Kaufman, P.B. Measurement of Cell Membrane Stability Evaluated by Electrolyte Leakage as a Drought and Heat Tolerance Test in Rice (Oryza sativa L.). J. Fac. Agric. Kyushu Univ. 1995, 40, 233–240. [Google Scholar] [CrossRef]
- Rehman, S.U.; Bilal, M.; Rana, R.M.; Tahir, M.N.; Shah, M.K.N.; Ayalew, H.; Yan, G. Cell membrane stability and chlorophyll content variation in wheat (Triticum aestivum) genotypes under conditions of heat and drought. Crop Pasture Sci. 2016, 67, 712–718. [Google Scholar] [CrossRef]
- Magwanga, R.O.; Lu, P.; Kirungu, J.N.; Dong, Q.; Cai, X.; Zhou, Z.; Wang, X.; Hou, Y.; Xu, Y.; Peng, R.; et al. Knockdown of cytochrome P450 genes Gh_D07G1197 and Gh_A13G2057 on chromosomes D07 and A13 reveals their putative role in enhancing drought and salt stress tolerance in Gossypium hirsutum. Genes 2019, 10, 226. [Google Scholar] [CrossRef] [Green Version]
- Majidi, M.M.; Rashidi, F.; Sharafi, Y. Physiological traits related to drought tolerance in Brassica. Int. J. Plant Prod. 2015, 9, 541–560. [Google Scholar] [CrossRef]
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of salinity stress on plants and its tolerance strategies: A review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef]
- Flowers, T.J. Improving crop salt tolerance. J. Exp. Bot. 2004, 55, 307–319. [Google Scholar] [CrossRef]
- Hazarika, P.; Rajam, M.V. Biotic and abiotic stress tolerance in transgenic tomatoes by constitutive expression of S-adenosylmethionine decarboxylase gene. Physiol. Mol. Biol. Plants 2011, 17, 115–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhandari, B.; Roesler, W.J.; DeLisio, K.D.; Klemm, D.J.; Ross, N.S.; Miller, R.E. A functional promoter flanks an intronless glutamine synthetase gene. J. Biol. Chem. 1991, 266, 7784–7792. [Google Scholar] [CrossRef]
- He, S.; Zou, M.; Guo, B. The roles and evolutionary patterns of intronless genes in deuterostomes. Comp. Funct. Genom. 2011, 2011, 680673. [Google Scholar] [CrossRef] [Green Version]
- Shiraku, M.L.; Magwanga, R.O.; Cai, X.; Kirungu, J.N.; Xu, Y.; Mehari, T.G.; Hou, Y.; Wang, Y.; Agong, S.G.; Peng, R.; et al. Functional Characterization of GhACX3 Gene Reveals Its Significant Role in Enhancing Drought and Salt Stress Tolerance in Cotton. Front. Plant Sci. 2021, 12, 1120. [Google Scholar] [CrossRef]
- Zhou, M.L.; Ma, J.T.; Pang, J.F.; Zhang, Z.L.; Tang, Y.X.; Wu, Y.M. Regulation of plant stress response by dehydration responsive element binding (DREB) transcription factors. Afr. J. Biotechnol. 2010, 9, 9255–9269. [Google Scholar] [CrossRef]
- Gao, J.; Lan, T. Functional characterization of the late embryogenesis abundant (LEA) protein gene family from Pinus tabuliformis (Pinaceae) in Escherichia coli. Sci. Rep. 2016, 6, 19467. [Google Scholar] [CrossRef] [Green Version]
- Tunnacliffe, A.; Wise, M.J. The continuing conundrum of the LEA proteins. Naturwissenschaften 2007, 94, 791–812. [Google Scholar] [CrossRef]
- Chakrabortee, S.; Boschetti, C.; Walton, L.J.; Sarkar, S.; Rubinsztein, D.C.; Tunnacliffe, A. Hydrophilic protein associated with desiccation tolerance exhibits broad protein stabilization function. Proc. Natl. Acad. Sci. USA 2007, 104, 18073–18078. [Google Scholar] [CrossRef] [Green Version]
- Heidari, P.; Ahmadizadeh, M.; Izanlo, F.; Nussbaumer, T. In silico study of the CESA and CSL gene family in Arabidopsis thaliana and Oryza sativa: Focus on post-translation modifications. Plant Gene 2019, 19, 100189. [Google Scholar] [CrossRef]
