
Analysis of Genetic Diversity, Population Structure and Association Mapping for Late Blight Resistance in Potato (Solanum tuberosum L.) Accessions Using SSR Markers
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Evaluation of Resistance to Late Blight
2.3. Genomic DNA Extraction
2.4. SSR Fingerprinting
2.5. Data Analysis
2.6. Association Mapping
3. Results
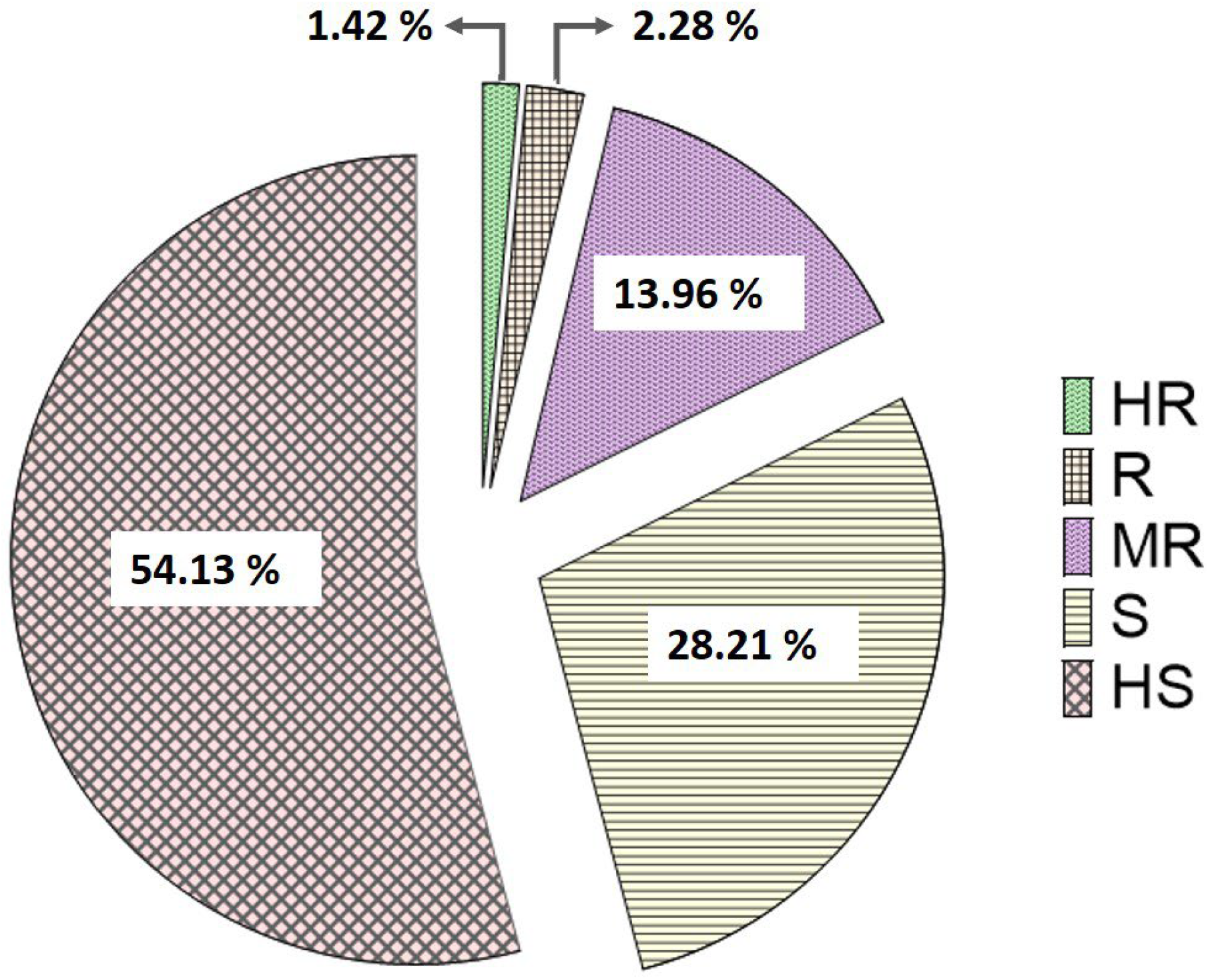
3.1. Assessment of Potato Late Blight
3.2. SSR Fingerprinting/Allelic Diversity
3.3. Population Structure
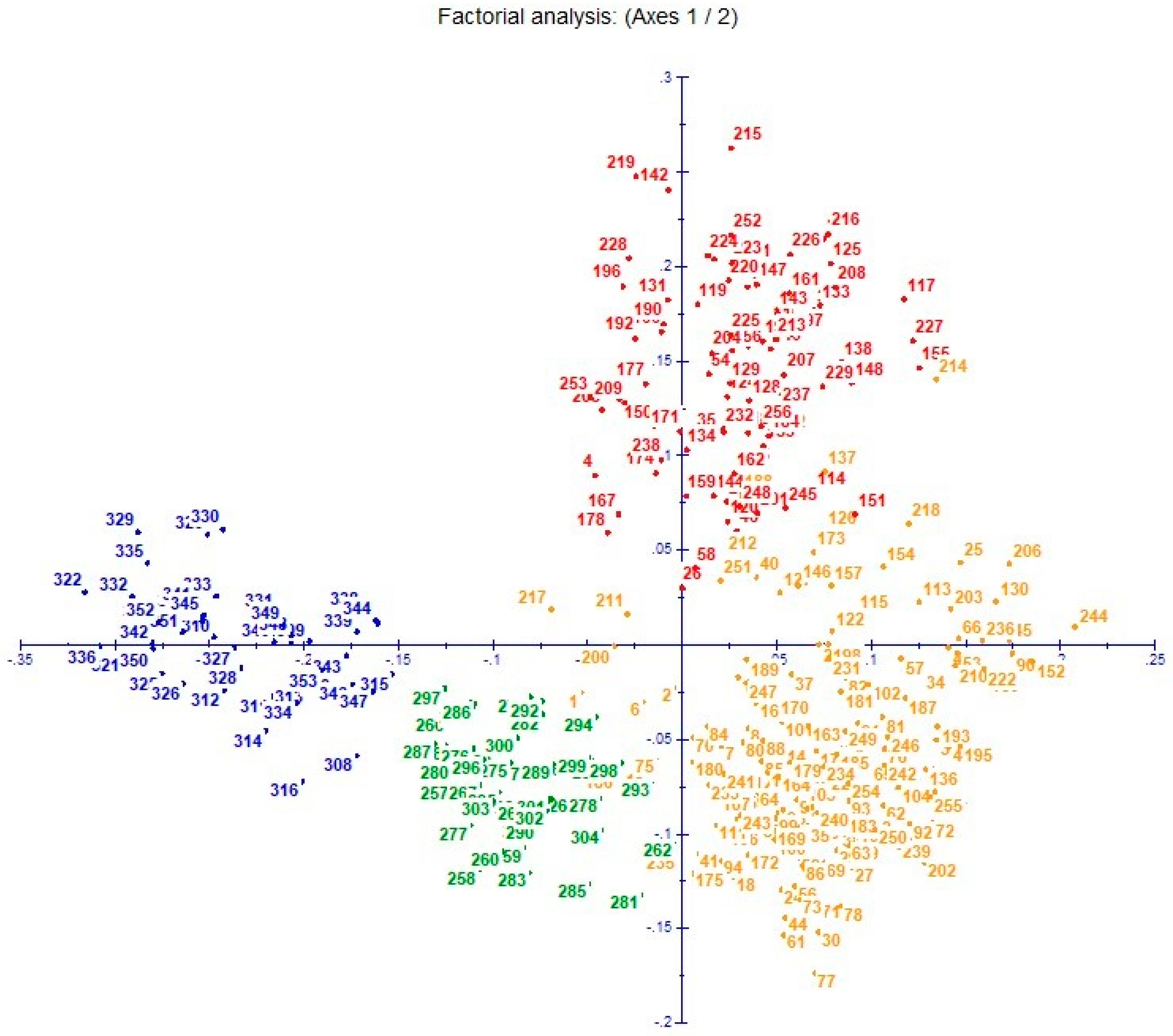
3.4. Cluster Analysis
3.5. Association Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Singh, B.; Goutam, U.; Kukreja, S.; Sharma, J.; Sood, S.; Bhardwaj, V. Potato biofortification: An effective way to fight global hidden hunger. Physiol. Mol. Biol. Plants 2021, 27, 2297–2313. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Kukreja, S.; Goutam, U. Impact of heat stress on potato (Solanum tuberosum L.): Present scenario and future opportunities. J. Hortic. Sci. Biotechnol. 2020, 95, 407–424. [Google Scholar] [CrossRef]
- Bradshaw, J.E. A brief history of the impact of potato genetics on the breeding of tetraploid potato cultivars for tuber propagation. Potato Res. 2022, 65, 461–501. [Google Scholar] [CrossRef]
- Ruiz de Arcaute, R.; Carrasco, A.; Ortega, F.; Rodriguez-Quijano, M.; Carrillo, J.M. Evaluation of genetic resources in a potato breeding program for chip quality. Agronomy 2022, 12, 1142. [Google Scholar] [CrossRef]
- Haverkort, A.J.; Boonekamp, P.M.; Hutten, R.; Jacobsen, E.; Lotz, L.A.P.; Kessel, G.J.T.; Vossen, J.H.; Visser, R.G.F. Durable late blight resistance in potato through dynamic varieties obtained by cisgenesis: Scientific and societal advances in the DuRPh Project. Potato Res. 2016, 59, 35–66. [Google Scholar] [CrossRef] [Green Version]
- Paluchowska, P.; Sliwka, J.; Yin, Z. Late blight resistance genes in potato breeding. Planta 2022, 255, 127. [Google Scholar] [CrossRef]
- Curran, D.; Froling, M. Large-scale mortality shocks and the Great Irish Famine 1845–1852. Econ. Model. 2010, 27, 1302–1314. [Google Scholar] [CrossRef]
