1. Introduction
The prolonged and excessive use of synthetic herbicides has often caused the emergence of environmental problems, such as the appearance of weed resistant biotypes [
1], or the accumulation in soils and ground water with adverse effects in living organisms and human health [
2,
3]. Herbicides based on natural products, especially plant secondary metabolites, are increasing in importance [
4], and the study of natural products produced by rare plants could offer the chance to identify new herbicidal compounds. In this regard, essential oils (EOs) and their components, generally having low persistence in the field as well as low incidence of resistance in weeds [
5], could be particularly attractive. EOs are natural complexes of semi-volatile and volatile aromatic compounds [
6]. They are produced by plants to attract pollinators and defend against predator attacks, and their composition can vary within the same species according to environmental conditions and harvest site characteristics [
7]. Those compounds are present in small amounts in different plant organs (leaves, stems, flowers, roots, or fruits) and their use has expanded in the last decades mainly for perfume, cosmetics, and food industries [
8]. They have also already been studied for their phytotoxic activities [
9,
10,
11]. In the European market, no commercial herbicides are available based on EOs, although European patent EP2684457A1 has been registered, based on oregano essential oils [
12].
One of the main botanical family producing EOs is Lamiaceae (formerly Labiatae), including 7886 species, thus being the largest family of the Lamiales order (
http://www.theplantlist.org/, accessed on 1 December 2022). Most of the species belonging to this family are aromatic. Lamiaceae species are largely present in the whole Mediterranean area, including Italy. EOs extracted from Lamiaceae have many biological applications and activities such as antioxidant, anti-inflammatory, antimicrobial, fungicidal, insecticidal [
7], and proved to be effective in inhibiting seed germination [
13,
14,
15].
Many Lamiaceae species were never or little considered for the herbicidal activity of their EOs, and neither the origin of the plants has been correlated with the biological activities. Hence, we tested the EOs extracted from some Lamiaceae species from the Apulia region (South Italy) having scarce or not exhaustive scientific literature.
EOs’ phytotoxic effects were often related to the inhibition of seed germination and to the decrease of the chlorophyll content [
9,
11].
