
The Relationship between Core Rhizosphere Taxa and Peanut Nodulation Capacity under Different Cover Crop Amendments
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Material Collection
2.2. Pot Experiment and Sampling
2.3. Determination of Soil and Plant Chemical Properties
2.4. DNA Extraction, Sequencing and Bioinformatics Analysis
2.5. Statistical Analysis
3. Results
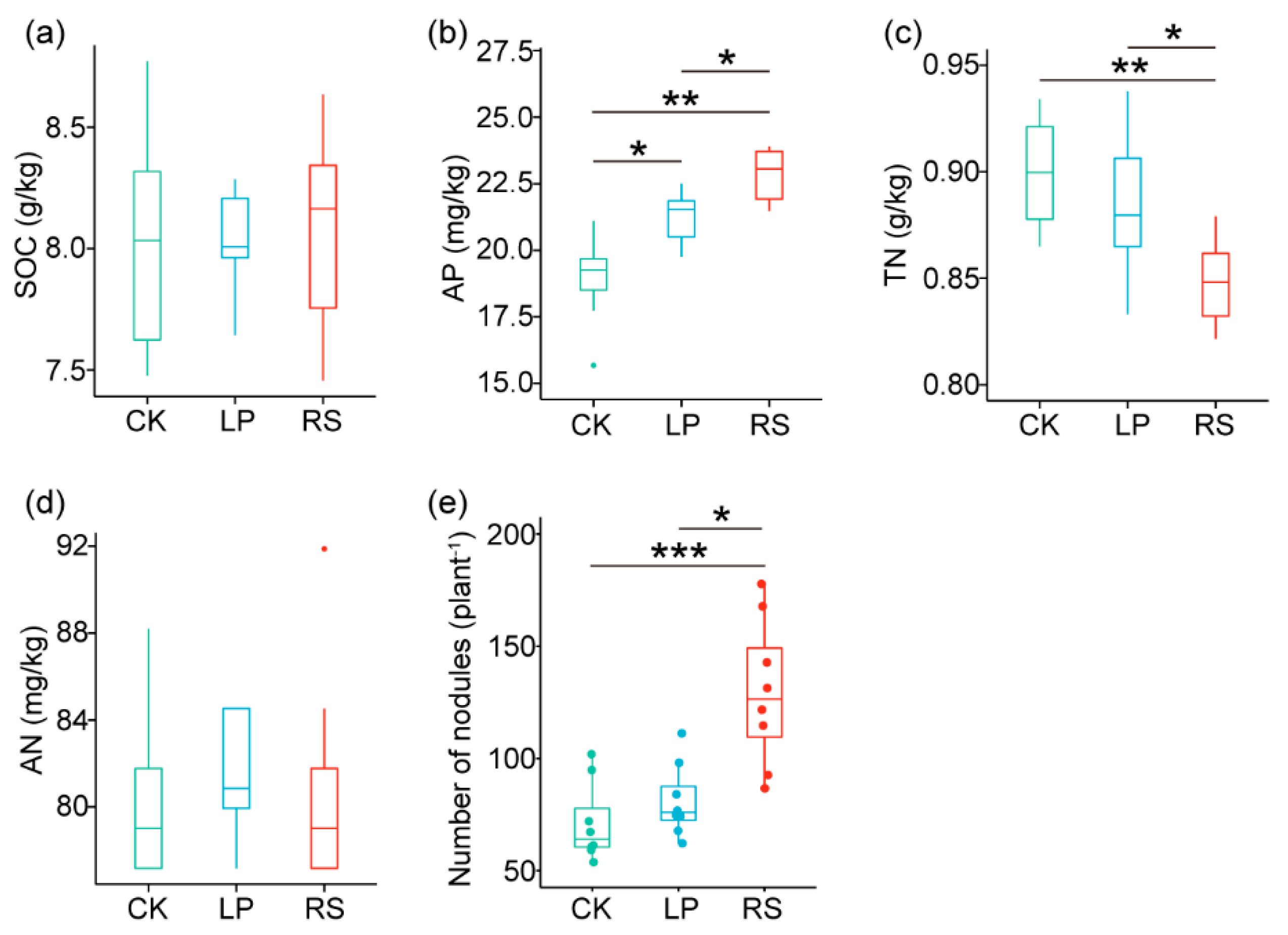
3.1. Cover Crop Nutrient Contents, Rhizosphere Soil Chemical Properties, and Peanut Nodulation Ability
3.2. Rhizosphere Soil Microbial Community Diversity and Structure
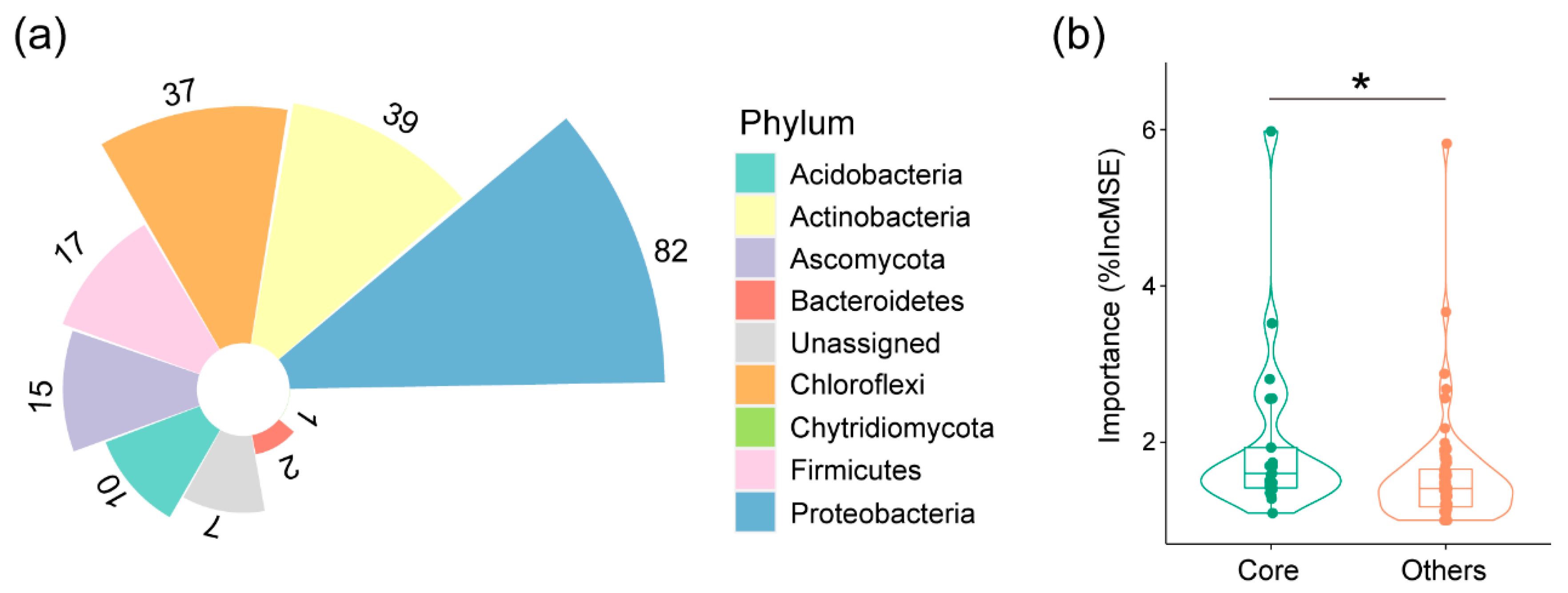
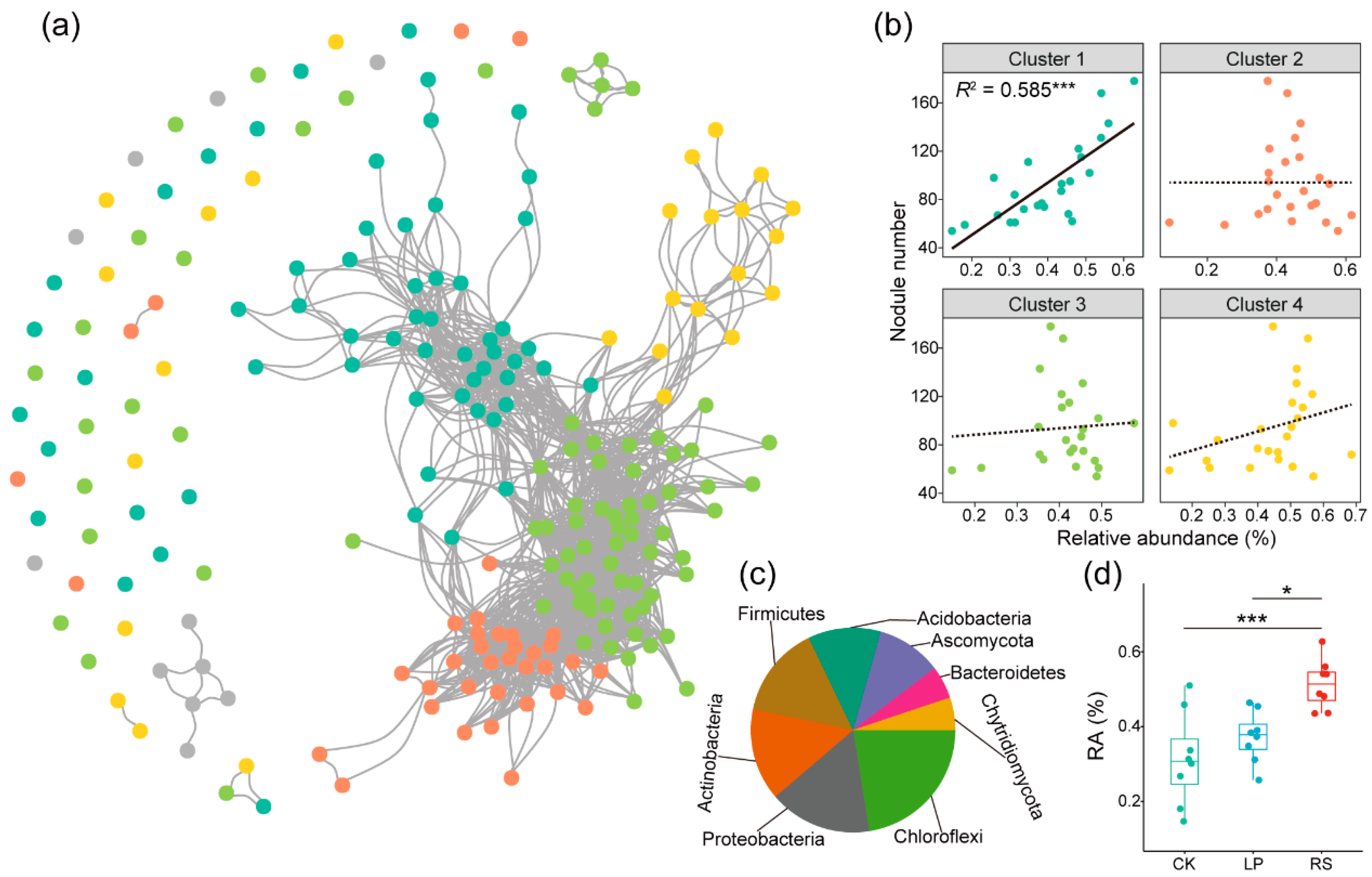
3.3. Correlation between Core Microbiota and Peanut Nodulation Ability
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tilman, D.; Cassman, K.G.; Matson, P.A.; Naylor, R.; Polasky, S. Agricultural sustainability and intensive production practices. Nature 2002, 418, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Godfray, H.C.; Beddington, J.R.; Crute, I.R.; Haddad, L.; Lawrence, D.; Muir, J.F.; Pretty, J.; Robinson, S.; Thomas, S.M.; Toulmin, C. The challenge of food security. Science 2010, 327, 812–818. [Google Scholar] [CrossRef] [Green Version]
- Bender, S.F.; Wagg, C.; van der Heijden, M.G. An underground revolution: Biodiversity and soil ecological engineering for agricultural sustainability. Trends Ecol. Evol. 2016, 31, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Li, P.; Wu, M.; Liu, K.; Evangelos, P.; Liu, J.; Liu, M.; Li, Z. Variation in rhizosphere microbial communities and its association with the nodulation ability of peanut. Arch. Agron. Soil Sci. 2022. [Google Scholar] [CrossRef]