- Bachmair, A.; Finley, D.; Varshavsky, A. In vivo half-life of a protein is a function of its amino-terminal residue. Science 1986, 234, 179–186. [Google Scholar] [CrossRef]
- Gong, B.; Li, X.; Vandenlangenberg, K.M.; Wen, D.; Sun, S.; Wei, M.; Li, Y.; Yang, F.; Shi, Q.; Wang, X. Overexpression of S-adenosyl-l-methionine synthetase increased tomato tolerance to alkali stress through polyamine metabolism. Plant Biotechnol. J. 2014, 12, 694–708. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Kukreja, S.; Salaria, N.; Thakur, K.; Gautam, S.; Taunk, J.; Goutam, U. VIGS: A flexible tool for the study of functional genomics of plants under abiotic stresses. J. Crop Improv. 2019, 33, 567–604. [Google Scholar] [CrossRef]
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and Salinity Stress Responses and Microbe-Induced Tolerance in Plants. Front. Plant Sci. 2020, 11, 1750. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N.; Rivero, R.M.; Shulaev, V.; Blumwald, E.; Mittler, R. Abiotic and biotic stress combinations. New Phytol. 2014, 203, 32–43. [Google Scholar] [CrossRef]
- Zago, E.; Morsa, S.; Dat, J.F.; Alard, P.; Ferrarini, A.; Inzé, D.; Delledonne, M.; Van Breusegem, F. Nitric oxide- and hydrogen peroxide-responsive gene regulation during cell death induction in tobacco. Plant Physiol. 2006, 141, 404–411. [Google Scholar] [CrossRef] [Green Version]
- Asada, K. The water-water cycle in chloroplasts: Scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Biol. 1999, 50, 601–639. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reaction | Plasmid 1 | Plasmid 2 |
|---|---|---|
| Positive Control | pGBKT7-53 | pGADT7-T |
| Negative Control | pGBKT7-Lam | GADT7-T |
| BD (Target gene) | pGBKT7-GhCBL10 | |
| Empty vector | pGADT7 |
| Transcript ID | Length (aa) | MW (kDa) | Charge | PI | GRAVY | Instability Index | Subcellular Localization |
|---|---|---|---|---|---|---|---|
| Gh_A02G0578.1 | 393 | 43.061 | −4 | 5.941 | −0.308 | Stable | Cytoplasm |
| Gh_A04G0603.1 | 390 | 42.855 | 3.5 | 6.983 | −0.326 | Stable | Cytoplasm |
| Gh_A07G1193.1 | 256 | 28.12 | 7 | 8.974 | −0.326 | Stable | Cytoplasm |
| Gh_A08G1067.1 | 393 | 43.091 | −5 | 5.772 | −0.325 | Stable | Cytoplasm |
| Gh_A09G1368.1 | 390 | 42.61 | 4 | 7.118 | −0.299 | Stable | Cytoskeleton |
| Gh_A11G0966.1 | 393 | 43.026 | −6.5 | 5.579 | −0.36 | Stable | Cytoplasm |
| Gh_A11G2886.1 | 390 | 42.682 | 2 | 6.786 | −0.332 | Stable | Cytoplasm |
| Gh_A12G1098.1 | 393 | 43.071 | −6 | 5.594 | −0.335 | Stable | Cytoplasm |
| Gh_D02G0636.1 | 393 | 43.044 | −4.5 | 5.909 | −0.3 | Stable | Cytoplasm |
| Gh_D04G1064.1 | 390 | 42.812 | 3.5 | 6.983 | −0.306 | Stable | Cytoplasm |
| Gh_D07G1294.1 | 393 | 43.039 | −6 | 5.594 | −0.328 | Stable | Cytoplasm |
| Gh_D08G1348.1 | 393 | 43.04 | −5.5 | 5.618 | −0.338 | Stable | Cytoskeleton |
| Gh_D09G1369.1 | 390 | 42.695 | 4 | 7.118 | −0.294 | Stable | Cytoskeleton |
| Gh_D11G1117.1 | 393 | 43.062 | −6 | 5.596 | −0.358 | Stable | Cytoplasm |
| Gh_D11G3272.1 | 390 | 42.616 | 2 | 6.786 | −0.32 | Stable | Cytoskeleton |
| Gh_D12G1222.1 | 393 | 43.042 | −6 | 5.594 | −0.321 | Stable | Cytoplasm |
| Transcript ID | Name | Gene Description | Chr | Starting | Ending | Length |
|---|---|---|---|---|---|---|