In this study, we aimed to find EOs with good herbicidal activity to be employed for weed management in organic farming systems. As the main phytotoxic effects occur mostly by leaf damages, germination, and radicle inhibition, we tested, in in vitro conditions, the herbicidal activity of 13 EOs extracted from plants of six Lamiaceae species, evaluating the effects against cress and branched broomrape seed germination, tomato radicle growth, lambsquarters leaf disk phytotoxicity symptoms, and chlorophyll content. We also evaluated if the oils extracted from plants of the same species but originating from different sites were distinguishable by their biological activity and chemical composition.
4. Discussion
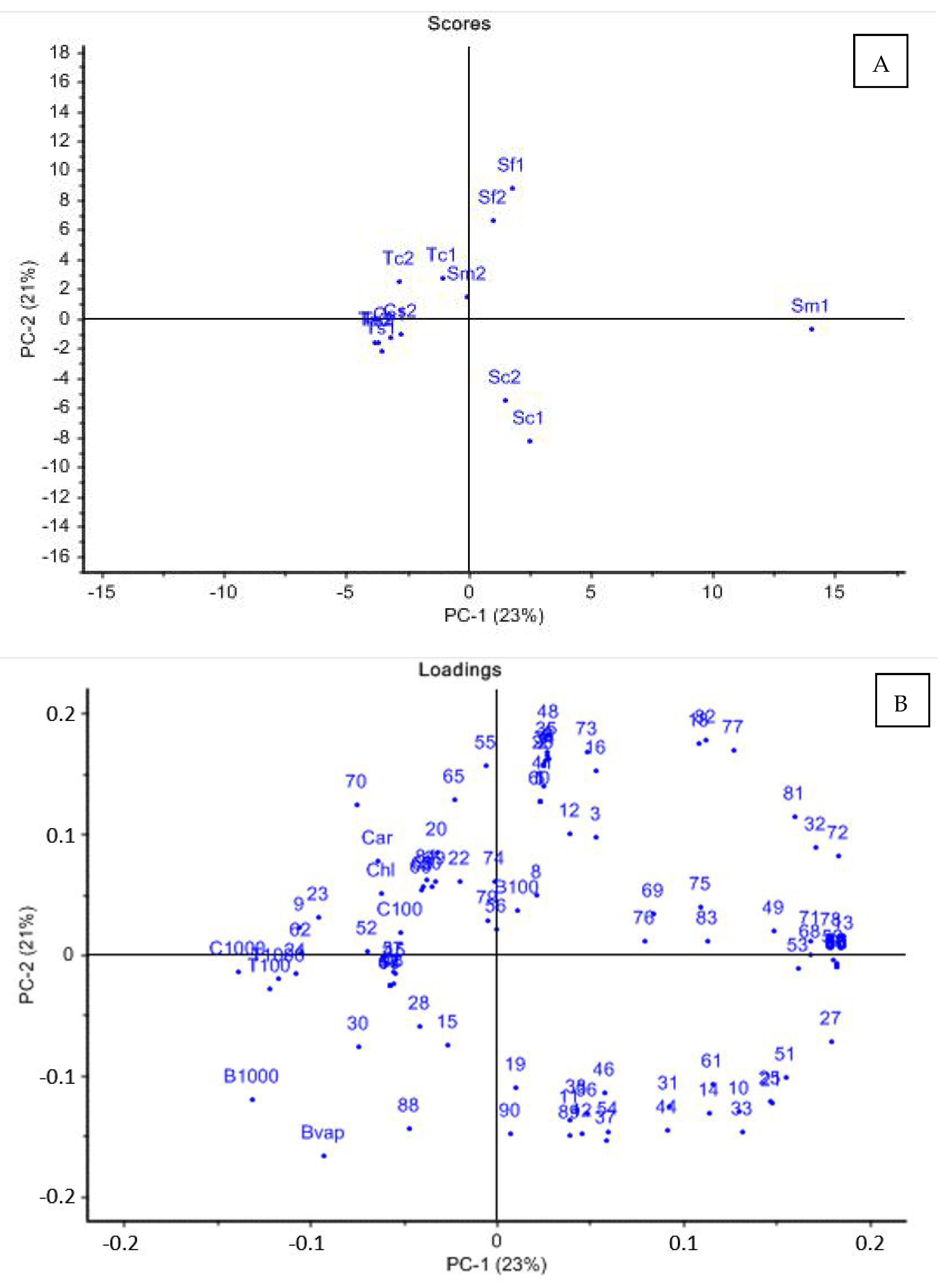
The results of our study in vitro confirm the potential biological activity of essential oils from aromatic Lamiaceae wild plants of Italian origin.
We tested 13 essential oils extracted each from plants of six Lamiacee species (
Table 1): in particular, two oils extracted from
Clinopodium suaveolens (Sm.) Kuntze,
Salvia fruticosa Mill. subsp.
thomasii (Lacaita) Brullo, Guglielmo, Pavone & Terrasi,
Satureja cuneifolia Ten.,
Satureja montana L. subsp.
montana,
Thymbra capitata (L.) Cav., and three oils from
Thymus spinulosus Ten. Each oil is characterized by two or three major phytochemical constituents, all monoterpenes, and many others present in low amount (
Table 2). As overall observation, the tested essential oils have herbicidal activity against germination and early radicle growth of two common test species (
L. sativum subsp.
sativum and
S. lycopersicum) and one parasitic species (
P. ramosa).
Within the same species, oils from
T. spinulosus,
C. suaveolens, and
S. cuneifolia gave consistent results among the different experiments and showed the same strong herbicidal activities. Oils from
S. fruticosa subsp.
thomasii gave consistent results too but proved to be the less active tested ones. Indeed, all oils from
T. spinulosus (Ts1, Ts2, Ts3) at 1000 ppm totally inhibited the initial growth of tomato radicles and the germination of cress and broomrape seeds. Even vapors from the three oils totally inhibited broomrape seed germination (
Table 6). In addition, Ts3 had a light activity on tomato radicle at the lowest tested dose of 100 ppm (
Table 4). Both essential oils from
C. suaveolens (Cs1, Cs2) at 1000 ppm showed a strong herbicidal activity (total or near total inhibition of cress and broomrape germination, by direct contact and vapors) with the only exception of the effect on the tomato radicle elongation (total inhibition from Cs1 and no activity from Cs2). Essential oils from
S. cuneifolia (Sc1, Sc2), both in direct contact and as vapors, caused great injury to broomrape seeds hampering germination, while they did not show any activity in the other tests. At the tested concentrations, both
S. fruticosa subsp.
thomasii oils (Sf1 and Sf2) proved to be the less effective oils, having a very light inhibitory effect on broomrape, but no effect in the other experiments.