- Suzaki, T.; Kawaguchi, M. Root nodulation: A developmental program involving cell fate conversion triggered by symbiotic bacterial infection. Curr. Opin. Plant Biol. 2014, 21, 16–22. [Google Scholar] [CrossRef]
- Dong, W.; Zhu, Y.; Chang, H.; Wang, C.; Yang, J.; Shi, J.; Gao, J.; Yang, W.; Lan, L.; Wang, Y.; et al. An SHR-SCR module specifies legume cortical cell fate to enable nodulation. Nature 2021, 589, 586–590. [Google Scholar] [CrossRef]
- Poole, P.; Ramachandran, V.; Terpolilli, J. Rhizobia: From saprophytes to endosymbionts. Nat. Rev. Microbiol. 2018, 16, 291–303. [Google Scholar] [CrossRef]
- Wang, X.; Feng, H.; Wang, Y.; Wang, M.; Xie, X.; Chang, H.; Wang, L.; Qu, J.; Sun, K.; He, W.; et al. Mycorrhizal symbiosis modulates the rhizosphere microbiota to promote rhizobia-legume symbiosis. Mol. Plant 2021, 14, 503–516. [Google Scholar] [CrossRef]
- Benezech, C.; Le Scornet, A.; Gourion, B. Medicago-Sinorhizobium-Ralstonia: A Model System to Investigate Pathogen-Triggered Inhibition of Nodulation. Mol. Plant Microbe Interact. 2021, 34, 499–503. [Google Scholar] [CrossRef]
- Schloss, P.D.; Handelsman, J. The last word: Books as a statistical metaphor for microbial communities. Annu. Rev. Microbiol. 2007, 61, 23–34. [Google Scholar] [CrossRef]
- Wertz, S.; Degrange, V.; Prosser, J.I.; Poly, F.; Commeaux, C.; Guillaumaud, N.; Roux, X.L. Decline of soil microbial diversity does not influence the resistance and resilience of key soil microbial functional groups following a model disturbance. Environ. Microbiol. 2007, 9, 2211–2219. [Google Scholar] [CrossRef]
