
Drought Tolerance of Mungbean Is Improved by Foliar Spray of Nanoceria
 ,
,
Abstract
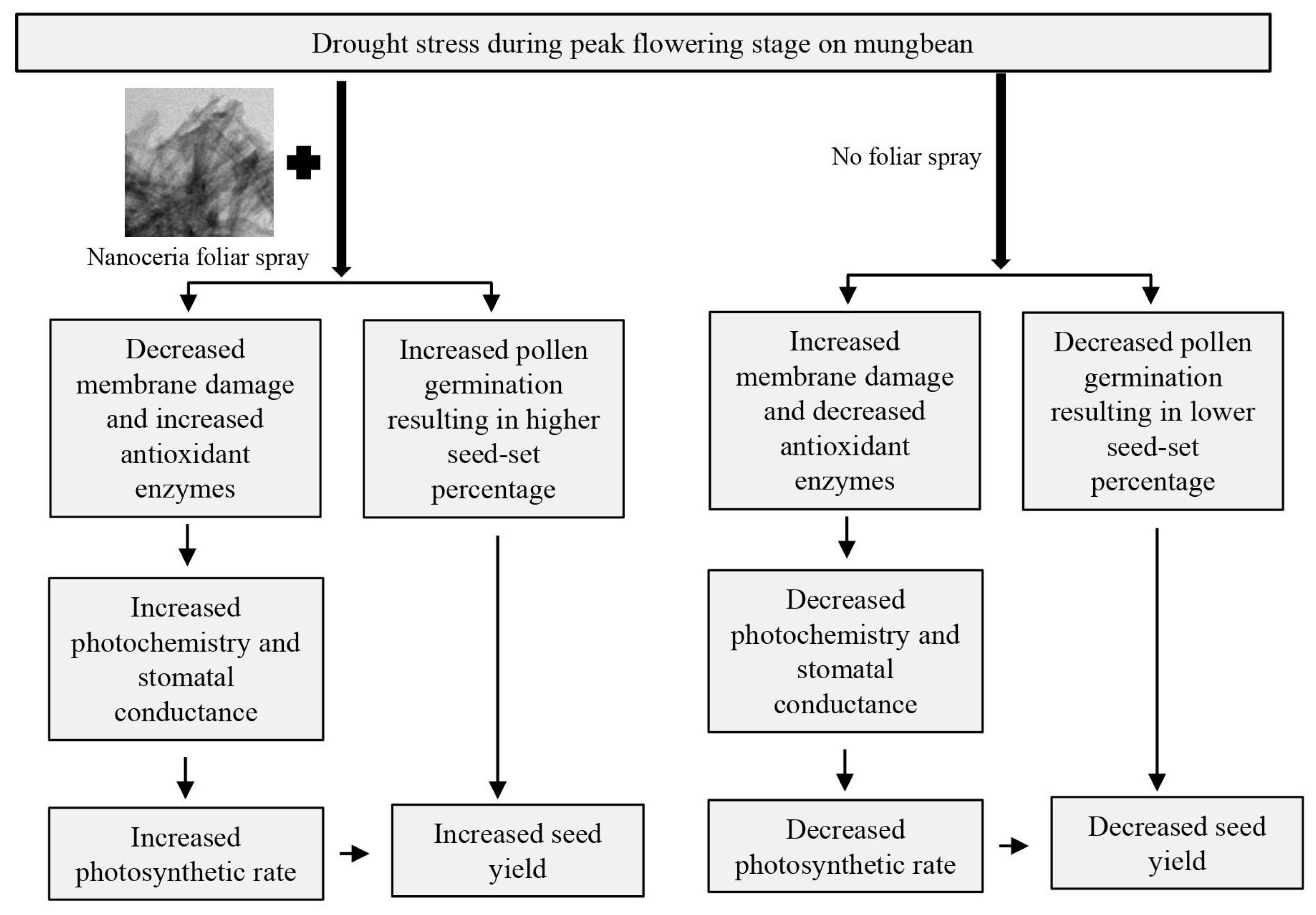
:1. Introduction
2. Materials and Methods
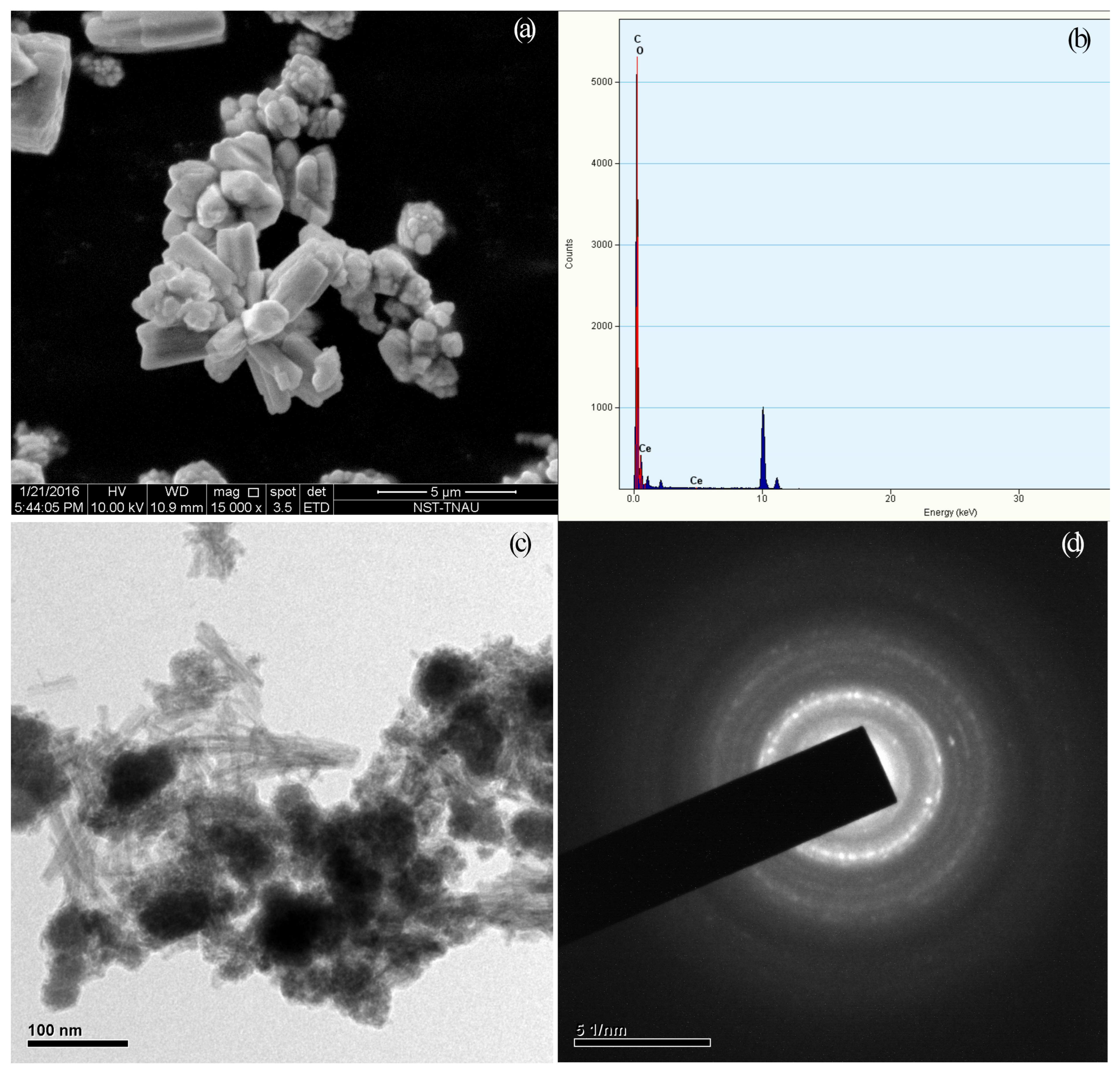
2.1. Synthesis and Characterization of Nanoceria
2.2. Experiment 1: Identification of Optimum Concentration of Nanoceria for the Foliar Spray—Pot Culture Experiment
2.3. Experiment 2: Quantification of Impacts of Nanoceria on Photosynthesis-Associated Traits, Reproductive Success, and Seed Yield of Mungbean—A field Experiment
2.3.1. Imposition of Drought Stress and Foliar Spray of Nanoceria