Within the same species, oils from
T. capitata (Tc1 and Tc2) and
S. montana subsp.
montana (Sm1 and Sm2) gave inconsistent results and showed different herbicidal activities. In particular, Tc2 was the most powerful oil of the 13 tested, as it was strongly active in all experiments at 1000 ppm, totally injuring cress and broomrape seeds (both by direct contact and by vapors), completely inhibiting the growth of tomato radicles, and additionally, it was the only oil out of 13 active at the lowest tested concentration, causing a nearly complete inhibition of cress seed germination at 100 ppm (
Table 3) and reducing the chlorophyll and carotenoids content in lambsquarters leaf disks (
Table 5). In contrast, Tc1 proved to be among the less active tested oils, being only partially active against broomrape seed germination at 1000 ppm. Similar observations can be shared for
S. montana subsp.
montana EOs, where Sm2 was strongly active on tomato radicles, cress and broomrape seed germination (both by direct contact and by vapors), while on the other hand Sm1 was nearly inactive in all experiments.
The good or scarce activity of the tested oils could be related to the presence or absence of those compounds which were found to be positively correlated to the biological activities (
Table 7). Indeed, compounds 15, 23, 24, 28, 30, 57, 62, 65, 66, and 84 were contained in the most active oils (Cs1, Cs2, Tc2, Sm2, Ts1, Ts2, and Ts3) and in Tc1 (less active), while they were absent or scarcely present in the less active oils (Sc1, Sc2, Sm1, Sf1, and Sf2) (
Table 2).
In the case of
T. spinulosus, the explanation of the consistent results among the three extracted oils could be that the extracted oils were stable in composition, regardless of environmental, climatic, and vegetational differences of the harvest sites [
16]. For three
T. spinulosus harvest sites having the same macroclimate features, differences concerning bioclimatic, geological, pedological characteristics, and plant communities were found. EOs Ts1, Ts2, and Ts3 composition consists of a total of 27 compounds for all three sites (
Table 2). The environmental differences very lightly influenced the chemical composition of EOs. Indeed, in the previous research, low differences were observed on the abundance and patterns among the three sites, since they share 25 compounds; only one compound (hotrienol) was exclusive to Ts1, and one to Ts2 (caryophyllene-oxide). The phytochemicals having the highest abundance in all of the 3 samples were thymol (62), p-cymene (24) and β-
cis-ocimene (28). Few differences in percentages were observed among the three oils for thymol (42.9, 48.8, and 45.9%, respectively), and β-ocimene (15.4, 11.7, and 10.1%, respectively). All the other compounds had very slight quantitative differences. The three most abundant constituents (24, 28, 62) were correlated to the biological activities C1000, T100, T1000, B1000 (
Table 7) and that could explain the good phytotoxic effects of those oils.
The EOs Cs1 and Cs2 extracted from
C. suaveolens had the very same composition, but there were some differences in terms of composition percentage between them. In particular, pulegone (58), δ-terpineol (45), and isopulegone-trans were found to be the most abundant constituents in both oils (
Table 2), the first being more abundant in Cs1 and the latter more abundant in Cs2 [
17,
20]. None of the phytochemicals correlated to the biological activities were found as abundant in the two oils, while β-pinene (15), α-terpinene (23), p-cymene (24), and β-
cis-ocimene (28), as a whole positively correlated with C1000, T100, T1000, and B1000 (
Table 7), were present in small amount with negligible quantitative differences, and that could explain the good phytotoxic effects of those oils.