- Griffiths, B.S.; Hallett, P.D.; Kuan, H.L.; Gregory, A.S.; Watts, C.W.; Whitmore, A.P. Functional resilience of soil microbial communities depends on both soil structure and microbial community composition. Biol. Fertil. Soils 2008, 44, 745–754. [Google Scholar] [CrossRef]
- Philippot, L.; Andersson, S.G.; Battin, T.J.; Prosser, J.I.; Schimel, J.P.; Whitman, W.B.; Hallin, S. The ecological coherence of high bacterial taxonomic ranks. Nat. Rev. Microbiol. 2010, 8, 523–529. [Google Scholar] [CrossRef]
- Peter, H.; Beier, S.; Bertilsson, S.; Lindström, E.S.; Langenheder, S.; Tranvik, L.J. Function-specific response to depletion of microbial diversity. ISME J. 2011, 5, 351–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shade, A.; Handelsman, J. Beyond the Venn diagram: The hunt for a core microbiome. Environ. Microbiol. 2012, 14, 4–12. [Google Scholar] [CrossRef]
- Gijsman, A.J.; Hoogenboom, G.; Parton, W.J.; Kerridge, P.C. Modifying DSSAT crop models for low-input agricultural systems using a soil organic matter-residue module from CENTURY. Agron. J. 2002, 94, 462–474. [Google Scholar] [CrossRef]
- Basu, M.; Bhadoria, P.B.; Mahapatra, S.C. Growth, nitrogen fixation, yield and kernel quality of peanut in response to lime, organic and inorganic fertilizer levels. Bioresour. Technol. 2008, 99, 4675–4683. [Google Scholar] [CrossRef]
- Nascente, A.S.; Li, Y.C.; Crusciol, C.A. Cover crops and no-till effects on physical fractions of soil organic matter. Soil Tillage Res. 2013, 130, 52–57. [Google Scholar] [CrossRef]
- Larkin, R.P. Soil health paradigms and implications for disease management. Annu. Rev. Phytopathol. 2015, 53, 199–221. [Google Scholar] [CrossRef]
- Kanders, M.J.; Berendonk, C.; Fritz, C.; Watson, C.; Wichern, F. Catch crops store more nitrogen below-ground when considering Rhizodeposits. Plant Soil 2017, 417, 287–299. [Google Scholar] [CrossRef]