2.3.2. Chlorophyll Index, Thylakoid Membrane Damage, and Gas Exchange
2.3.3. Pod-set Percentage, Pod Weight, and Seed Yield
2.4. Experiment 3: Confirming the Effects of Nanoceria on Photosynthesis-Associated Traits, Reproductive Success, and Seed Yield of Mungbean—A Pot Culture Experiment
2.4.1. Chlorophyll Content, Thylakoid Membrane Damage, Gas Exchange, and Chlorophyll a Fluorescence Traits
2.4.2. Quantification of Oxidants, Antioxidant Enzyme Activity, and Membrane Integrity
Superoxide Content, Hydrogen Peroxide Content, and Membrane Stability Index
Antioxidant Enzyme Quantification
2.4.3. Pollen Viability, Pod-set Percentage, Individual Seed Weight, and Seed Yield
3. Statistical Analysis
4. Results
4.1. Synthesis and Characterization of Nanoceria
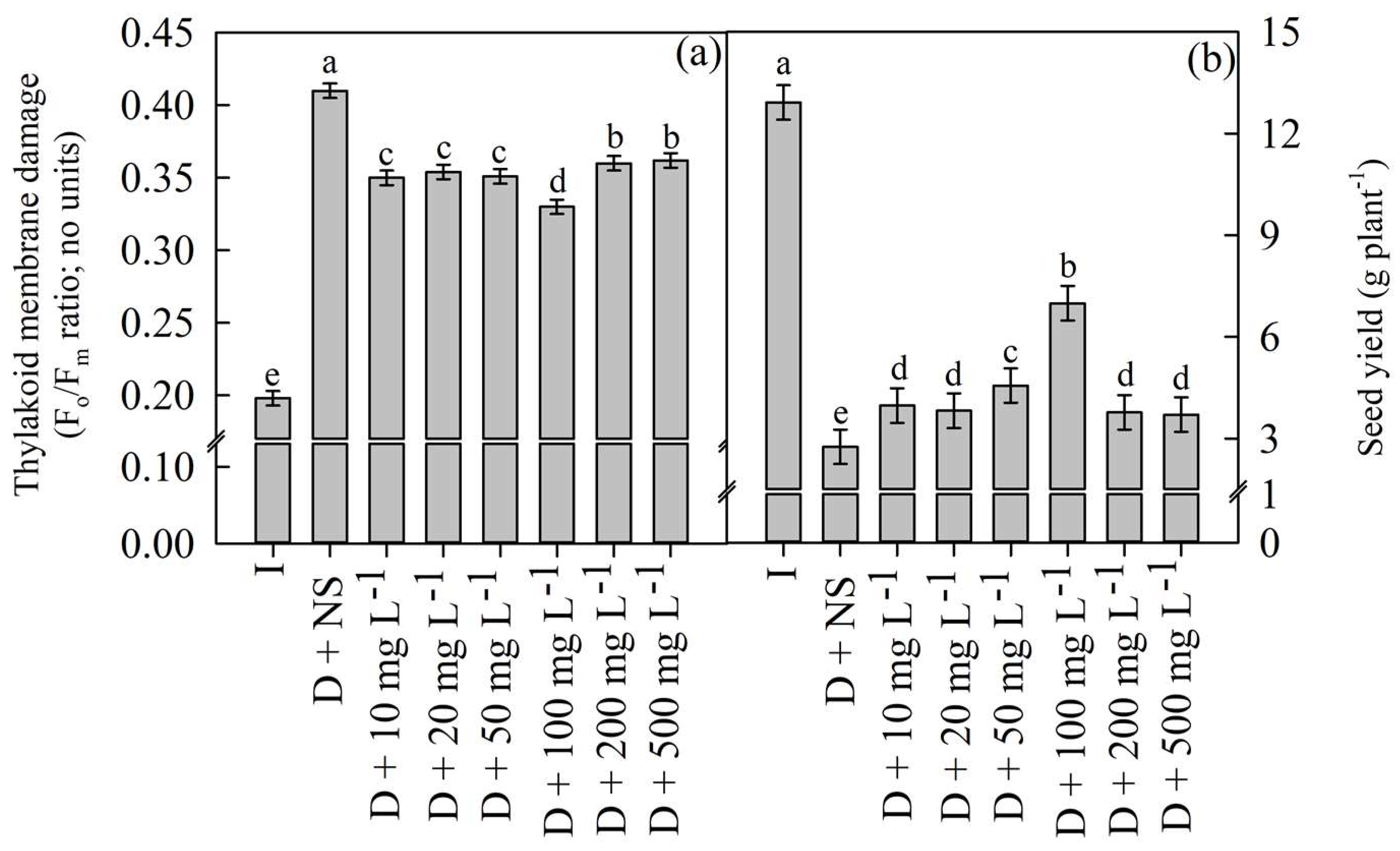
4.2. Experiment 1: Identification of Optimum Concentration of Nanoceria for the Foliar Spray by Assessing Thylakoid Membrane Damage (Fo/Fm Ratio; No Units) and Seed Yield (g Plant−1)—Pot Culture Experiment
4.3. Experiment 2: Quantification of Impacts of Nanoceria on Photosynthesis-Associated Traits, Reproductive Success, and Seed Yield of Mungbean—A Field Experiment