- Álvarez, M.F.; Angarita, M.; Delgado, M.C.; García, C.; Jiménez-Gomez, J.; Gebhardt, C.; Mosquera, T. Identification of novel associations of candidate genes with resistance to late blight in Solanum tuberosum group Phureja. Front. Plant Sci. 2017, 8, 1040. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Zou, M.; Zhao, L.; Xia, Z.; Wang, J. Genome-Wide association mapping of late blight tolerance trait in potato (Solanum tuberosum L.). Front. Genet. 2021, 12, 1720. [Google Scholar] [CrossRef]
- van Eck, H.J. Genetics of morphological and tuber traits. In Potato Biology and Biotechnology; Elsevier Science BV: Amsterdam, The Netherlands, 2007; pp. 91–115. [Google Scholar] [CrossRef]
- Govindaraj, M.; Vetriventhan, M.; Srinivasan, M. Importance of genetic diversity assessment in crop plants and its recent advances: An overview of its analytical perspectives. Genet. Res. Int. 2015, 2015, 431487. [Google Scholar] [CrossRef] [Green Version]
- Moisan-Thiery, M.; Marhadour, S.; Kerlan, M.C.; Dessenne, N.; Perramant, M.; Gokelaere, T.; Le Hingrat, Y. Potato cultivar identification using simple sequence repeats markers (SSR). Potato Res. 2005, 48, 191–200. [Google Scholar] [CrossRef]
- Duan, S.; Liu, B.; Zhang, Y.; Li, G.; Guo, X. Genome-wide identification and abiotic stress-responsive pattern of heat shock transcription factor family in Triticum aestivum L. BMC Genom. 2019, 20, 257. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, J.K.; Ali, N.; Devi, S.; Kumar, V.; Zinta, R.; Chakrabarti, S.K. Development of microsatellite markers set for identification of indian potato varieties. Sci. Hortic. 2018, 231, 22–30. [Google Scholar] [CrossRef]
- Salimi, H.; Bahar, M.; Mirloh, A.; Talebi, M. Assessment of the genetic diversity among potato cultivars from different geographical areas using the genomic and EST microsatellites. Iran. J. Biotechnol. 2016, 14, 270. [Google Scholar] [CrossRef] [Green Version]
- Tillault, A.S.; Yevtushenko, D.P. Simple sequence repeat analysis of new potato varieties developed in Alberta, Canada. Plant Direct 2019, 3, e00140. [Google Scholar] [CrossRef] [Green Version]
- Ghislain, M.; Spooner, D.M.; Rodríguez, F.; Villamón, F.; Núñez, J.; Vásquez, C.; Waugh, R.; Bonierbale, M. Selection of highly informative and user-friendly microsatellites (SSRs) for genotyping of cultivated potato. Theor. Appl. Genet. 2004, 108, 881–890. [Google Scholar] [CrossRef]