- Wen, L.; Lee-Marzano, S.; Ortiz-Ribbing, L.M.; Gruver, J.; Hartman, G.L.; Eastburn, D.M. Suppression of soilborne diseases of soybean with cover crops. Plant Dis. 2017, 101, 1918–1928. [Google Scholar] [CrossRef] [Green Version]
- Xiang, X.; Zhang, J.; Li, G.; Leng, K.; Sun, L.; Qin, W.; Peng, C.; Xu, C.; Liu, J.; Jiang, Y. Positive feedback between peanut and arbuscular mycorrhizal fungi with the application of hairy vetch in Ultisol. Front. Microbiol. 2022, 13, 1002459. [Google Scholar] [CrossRef] [PubMed]
- Barel, J.M.; Kuyper, T.W.; de Boer, W.; Douma, J.C.; de Deyn, G.B. Legacy effects of diversity in space and time driven by winter cover crop biomass and nitrogen concentration. J. Appl. Ecol. 2018, 55, 299–310. [Google Scholar] [CrossRef] [Green Version]
- Manici, L.M.; Caputo, F.; Nicoletti, F.; Leteo, F.; Campanelli, G. The impact of legume and cereal cover crops on rhizosphere microbial communities of subsequent vegetable crops for contrasting crop decline. Biol. Control 2018, 120, 17–25. [Google Scholar] [CrossRef]
- Liu, X.; Hannula, S.E.; Li, X.; Hundscheid, M.P.; klein Gunnewiek, P.J.; Clocchiatti, A.; Ding, W.; de Boer, W. Decomposing cover crops modify root-associated microbiome composition and disease tolerance of cash crop seedlings. Soil Biol. Biochem. 2021, 160, 108343. [Google Scholar] [CrossRef]
- Li, P.; Liu, J.; Jiang, C.; Wu, M.; Liu, M.; Wei, S.; Qiu, C.; Li, G.; Xu, C.; Li, Z. Trade-off between potential phytopathogenic and non-phytopathogenic fungi in the peanut monoculture cultivation system. Appl. Soil Ecol. 2020, 148, 103508. [Google Scholar] [CrossRef]