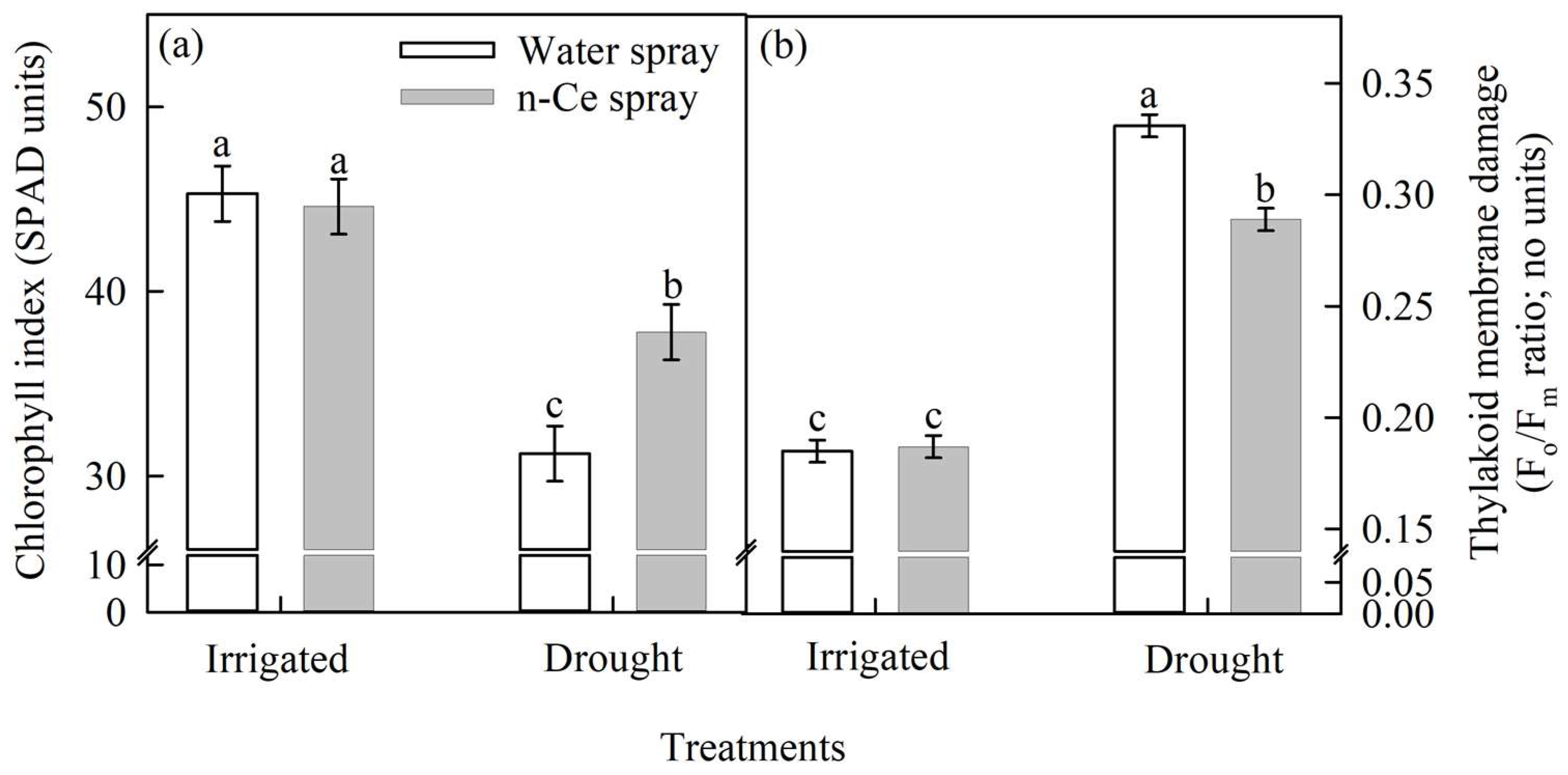
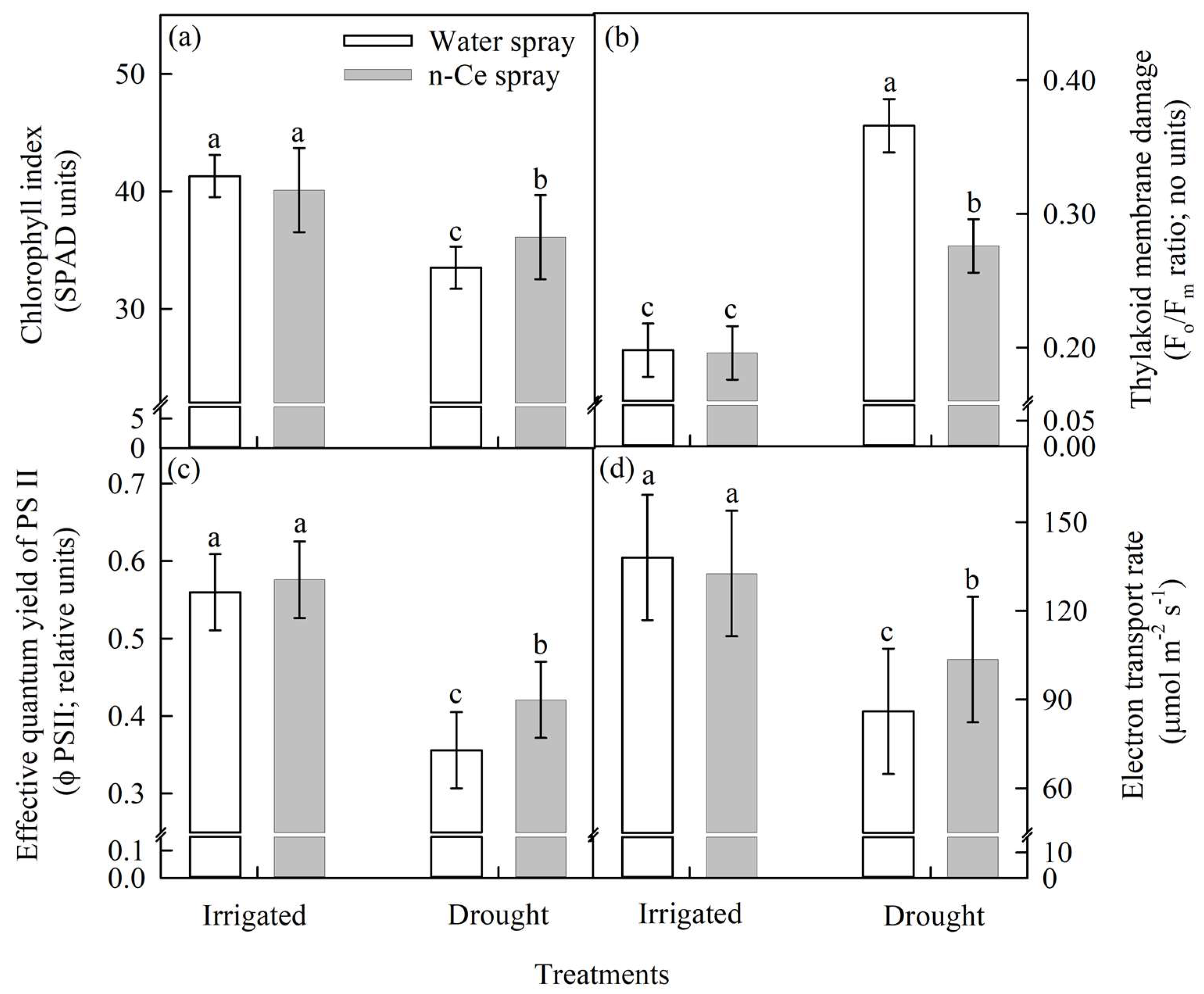
4.3.1. Chlorophyll Index (SPAD units) and Thylakoid Membrane Damage (Fo/Fm ratio)
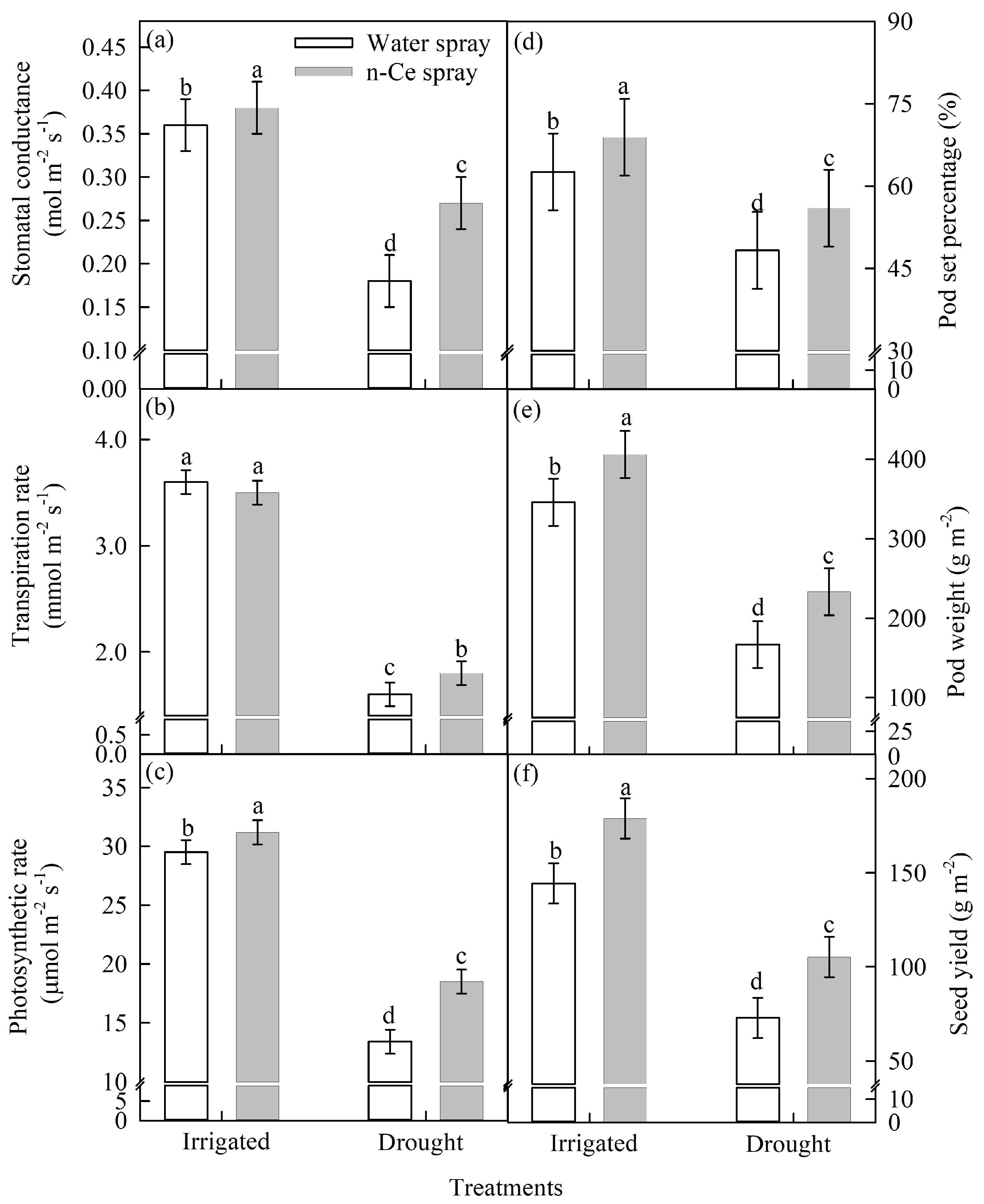
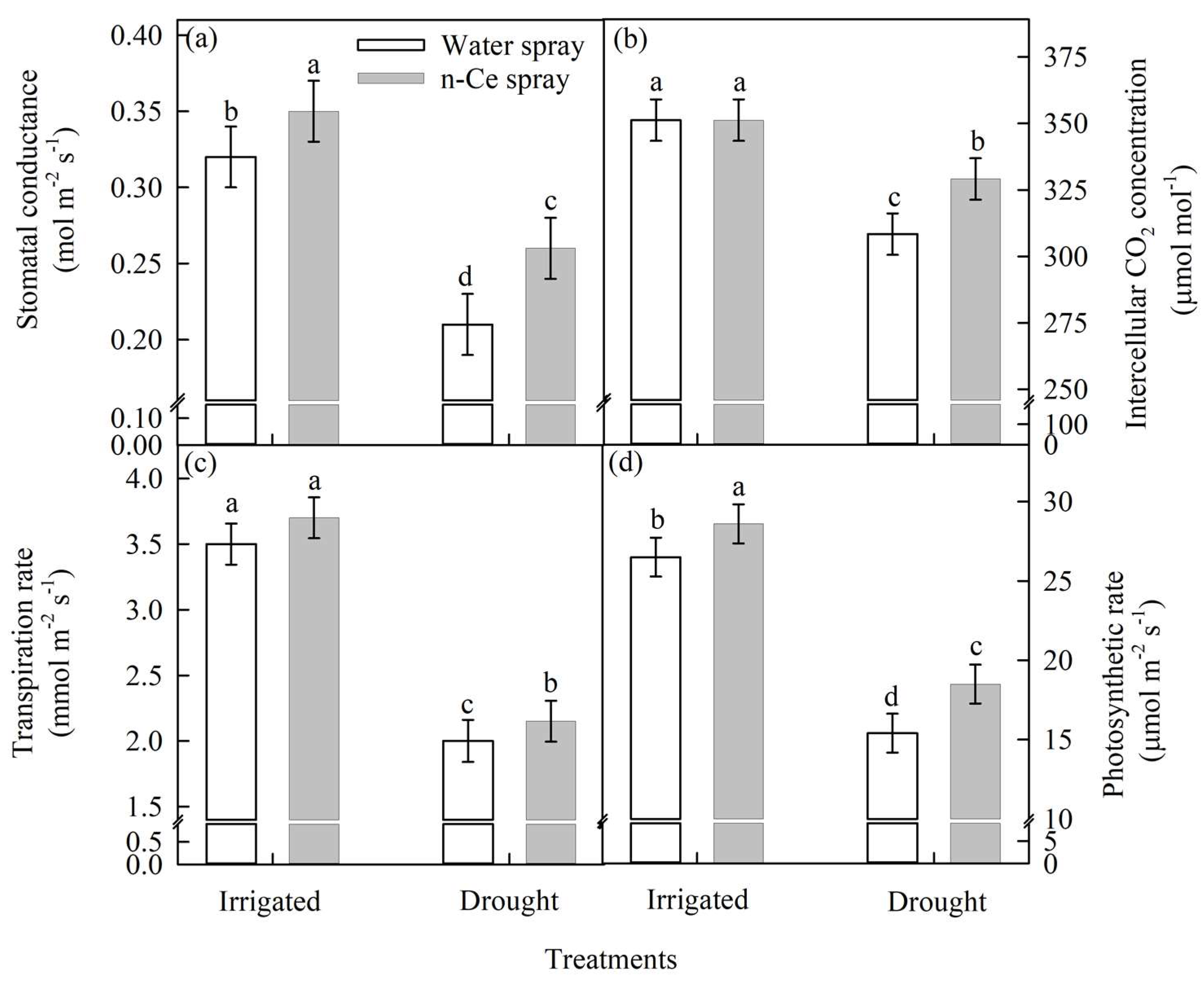
4.3.2. Stomatal Conductance (mol m−2 s−1), Transpiration Rate (mmol m−2 s−1), and Photosynthetic Rate (µmol m−2 s−1)
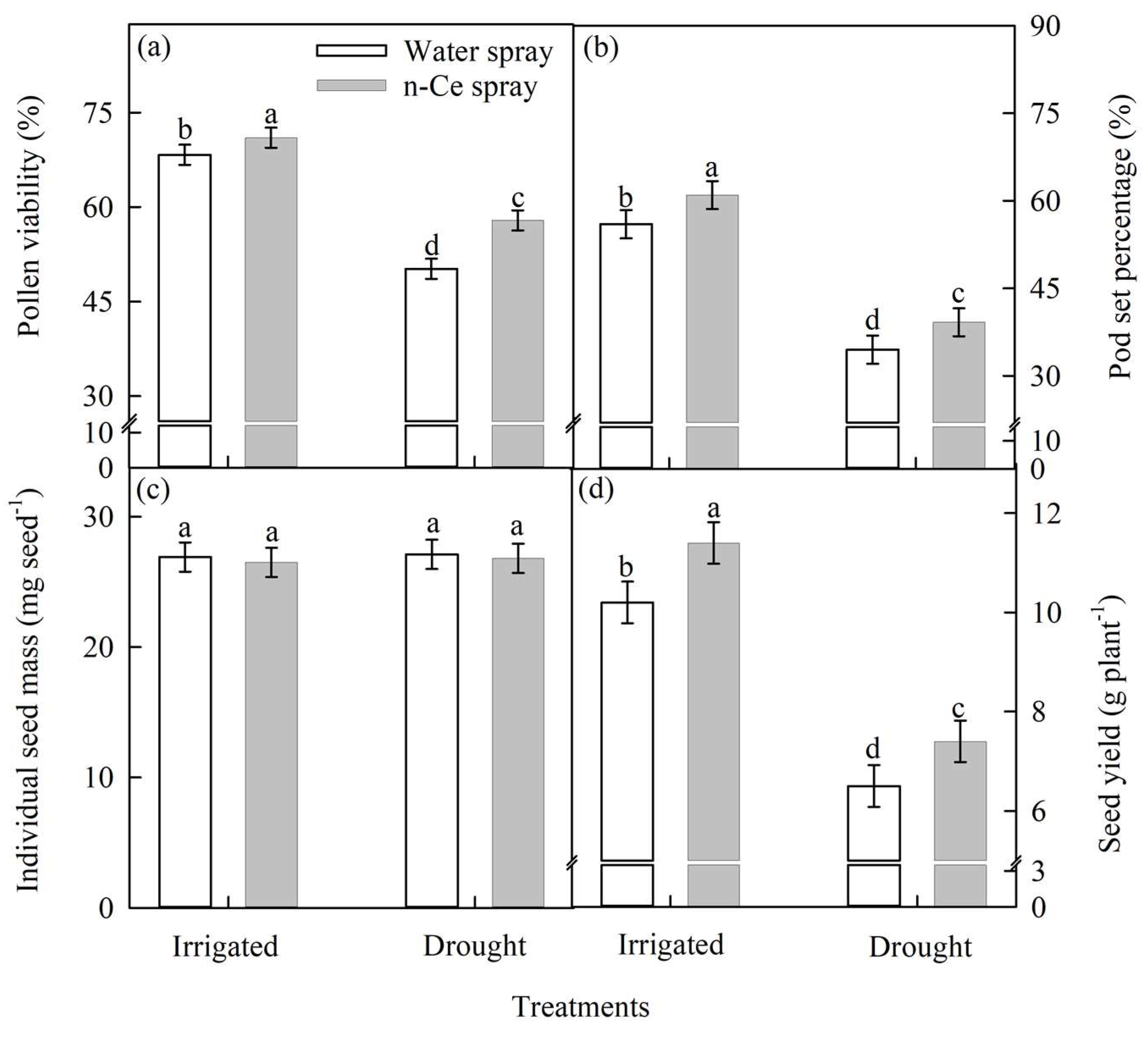
4.3.3. Pod-Set Percentage, Pod Weight (g m−2), and Seed Yield (g m−2)
4.4. Experiment 3: Confirming the Effects of Nanoceria on Oxidative Damage, Photosynthesis-Associated Traits, Reproductive Success, and Seed Yield of Mungbean—A Pot Culture Experiment