- Ghislain, M.; Núñez, J.; Del Rosario Herrera, M.; Pignataro, J.; Guzman, F.; Bonierbale, M.; Spooner, D.M. Robust and highly informative microsatellite-based genetic identity kit for potato. Mol. Breed. 2009, 23, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Liao, H.; Guo, H. Using SSR to evaluate the genetic diversity of potato cultivars from Yunnan province (SW China). Acta Biol. Crac. Ser. Bot. 2014, 56, 16–27. [Google Scholar] [CrossRef]
- Song, X.; Zhang, C.; Li, Y.; Feng, S.; Yang, Q.; Huang, S. SSR analysis of genetic diversity among 192 diploid potato cultivars. Hortic. Plant J. 2016, 2, 163–171. [Google Scholar] [CrossRef]
- Bali, S.; Patel, G.; Novy, R.; Vining, K.; Brown, C.; Holm, D.; Porter, G.; Endelman, J.; Thompson, A.; Sathuvalli, V. Evaluation of genetic diversity among russet potato clones and varieties from breeding programs across the United States. PLoS ONE 2018, 13, e0201415. [Google Scholar] [CrossRef]
- Tiwari, J.K.; Ali, N.; Devi, S.; Zinta, R.; Kumar, V.; Chakrabarti, S.K. Analysis of allelic variation in wild potato (Solanum) species by simple sequence repeat (SSR) markers. 3 Biotech 2019, 9, 262. [Google Scholar] [CrossRef] [PubMed]
- Yuen, J.E.; Forbes, G.A. Estimating the level of susceptibility to Phytophthora infestans in potato genotypes. Phytopathology 2009, 99, 782–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, C.L.; Madden, L.V. Introduction to Plant Disease Epidemiology; John Wiley & Sons: New York, NY, USA, 1990. [Google Scholar]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dice, L.D. Measures of the amount of ecologic association between species. Ecology 1945, 26, 297–302. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.A.; vonHoldt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the evanno method. Conserv. Genet. Resour. 2011, 4, 359–361. [Google Scholar] [CrossRef]
- Forbes, G.; Pérez, W.; Piedra, J.A. Field Assessment of Resistance in Potato to Phytophthora Infestans: International Cooperators Guide; International Potato Center: Lima, Peru, 2014. [Google Scholar]
- Raker, C.M.; Spooner, D.M. Chilean tetraploid cultivated potato, Solanum tuberosum, is distinct from the Andean populations. Crop Sci. 2002, 42, 1451–1458. [Google Scholar] [CrossRef] [Green Version]
- Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogel, J.; Tingey, S.; Rafalski, A. The comparison of RFLP, RAPD, AFLP and SSR (Microsatellite) markers for germplasm analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- Gavrilenko, T.; Antonova, O.; Ovchinnikova, A.; Novikova, L.; Krylova, E.; Mironenko, N.; Pendinen, G.; Islamshina, A.; Shvachko, N.; Kiru, S.; et al. A microsatellite and morphological assessment of the russian national cultivated potato collection. Genet. Resour. Crop Evol. 2010, 57, 1151–1164. [Google Scholar] [CrossRef]