- Liu, W.; Wang, Q.; Wang, B.; Wang, X.; Franks, A.E.; Teng, Y.; Li, Z.; Luo, Y. Changes in the abundance and structure of bacterial communities under long-term fertilization treatments in a peanut monocropping system. Plant Soil 2015, 395, 415–427. [Google Scholar] [CrossRef]
- Jani, A.D.; Mulvaney, M.J.; Erickson, J.E.; Leon, R.G.; Wood, C.W.; Rowland, D.L.; Enloe, H.A. Peanut nitrogen credits to winter wheat are negligible under conservation tillage management in the southeastern USA. Field Crops Res. 2020, 249, 107739. [Google Scholar] [CrossRef]
- Wang, H.; Ma, C.; Xu, F.; Lu, F.; Zhang, W.; Dai, C. Root endophyte-enhanced peanut-rhizobia interaction is associated with regulation of root exudates. Microbiol. Res. 2021, 250, 126765. [Google Scholar] [CrossRef]
- Fan, K.; Delgado-Baquerizo, M.; Guo, X.; Wang, D.; Wu, Y.; Zhu, M.; Yu, W.; Yao, Y.; Zhu, Y.; Chu, H. Suppressed N fixation and diazotrophs after four decades of fertilization. Microbiome 2019, 7, 143. [Google Scholar] [CrossRef]
- Pansu, M.; Gautheyrou, J. Handbook of Soil Analysis: Mineralogical, Organic and Inorganic Methods; Springer: Berlin, Germany, 2007; pp. 327–370. [Google Scholar] [CrossRef]
- Biddle, J.F.; Fitz-Gibbon, S.; Schuster, S.C.; Brenchley, J.E.; House, C.H. Metagenomic signatures of the Peru Margin subseafloor biosphere show a genetically distinct environment. Proc. Natl. Acad. Sci. USA 2008, 105, 10583–10588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Cui, W.; Feng, X.; Zhong, Z.; Li, Y.; Chen, W.; Chen, W.; Shao, X.; Tian, C. Rhizobia inhabiting nodules and rhizosphere soils of alfalfa: A strong selection of facultative microsymbionts. Soil Biol. Biochem. 2018, 116, 340–350. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Adékambi, T.; Shinnick, T.M.; Raoult, D.; Drancourt, M. Complete rpoB gene sequencing as a suitable supplement to DNA-DNA hybridization for bacterial species and genus delineation. Int. J. Syst. Evol. Microbiol. 2008, 58, 1807–1814. [Google Scholar] [CrossRef] [Green Version]
- Vos, M.; Quince, C.; Pijl, A.S.; de Hollander, M.; Kowalchuk, G.A. A comparison of rpoB and 16S rRNA as markers in pyrosequencing studies of bacterial diversity. PLoS ONE 2012, 7, e30600. [Google Scholar] [CrossRef]
- Fish, J.A.; Chai, B.; Wang, Q.; Sun, Y.; Brown, C.T.; Tiedje, J.M.; Cole, J.R. FunGene: The functional gene pipeline and repository. Front. Microbiol. 2013, 4, 291. [Google Scholar] [CrossRef] [Green Version]
- Vegan: Community Ecology Package, R Package Version 2.5-4; R Foundation: Vienna, Austria, 2022. Available online: https://cran.r-project.org/package=vegan(accessed on 16 June 2019).
- de Cáceres, M.; Legendre, P.; Moretti, M. Improving indicator species analysis by combining groups of sites. Oikos 2010, 119, 1674–1684. [Google Scholar] [CrossRef]
- Jiao, S.; Chen, W.; Wei, G. Core microbiota drive functional stability of soil microbiome in reforestation ecosystems. Glob. Change Biol. 2022, 28, 1038–1047. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Prasad, A.M.; Iverson, L.R.; Liaw, A. Newer classification and regression tree techniques: Bagging and random forests for ecological prediction. Ecosystems 2006, 9, 181–199. [Google Scholar] [CrossRef]
- rfPermute: Estimate Permutation p-Values for Random Forest Importance Metrics, R Package Version 2.5; R Foundation: Vienna, Austria, 2022. Available online: https://CRAN.R-project.org/package=rfPermute(accessed on 10 September 2021).