4.4.1. Chlorophyll Index, Thylakoid Membrane Damage, the Effective Quantum Yield of PSII (relative units), and Electron Transport Rate (µmol m−2 s−1)
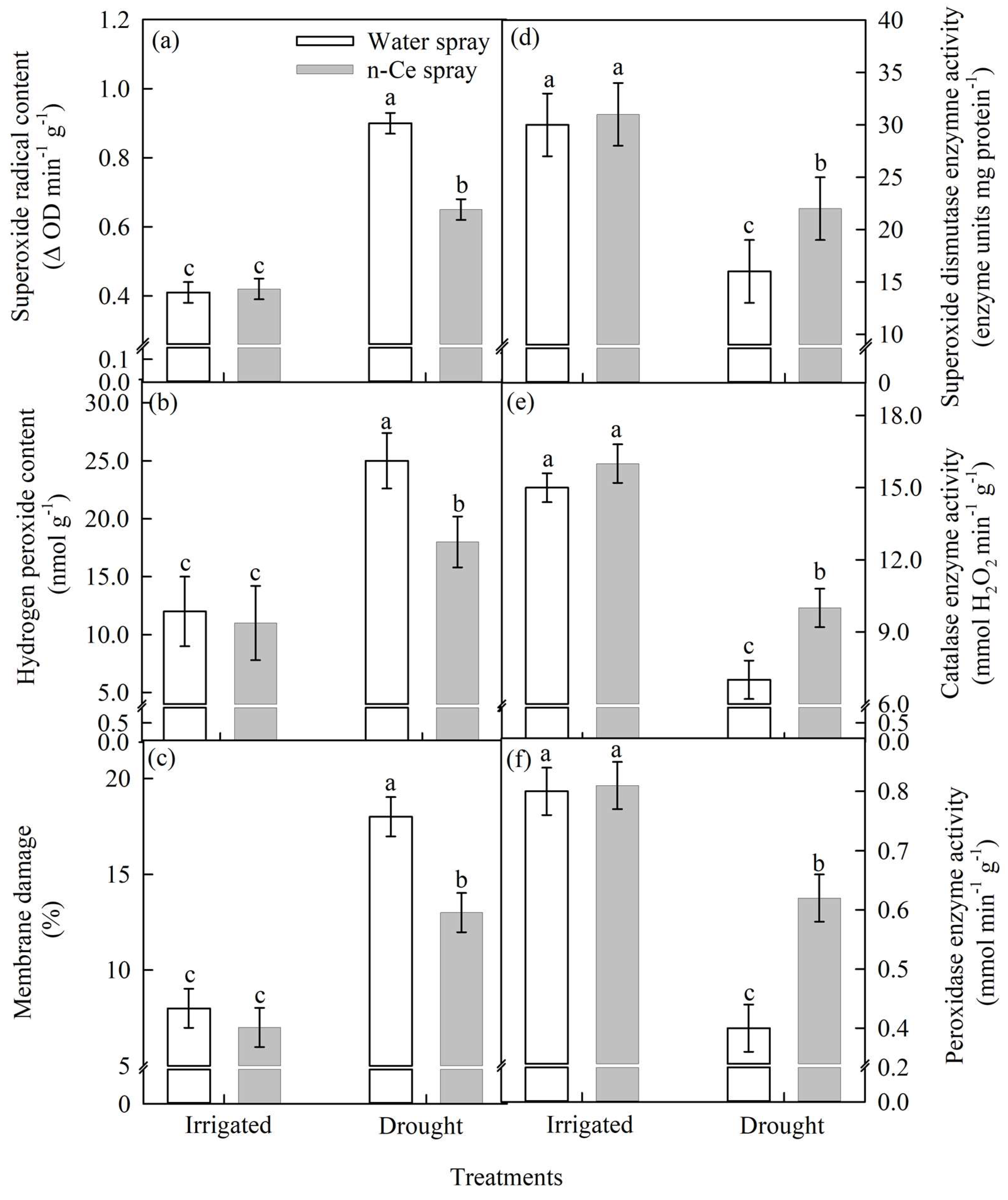
4.4.2. Oxidants, Membrane Damage, and Antioxidant Enzyme Activity
Superoxide Radical Content (Δ OD min−1 g−1), Hydrogen Peroxide Content (nmol g−1), and Membrane Damage (%)
Superoxide Dismutase (enzyme units mg protein−1), Catalase (mmol H2O2 min−1 g−1), and Peroxidase (mmol H2O2 min−1 g−1) Enzyme Activity
4.4.3. Stomatal Conductance, Intercellular CO2 Concentration, Transpiration Rate, and Photosynthetic Rate
4.4.4. Pollen Viability (%), Pod-Set Percentage, Individual Seed Mass (mg seed−1), and Seed Yield (g plant−1)
5. Discussion
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Searchinger, T.; Waite, R.; Hanson, C.; Ranganathan, J.; Dumas, P.; Matthews, E. Creating a Sustainable Food Future—A Menu of Solutions to Feed Nearly 10 Billion People by 2050. Final Report, World Resources Report. July 2019. Available online: https://wrr-food.wri.org/sites/default/files/2019-07/WRR_Food_Full_Report_0.pdf (accessed on 16 April 2020).