- Schönhals, E.M.; Ortega, F.; Barandalla, L.; Aragones, A.; Ruiz de Galarreta, J.I.; Liao, J.C.; Sanetomo, R.; Walkemeier, B.; Tacke, E.; Ritter, E.; et al. Identification and reproducibility of diagnostic dna markers for tuber starch and yield optimization in a novel association mapping population of potato (Solanum tuberosum L.). Theor. Appl. Genet. 2016, 129, 767–785. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, J.; Singh, B.; Gopal, J.; Poonam, P.; Patil, V.U. Molecular characterization of the Indian Andigena Potato core collection using microsatellite markers. Afr. J. Biotechnol. 2013, 12, 1025–1033. [Google Scholar]
- Deperi, S.I.; Tagliotti, M.E.; Bedogni, M.C.; Huarte, M.A. Association mapping to detect phytophthora infestans resistance in tetraploid potato. Crop Sci. 2022, 62, 216–234. [Google Scholar] [CrossRef]
- Spooner, D.M.; Núñez, J.; Trujillo, G.; Herrera, M.D.R.; Guzmán, F.; Ghislain, M. Extensive simple sequence repeat genotyping of potato landraces supports a major reevaluation of their gene pool structure and classification. Proc. Natl. Acad. Sci. USA 2007, 104, 19398–19403. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No | Primer | Repeat Motif | Ta (°C) | New Alleles | Range | PIC |
|---|---|---|---|---|---|---|
| 1. | STG0001 | (CT)n | 58 | 15 | 119–157 | 0.826 |
| 2. | STG0010 | (TG)n | 58 | 14 | 151–169 | 0.675 |
| 3. | STG0016 | (AGA)n | 55 | 15 | 118–198 | 0.877 |
| 4. | STG0025 | (AAAC)n | 56 | 15 | 187–205 | 0.899 |
| 5. | STI001 | (AAT)n | 60 | 11 | 170–198 | 0.739 |
| 6. | STI003 | (ACC)n | 60 | 20 | 125–222 | 0.881 |
| 7. | STI004 | (AAG)n | 60 | 16 | 70–102 | 0.852 |
| 8. | STI0012 | (ATT)n | 56 | 12 | 162–189 | 0.801 |
| 9. | STI0030 | (ATT)n | 58 | 15 | 83–131 | 0.907 |
| 10. | STI0032 | (GGA)n | 61 | 10 | 108–159 | 0.716 |
| 11. | STI0033 | (AGG)n | 61 | 12 | 105–139 | 0.829 |
| 12. | STM0019a,b | (AT)n (GT)n (AT)n (GT)n (GC)n (GT)n | 55 | 17 | 89–207 | 0.874 |
| 13. | STM0031 | (AC)n…(AC)n GCAC (AC)n (GCAC)n | 53 | 13 | 162–192 | 0.856 |
| 14. | STM0037 | (TC)n (AC)n AA (AC)n (AT)n | 52 | 14 | 66–178 | 0.724 |
| 15. | STM1052 | (AT)n GT (AT)n (GT)n | 55 | 15 | 207–264 | 0.879 |