Also for
S. cuneifolia, the explanation of the consistent results between Sc1 and Sc2 could be that the extracted oils were stable in composition, regardless of environmental, climatic, and vegetational differences of the harvest sites [
16]. For
S. cuneifolia at both investigated sites, with the same Mediterranean macroclimate, there were environmental, bioclimatic, geological, lithological and ecological, pedological, and vegetational differences. Even so, those different environmental conditions very little affected the phytochemical properties of the two oils. In total, for both oils, 36 compounds were identified with very few quantitative differences (
Table 2). In particular, the most abundant compounds in Sc1 and Sc2 were α-pinene (10) and α-terpineol (51), the latter showing a small abundance difference (11 and 17.1%, respectively). All the other minor compounds had small to irrelevant abundance differences. None of the phytochemicals positively correlated to the biological activities were found as abundant in the two oils, and that could explain the limited phytotoxic effects of those oils. Moreover, p-cymene (24), which was found to be positively correlated with B1000 (
Table 7), was present in different small amount in Sc1 and Sc2 (1.55 and 0.63%, respectively), and that could explain the broomrape seeds injury occurred (higher in Sc1 than in Sc2).
Despite the aligned (not promising) results obtained from
S. fruticosa subsp.
thomasii extracted oils, Sf1 and Sf2 were different in composition due to environmental differences of the harvest sites, being Sf1 collected from a less disturbed site than Sf2. That was reflected in a higher number of compounds (48) identified in Sf1 than in Sf2 (42 compounds), with six exclusive to Sf1, and 42 common (
Table 2). The most abundant components of
S. fruticosa subsp.
thomasii oils were eucalyptol (26) (40.2 to 60.9%), camphor (41) (1.9 to 14.9%), and α-pinene (10) (3.6 to 5%). Those data confirm the ones available in literature on the genus
Salvia, which show eucalyptol to be usually the most abundant compound [
21,
22]. Like for
S. cuneifolia, none of the phytochemicals positively correlated to the biological activities were found as abundant in the two oils, and that could explain the limited phytotoxic effects of those oils. P-cymene (24), which was found to be positively correlated with B1000 (
Table 7), was present in a small amount, and that could explain the broomrape seeds injury occurred.
T. capitata oils Tc1 and Tc2 shared 50 compounds, and Tc1 had 51 (
Table 2). The most abundant compounds were carvacrol (63), thymol (62), ϒ-terpinene (29), and p-cymene (24), the last three being more abundant in Tc2 than in Tc1 (
Table 2). In
S. montana subsp.
montana, the phytochemicals with the highest abundance in both samples were α-pinene (10), thymol (62), and ϒ-terpinene (29), the last two being much more abundant in Sm2 than in Sm1. Sm1 and Sm2 shared 55 compounds, whereas 3 were exclusive of Sm1 and 3 of Sm2. Thus, for
T. capitata and
S. montana subsp.
montana, the different biological activities among oils of the same species could be attributed to the different chemical composition of the extracted oils due to environmental and vegetational differences between the harvest sites [
17]. The variability among provenances is also confirmed by Angelini et al. who tested EOs from
S. montana subsp.
montana and their main compounds on different weeds and crops, finding that carvacrol was the most abundant (57%) and effective [
23]. Tc1 and Tc2 contained compounds 15, 23, 24, 28, 30, 62, 65, 66, and 84, positively correlated as a whole to all the tested activities (C100, C1000, T100, T1000, B1000, Chl, and Car) but Bvap (
Table 7). Most of those compounds were more abundant in Tc2 than in Tc1. That could explain the phytotoxicity of Tc2 in all experiments including C100, Chl, and Car, positively correlated to thymol acetate (65) and carvacrol acetate (66). Therefore, compounds 65 and 66 seems to be responsible for those activities. Compounds 23, 24, 57, 62, 65, and 84 (positively correlated as a whole to the tested activities C100, C1000, T100, T1000, B1000, C100, Chl, and Car) were also more abundant in Sm2 than in Sm1 and that could explain the good phytotoxic effect of Sm2.
The active compounds abovementioned are monoterpenes and monoterpenoids. Some of them had been studied individually or in combination and proved to be very active compounds. Vasilakoglou et al. found that carvacrol and thymol were very phytotoxic components (the most phytotoxic among 19 they tested), completely inhibiting rigid ryegrass germination and root length at 160 nL/cm
3 [
24]. Thymol was found to have high inhibitory effect against seeds of several weeds [
25]. That is congruent and could explain the high phytotoxicity of
T. spinulosus,
T. capitata, and
S. montana subsp.
montana oils, and even the different activity between oils extracted from the same species. Indeed, all tested
T. spinulosus oils where highly active and rich in thymol, as well as Tc2 and Sm2 which were much more active than the corresponding Tc1 and Sm1, the former being richer in thymol than the latter. In Sm1, α-pinene was much more abundant than in Sm2, but its phytotoxic activity has been proven to be very light on
Lolium rigidum Gaudin at all tested doses [
24], in accordance with the result of our experiments in which Sm1 was not active. The promising results from our experiments regarding
T. capitata also confirm previous studies in which extracted oils showed variability among provenances and the species resulted weed-killer against
Sinapis arvensis [
26],
Erigeron bonariensis in pre- and post-emergence assays in greenhouse conditions [
14],
Avena fatua L.,
Echinochloa crus-galli (L.)