- Ma, X.; Geisler-Lee, J.; Deng, Y.; Kolmakov, A. Interactions between engineered nanoparticles (ENPs) and plants: Phytotoxicity, uptake and accumulation. Sci. Total Environ. 2010, 408, 3053–3061. [Google Scholar] [CrossRef] [PubMed]
- Djanaguiraman, M.; Nair, R.; Giraldo, J.P.; Prasad, P.V.V. Cerium oxide nanoparticles decrease drought-induced oxidative damage in sorghum leading to higher photosynthesis and grain yield. ACS Omega 2018, 3, 14406–14416. [Google Scholar] [CrossRef] [PubMed]
- Kebede, E. Contribution, utilization, and improvement of legumes-driven biological nitrogen fixation in agricultural systems. Front. Sustain. Food Syst. 2021, 5, 767998. [Google Scholar] [CrossRef]
- Gowda, C.L.L.; Parthasarathy Rao, P.; Bhagavatula, S. Global trends in production and trade of major grain legumes. In Proceedings of the International Conference on Grain Legumes: Quality Improvement, Value Addition and Trade, Kanpur, India, 14–16 February 2009; pp. 282–301. [Google Scholar]
- Foyer, C.H.; Lam, H.M.; Nguyen, H.T.; Siddique, K.H.; Varshney, R.K.; Colmer, T.D.; Cowling, W.; Bramley, H.; Mori, T.A.; Hodgson, J.M.; et al. Neglecting legumes has compromised human health and sustainable food production. Nat. Plants 2016, 2, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Ayachit, G.; Sahoo, L. Screening of mungbean for drought tolerance and transcriptome profiling between drought-tolerant and susceptible genotype in response to drought stress. Plant Physiol. Biochem. 2020, 157, 229–238. [Google Scholar] [CrossRef]
- Nair, R.M.; Yang, R.-Y.; Easdown, W.J.; Thavarajah, D.; Thavarajah, P.; Hughes, J.A.; Keatinge, J. Biofortification of mungbean (Vigna radiata) as a whole food to enhance human health. J. Sci. Food Agric. 2013, 93, 1805–1813. [Google Scholar] [CrossRef]
- Ripley, B.; Frole, K.; Gilbert, M. Differences in drought sensitivities and photosynthetic limitations between co-occurring C3 and C4 (NADP-ME) Panicoid grasses. Ann. Bot. 2010, 105, 493–503. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.; Lata, C.; Chauhan, P.S.; Nautiyal, C.S. Pseudomonas putida attunes morphophysiological, biochemical and molecular responses in Cicer arietinum L. during drought stress and recovery. Plant Physiol. Biochem. 2016, 99, 108–117. [Google Scholar] [CrossRef]
- Kaushal, M.; Wani, S.P. Rhizobacterial-plant interactions: Strategies ensuring plant growth promotion under drought and salinity stress. Agric. Ecosyst. Environ. 2016, 231, 68–78. [Google Scholar] [CrossRef]
- Flexas, J.; Bota, J.; Galmes, J.; Medrano, H.; Ribas-Carbo, M. Keeping a positive carbon balance under adverse conditions: Responses of photosynthesis and respiration to water stress. Physiol. Plant. 2006, 127, 343–352. [Google Scholar] [CrossRef]
- Sarkar, S.; Khatun, M.; Era, F.M.; Islam, A.K.M.M.; Anwar, M.P.; Danish, S.; Datta, R.; Islam, A.K.M.A. Abiotic stresses: Alteration of composition and grain quality in food legumes. Agronomy 2021, 11, 2238. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Tezara, W. Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: A critical evaluation of mechanisms and integration of processes. Ann. Bot. 2009, 103, 561–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, A.; Selim, M.M.; Alderfasi, A.A.; Afzal, M. Effect of drought stress on mung bean (Vigna radiata L.) under arid climatic conditions of Saudi Arabia. In Ecosystem and Sustainable Development; Miralles, I., Garcia, J.L., Brebbia, C.A., Eds.; WIT Press: Southampton, UK, 2015; pp. 185–193. [Google Scholar]
- Boghossian, A.A.; Sen, F.; Gibbons, B.M.; Sen, S.; Faltermeier, S.M.; Giraldo, J.P.; Zhang, C.T.; Zhang, J.; Strano, M.S. Application of nanoparticle antioxidants to enable hyperstable chloroplasts for solar energy harvesting. Adv. Energy Mater. 2013, 3, 881–893. [Google Scholar] [CrossRef]
- Reed, K.; Cormack, A.; Kulkarni, A.; Mayton, M.; Sayle, D.; Klaessig, F.; Stadler, B. Exploring the properties and applications of nanoceria: Is there still plenty of room at the bottom? Environ. Sci. Nano 2014, 1, 390–405. [Google Scholar] [CrossRef] [Green Version]
- Djanaguiraman, M.; Prasad, P.V.V.; Schapaugh, W.T. High day-or nighttime temperature alters leaf assimilation, reproductive success, and phosphatidic acid of pollen grain in soybean [Glycine max (L.) Merr.]. Crop Sci. 2013, 53, 1594–1604. [Google Scholar] [CrossRef]
- Chaitanya, K.S.K.; Naithani, S.C. Role of superoxide, lipid peroxidation and superoxide dismutase in membrane perturbation during loss of viability in seeds of Shorea robusta Gaertn. f. New Phytol. 1994, 126, 623–627. [Google Scholar] [CrossRef]
- Patterson, B.D.; Payne, L.A.; Cnen, Y.; Graham, D. An inhibitor of catalase induced by cold in chilling-sensitive plants. Plant Physiol. 1984, 76, 1014–1018. [Google Scholar] [CrossRef] [Green Version]
- Sairam, R.K.; Deshmukh, P.S.; Shukla, D.S. Tolerance of drought and temperature stress in relation to increased antioxidant enzyme activity in wheat. J. Agron. Crop Sci. 1997, 178, 171–178. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef]