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Baquerizo, M.; Reith, F.; Dennis, P.G.; Hamonts, K.; Powell, J.R.; Young, A.; Singh, B.K.; Bissett, A. Ecological drivers of soil microbial diversity and soil biological networks in the Southern Hemisphere. Ecology 2018, 99, 583–596. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, M.; Hastings, A.; Cheng, K.; Yue, Q.; Chadwick, D.; Espenberg, M.; Truu, J.; Rees, R.M.; Smith, P. A critical review of the impacts of cover crops on nitrogen leaching, net greenhouse gas balance and crop productivity. Glob. Change Biol. 2019, 25, 2530–2543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, K.; Wu, Y. Rhizosphere calcareous soil P-extraction at the expense of organic carbon from root-exuded organic acids induced by phosphorus deficiency in several plant species. J. Soil Sci. Plant Nutr. 2014, 60, 640–650. [Google Scholar] [CrossRef]
- Han, Q.; Ma, Q.; Chen, Y.; Tian, B.; Xu, L.; Bai, Y.; Chen, W.; Li, X. Variation in rhizosphere microbial communities and its association with the symbiotic efficiency of rhizobia in soybean. ISME J. 2020, 14, 1915–1928. [Google Scholar] [CrossRef] [Green Version]
- Barberán, A.; Ramirez, K.S.; Leff, J.W.; Bradford, M.A.; Wall, D.H.; Fierer, N. Why are some microbes more ubiquitous than others? Predicting the habitat breadth of soil bacteria. Ecol. Lett. 2014, 17, 794–802. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Shi, W.; Kronzucker, H.J. How plant root exudates shape the nitrogen cycle. Trends Plant Sci. 2017, 22, 661–673. [Google Scholar] [CrossRef]
- Chen, Y.; Bonkowski, M.; Shen, Y.; Griffiths, B.S.; Jiang, Y.; Wang, X.; Sun, B. Root ethylene mediates rhizosphere microbial community reconstruction when chemically detecting cyanide produced by neighbouring plants. Microbiome 2020, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Nievas, F.; Bogino, P.; Nocelli, N.; Giordano, W. Genotypic analysis of isolated peanut-nodulating rhizobial strains reveals differences among populations obtained from soils with different cropping histories. Appl. Soil Ecol. 2012, 53, 74–82. [Google Scholar] [CrossRef]
- Lee, S.M.; Kong, H.G.; Song, G.C.; Ryu, C.M. Disruption of Firmicutes and Actinobacteria abundance in tomato rhizosphere causes the incidence of bacterial wilt disease. ISME J. 2021, 15, 330–347. [Google Scholar] [CrossRef]
- Jin, J.; Krohn, C.; Franks, A.E.; Wang, X.; Wood, J.L.; Petrovski, S.; McCaskill, M.; Batinovic, S.; Xie, X.; Tang, C. Elevated atmospheric CO2 alters the microbial community composition and metabolic potential to mineralize organic phosphorus in the rhizosphere of wheat. Microbiome 2022, 10, 12. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.; Delgado-Baquerizo, M.; Guo, X.; Wang, D.; Zhu, Y.; Chu, H. Biodiversity of key-stone phylotypes determines crop production in a 4-decade fertilization experiment. ISME J. 2021, 15, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Blundell, R.; Schmidt, J.E.; Igwe, A.; Cheung, A.L.; Vannette, R.L.; Gaudin, A.C.; Casteel, C.L. Organic management promotes natural pest control through altered plant resistance to insects. Nat. Plants 2020, 6, 483–491. [Google Scholar] [CrossRef]