| Gh_D06G1756.1 | PRA1B1 | PRA1 family protein B1 | D06 | 57,193,276 | 57,193,932 | 657 |
| Gh_A11G0688.1 | DSP8 | Putative dual-specificity protein DSP8 phosphatase | A11 | 6,717,943 | 6,719,957 | 945 |
| Gh_A07G1725.1 | CAB-151 | Chlorophyll a-b binding protein 151, chloroplastic | A07 | 70,403,379 | 70,404,266 | 798 |
| Gh_AO8G1067.1 | SAMS2 | S-adenosylmethionine synthase-2 | A08 | 73,601,857 | 73,603,038 | 1182 |
| Gh_D12G0158.1 | PYD3 | Beta-ureidopropionase | D12 | 2,003,668 | 2,006,254 | 1251 |
| Gh_D04G1908.1 | RPL34 | 60S ribosomal protein L34 | D04 | 51,393,192 | 51,394,076 | 363 |
| Gh_D02G0037.1 | UBC28 | Ubiquitin-conjugating enzyme E2 28 | D02 | 190,259 | 192,000 | 447 |
| Gh_D06G1538.1 | PSAF | Photosystem I reaction center subunit III, chloroplastic | D06 | 51,265,731 | 51,266,405 | 675 |
| Gh_D08G1752.1 | LON2 | Lon protease homolog 2, peroxisomal | D08 | 53,762,846 | 53,770,001 | 2670 |
| Gh_D02G0914.1 | PAH2 | Phosphatidate phosphatase PAH2 | D02 | 19,402,617 | 19,409,204 | 2934 |
| Gh_A11G2956.1 | BEE3 | Transcription factor BEE 3 | scaffold2723_A11 | 67,019 | 68,759 | 708 |
| Gh_D12G0965.1 | Rnf25 | E3 ubiquitin-protein ligase RNF25 | D12 | 35,117,734 | 35,120,355 | 1026 |
| Gh_A13G2030.1 | RAX2 | Transcription factor RAX2 | A13 | 79,732,246 | 79,733,388 | 903 |
| Gh_A12G2413.1 | ALMT9 | Aluminum-activated malate transporter 9 | A12 | 86,624,248 | 86,627,577 | 1839 |
| Gh_D11G0245.1 | ARF9 | Auxin response factor 9 | D11 | 2,017,754 | 2,033,373 | 3696 |
| Gh_D11G2402.1 | NA | NA | D11 | 47,820,689 | 47,823,871 | 1290 |
| Gh_D09G1701.1 | NA | NA | D09 | 44,755,344 | 44,757,734 | 2070 |
| Gh_A05G3519.1 | At1g54200 | Protein BIG GRAIN 1-like B | A05 | 90,846,177 | 90,847,466 | 1290 |
| Gh_D08G0705.1 | NA | Ent-copalyl diphosphate synthase, chloroplastic | D08 | 9,782,732 | 9,788,296 | 2538 |
| Gh_A04G1028.1 | At4g26680 | Pentatricopeptide repeat-containing, containing protein At4g26680 | A04 | 60,318,577 | 60,320,187 | 1611 |
| Gh_D05G3560.1 | RH32 | DEAD-box ATP-dependent RNA helicase 32 | D05 | 58,950,106 | 58,954,430 | 2262 |
| Gh_D13G0219.1 | AN11010 | Putative GTPase-activating protein | D13 | 2,155,213 | 2,163,523 | 2538 |
| Gh_A12G0039.1 | NA | NA | A12 | 598,420 | 600,787 | 1281 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kilwake, J.W.; Umer, M.J.; Wei, Y.; Mehari, T.G.; Magwanga, R.O.; Xu, Y.; Hou, Y.; Wang, Y.; Shiraku, M.L.; Kirungu, J.N.; et al. Genome-Wide Characterization of the SAMS Gene Family in Cotton Unveils the Putative Role of GhSAMS2 in Enhancing Abiotic Stress Tolerance. Agronomy 2023, 13, 612. https://doi.org/10.3390/agronomy13020612
Kilwake JW, Umer MJ, Wei Y, Mehari TG, Magwanga RO, Xu Y, Hou Y, Wang Y, Shiraku ML, Kirungu JN, et al. Genome-Wide Characterization of the SAMS Gene Family in Cotton Unveils the Putative Role of GhSAMS2 in Enhancing Abiotic Stress Tolerance. Agronomy. 2023; 13(2):612. https://doi.org/10.3390/agronomy13020612
Chicago/Turabian StyleKilwake, Joseph Wanjala, Muhammad Jawad Umer, Yangyang Wei, Teame Gereziher Mehari, Richard Odongo Magwanga, Yanchao Xu, Yuqing Hou, Yuhong Wang, Margaret Linyerera Shiraku, Joy Nyangasi Kirungu, and et al. 2023. "Genome-Wide Characterization of the SAMS Gene Family in Cotton Unveils the Putative Role of GhSAMS2 in Enhancing Abiotic Stress Tolerance" Agronomy 13, no. 2: 612. https://doi.org/10.3390/agronomy13020612