- Samantary, S. Biochemical responses of Cr-tolerant and Cr-sensitive mung bean cultivars grown on varying levels of chromium. Chemosphere 2002, 47, 1065–1072. [Google Scholar] [CrossRef] [PubMed]
- Castillo, F.; Penel, I.; Greppin, H. Peroxidase release induced by ozone in Sedum album leaves. Plant Physiol. 1984, 74, 846–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- SAS Institute. The SAS Users Guide, Version 9.3; SAS Inst.: Cary, NC, USA, 2008. [Google Scholar]
- Paramo, L.A.; Feregrino-Perez, A.A.; Guevara, R.; Mendoza, S.; Esquivel, K. Nanoparticles in agroindustry: Applications, toxicity, challenges, and trends. J. Nanomat. 2020, 10, 1654. [Google Scholar] [CrossRef]
- Singh, A.K. Introduction to Nanoparticles and Nanotoxicology; Academic Press: Cambridge, MA, USA, 2016; pp. 1–18. [Google Scholar]
- Peng, X.J.; Luan, Z.K.; Zing, J.; Di, Z.H.; Li, Y.H.; Tian, B.H. Ceria nanoparticles supported on carbon nanotubes for the removal of arsenate from water. Mater. Lett. 2005, 59, 399–403. [Google Scholar] [CrossRef]
- Sardesai, N.P.; Daniel, A.; Silvana, A. Electroanalytical evaluation of antioxidant activity of cerium oxide nanoparticles by nanoparticle collisions at microelectrodes. J. Am. Chem. Soc. 2013, 135, 16770–16773. [Google Scholar] [CrossRef] [PubMed]
- Heckert, E.G.; Seal, S.; Self, W.T. Fenton-like reaction catalyzed by the rare earth inner transition metal cerium. Environ. Sci. Tech. 2008, 42, 5014–5019. [Google Scholar] [CrossRef] [Green Version]
- Havaux, M. Characterization of thermal damage to the photosynthetic electron transport system in potato leaves. Plant Sci. 1993, 94, 19–33. [Google Scholar] [CrossRef]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought-from genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Cornic, G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants. Plant Cell Environ. 2002, 25, 275–294. [Google Scholar] [CrossRef] [Green Version]
- Krause, G.H. Photoinhibition of photosynthesis: An evaluation of damaging and protective mechanisms. Physiol. Plant. 1988, 74, 566–574. [Google Scholar] [CrossRef]
- Sgherri, C.L.M.; Pinzino, C.; Navari-Izzo, F. Sunflower seedlings subjected to increasing stress by water deficit: Changes in O2− production related to the composition of thylakoid membranes. Physiol. Plant. 1996, 96, 446–452. [Google Scholar] [CrossRef]
- Nguyen, G.N.; Hailstones, D.L.; Wilkes, M.; Sutton, B.G. Drought-Induced oxidative conditions in rice anthers leading to a programmed cell death and pollen abortion. J. Agron. Crop Sci. 2009, 195, 157–164. [Google Scholar] [CrossRef]
- Saini, H.S. Effects of water stress on male gametophyte development in plants. Sex. Plant Reprod. 1997, 10, 67–73. [Google Scholar] [CrossRef]
- Pang, J.; Turner, N.C.; Khan, T.; Du, Y.L.; Xiong, J.L.; Colmer, T.D.; Devilla, R.; Stefanova, K.; Siddique, K.H.M. Response of chickpea (Cicer arietinum L.) to terminal drought: Leaf stomatal conductance, pod abscisic acid concentration, and seed set. J. Exp. Bot. 2017, 68, 1973–1985. [Google Scholar]
- Saini, H.S.; Westgate, M.E. Reproductive development in grain crops during drought. Adv. Agron. 2000, 68, 59–96. [Google Scholar]
- Djanaguiraman, M.; Perumal, R.; Jagadish, S.V.K.; Ciampitti, I.A.; Welti, R.; Prasad, P.V.V. Sensitivity of sorghum pollen and pistil to high-temperature stress. Plant Cell Environ. 2018, 41, 1065–1082. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Details |
|---|---|
| T1 | Irrigated control and no spray |
| T2 | Drought stressed and no spray |
| T3 | Drought stressed and sprayed with nanoceria @ 10 mg L−1 |
| T4 | Drought stressed and sprayed with nanoceria @ 20 mg L−1 |
| T5 | Drought stressed and sprayed with nanoceria @ 50 mg L−1 |
| T6 | Drought stressed and sprayed with nanoceria @ 100 mg L−1 |
| T7 | Drought stressed and sprayed with nanoceria @ 200 mg L−1 |
| T8 | Drought stressed and sprayed with nanoceria @ 500 mg L−1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maduraimuthu, D.; Alagarswamy, S.; Prabhakaran, J.; Karuppasami, K.M.; Venugopal, P.B.R.; Koothan, V.; Natarajan, S.; Dhashnamurthi, V.; Veerasamy, R.; Rathinavelu, S.; et al. Drought Tolerance of Mungbean Is Improved by Foliar Spray of Nanoceria. Agronomy 2023, 13, 201. https://doi.org/10.3390/agronomy13010201
Maduraimuthu D, Alagarswamy S, Prabhakaran J, Karuppasami KM, Venugopal PBR, Koothan V, Natarajan S, Dhashnamurthi V, Veerasamy R, Rathinavelu S, et al. Drought Tolerance of Mungbean Is Improved by Foliar Spray of Nanoceria. Agronomy. 2023; 13(1):201. https://doi.org/10.3390/agronomy13010201
Chicago/Turabian StyleMaduraimuthu, Djanaguiraman, Senthil Alagarswamy, Jeyakumar Prabhakaran, Kalarani M. Karuppasami, Prasad B. R. Venugopal, Vanitha Koothan, Sritharan Natarajan, Vijayalakshmi Dhashnamurthi, Ravichandran Veerasamy, Sivakumar Rathinavelu, and et al. 2023. "Drought Tolerance of Mungbean Is Improved by Foliar Spray of Nanoceria" Agronomy 13, no. 1: 201. https://doi.org/10.3390/agronomy13010201