| 16. | STM1053 | (TA)n (ATC)n | 53 | 8 | 162–190 | 0.786 |
| 17. | STM1064 | (TA)n (TG)n GT (TG)n | 52 | 12 | 163–210 | 0.838 |
| 18. | STM1104 | (TCT)n | 53 | 12 | 160–182 | 0.759 |
| 19. | STM1106 | (ATT)n | 51 | 14 | 152–192 | 0.932 |
| 20. | STM5114 | (ACC)n | 60 | 10 | 280–300 | 0.694 |
| 21. | STM5121 | (TGT)n | 50 | 10 | 278–291 | 0.804 |
| 22. | STM5127 | (TCT)n | 55 | 11 | 238–272 | 0.855 |
| 23. | STPoAc58 | (TA)n | 57 | 12 | 226–250 | 0.861 |
| 24. | STU6SNRN | (TGG)5 | 55 | 15 | 129–204 | 0.763 |
| 25. | STIKA | (T)12(A)9ATTCTTGTT(TA)2CA(TA)7 | 55 | 25 | 175–247 | 0.816 |
| Marker | Allele Size (bp) with Their Absolute Frequencies (In Brackets) | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| STG0001 | 119 (149) | 122 (134) | 125 (194) | 126 (237) | 129 (122) | 131 (170) | 132 (154) | 134 (73) | 135 (132) | 137 (106) | 138 (108) | 139 (28) | 140 (136) | 142 (157) | 157 (190) | |||||
| STG0010 | 151 (57) | 153 (114) | 154 (240) | 156 (180) | 158 (149) | 159 (185) | 162 (254) | 163 (280) | 164 (313) | 165 (242) | 166 (259) | 167 (194) | 168 (51) | 169 (25) | ||||||
| STG0016 | 118 (210) | 122 (142) | 126 (52) | 128 (154) | 131 (192) | 135 (226) | 138 (20) | 140 (28) | 148 (84) | 154 (182) | 156 (39) | 168 (26) | 180 (22) | 188 (26) | 198 (92) | |||||
| STG0025 | 187 (36) | 189 (97) | 190 (154) | 193 (82) | 194 (84) | 196 (66) | 197 (230) | 198 (204) | 199 (31) | 200 (56) | 201 (137) | 202 (136) | 203 (16) | 204 (22) | 205 (7) | |||||
| STI001 | 170 (160) | 173 (103) | 176 (133) | 177 (159) | 180 (309) | 183 (155) | 185 (286) | 188 (159) | 191 (221) | 194 (72) | 198 (22) | |||||||||
| STI003 | 125 (28) | 130 (146) | 135 (45) | 140 (225) | 147 (22) | 150 (213) | 155 (137) | 158 (59) | 162 (214) | 165 (62) | 171 (229) | 175 (52) | 180 (148) | 183 (66) | 192 (96) | 201 (105) | 205 (26) | 207 (25) | 215 (40) | 222 (32) |
| STI004 | 70 (121) | 72 (178) | 74 (216) | 75 (252) | 78 (133) | 80 (57) | 83 (27) | 85 (153) | 87 (148) | 89 (55) | 91 (42) | 93 (165) | 96 (73) | 98 (134) | 99 (133) | 102 (31) | ||||
| STI0012 | 162 (121) | 165 (238) | 167 (221) | 170 (263) | 173 (92) | 176 (18) | 178 (53) | 180 (114) | 183 (251) | 185 (42) | 186 (110) | 189 (90) | ||||||||
| STI0030 | 83 (203) | 85 (128) | 87 (106) | 89 (124) | 91 (37) | 96 (244) | 98 (53) | 102 (48) | 104 (82) | 105 (106) | 109 (8) | 110 (76) | 113 (12) | 128 (32) | 131 (21) | |||||