P. Beauv. subsp.
crus-galli,
Portulaca oleracea L., and
Amaranthus retroflexus L. [
15].
As for pulegone, the most abundant monoterpene constituent of oils from
C. suaveolens, previous results are consistent with ours. Indeed, in the literature pulegone was found to be a very toxic compound for cucumber (
Cucumis sativus L.), inhibiting root and mitochondrial respiration for concentrations ranging from 50 to 900 ppm [
27]. It also showed inhibitory activity in the germination, seedling, and shoot growth of the test plant
Lactuca sativa [
28]. Pulegone was the main constituent (84%) of
Minthostachys mollis (Benth.) Griseb., which showed inhibitory effects on germination, shoot, and root elongation of the tested species lettuce, tomato, cucumber, and
Bidens pilosa L. [
29].
Alpha-pinene was the main constituent of Sc1 and Sc2 from
Satureja cuneifolia, and Sm1. In Singh et al., the compound inhibited early root growth and caused oxidative damage in root tissue in five test species, and results were concentration dependent [
30]. In Abrahim et al., α-pinene concentrations of 0.05–1.0 mM stimulated respiration while at concentrations higher than 1.0 mM, α-pinene inhibited respiration; moreover, α-pinene had less activity than camphor and eucalyptol in inhibiting seed germination and primary root growth, despite the fact that it had a higher activity on the oxidative metabolism of mitochondria [
31]. In Vasilakoglou et al., α-pinene phytotoxic activity has been proved to be very light on
L. rigidum at dose of 640 nL/cm
3 [
24]. At the concentration we tested, both
Satureja cuneifolia oils and Sm1 were active against broomrape seed germination, but not active in the other experiments. That indicates the need to test further those oils at different doses to better understand their potentiality. The same need for further investigations at different doses is valid for
S. fruticosa subsp.
thomasii oils, which were very partially active only on broomrape seeds, although they contained a high amount of eucalyptol, which proved to be active in other studies. Eucalyptol is one of the most studied monoterpenes, but some previous results are in contrast with ours. It severely affected the germination, speed of germination, seedling growth, chlorophyll content, and respiratory activity of
Ageratum conyzoides (bill goat weed). After two weeks of exposure, plants wilted [
32]. Eucalyptol also inhibited germination of
Brassica rapa L. subsp.
campestris (L.) A.R. Clapham seeds at high concentrations [
33]. Angelini et al. tested in vitro the EO extracted from
Salvia rosmarinus Schleid. (whose main compound was eucalyptol 47%) on three weeds, and it completely inhibited their germination [
23].
P. ramosa is one of the most widespread and troublesome parasitic weeds, mostly present in the Mediterranean area. Seeds proved to be sensitive to all the tested oils at the highest tested dose by direct contact (1000 ppm) and insensitive at the lowest (100 ppm) (
Table 6). Concentrations lower than 1000 ppm should be investigated to find the lowest active dose. As for vapors, broomrape seeds were completely injured by Cs1, Tc2, Sm2, Sc1, Sc2, Ts1, Ts2, and Ts3. Injury was very high for
C. suaveolens Cs2. Those results are promising as those oils could be supplied by micro-irrigation or fumigations to the soil, where broomrape seeds stay. Often EOs face a drop of effectiveness when used row in the field, due to their volatility; that could be overturned in an advantage if vapors are to be used as soil fumigants for broomrape control. Moreover, from our analysis thymol (62) and p-cymene (24) are correlated to the broomrape seed inhibition, so new opportunities should be tested.
As for the leaf disk bioassay, only oil Tc2 from T. capitata gave a reduction in the chlorophyll content. It is likely that the leaf disks were not covered for the most part by the essential oils. Therefore, segments without coverage would have increased injury with a more complete exposition.


 ,
, 



{kind=link}