- Gumiere, T.; Rousseau, A.N.; da Costa, D.P.; Cassetari, A.; Cotta, S.R.; Andreote, F.D.; Gumiere, S.J.; Pavinato, P.S. Phosphorus source driving the soil microbial interactions and improving sugarcane development. Sci. Rep. 2019, 9, 4400. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Liu, M.; Li, G.; Liu, K.; Liu, T.; Wu, M.; Saleem, M.; Li, Z. Phosphorus availability increases pathobiome abundance and invasion of rhizosphere microbial networks by Ralstonia. Environ. Microbiol. 2021, 23, 5992–6003. [Google Scholar] [CrossRef]
- Wagg, C.; Schlaeppi, K.; Banerjee, S.; Kuramae, E.E.; van der Heijden, M.G. Fungal-bacterial diversity and microbiome complexity predict ecosystem functioning. Nat. Commun. 2019, 10, 4841. [Google Scholar] [CrossRef] [Green Version]
- Delgado-Baquerizo, M.; Reich, P.B.; Trivedi, C.; Eldridge, D.J.; Abades, S.; Alfaro, F.D.; Bastida, F.; Berhe, A.A.; Cutler, N.A.; Gallardo, A.; et al. Multiple elements of soil biodiversity drive ecosystem functions across biomes. Nat. Ecol. Evol. 2020, 4, 210–220. [Google Scholar] [CrossRef]
- Alberty, R.A. Thermodynamics of the mechanism of the nitrogenase reaction. Biophys. Chem. 2005, 114, 115–120. [Google Scholar] [CrossRef]
- Zhang, J.; Zheng, M.; Zhang, Y.; Wang, J.; Shen, H.; Lin, Y.; Tang, X.; Hui, D.; Lambers, H.; Sardans, J.; et al. Soil phosphorus availability affects diazotroph communities during vegetation succession in lowland subtropical forests. Appl. Soil Ecol. 2021, 166, 104009. [Google Scholar] [CrossRef]
- Che, R.; Qin, J.; Tahmasbian, I.; Wang, F.; Zhou, S.; Xu, Z.; Cui, X. Litter amendment rather than phosphorus can dramatically change inorganic nitrogen pools in a degraded grassland soil by affecting nitrogen-cycling microbes. Soil Biol. Biochem. 2018, 120, 145–152. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Y.; Yan, X.; Li, J.; Jiao, N.; Hu, S. Biochar amendments increase the yield advantage of legume-based intercropping systems over monoculture. Agric. Ecosyst. Environ. 2017, 237, 16–23. [Google Scholar] [CrossRef]
- Rondon, M.A.; Lehmann, J.; Ramírez, J.; Hurtado, M. Biological nitrogen fixation by common beans (Phaseolus vulgaris L.) increases with bio-char additions. Biol. Fertil. Soils 2007, 43, 699–708. [Google Scholar] [CrossRef]
- Van Zwieten, L.; Rose, T.; Herridge, D.; Kimber, S.; Rust, J.; Cowie, A.; Morris, S. Enhanced biological N2 fixation and yield of faba bean (Vicia faba L.) in an acid soil following biochar addition: Dissection of causal mechanisms. Plant Soil 2015, 395, 7–20. [Google Scholar] [CrossRef] [Green Version]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant-microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leng, K.; Liu, J.; Li, G.; Wang, P.; Zhou, P.; Sun, M.; Liu, K.; Zhang, J.; Ma, T.; Xiang, X. The Relationship between Core Rhizosphere Taxa and Peanut Nodulation Capacity under Different Cover Crop Amendments. Agronomy 2023, 13, 311. https://doi.org/10.3390/agronomy13020311
Leng K, Liu J, Li G, Wang P, Zhou P, Sun M, Liu K, Zhang J, Ma T, Xiang X. The Relationship between Core Rhizosphere Taxa and Peanut Nodulation Capacity under Different Cover Crop Amendments. Agronomy. 2023; 13(2):311. https://doi.org/10.3390/agronomy13020311
Chicago/Turabian StyleLeng, Ke, Jia Liu, Guilong Li, Ping Wang, Panling Zhou, Mingzhu Sun, Kailou Liu, Jinyi Zhang, Tingting Ma, and Xingjia Xiang. 2023. "The Relationship between Core Rhizosphere Taxa and Peanut Nodulation Capacity under Different Cover Crop Amendments" Agronomy 13, no. 2: 311. https://doi.org/10.3390/agronomy13020311