| STI0032 | 108 (190) | 111 (116) | 114 (115) | 116 (206) | 117 (107) | 119 (224) | 122 (173) | 123 (217) | 126 (70) | 159 (328) | ||||||||||
| STI0033 | 105 (159) | 108 (95) | 110 (302) | 113 (73) | 116 (118) | 119 (43) | 122 (126) | 125 (149) | 128 (227) | 131 (142) | 135 (23) | 139 (19) | ||||||||
| STM0019a,b | 89 (156) | 91 (242) | 98 (92) | 100 (102) | 103 (16) | 110 (138) | 112 (153) | 119 (20) | 153 (20) | 163 (44) | 165 (19) | 171 (17) | 180 (15) | 190 (155) | 197 (100) | 204 (201) | 207 (214) | |||
| STM0031 | 162 (159) | 165 (90) | 167 (168) | 168 (155) | 170 (22) | 172 (28) | 177 (63) | 179 (73) | 183 (126) | 185 (207) | 186 (219) | 187 (178) | 192 (28) | |||||||
| STM0037 | 66 (203) | 68 (303) | 70 (214) | 72 (110) | 74 (232) | 76 (246) | 79 (44) | 81 (97) | 84 (139) | 86 (30) | 90 (282) | 110 (180) | 156 (65) | 178 (174) | ||||||
| STM1052 | 207 (171) | 208 (253) | 210 (81) | 217 (108) | 221 (26) | 224 (164) | 226 (188) | 227 (81) | 242 (9) | 249 (106) | 251 (159) | 253 (85) | 255 (26) | 258 (30) | 262 (28) | |||||
| STM1053 | 162 (64) | 166 (82) | 168 (280) | 169 (59) | 171 (309) | 172 (135) | 175 (37) | 190 (81) | ||||||||||||
| STM1064 | 163 (80) | 178 (28) | 180 (119) | 183 (32) | 187 (322) | 189 (264) | 191 (154) | 193 (107) | 195 (89) | 197 (28) | 201 (27) | 210 (50) | ||||||||
| STM1104 | 160 (119) | 164 (148) | 165 (257) | 168 (330) | 169 (227) | 171 (63) | 172 (178) | 173 (109) | 175 (205) | 177 (90) | 178 (36) | 182 (14) | ||||||||
| STM1106 | 152 (171) | 157 (166) | 160 (188) | 164 (68) | 165 (49) | 168 (87) | 169 (64) | 171 (24) | 173 (51) | 175 (36) | 178 (20) | 184 (12) | 189 (30) | 192 (51) | ||||||
| STM5114 | 280 (145) | 283 (178) | 285 (274) | 288 (322) | 290 (172) | 291 (71) | 294 (196) | 296 (269) | 297 (59) | 300 (43) | ||||||||||
| STM5121 | 278 (62) | 282 (218) | 283 (144) | 284 (60) | 285 (289) | 286 (148) | 287 (88) | 288 (216) | 289 (70) | 291 (74) | ||||||||||
| STM5127 | 238 (255) | 239 (93) | 241 (163) | 242 (173) | 247 (92) | 249 (71) | 251 (121) | 254 (37) | 268 (114) | 270 (121) | 272 (108) | |||||||||
| STPoAc58 | 226 (134) | 230 (201) | 231 (185) | 232 (212) | 233 (183) | 235 (109) | 238 (39) | 241 (52) | 243 (26) | 245 (75) | 248 (111) | 250 (59) | ||||||||
| STU6SNRN | 129 (25) | 134 (19) | 138 (18) | 145 (188) | 179 (249) | 181 (237) | 182 (145) | 189 (207) | 190 (171) | 197 (234) | 198 (217) | 199 (155) | 200 (202) | 201 (172) | 204 (13) | |||||
| STIKA | 175 (217) | 184 (40) | 186 (77) | 189 (133) | 190 (144) | 192 (103) | 193 (176) | 194 (273) | 195 (201) | 196 (99) | 198 (145) | 199 (156) | 203 (33) | 211 (14) | 214 (248) | 219 (181) | 222 (282) | 223 (94) | 225 (131) | 230 (182) |
| 234 (39) | 235 (52) | 242 (152) | 245 (84) | 247 (25) | ||||||||||||||||
| Pop-1 | Pop-2 | Pop-3 | Pop-4 | |
|---|---|---|---|---|
| Pop-1 | - | 0.0714 | 0.0978 | 0.0355 |
| Pop-2 | 0.0714 | - | 0.0994 | 0.0634 |
| Pop-3 | 0.0978 | 0.0994 | - | 0.0980 |
| Pop-4 | 0.0355 | 0.0634 | 0.0980 | - |
| Model | Marker | Chromosome Number | Marker F-Value | Marker p-Value | Marker R2 |
|---|---|---|---|---|---|
| GLM | STM5127_254 | 1 | 7.9058 | 0.0053 | 0.0254 |
| GLM | STG0016_188 | 1 | 7.0407 | 0.0084 | 0.0227 |
| GLM | STI0032_111 | 5 | 7.1891 | 0.0077 | 0.0232 |
| GLM | STM0019a,b_103 | 6 | 9.6300 | 0.0021 | 0.0308 |
| GLM | STM0031_192 | 7 | 9.2656 | 0.0025 | 0.0297 |
| GLM | STM5121_278 | 12 | 10.5743 | 0.0013 | 0.0337 |
| MLM | STM5127_254 | 1 | 7.8669 | 0.0054 | 0.0259 |
| MLM | STM0019a,b_103 | 6 | 9.4861 | 0.0023 | 0.0312 |
| MLM | STM5121_278 | 12 | 8.6441 | 0.0035 | 0.0285 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhardwaj, V.; Kumar, A.; Sharma, S.; Singh, B.; Poonam; Sood, S.; Dipta, B.; Singh, R.; Siddappa, S.; Thakur, A.K.; et al. Analysis of Genetic Diversity, Population Structure and Association Mapping for Late Blight Resistance in Potato (Solanum tuberosum L.) Accessions Using SSR Markers. Agronomy 2023, 13, 294. https://doi.org/10.3390/agronomy13020294
Bhardwaj V, Kumar A, Sharma S, Singh B, Poonam, Sood S, Dipta B, Singh R, Siddappa S, Thakur AK, et al. Analysis of Genetic Diversity, Population Structure and Association Mapping for Late Blight Resistance in Potato (Solanum tuberosum L.) Accessions Using SSR Markers. Agronomy. 2023; 13(2):294. https://doi.org/10.3390/agronomy13020294
Chicago/Turabian StyleBhardwaj, Vinay, Ashwani Kumar, Sanjeev Sharma, Baljeet Singh, Poonam, Salej Sood, Bhawna Dipta, Rajender Singh, Sundaresha Siddappa, Ajay Kumar Thakur, and et al. 2023. "Analysis of Genetic Diversity, Population Structure and Association Mapping for Late Blight Resistance in Potato (Solanum tuberosum L.) Accessions Using SSR Markers" Agronomy 13, no. 2: 294. https://doi.org/10.3390/agronomy13020294